
gjSOX9 Cloning, Expression, and Comparison with gjSOXs Family Members in Gekko japonicus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Tissues Collected
2.2. Total RNA Extraction and cDNA Synthesis of gjSOX9
2.3. cDNA Fragment Cloning and Sequence Analyses
2.4. Cloning of the Full Length of gjSOX9 cDNA Using RACE Technology
2.5. gjSOX9 Expression Pattern Analysis
2.6. Physicochemical Property Analyses and Phylogenetic Reconstruction of gjSOX9
2.7. Analysis of Conserved Regions of gjSOX9 and the gjSOX Family
2.8. gjSOX9 Evolution within the gjSOXs Family and Family Orthologs Among Species
3. Results
3.1. gjSOX9 Sequence and Physiochemical Properties
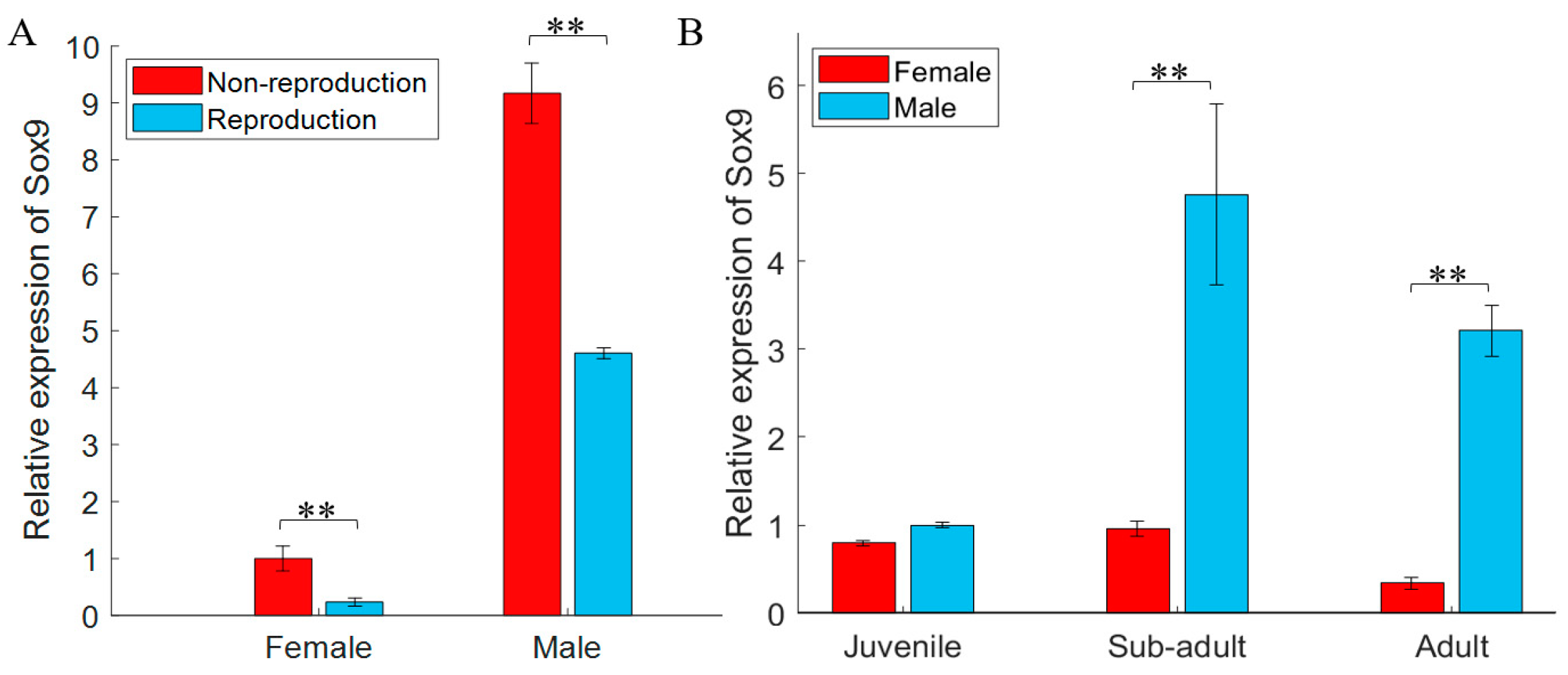
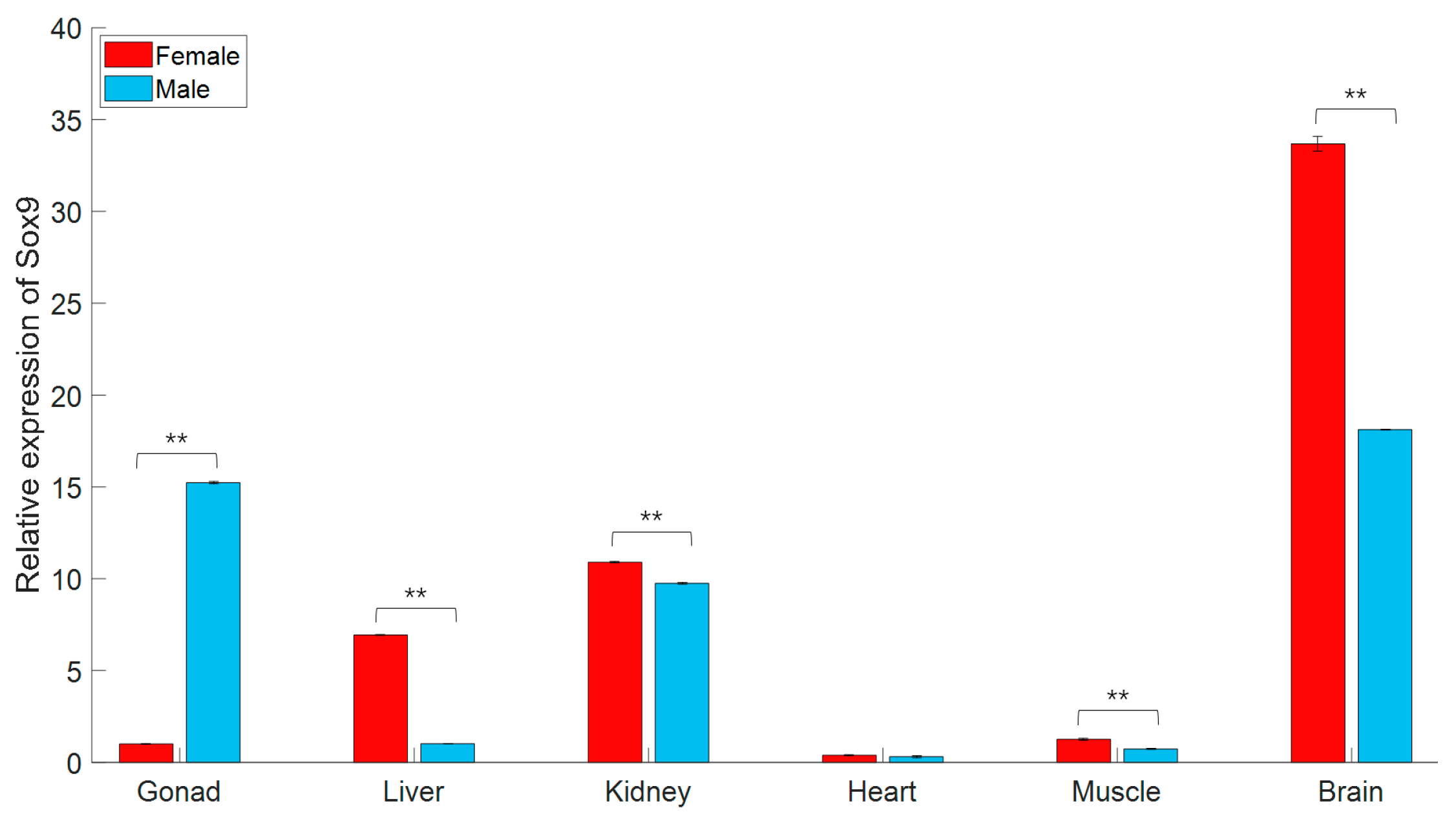
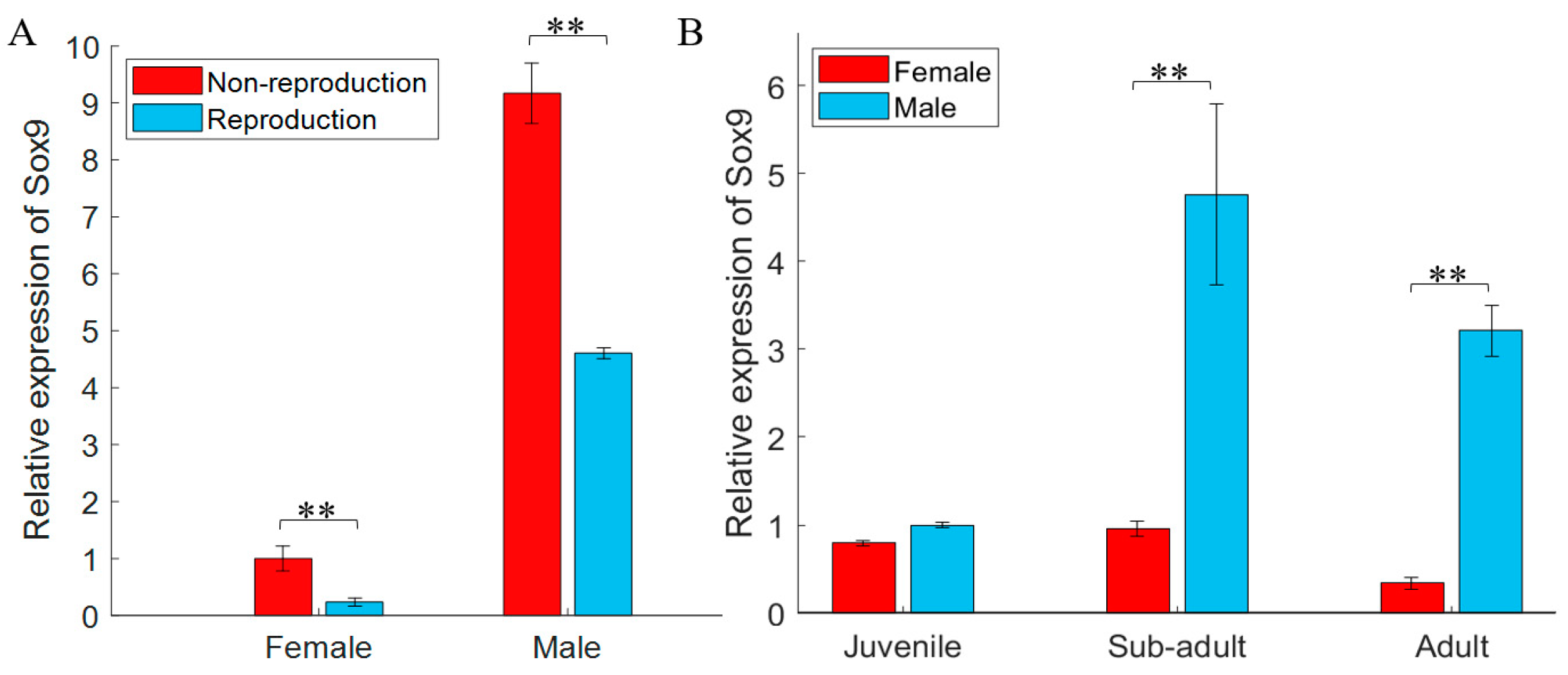
3.2. gjSOX9 Expression Patterns in G. japonicus
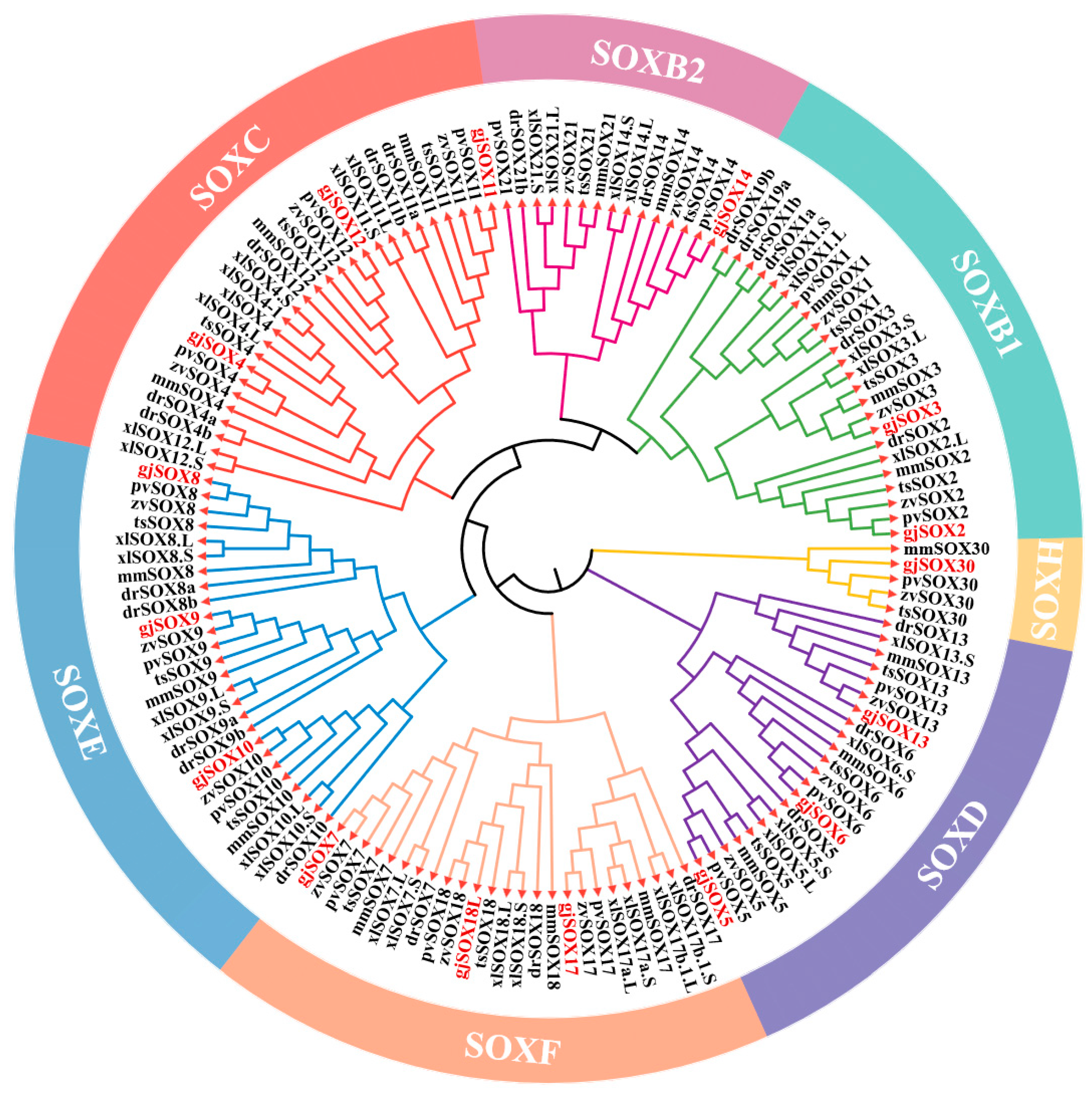
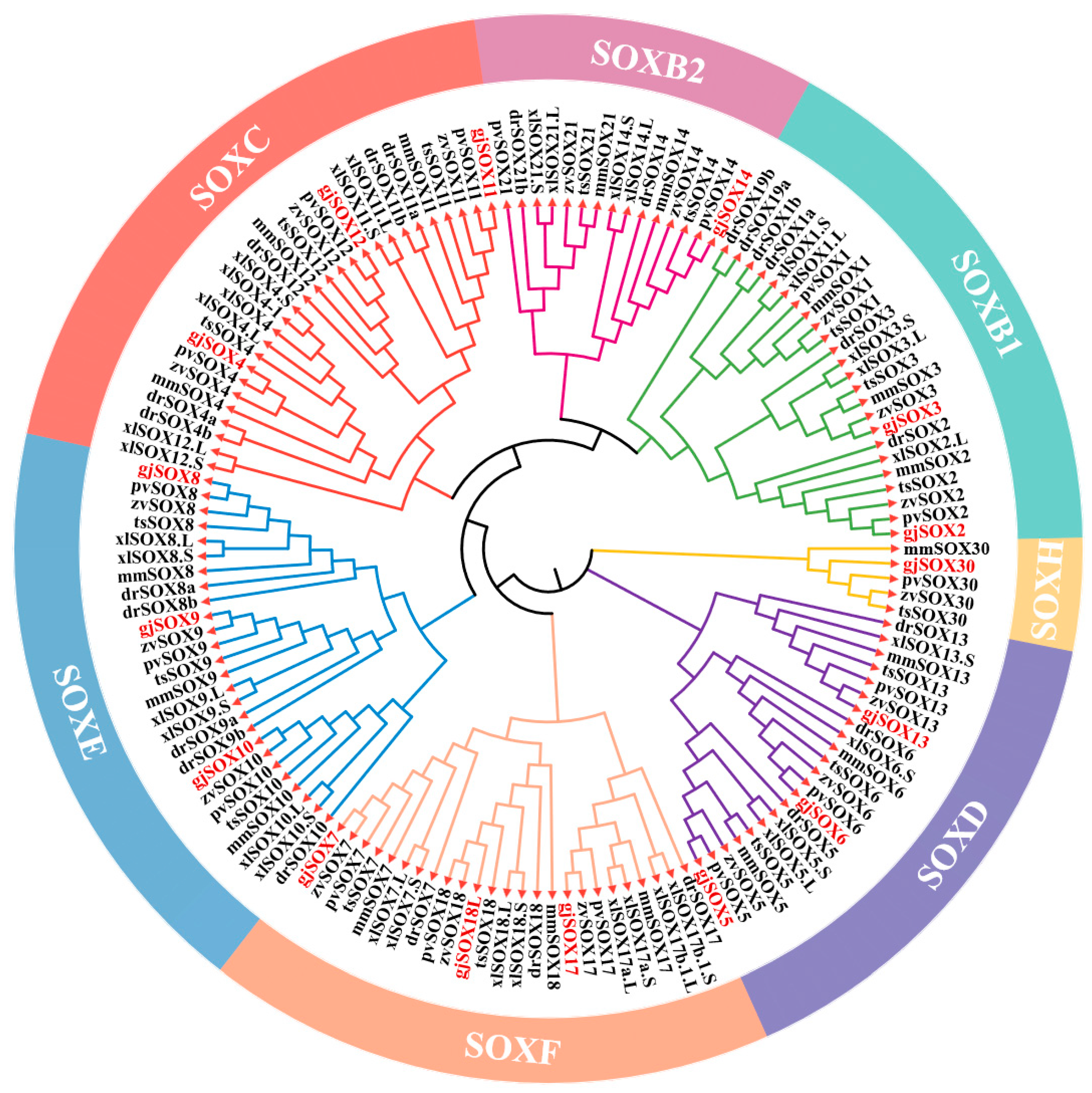
3.3. gjSOX9 Phylogenetic Analyses
3.4. Comparison with gjSOX9 and gjSOX Family
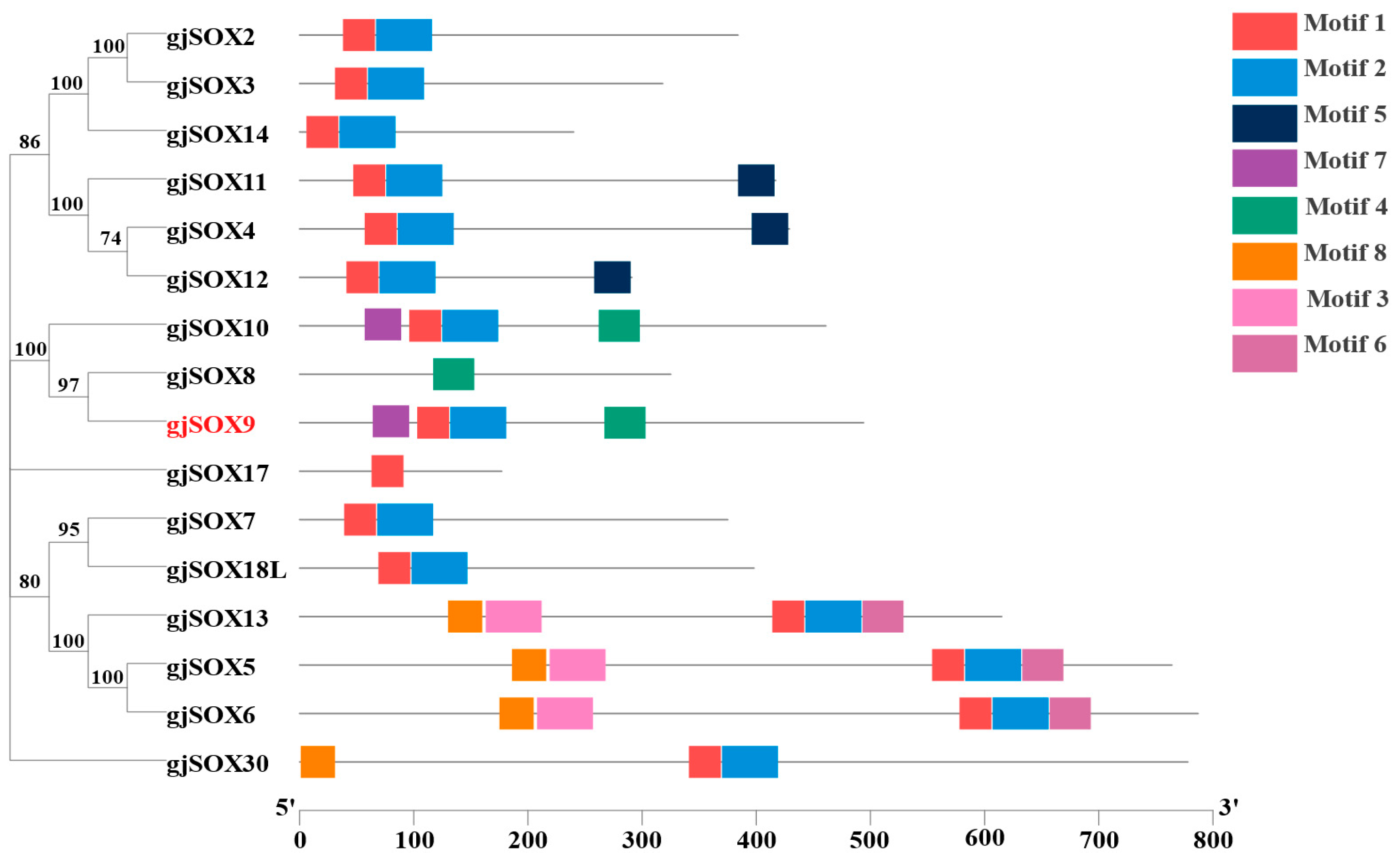
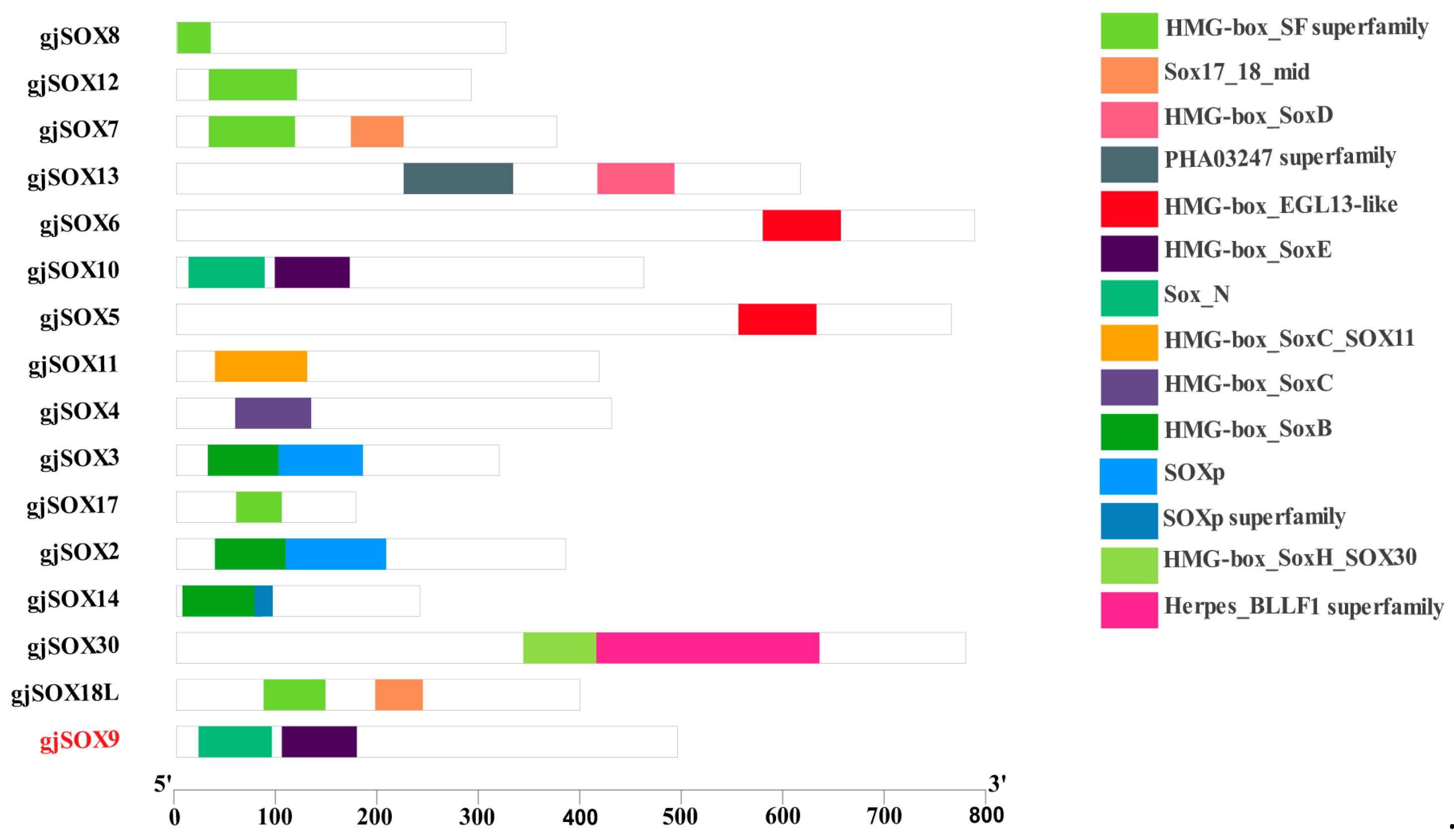
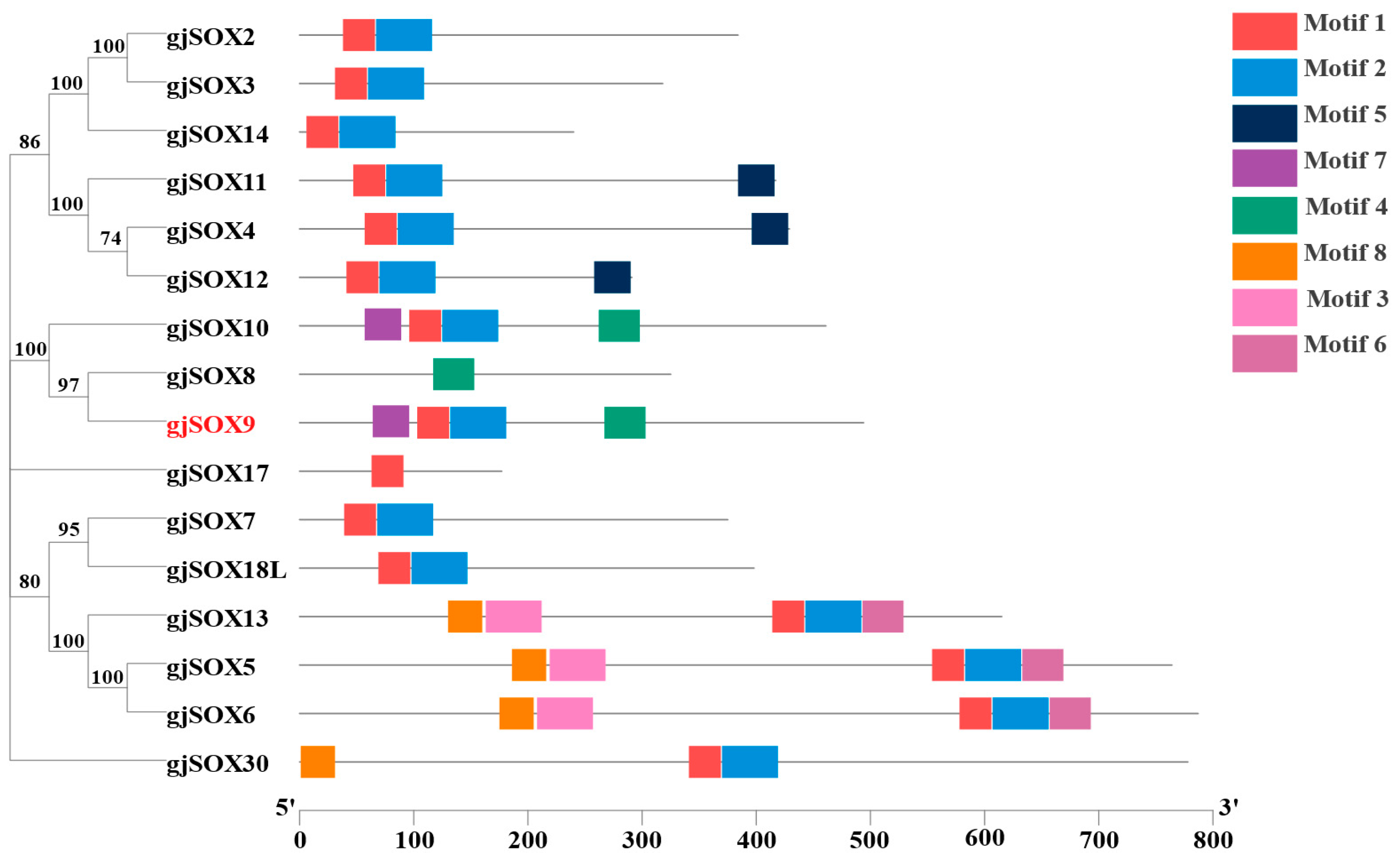
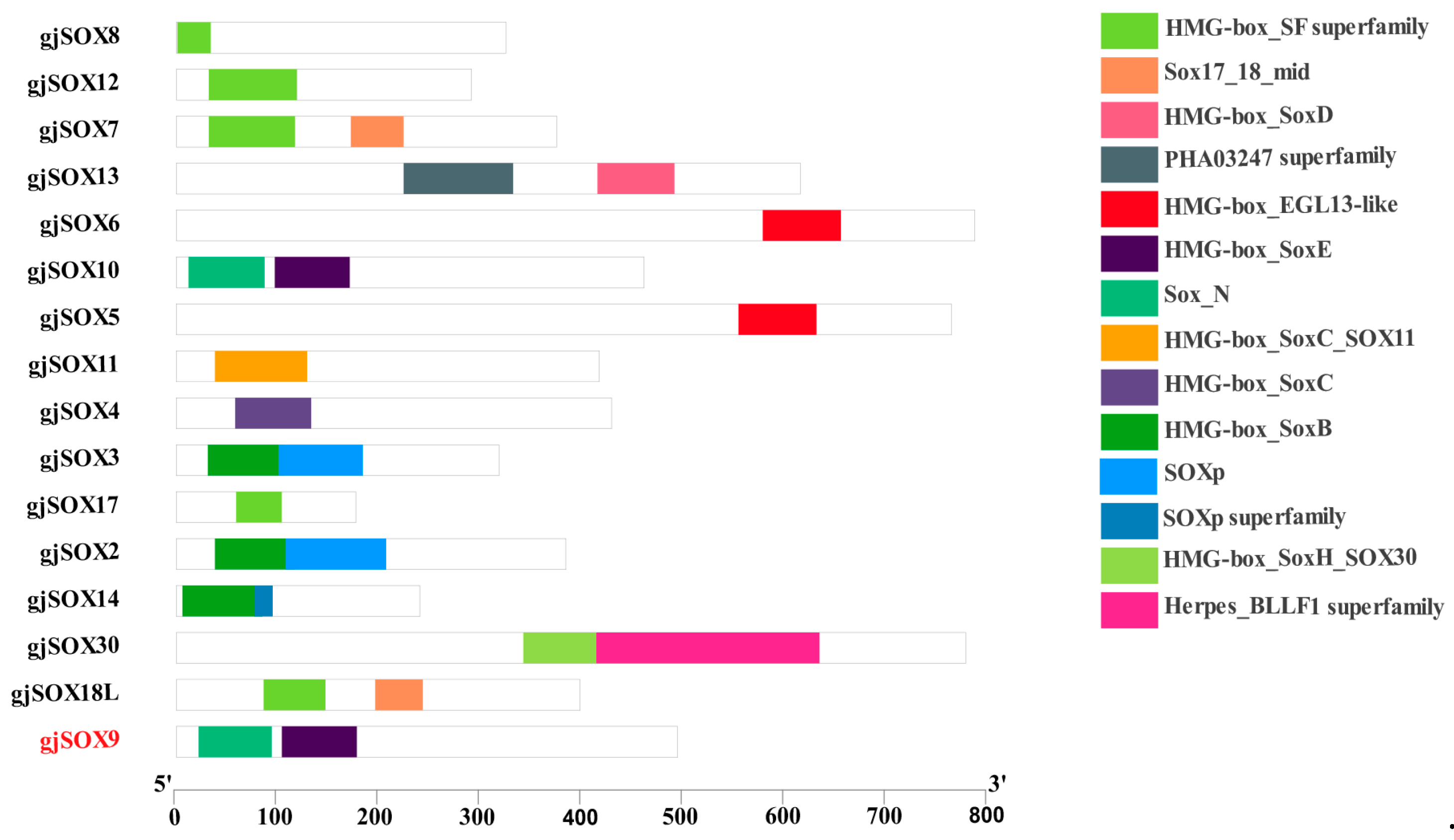
3.5. Motif and Domain Analyses of gjSOX9 and gjSOX Family
3.6. Evolution Analyses of gjSOX9 and Its Family Members among Species
4. Discussion
4.1. gjSOX9 cDNAs Sequence Analyses
4.2. gjSOX9 Expression Patterns in G. japonicus
4.3. Variation in Conserved Region in gjSOX9 and Its Family Members
4.4. Evolutionary Analyses of SOX9 and Its Family Members among Species
4.5. Future Investigations of gjSOX9 in G. japonicus
