
Associations between Sperm Epigenetic Age and Semen Parameters: An Evaluation of Clinical and Non-Clinical Cohorts

 and
and
Abstract
1. Introduction
2. Methods
2.1. Study Populations
2.2. Semen Sample Collection and Measurement of Semen Parameters
2.3. Semen Preparation and DNA Isolation
2.4. Sperm Clock Development in the Non-Clinical Cohort
2.5. Statistical Analysis
3. Results
3.1. Demographics
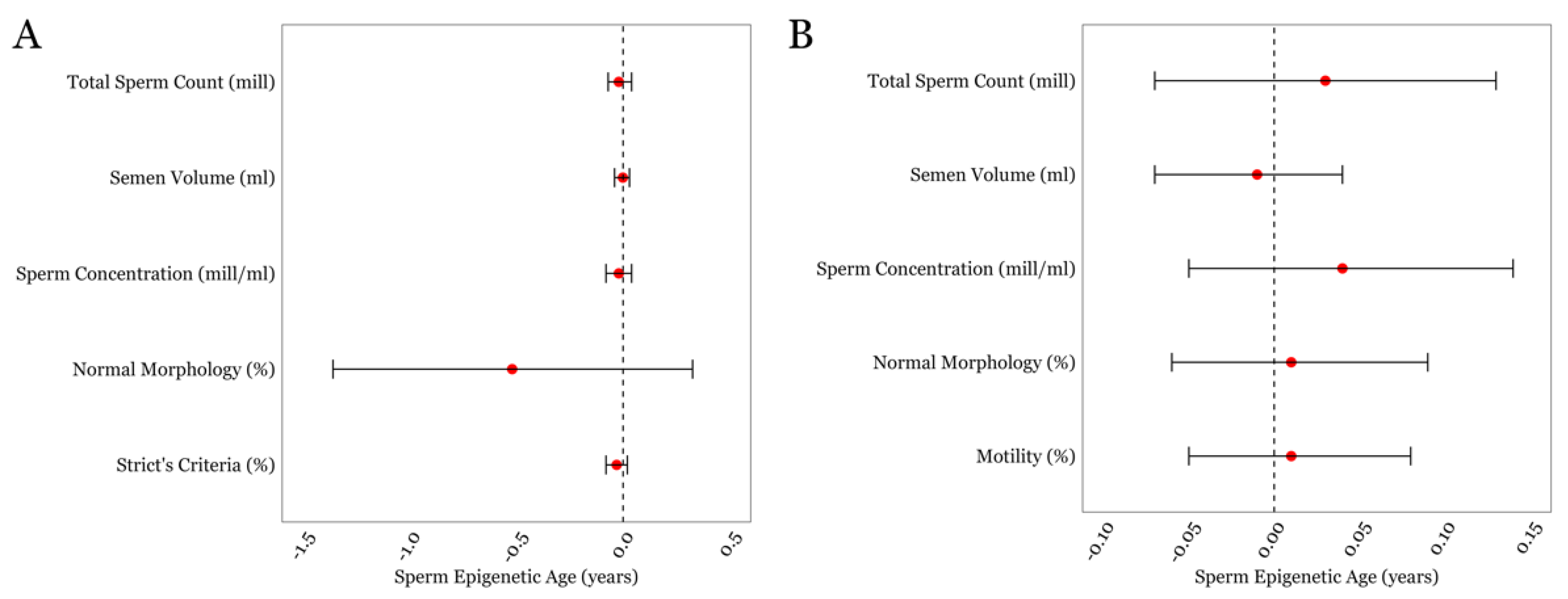
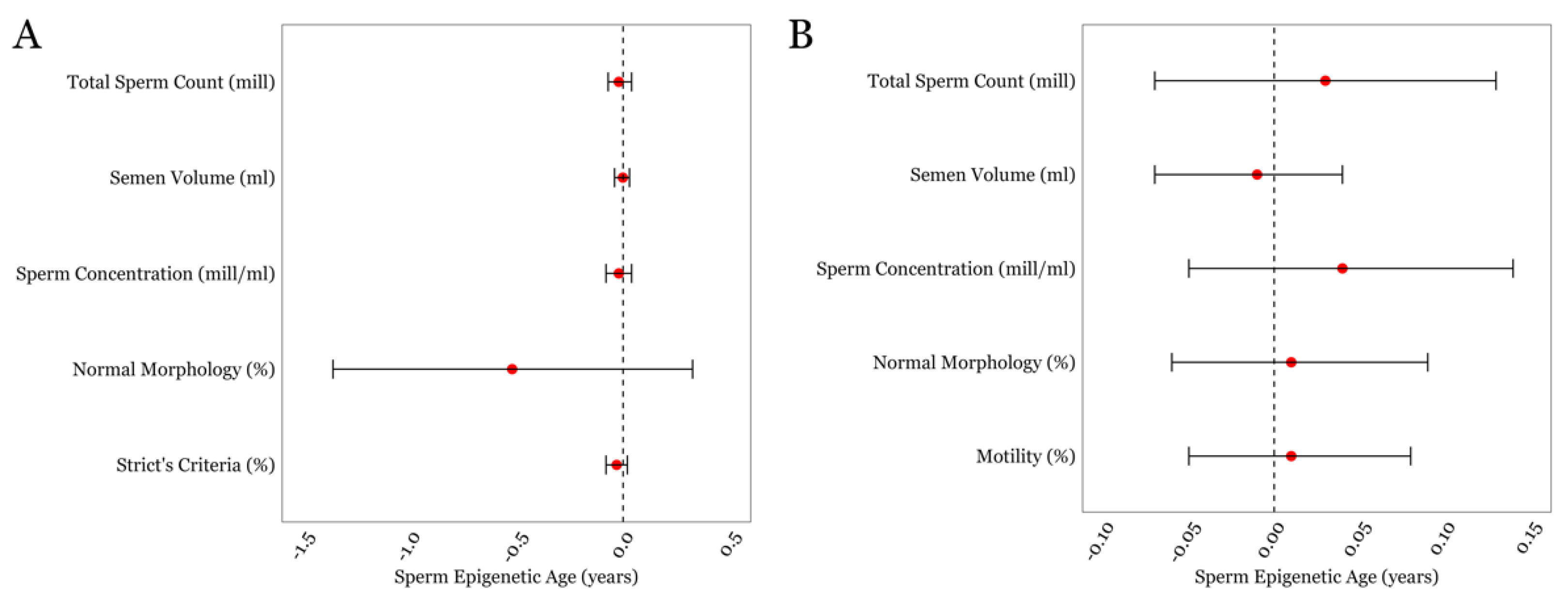
