
ZmNLR-7-Mediated Synergistic Regulation of ROS, Hormonal Signaling, and Defense Gene Networks Drives Maize Immunity to Southern Corn Leaf Blight

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analysis of ZmNLR-7
2.2. Vector Construction and Subcellular Localization
2.3. Pathogen Cultivation
2.4. Plant Materials and Treatments
2.5. RNA Extraction and RT-PCR Analysis
2.6. EMS Mutant Identification
2.7. EMS Mutant Identification Physiological Parameter Analysis of ZmNLR-7
2.8. RNA Sequencing (RNA-Seq) Analysis
2.9. Statistical Analysis
3. Results
3.1. Bioinformatics Analysis of the ZmNLR-7 Gene
3.2. Analysis of Subcellular Localization of ZmNLR-7
3.3. Analysis of the Expression Pattern of ZmNLR-7
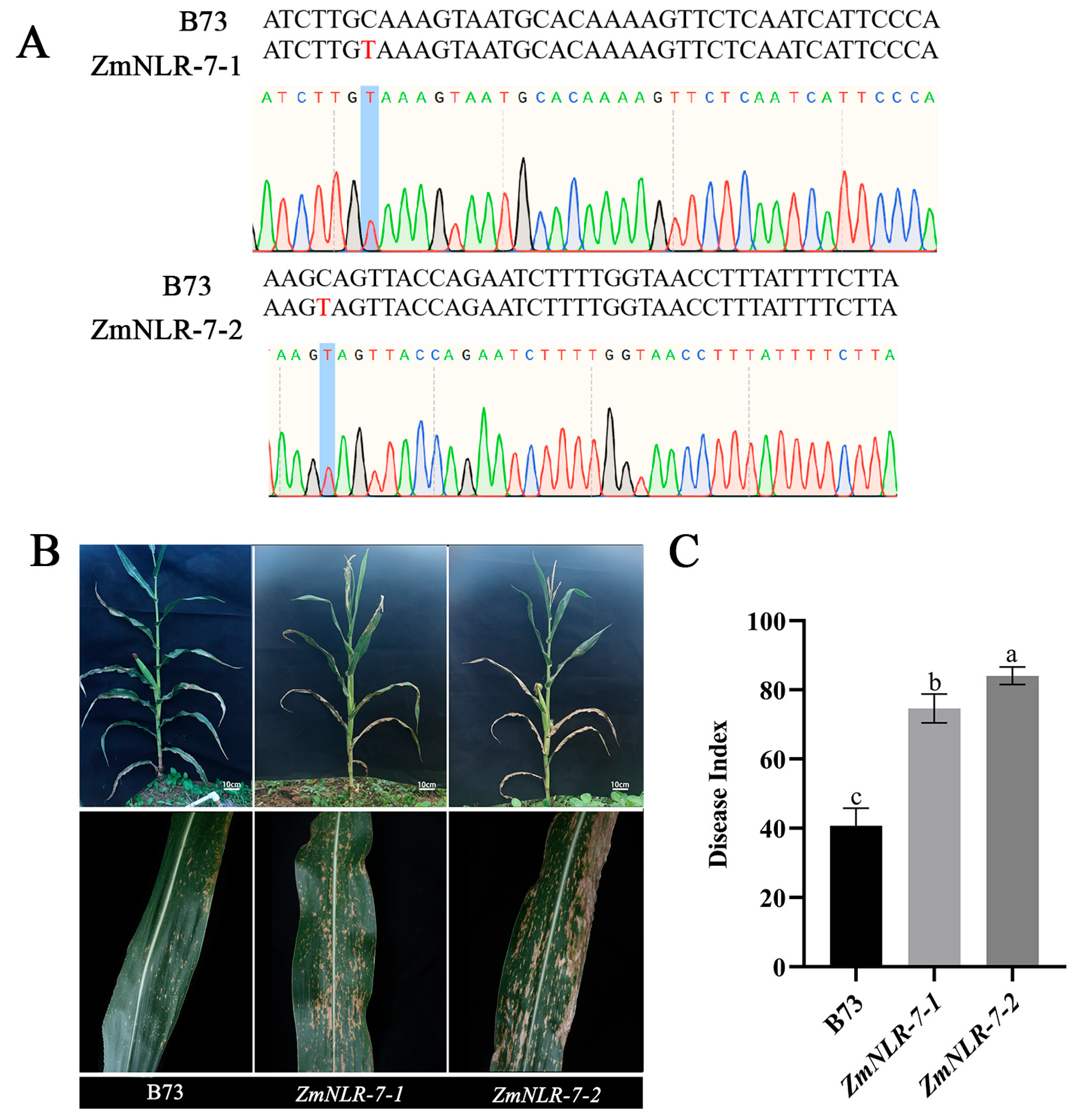
3.4. Obtaining Homozygous Mutants of ZmNLR-7 Maize and Phenotypic Analysis of Response to Infection with Bipolaris Maydis
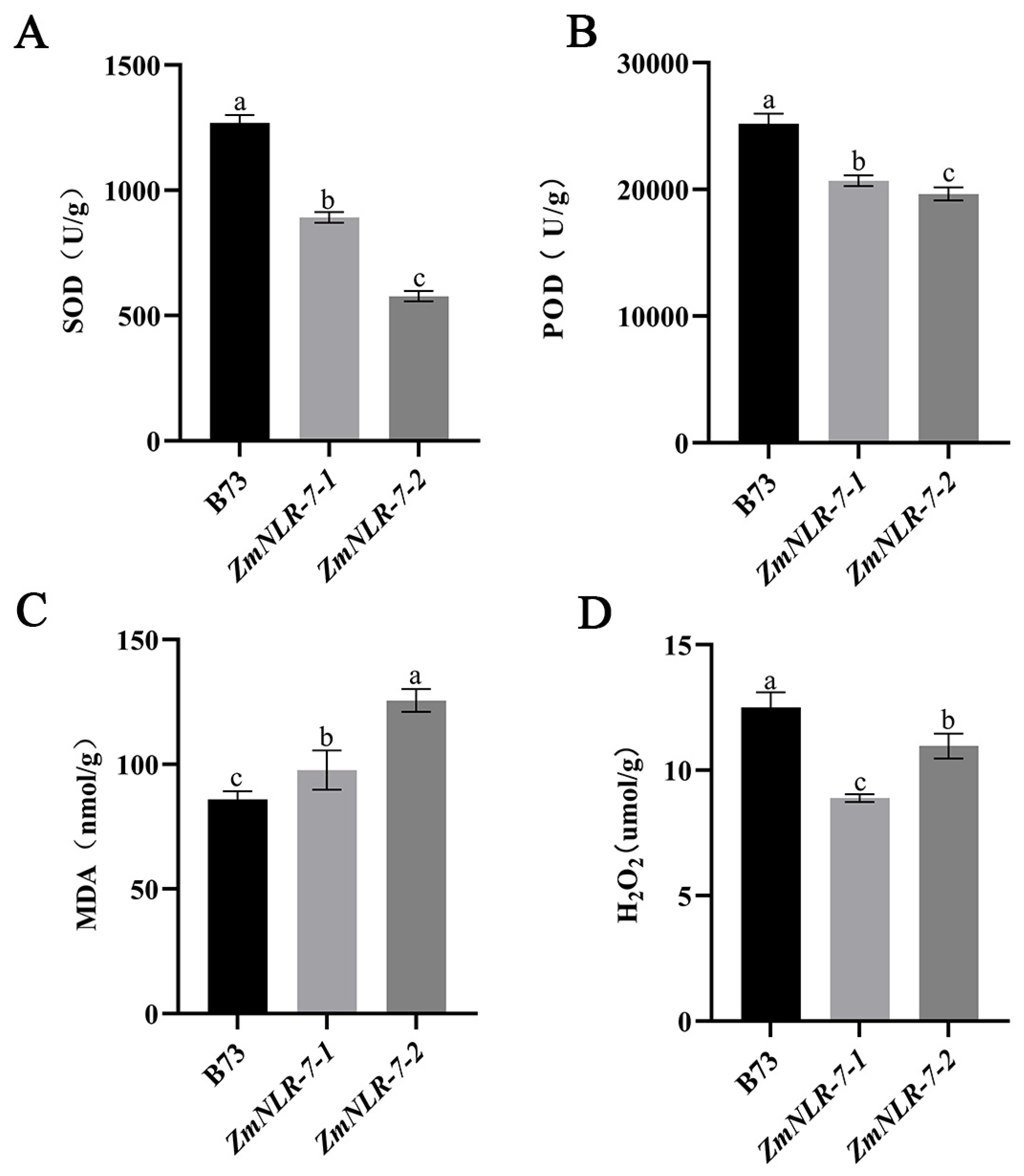
3.5. Determination of Physiological Indicators of ZmNLR-7 Maize Mutants in Response to Infection with Bipolaris Maydis
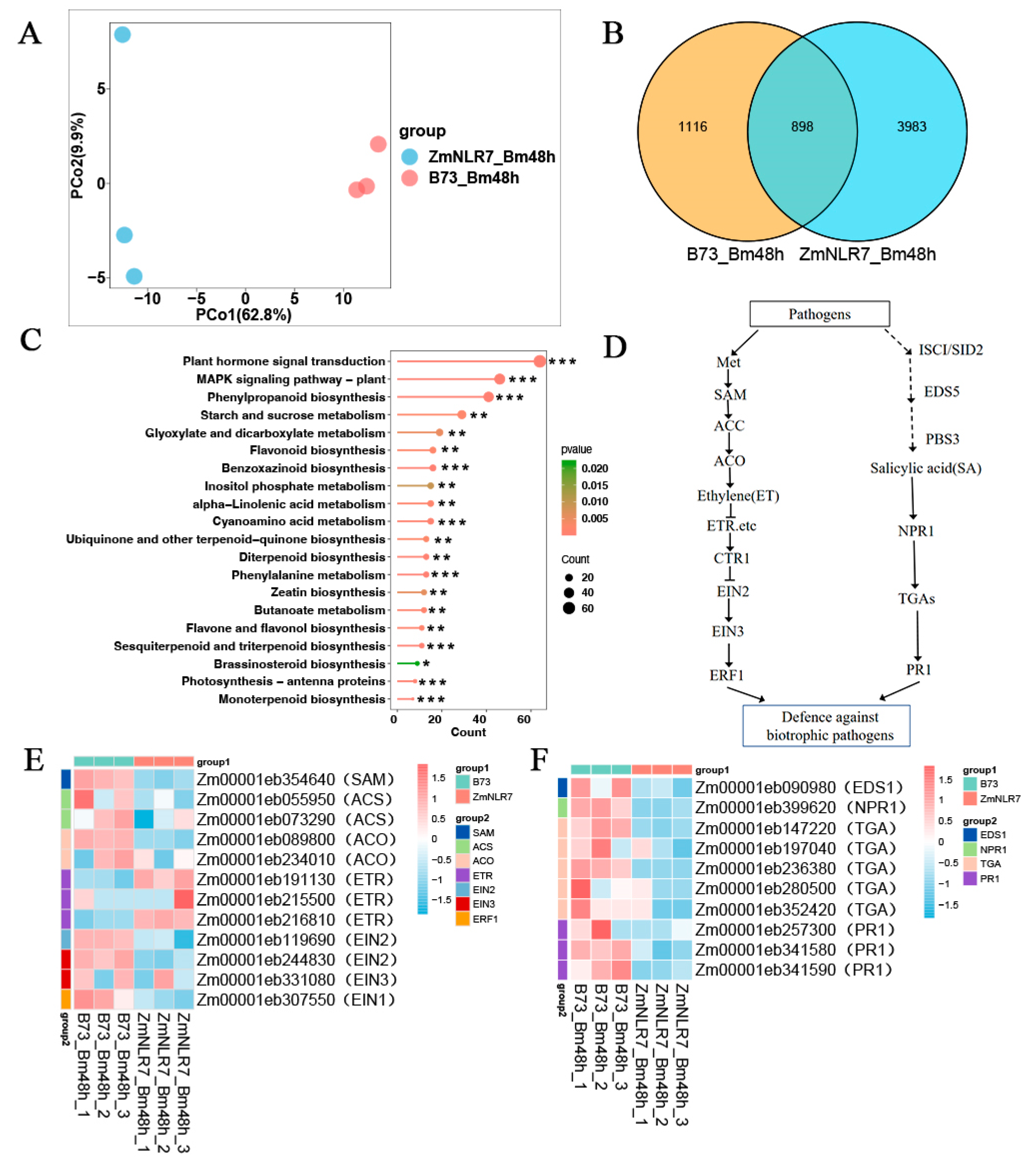
3.6. Transcriptomic Analysis of ZmNLR-7 Maize Mutant in Response to Bipolaris Maydis Infection
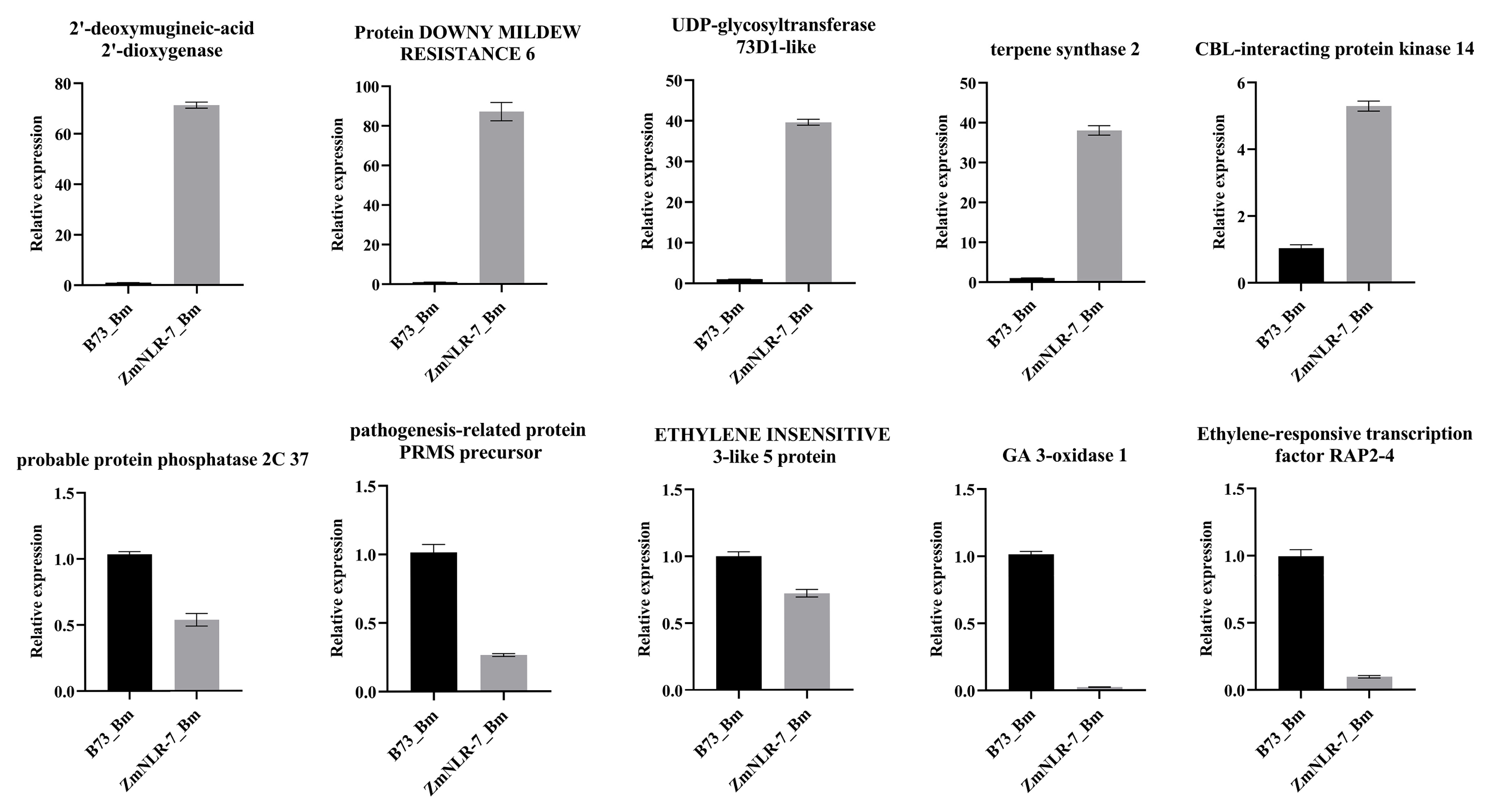
3.7. Fluorescence Quantitative Verification of ZmNLR-7 Transcriptomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Niu, H.; Liu, C.; Wang, H.; Yin, W.; Xia, X. PTI-ETI synergistic signal mechanisms in plant immunity. Plant Biotechnol. J. 2024, 22, 2113–2128. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Menke, F.L.; Yoshioka, K.; Moder, W.; Shirano, Y.; Klessig, D.F. High humidity suppresses ssi4-mediated cell death and disease resistance upstream of MAP kinase activation, H2O2 production and defense gene expression. Plant J. 2004, 39, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.-M.; He, S.Y.; Xin, X.-F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Ahn, H.-K.; Ding, P.; Jones, J.D.G. Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.-S.; Zhu, Y.-X.; Zhang, Y.-F.; Zhong, X.; Pan, K.-Y.; Jiang, Y.; Wen, C.-K.; Yang, Z.-N.; Yao, X. Ethylene activates the EIN2-EIN3/EIL1 signaling pathway in tapetum and disturbs anther development in Arabidopsis. Cells 2022, 11, 3177. [Google Scholar] [CrossRef] [PubMed]
- Duxbury, Z.; Wu, C.-H.; Ding, P. A comparative overview of the intracellular guardians of plants and animals: NLRs in innate immunity and beyond. Annu. Rev. Plant Biol. 2021, 72, 155–184. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Liu, Y.-S.; Budai-Hadrian, O.; Sela, M.; Carmel-Goren, L.; Zamir, D.; Fluhr, R. Comparative genetics of nucleotide binding site-leucine rich repeat resistance gene homologues in the ge-nomes of two dicotyledons: Tomato and Arabidopsis. Genetics 2000, 155, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Wendel, J.; Fluhr, R. Divergent evolution of plant NBS-LRR resistance gene homologues in dicot and cereal genomes. J. Mol. Evol. 2000, 50, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Tamborski, J.; Krasileva, K.V. Evolution of plant NLRs: From natural history to precise modifications. Annu. Rev. Plant Biol. 2020, 71, 355–378. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.A.; Takemoto, D. Plant innate immunity–direct and indirect recognition of general and specific pathogen-associated molecules. Curr. Opin. Immunol. 2004, 16, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.B.; Bogdanove, A.J.; Sessa, G. Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 2003, 54, 23–61. [Google Scholar] [CrossRef] [PubMed]
- Danot, O.; Marquenet, E.; Vidal-Ingigliardi, D.; Richet, E. Wheel of life, wheel of death: A mechanistic insight into signaling by STAND proteins. Structure 2009, 17, 172–182. [Google Scholar] [CrossRef] [PubMed]
- van der Biezen, E.A.; Jones, J.D. The NB-ARC domain: A novel signalling motif shared by plant resistance gene products and regulators of cell death in animals. Curr. Biol. 1998, 8, R226–R228. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Xie, L.; Chakraborty, S.; Wang, A.; Matny, O.; Jugovich, M.; Kolmer, J.A.; Richardson, T.; Bhatt, D.; Hoque, M.; et al. A five-transgene cassette confers broad-spectrum resistance to a fungal rust pathogen in wheat. Nat. Biotechnol. 2021, 39, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Hatta, M.A.M.; Arora, S.; Ghosh, S.; Matny, O.; Smedley, M.A.; Yu, G.; Chakraborty, S.; Bhatt, D.; Xia, X.; Steuernagel, B.J. The wheat Sr22, Sr33, Sr35 and Sr45 genes confer resistance against stem rust in barley. Plant Biotechnol. J. 2021, 19, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Dracatos, P.M.; Lu, J.; Sánchez-Martín, J.; Wulff, B.B. Resistance that stacks up: Engineering rust and mildew disease control in the cereal crops wheat and barley. Plant Biotechnol. J. 2023, 21, 1938–1951. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, C.; Zhao, X.; Liu, J.; Cui, T.; Ren, Y.; Niu, Y.; Tang, Z. Research Progress on Pyramiding Breeding of Disease-Resistant Related Genes in Wheat. Shanxi Agric. Sci. 2017, 45, 308–313. [Google Scholar]
- Feng, Y.; Ma, Y.; Feng, F.; Chen, X.; Qi, W.; Ma, Z.; Song, R. Accumulation of 22 kDa α-zein-mediated nonzein protein in protein body of maize endosperm. New Phytol. 2022, 233, 265–281. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Cao, Q.; Dong, J.; Qiao, M.; Wang, Z.; Zhang, Z.; Sun, H.; Xie, H.; Ge, M.; Zhang, Y.; et al. Transcription factor ZmNLP8 modulates nitrate utilization by transactivating ZmNiR1.2 in maize. Plant J. 2025, 122, e70263. [Google Scholar] [CrossRef] [PubMed]
- Hang, T.; Ling, X.; He, C.; Xie, S.; Jiang, H.; Ding, T. Isolation of the zmers4 gene from maize and its functional analysis in transgenic plants. Front. Microbiol. 2021, 12, 632908. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Ge, L.; Ye, X.; Xu, L.; Si, W.; Ding, T. ZmGLP1, a germin-like protein from maize, plays an important role in the regulation of pathogen resistance. Int. J. Mol. Sci. 2022, 23, 14316. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Jiang, H.; Song, Z.; Liu, W.; Rao, S.; Jiang, H.; Wu, G.; Ding, T. Overexpression of ZmEREB211 confers enhanced susceptibility to Pseudomonas syringae pv. tomato DC3000 in Arabidopsis. Plant Sci. 2025, 356, 112482. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.-C.; Zhang, Y.