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Akinyemi, M.O.; Finucan, J.; Grytsay, A.; Osaiyuwu, O.H.; Adegbaju, M.S.; Ogunade, I.M.; Thomas, B.N.; Peters, S.O.; Morenikeji, O.B. Molecular evolution and inheritance pattern of sox gene family among bovidae. Genes 2022, 13, 1783. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, R.; Gonen, N.; Sinclair, A. Sox genes and their role in disorders of sex development. Sex. Dev. 2022, 16, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Anitha, A.; Senthilkumaran, B. Role of sox family genes in teleostean reproduction-an overview. Biol. Reprod. 2021, 1, 22–31. [Google Scholar] [CrossRef]
- Bowles, J.; Schepers, G.; Koopman, P. Phylogeny of the sox family of developmental transcription factors based on sequence and structural indicators. Dev. Biol. 2000, 227, 239–255. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Luo, X.; Qu, C.; Xu, T.; Zou, G.; Liang, H. The important role of sex-related sox family genes in the sex reversal of the chinese soft-shelled turtle (Pelodiscus sinensis). Biology 2022, 11, 83. [Google Scholar] [CrossRef] [PubMed]
- Gonen, N.; Lovell-Badge, R. The regulation of sox9 expression in the gonad. Curr. Top. Dev. Biol. 2019, 134, 223–252. [Google Scholar]
- She, Z.Y.; Yang, W.X. Sox family transcription factors involved in diverse cellular events during development. Eur. J. Cell Biol. 2015, 94, 547–563. [Google Scholar] [CrossRef]
- Chiang, E.F.; Pai, C.I.; Wyatt, M.; Yan, Y.L.; Postlethwait, J.; Chung, B. Two sox9 genes on duplicated zebrafish chromosomes: Expression of similar transcription activators in distinct sites. Dev. Biol. 2001, 231, 149–163. [Google Scholar] [CrossRef]
- Hui, H.B.; Xiao, L.; Sun, W.; Zhou, Y.J.; Zhang, H.Y.; Ge, C.T. Sox9 is indispensable for testis differentiation in the red-eared slider turtle, a reptile with temperature-dependent sex determination. Zool. Res. 2021, 42, 721–725. [Google Scholar] [CrossRef]
- Gonen, N.; Futtner, C.R.; Wood, S.; Garcia-Moreno, S.A.; Salamone, I.M.; Samson, S.C.; Sekido, R.; Poulat, F.; Maatouk, D.M.; Lovell-Badge, R. Sex reversal following deletion of a single distal enhancer of sox9. Science 2018, 360, 1469–1473. [Google Scholar] [CrossRef]