3.2. SEA and General Semen Parameters in the LIFE and SEEDS cohorts
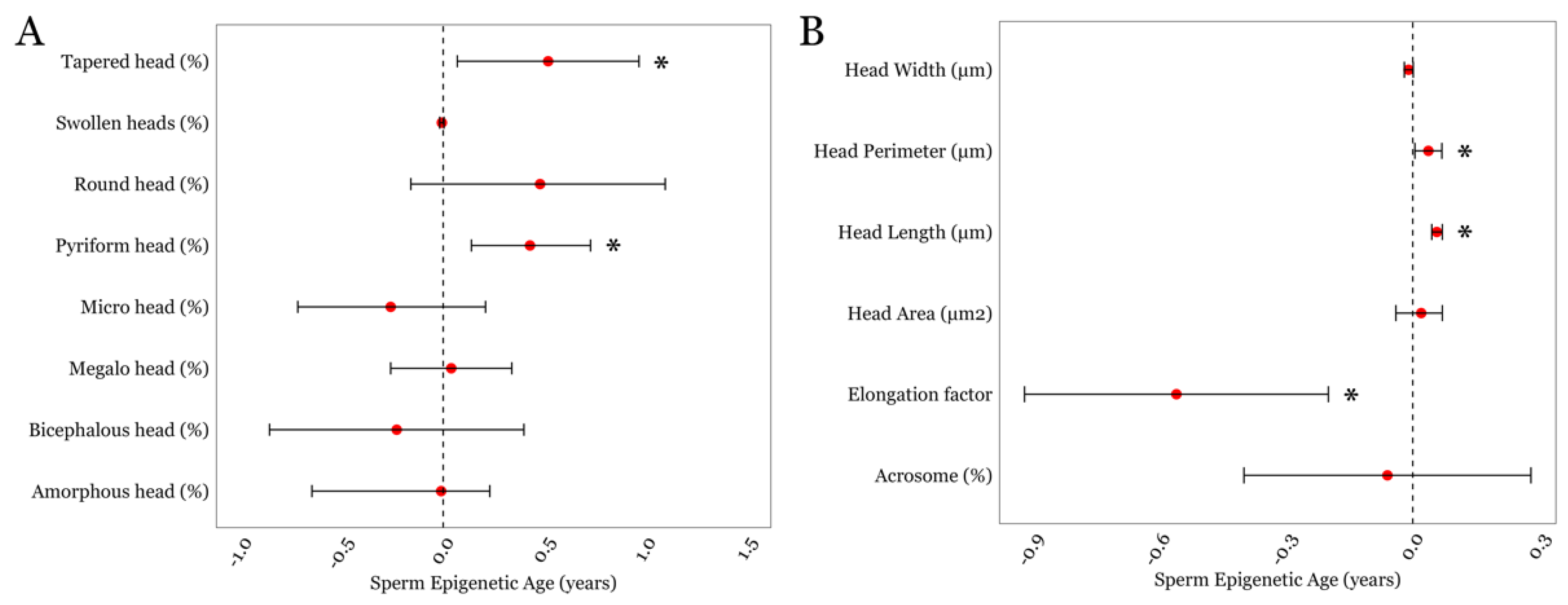
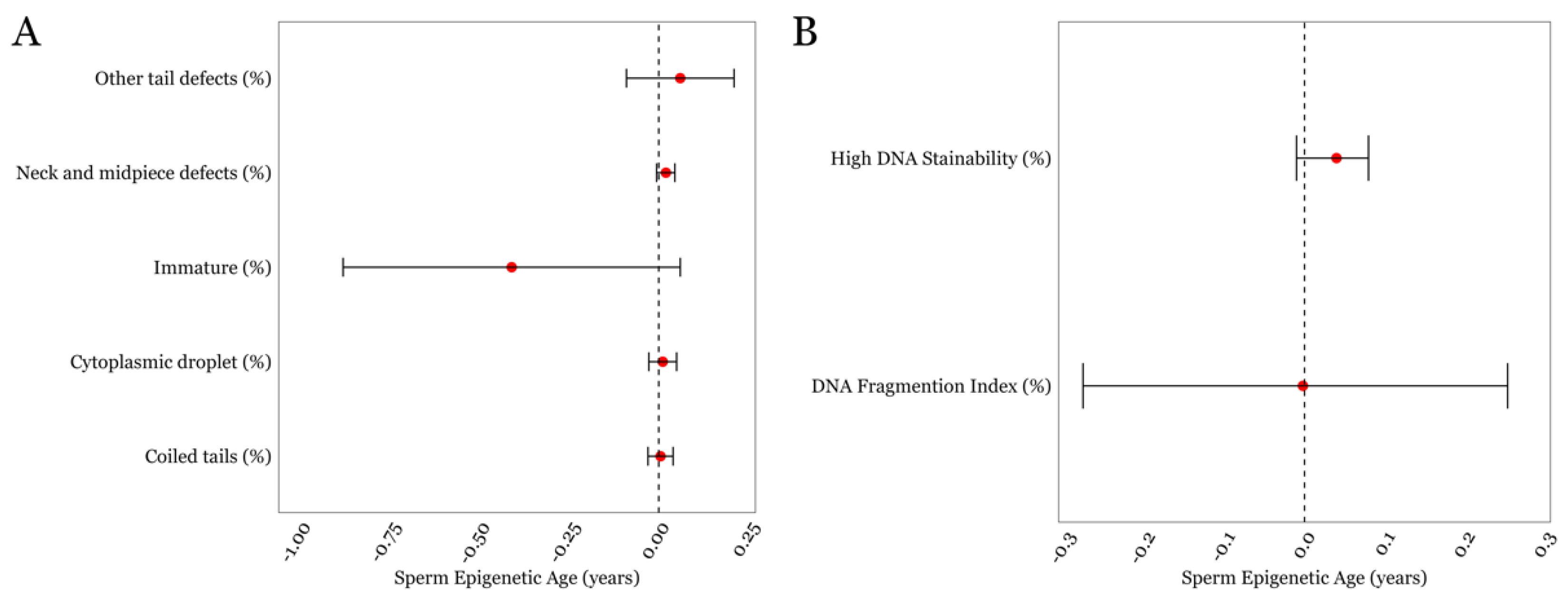
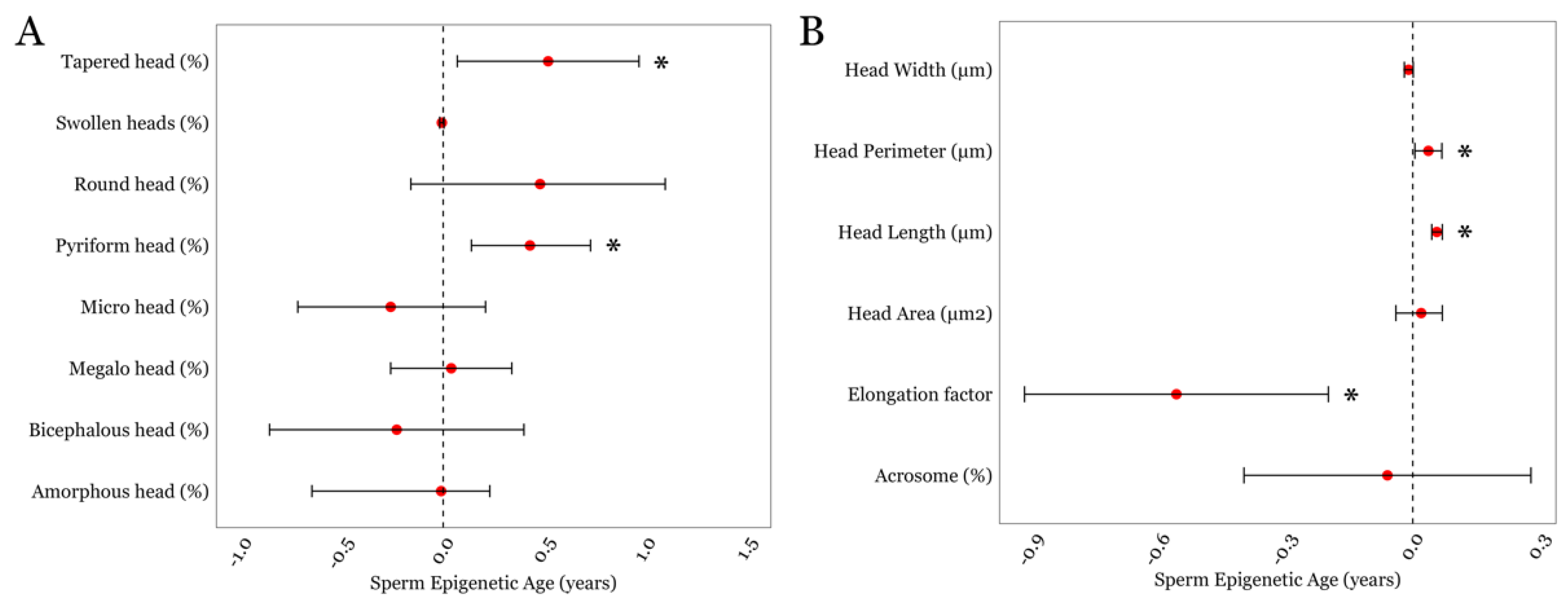
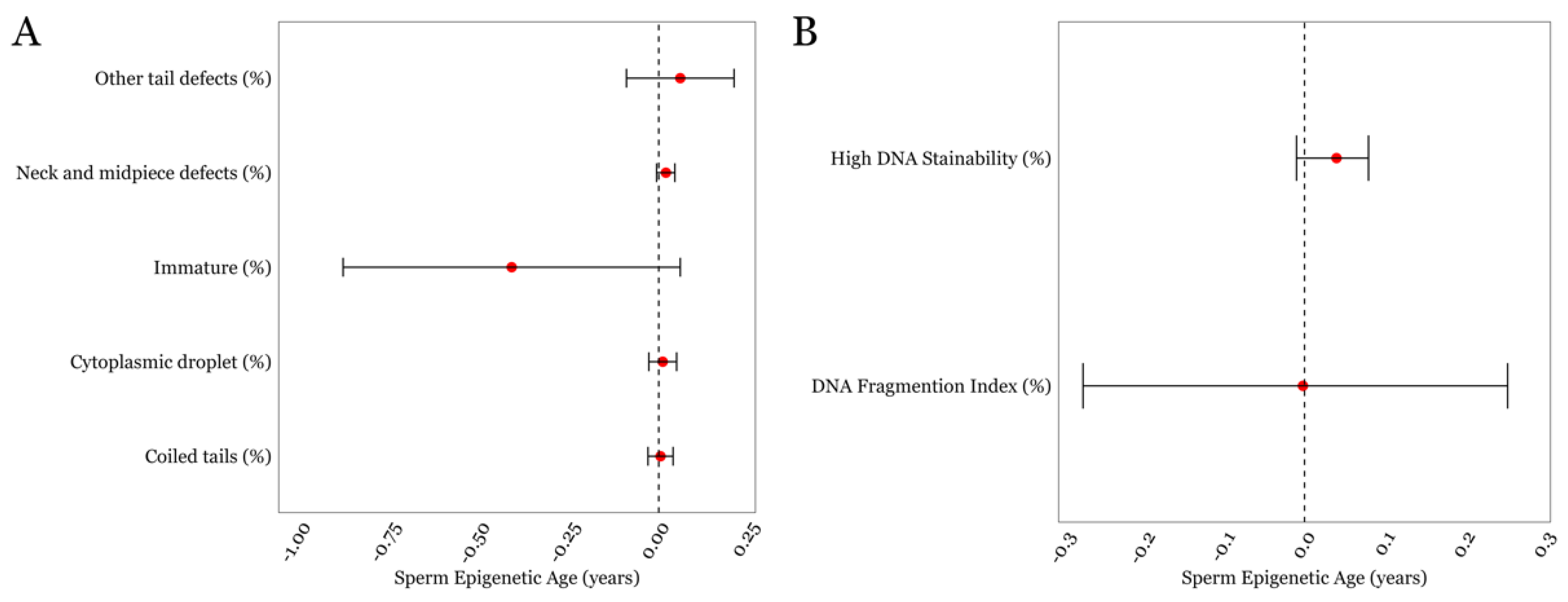
3.3. SEA and Detailed Semen Parameters in the LIFE cohort
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Infertility Prevalence Estimates, 1990–2021; World Health Organization: Geneva, Switzerland, 2023; p. 79.