-R.; Liu, Z.-G.; Li, X.-C.; Yu, Z.; Ping, B.-Y.; Sun, Y.-Q.; Burg, H.v.D.; Ma, F.-W.; Zhao, T.; et al. Deciphering Plant NLR Genomic Evolution: Synteny-Informed Classification Unveils Insights into TNL Gene Loss. Mol. Biol. Evol. 2025, 42, msaf015. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Jia, A.; Ma, S.; Sun, Y.; Chang, X.; Han, Z.; Chai, J. NLR signaling in plants: From resistosomes to second messengers. Trends Biochem. Sci. 2023, 48, 776–787. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.T.; Anderson, R.G.; Cherkis, K.A.; Law, T.F.; Liu, Q.L.; Machius, M.; Nimchuk, Z.L.; Yang, L.; Chung, E.-H.; El Kasmi, F.; et al. TIR-only protein RBA1 recognizes a pathogen effector to regulate cell death in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, E2053–E2062. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Lapin, D.; Liu, L.; Sun, Y.; Song, W.; Zhang, X.; Logemann, E.; Yu, D.; Wang, J.; Jirschitzka, J.; et al. Direct pathogen-induced assembly of an NLR immune receptor complex to form a holoenzyme. Science 2020, 370, eabe3069. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zhang, F.; Du, X.; Zhang, X.; Huang, X.; Li, Z.; Zhang, Y.; Gan, P.; Li, H.; Li, M.; et al. TaANK-TPR1 enhances wheat resistance against stripe rust via controlling gene expression and protein activity of NLR protein TaRPP13L1. Dev. Cell 2025, 60, p1702–p1718.e6. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.; Zhang, Y.; Qin, Z.; Wang, S.; Liu, C.; Cui, Z.; Liu, D.; Wang, H. Puccinia triticina effector Pt-1234 modulates wheat immunity by targeting transcription factor TaNAC069 via its C subdomain. Crop J. 2025, 13, 69–78. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, S.; Liu, X.; Wang, G.-F. The maize ZmVPS23-like protein relocates the nucleotide-binding leucine-rich repeat protein Rp1-D21 to endosomes and suppresses the defense response. Plant Cell 2023, 35, 2369–2390. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Li, M.; Dong, L.; Zhang, H.; Zhang, D.; Lu, P.; Wu, Q.; Xie, J.; Chen, Y.; Guo, G.; et al. An atypical NLR pair TdCNL1/TdCNL5 from wild emmer confers powdery mildew resistance in wheat. Nat. Genet. 2025, 57, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, H.; Ding, Z.; Yan, J.; Yu, H.; Pan, R.; Hu, J.; Guan, Y.; Hua, J. Low temperature enhances plant immunity via salicylic acid pathway genes that are repressed by ethylene. Plant Physiol. 2019, 182, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhu, K.; Shen, X.; Meng, J.; Huang, X.; Tan, Y.; Cardinale, F.; Liu, J.; Li, G.; Liu, J. Two interacting ethylene response factors negatively regulate peach resistance to Lasiodiplodia theobromae. Plant Physiol. 2023, 192, 3134–3151. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, C.; Du, J.; Sun, Y.; Hu, R.; Liu, S.; Xu, Q.; He, X.; Tang, C.-X.; Xu, R.; et al. DEAD-box protein SMA1 activates immunity likely through the formation of nuclear condensates with EDS1 in Arabidopsis. Cell Rep. 2025, 44, 115895. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, J.; Lu, W.; Deng, D. Gibberellin in plant height control: Old player, new story. Plant Cell Rep. 2017, 36, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Li, Y.; Cheng, Z.; Zheng, X.; Cai, C.; Wang, H.; Lu, K.; Zhu, C.; Ding, Y. Important Factors Controlling Gibberellin Homeostasis in Plant Height Regulation. J. Agric. Food Chem. 2023, 71, 15895–15907. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, B.; Yang, X.; Zhang, R.; Dong, S.; Liu, Y.; Jiang, H.; Wu, G.; Ding, T. ZmNLR-7-Mediated Synergistic Regulation of ROS, Hormonal Signaling, and Defense Gene Networks Drives Maize Immunity to Southern Corn Leaf Blight. Curr. Issues Mol. Biol. 2025, 47, 573. https://doi.org/10.3390/cimb47070573
Su B, Yang X, Zhang R, Dong S, Liu Y, Jiang H, Wu G, Ding T. ZmNLR-7-Mediated Synergistic Regulation of ROS, Hormonal Signaling, and Defense Gene Networks Drives Maize Immunity to Southern Corn Leaf Blight. Current Issues in Molecular Biology. 2025; 47(7):573. https://doi.org/10.3390/cimb47070573
Chicago/Turabian StyleSu, Bo, Xiaolan Yang, Rui Zhang, Shijie Dong, Ying Liu, Hubiao Jiang, Guichun Wu, and Ting Ding. 2025. "ZmNLR-7-Mediated Synergistic Regulation of ROS, Hormonal Signaling, and Defense Gene Networks Drives Maize Immunity to Southern Corn Leaf Blight" Current Issues in Molecular Biology 47, no. 7: 573. https://doi.org/10.3390/cimb47070573
APA StyleSu, B., Yang, X., Zhang, R., Dong, S., Liu, Y., Jiang, H., Wu, G., & Ding, T. (2025). ZmNLR-7-Mediated Synergistic Regulation of ROS, Hormonal Signaling, and Defense Gene Networks Drives Maize Immunity to Southern Corn Leaf Blight. Current Issues in Molecular Biology, 47(7), 573. https://doi.org/10.3390/cimb47070573