- Li, X.; Yu, H.; Wang, Y.; Liu, X.; Liu, Y.; Qu, J.; Wang, X. Roles of two sox9 genes during gonadal development in Japanese flounder: Sex differentiation, spermatogenesis and gonadal function maintenance. Int. J. Mol. Sci. 2018, 19, 512. [Google Scholar] [CrossRef] [PubMed]
- Voldoire, E.; Brunet, F.; Naville, M.; Volff, J.N.; Galiana, D. Expansion by whole genome duplication and evolution of the sox gene family in teleost fish. PLoS ONE 2017, 12, e0180936. [Google Scholar] [CrossRef]
- Vining, B.; Ming, Z.; Bagheri-Fam, S.; Harley, V. Diverse regulation but conserved function: Sox9 in vertebrate sex determination. Genes 2021, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Hou, C.C.; She, Z.Y.; Yang, W.X. The sox gene family: Function and regulation in testis determination and male fertility maintenance. Mol. Biol. Rep. 2013, 40, 2187–2194. [Google Scholar] [CrossRef]
- Rhen, T.; Schroeder, A. Molecular mechanisms of sex determination in reptiles. Sex. Dev. 2010, 4, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.E.; Wynn, S.L.; Sesay, A.; Cruz, C.; Cheung, M.; Gomez Gaviro, M.V.; Booth, S.; Gao, B.; Cheah, K.S.; Lovell-Badge, R.; et al. Sox9 induces and maintains neural stem cells. Nat. Neurosci. 2010, 13, 1181–1189. [Google Scholar] [CrossRef]
- Wang, L.; Cai, R.; Liu, F.; Lv, Y.; Zhang, Y.; Duan, S.; Izaz, A.; Zhou, J.; Wang, H.; Duan, R.; et al. Molecular cloning, characterization, mRNA expression changes and nucleocytoplasmic shuttling during kidney embryonic development of sox9 in Alligator sinensis. Gene 2020, 731, 144334. [Google Scholar] [CrossRef]
- Lefebvre, V.; Angelozzi, M.; Haseeb, A. Sox9 in cartilage development and disease. Curr. Opin. Cell Biol. 2019, 61, 39–47. [Google Scholar] [CrossRef]