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef]
- Boitrelle, F.; Shah, R.; Saleh, R.; Henkel, R.; Kandil, H.; Chung, E.; Vogiatzi, P.; Zini, A.; Arafa, M.; Agarwal, A. The Sixth Edition of the WHO Manual for Human Semen Analysis: A Critical Review and SWOT Analysis. Life 2021, 11, 1368. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.; Jorgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Jolles, M.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis of samples collected globally in the 20th and 21st centuries. Hum. Reprod. Update 2023, 29, 157–176. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.; Jorgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Bjorndahl, L.; Kirkman Brown, J.; Editorial Board Members of the WHO Laboratory Manual for the Examination and Processing of Human Semen. The sixth edition of the WHO Laboratory Manual for the Examination and Processing of Human Semen: Ensuring quality and standardization in basic examination of human ejaculates. Fertil. Steril. 2022, 117, 246–251. [Google Scholar] [CrossRef]
- Buck Louis, G.M.; Schisterman, E.F.; Sweeney, A.M.; Wilcosky, T.C.; Gore-Langton, R.E.; Lynch, C.D.; Boyd Barr, D.; Schrader, S.M.; Kim, S.; Chen, Z.; et al. Designing prospective cohort studies for assessing reproductive and developmental toxicity during sensitive windows of human reproduction and development—The LIFE Study. Paediatr. Perinat. Epidemiol. 2011, 25, 413–424. [Google Scholar] [CrossRef]
- Jungwirth, A.; Giwercman, A.; Tournaye, H.; Diemer, T.; Kopa, Z.; Dohle, G.; Krausz, C.; European Association of Urology Working Group on Male, I. European Association of Urology guidelines on Male Infertility: The 2012 update. Eur. Urol. 2012, 62, 324–332. [Google Scholar] [CrossRef]
- Bieniek, J.M.; Drabovich, A.P.; Lo, K.C. Seminal biomarkers for the evaluation of male infertility. Asian J. Androl. 2016, 18, 426–433. [Google Scholar] [CrossRef]
- Lewis, S.E. Is sperm evaluation useful in predicting human fertility? Reproduction 2007, 134, 31–40. [Google Scholar] [CrossRef]
- Guzick, D.S.; Overstreet, J.W.; Factor-Litvak, P.; Brazil, C.K.; Nakajima, S.T.; Coutifaris, C.; Carson, S.A.; Cisneros, P.; Steinkampf, M.P.; Hill, J.A.; et al. Sperm morphology, motility, and concentration in fertile and infertile men. N. Engl. J. Med. 2001, 345, 1388–1393. [Google Scholar] [CrossRef]
- Llavanera, M.; Delgado-Bermudez, A.; Ribas-Maynou, J.; Salas-Huetos, A.; Yeste, M. A systematic review identifying fertility biomarkers in semen: A clinical approach through Omics to diagnose male infertility. Fertil. Steril. 2022, 118, 291–313. [Google Scholar] [CrossRef]
- Waldenstrom, U. Postponing parenthood to advanced age. Ups. J. Med. Sci. 2016, 121, 235–243. [Google Scholar] [CrossRef]
- Oluwayiose, O.A.; Wu, H.; Saddiki, H.; Whitcomb, B.W.; Balzer, L.B.; Brandon, N.; Suvorov, A.; Tayyab, R.; Sites, C.K.; Hill, L.; et al. Sperm DNA methylation mediates the association of male age on reproductive outcomes among couples undergoing infertility treatment. Sci. Rep. 2021, 11, 3216. [Google Scholar] [CrossRef]
- Kaltsas, A.; Moustakli, E.; Zikopoulos, A.; Georgiou, I.; Dimitriadis, F.; Symeonidis, E.N.; Markou, E.; Michaelidis, T.M.; Tien, D.M.B.; Giannakis, I.; et al. Impact of Advanced Paternal Age on Fertility and Risks of Genetic Disorders in Offspring. Genes 2023, 14, 486. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P.; Djira, G.; Kasperson, K.; Christianson, J. Relationships between the age of 25,445 men attending infertility clinics and sperm chromatin structure assay (SCSA(R)) defined sperm DNA and chromatin integrity. Fertil. Steril. 2020, 114, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Lewis, S.E.M. DNA damage in testicular germ cells and spermatozoa. When and how is it induced? How should we measure it? What does it mean? Andrology 2023, 11, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- Ashapkin, V.; Suvorov, A.; Pilsner, J.R.; Krawetz, S.A.; Sergeyev, O. Age-associated epigenetic changes in mammalian sperm: Implications for offspring health and development. Hum. Reprod. Update 2023, 29, 24–44. [Google Scholar] [CrossRef]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging 2018, 10, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.B.; Gao, Y.; et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Pilsner, J.R.; Saddiki, H.; Whitcomb, B.W.; Suvorov, A.; Buck Louis, G.M.; Mumford, S.L.; Schisterman, E.F.; Oluwayiose, O.A.; Balzer, L.B. Sperm epigenetic clock associates with pregnancy outcomes in the general population. Hum. Reprod. 2022, 37, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.G.; Aston, K.I.; Cairns, B.; Smith, A.; Carrell, D.T. Paternal germ line aging: DNA methylation age prediction from human sperm. BMC Genom. 2018, 19, 763. [Google Scholar] [CrossRef] [PubMed]
- Oluwayiose, O.A.; Houle, E.; Wu, H.; Whitcomb, B.W.; Mumford, S.L.; Schisterman, E.F.; Suvorov, A.; Balzer, L.B.; Pilsner, J.R. Urinary phthalate metabolites and their mixtures are associated with advanced sperm epigenetic aging in a general population. Environ. Res. 2022, 214, 114115. [Google Scholar] [CrossRef]
- WHO. Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010; p. 271.
- Evenson, D.P.; Larson, K.L.; Jost, L.K. Sperm chromatin structure assay: Its clinical use for detecting sperm DNA fragmentation in male infertility and comparisons with other techniques. J. Androl. 2002, 23, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.G.; Noonan, E.; von Eckardstein, S.; Auger, J.; Baker, H.W.; Behre, H.M.; Haugen, T.B.; Kruger, T.; Wang, C.; Mbizvo, M.T.; et al. World Health Organization reference values for human semen characteristics. Hum. Reprod. Update 2010, 16, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; de Gannes, M.K.; Luchetti, G.; Pilsner, J.R. Rapid method for the isolation of mammalian sperm DNA. Biotechniques 2015, 58, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Suvorov, A.; Shershebnev, A.; Wu, H.; Medvedeva, Y.; Sergeyev, O.; Pilsner, J.R. Perinatal exposure to low dose 2,2′,4,4′-tetrabromodiphenyl ether (BDE-47) alters sperm DNA methylation in adult rats. Reprod. Toxicol. 2018, 75, 136–143. [Google Scholar] [CrossRef]
- Mohanty, G.; Tourzani, D.A.; Gervasi, M.G.; Houle, E.; Oluwayiose, O.; Suvorov, A.; Pilsner, J.R.; Visconti, P.E. Effects of preconception exposure to phthalates on mouse sperm capacitation parameters. Andrology 2023, 11, 1484–1494. [Google Scholar] [CrossRef]
- van der Laan, M.J.; Polley, E.C.; Hubbard, A.E. Super learner. Stat. Appl. Genet. Mol. Biol. 2007, 6, 25. [Google Scholar] [CrossRef]
- Evenson, D.P. Sperm Chromatin Structure Assay (SCSA(R)) for Fertility Assessment. Curr. Protoc. 2022, 2, e508. [Google Scholar] [CrossRef]
- Turner, K.A.; Rambhatla, A.; Schon, S.; Agarwal, A.; Krawetz, S.A.; Dupree, J.M.; Avidor-Reiss, T. Male Infertility is a Women’s Health Issue-Research and Clinical Evaluation of Male Infertility Is Needed. Cells 2020, 9, 990. [Google Scholar] [CrossRef]