- Zafar, I.; Iftikhar, R.; Ahmad, S.U.; Rather, M.A. Genome wide identification, phylogeny, and synteny analysis of sox gene family in common carp (Cyprinus carpio). Biotechnol. Rep. 2021, 30, e00607. [Google Scholar] [CrossRef]
- Yao, C.; Wan, H.; Zhang, Z.; Lin, J.; Wang, Y. Genome-wide identification and expression profile of the sox gene family in different tissues and during embryogenesis in the pacific white shrimp (Litopenaeus vannamei). Gene 2020, 763, 144956. [Google Scholar] [CrossRef]
- Wan, H.; Liao, J.; Zhang, Z.; Zeng, X.; Liang, K.; Wang, Y. Molecular cloning, characterization, and expression analysis of a sex-biased transcriptional factor sox9 gene of mud crab Scylla paramamosain. Gene 2021, 774, 145423. [Google Scholar] [CrossRef] [PubMed]
- Adolfi, M.C.; Carreira, A.C.; Jesus, L.W.; Bogerd, J.; Funes, R.M.; Schartl, M.; Sogayar, M.C.; Borella, M.I. Molecular cloning and expression analysis of dmrt1 and sox9 during gonad development and male reproductive cycle in the lambari fish, Astyanax altiparanae. Reprod. Biol. Endocrinol. 2015, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, Z.; Luo, L.; Ping, J.; Zhou, H.; Xie, L.; Zhang, Y. Latitudinal variation in the pattern of temperature-dependent sex determination in the Japanese Gecko, Gekko japonicus. Animals 2022, 12, 942. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.H. Temperature Dependent Sex Determination (TSD) for Gekko japonicus. Ph.D. Thesis, Nanjing Normal University, Nanjing, China, 2012. (In Chinese). [Google Scholar]
- Chen, J.C. Karyotype studies of three gecko species in genus Gekko. Asian Herpetol. Res. 1986, 5, 24–29. (In Chinese) [Google Scholar]
- Yoshida, M.; Itoh, M. Karyotype of the gecko, Gekko japonicus. Cis 1974, 17, 29–31. [Google Scholar]
- Liu, Y.; Zhou, Q.; Wang, Y.; Luo, L.; Yang, J.; Yang, L.; Liu, M.; Li, Y.; Qian, T.; Zheng, Y.; et al. Gekko japonicus genome reveals evolution of adhesive toe pads and tail regeneration. Nat. Commun. 2015, 6, 10033. [Google Scholar] [CrossRef]
- Xin, H.; Wang, X.; Tang, S.; Lu, W.; Li, Y.; Zhang, S.; Liang, S.; Chen, Q. Characterization and Expression Analysis of Sox9 in the Multiocellated Racerunner, Eremias multiocellata. Asian Herpetol. Res. 2014, 5, 104–112. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, 200–204. [Google Scholar] [CrossRef]
- Yang, M.; Derbyshire, M.K.; Yamashita, R.A.; Marchler-Bauer, A. NCBI’s conserved domain database and tools for protein domain analysis. Curr. Protoc. Bioinform. 2020, 69, e90. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.S.; Hu, W.; Liu, X.C.; Lin, H.R.; Zhu, Z.Y. Molecular cloning and mRNA expression pattern of sox9 during sex reversal in orange-spotted grouper (Epinephelus coioides). Aquaculture 2010, 306, 322–328. [Google Scholar] [CrossRef]
- Dumond, H.; Al-Asaad, I.; Chesnel, A.; Chardard, D.; Boizet-Bonhoure, B.; Flament, S.; Kuntz, S. Temporal and spatial sox9 expression patterns in the course of gonad development of the caudate amphibian Pleurodeles waltl. J. Exp. Zool. B Mol. Dev. Evol. 2011, 316b, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Heenan, P.; Zondag, L.; Wilson, M.J. Evolution of the sox gene family within the Chordate phylum. Gene 2016, 575, 385–392. [Google Scholar] [CrossRef]
- Schepers, G.E.; Teasdale, R.D.; Koopman, P. Twenty pairs of sox: Extent, homology, and nomenclature of the mouse and human sox transcription factor gene families. Dev. Cell 2002, 3, 167–170. [Google Scholar] [CrossRef]
- Koopman, P.; Schepers, G.; Brenner, S.; Venkatesh, B. Origin and diversity of the sox transcription factor gene family: Genome-wide analysis in Fugu rubripes. Gene 2004, 328, 177–186. [Google Scholar] [CrossRef]