- Buck Louis, G.M.; Sundaram, R.; Schisterman, E.F.; Sweeney, A.; Lynch, C.D.; Kim, S.; Maisog, J.M.; Gore-Langton, R.; Eisenberg, M.L.; Chen, Z. Semen quality and time to pregnancy: The Longitudinal Investigation of Fertility and the Environment Study. Fertil. Steril. 2014, 101, 453–462. [Google Scholar] [CrossRef]
- Kleshchev, M.; Osadchuk, L.; Osadchuk, A. Age-Related Changes in Sperm Morphology and Analysis of Multiple Sperm Defects. Front. Biosci. (Schol. Ed.) 2023, 15, 12. [Google Scholar] [CrossRef]
- Zini, A.; Phillips, S.; Courchesne, A.; Boman, J.M.; Baazeem, A.; Bissonnette, F.; Kadoch, I.J.; San Gabriel, M. Sperm head morphology is related to high deoxyribonucleic acid stainability assessed by sperm chromatin structure assay. Fertil. Steril. 2009, 91, 2495–2500. [Google Scholar] [CrossRef] [PubMed]
- Hammoud, S.S.; Nix, D.A.; Zhang, H.; Purwar, J.; Carrell, D.T.; Cairns, B.R. Distinctive chromatin in human sperm packages genes for embryo development. Nature 2009, 460, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Carrell, D.T.; Emery, B.R.; Hammoud, S. The aetiology of sperm protamine abnormalities and their potential impact on the sperm epigenome. Int. J. Androl. 2008, 31, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Bendayan, M.; Caceres, L.; Sais, E.; Swierkowski-Blanchard, N.; Alter, L.; Bonnet-Garnier, A.; Boitrelle, F. Human Sperm Morphology as a Marker of Its Nuclear Quality and Epigenetic Pattern. Cells 2022, 11, 1788. [Google Scholar] [CrossRef] [PubMed]
- Boitrelle, F.; Albert, M.; Petit, J.M.; Ferfouri, F.; Wainer, R.; Bergere, M.; Bailly, M.; Vialard, F.; Selva, J. Small human sperm vacuoles observed under high magnification are pocket-like nuclear concavities linked to chromatin condensation failure. Reprod. Biomed. Online 2013, 27, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.G., Jr.; Mauri, A.L.; Petersen, C.G.; Massaro, F.C.; Silva, L.F.; Felipe, V.; Cavagna, M.; Pontes, A.; Baruffi, R.L.; Oliveira, J.B.; et al. Large nuclear vacuoles are indicative of abnormal chromatin packaging in human spermatozoa. Int. J. Androl. 2012, 35, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Gao, X.; Li, W.; Jiang, S.; Li, R.; Hong, H.; Zhao, C.; Zhou, P.; Chen, H.; Bo, X.; et al. Stable H3K4me3 is associated with transcription initiation during early embryo development. Bioinformatics 2019, 35, 3931–3936. [Google Scholar] [CrossRef]
- Pepin, A.S.; Lafleur, C.; Lambrot, R.; Dumeaux, V.; Kimmins, S. Sperm histone H3 lysine 4 tri-methylation serves as a metabolic sensor of paternal obesity and is associated with the inheritance of metabolic dysfunction. Mol. Metab. 2022, 59, 101463. [Google Scholar] [CrossRef]
- Lismer, A.; Kimmins, S. Emerging evidence that the mammalian sperm epigenome serves as a template for embryo development. Nat. Commun. 2023, 14, 2142. [Google Scholar] [CrossRef]
- Lismer, A.; Dumeaux, V.; Lafleur, C.; Lambrot, R.; Brind’Amour, J.; Lorincz, M.C.; Kimmins, S. Histone H3 lysine 4 trimethylation in sperm is transmitted to the embryo and associated with diet-induced phenotypes in the offspring. Dev. Cell 2021, 56, 671–686.e676. [Google Scholar] [CrossRef]
- Jiang, H.; Huang, C.J. Aberrant protamination in sperm correlates to anomalous nuclear and cytoplasmic architectures in infertile males with sperm dysmorphology. Asian J. Androl. 2023, 26, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Houle, E.; Li, Y.; Schroder, M.; McRitchie, S.L.; Rahil, T.; Sites, C.K.; Jenkins Sumner, S.; Pilsner, J.R. Exploring the internal exposome of seminal plasma with semen quality and live birth: A Pilot Study. Syst. Biol. Reprod. Med. 2023, 69, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Amirjannati, N.; Henkel, R.; Hosseini, E.; Choopanian, P.; Moghadasfar, H.; Arjmand, B.; Asgharpour Sarouey, L.; Haji Parvaneh, A.; Gilany, K. The Amino Acid Profile in Seminal Plasma of Normozoospermic Men: A Correlation Analysis with Spermiogram Parameters and Total Antioxidant Capacity. J. Reprod. Infertil. 2023, 24, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Rosati, A.J.; Whitcomb, B.W.; Brandon, N.; Buck Louis, G.M.; Mumford, S.L.; Schisterman, E.F.; Pilsner, J.R. Sperm mitochondrial DNA biomarkers and couple fecundity. Hum. Reprod. 2020, 35, 2619–2625. [Google Scholar] [CrossRef] [PubMed]
- Deonandan, R.; Jaleel, M. Global decline in semen quality: Ignoring the developing world introduces selection bias. Int. J. Gen. Med. 2012, 5, 303–306. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| LIFE | SEEDS | |||
|---|---|---|---|---|
| (Mean ± SD or n (%)) | ||||
| Age (years) | 31.9 ± 4.8 | 36.1 ± 6.5 | ||
| BMI (kg/m2) 1 | 29.9 ± 5.7 | 29.7 ± 5.5 | ||
| Normal (18–25) | 67 (18) | 38 (20) | ||
| Overweight (>25 <30) | 156 (42) | 80 (42) | ||
| Obese (>30) | 151 (40) | 74 (38) | ||
| Race 1 | ||||
| White (non-Hispanic) | 307 (81) | 139 (72) | ||
| Black (non-Hispanic) | 15 (4) | 8 (4) | ||
| Hispanic | 32 (9) | 11 (6) | ||
| Others/Unknown | 23 (6) | 34 (18) | ||
| Smoking status 2 | ||||
| No | 295 (79) | 162 (84) | ||
| Yes | 80 (21) | 30 (16) | ||
| Semen Parameters | Mean ± SD | % < WHO 3 | Mean ± SD | % < WHO |
| Sperm count (millions) | 74.5 ± 55 | 7.4 | 92.9 ± 27.7 | 4.6 |
| Sperm volume (mL) | 3.4 ± 1.7 | 8.7 | 2.96 ± 1.4 | 18.2 |
| Sperm concentration (millions/mL) | 228.4 ± 117 | 7.7 | 78.4 ± 84.4 | 9.1 |
| Normal morphology (%) | 30 ± 12.2 | 1.1 | 6.4 ± 4.5 | 42.2 |
| Sperm motility (%) 4 | --- | 56.9 ± 20.0 | 9.1 | |
| DNA fragmentation index (%) 5 | 15.3 ± 10.4 | 13.3 | --- | |
| High DNA stainability (%) 5 | 7.4 ± 5.1 | 10.3 | --- | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawant, S.; Oluwayiose, O.A.; Nowak, K.; Maxwell, D.L.; Houle, E.; Paskavitz, A.L.; Saddiki, H.; Bertolla, R.P.; Pilsner, J.R. Associations between Sperm Epigenetic Age and Semen Parameters: An Evaluation of Clinical and Non-Clinical Cohorts. Curr. Issues Mol. Biol. 2024, 46, 1567-1578. https://doi.org/10.3390/cimb46020101
Sawant S, Oluwayiose OA, Nowak K, Maxwell DL, Houle E, Paskavitz AL, Saddiki H, Bertolla RP, Pilsner JR. Associations between Sperm Epigenetic Age and Semen Parameters: An Evaluation of Clinical and Non-Clinical Cohorts. Current Issues in Molecular Biology. 2024; 46(2):1567-1578. https://doi.org/10.3390/cimb46020101
Chicago/Turabian StyleSawant, Savni, Oladele A. Oluwayiose, Karolina Nowak, DruAnne L. Maxwell, Emily Houle, Amanda L. Paskavitz, Hachem Saddiki, Ricardo P. Bertolla, and J. Richard Pilsner. 2024. "Associations between Sperm Epigenetic Age and Semen Parameters: An Evaluation of Clinical and Non-Clinical Cohorts" Current Issues in Molecular Biology 46, no. 2: 1567-1578. https://doi.org/10.3390/cimb46020101
APA StyleSawant, S., Oluwayiose, O. A., Nowak, K., Maxwell, D. L., Houle, E., Paskavitz, A. L., Saddiki, H., Bertolla, R. P., & Pilsner, J. R. (2024). Associations between Sperm Epigenetic Age and Semen Parameters: An Evaluation of Clinical and Non-Clinical Cohorts. Current Issues in Molecular Biology, 46(2), 1567-1578. https://doi.org/10.3390/cimb46020101