- Baudouin-Gonzalez, L.; Schoenauer, A.; Harper, A.; Blakeley, G.; Seiter, M.; Arif, S.; Sumner-Rooney, L.; Russell, S.; Sharma, P.P.; McGregor, A.P. The evolution of sox gene repertoires and regulation of segmentation in arachnids. Mol. Biol. Evol. 2021, 38, 3153–3169. [Google Scholar] [CrossRef]
- Huang, X.; Wu, C.; Gong, K.; Chen, Q.; Gu, Q.; Qin, H.; Zhao, C.; Yu, T.; Yang, L.; Fu, W.; et al. Sox gene family revealed genetic variations in autotetraploid Carassius auratus. Front. Genet. 2020, 11, 804. [Google Scholar] [CrossRef]
- Wegner, M. All purpose SOX: The many roles of SOX proteins in gene expression. Int. J. Biochem. Cell Biol. 2010, 42, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Liu, J.; Rao, Y. Whole genome resequencing helps study important traits in chickens. Genes 2023, 14, 1198. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Huang, L.; Bai, H.; Wang, Z.; Bi, Y.; Chen, G.; Jiang, Y.; Chang, G. Genome-wide association study of potential meat quality trait loci in Ducks. Genes 2022, 13, 986. [Google Scholar] [CrossRef] [PubMed]
- Dehshahri, A.; Biagioni, A.; Bayat, H.; Lee, E.H.C.; Hashemabadi, M.; Fekri, H.S.; Zarrabi, A.; Mohammadinejad, R.; Kumar, A.P. Editing sox genes by CRISPR-Cas: Current insights and future perspectives. Int. J. Mol. Sci. 2021, 22, 11321. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence | Usage |
|---|---|---|
| gjSOX9F1 | TTCCGAGAYGTGGACATTGG | partial sequences acquired |
| gjSOX9R1 | GGGCCTYTGGWTGGGRYTCATGT | partial sequences acquired |
| gjSOX9-5′GSP1 | CAGATTCACCCCAGTCTTCGTC | 5′RACE |
| gjSOX9-5′GSP2 | CCCACATTGACTTCCGAGACG | 5′RACE |
| gjSOX9-3′GSP3 | TCCAGTCGTAGCCCTTCAGCACCT | 3′RACE |
| gjSOX9-3′GSP4 | CTTGTCCTCGTCGCTCTCCTTCTTC | 3′RACE |
| UPM | CTAATACGACTCACTATAGGGCAAGC AGTGGTATCAACGCAGAGT | 5′/3′RACE |
| NUP | CTAATACGACTCACTATAGGGC | 5′/3′RACE |
| M13F | GTAAAACGACGGCCAGT | clone analysis |
| M13R | CAGGAAACAGCTATGAC | clone analysis |
| gjSOX9-RT-F | CGTCTGGATGTGTAAGC | qPCR |
| gjSOX9-RT-R | TGATGTGTGTCCTCTGC | qPCR |
| Actin-F | CCCCAAAGCCAACAGAGA | qPCR |
| Actin-R | ACGCCATCACCAGAGTCCA | qPCR |
| Item Predicted | Software/Database | Web Site |
|---|---|---|
| ORF (open reading frame) | ORF finder | www.ncbi.nlm.nih.gov/projects/gorf (accessed on 21 October 2023) |
| Protein sequence analyses | Translate tool | http://web.expasy.org/translate/ (accessed on 21 October 2023) |
| Physicochemical properties | ProtParam | http://web.expasy.org/protparam (accessed on 21 October 2023) |
| Protein transmembrane | TMHMM2.0 | https://services.healthtech.dtu.dk/services/TMHMM-2.0/ (accessed on 21 October 2023) |
| Hydrophilic analysis | ProtScale | www.expasy.org/cgi-bin/protscale.pl (accessed on 21 October 2023) |
| Signal peptide | SingalP4.1 | https://services.healthtech.dtu.dk/services/SignalP-4.1 (accessed on 21 October 2023) |
| Protein motif | MEME5.5.2 | http://meme-suite.org/meme/tools/meme (accessed on 21 October 2023) |
| Conserved domain | CDD | www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml (accessed on 21 October 2023) |
| Subcellular localization | Cell-PLoc 2.0 | www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2 (accessed on 21 October 2023) |
| SOX Family | Accession No. | Size (aa) | MW (kDa) | pI | HI* | Subcellular Localization |
|---|---|---|---|---|---|---|
| gjSOX2 | XP_015262440.1 | 384 | 40.74 | 9.92 | −0.57 | Cytoplasm; nucleus |
| gjSOX3 | XP_015267683.1 | 318 | 34.01 | 9.62 | −0.61 | Cytoplasm; nucleus |
| gjSOX4 | XP_015271872.1 | 420 | 44.34 | 6.74 | −0.67 | Nucleus |
| gjSOX5 | XP_015268168.1 | 764 | 84.30 | 6.12 | −0.77 | Nucleus |
| gjSOX6 | XP_015277789.1 | 787 | 87.40 | 6.54 | −0.82 | Nucleus |
| gjSOX7 | XP_015284766.1 | 375 | 41.46 | 6.16 | −0.68 | Nucleus |
| gjSOX8 | XP_015279835.1 | 325 | 35.64 | 6.28 | −1.02 | Cytoplasm; nucleus |
| gjSOX9 | WGV33816.1 | 494 | 55.35 | 6.31 | −1.09 | Nucleus |
| gjSOX10 | XP_015261463.1 | 461 | 49.89 | 6.20 | −0.84 | Nucleus |
| gjSOX11 | XP_015262936.1 | 417 | 45.62 | 5.09 | −0.85 | Cytoplasm; nucleus |
| gjSOX12 | XP_015279104.1 | 291 | 32.88 | 8.82 | −0.87 | Cytoplasm; nucleus |
| gjSOX13 | XP_015262805.1 | 615 | 69.24 | 6.31 | −0.87 | Nucleus |
| gjSOX14 | XP_015276981.1 | 240 | 26.64 | 9.68 | −0.61 | Cytoplasm; nucleus |
| gjSOX17 | XP_015282499.1 | 177 | 18.63 | 5.94 | −0.55 | Nucleus |
| gjSOX18L | XP_015284209.1 | 398 | 44.32 | 8.66 | −0.76 | Nucleus |
| gjSOX30 | XP_015272912.1 | 778 | 85.68 | 5.75 | −0.76 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Zhao, R.; Xu, Z.; Fu, C.; Xie, L.; Li, S.; Wang, X.; Zhang, Y. gjSOX9 Cloning, Expression, and Comparison with gjSOXs Family Members in Gekko japonicus. Curr. Issues Mol. Biol. 2023, 45, 9328-9341. https://doi.org/10.3390/cimb45110584
Huang X, Zhao R, Xu Z, Fu C, Xie L, Li S, Wang X, Zhang Y. gjSOX9 Cloning, Expression, and Comparison with gjSOXs Family Members in Gekko japonicus. Current Issues in Molecular Biology. 2023; 45(11):9328-9341. https://doi.org/10.3390/cimb45110584
Chicago/Turabian StyleHuang, Xingze, Ruonan Zhao, Zhiwang Xu, Chuyan Fu, Lei Xie, Shuran Li, Xiaofeng Wang, and Yongpu Zhang. 2023. "gjSOX9 Cloning, Expression, and Comparison with gjSOXs Family Members in Gekko japonicus" Current Issues in Molecular Biology 45, no. 11: 9328-9341. https://doi.org/10.3390/cimb45110584
APA StyleHuang, X., Zhao, R., Xu, Z., Fu, C., Xie, L., Li, S., Wang, X., & Zhang, Y. (2023). gjSOX9 Cloning, Expression, and Comparison with gjSOXs Family Members in Gekko japonicus. Current Issues in Molecular Biology, 45(11), 9328-9341. https://doi.org/10.3390/cimb45110584