
Genome-Wide Association Study of Partial Resistance to P. sojae in Wild Soybeans from Heilongjiang Province, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Medium and P. sojae Strain Preparation
2.3. Pot Experiment
2.4. Inoculation Identification
2.5. Genotype Data and Quality Control
2.6. Population Structure and Linkage Disequilibrium Analysis
2.7. Genome-Wide Association Analysis
2.8. Fluorescence Quantitative PCR Detection
3. Results
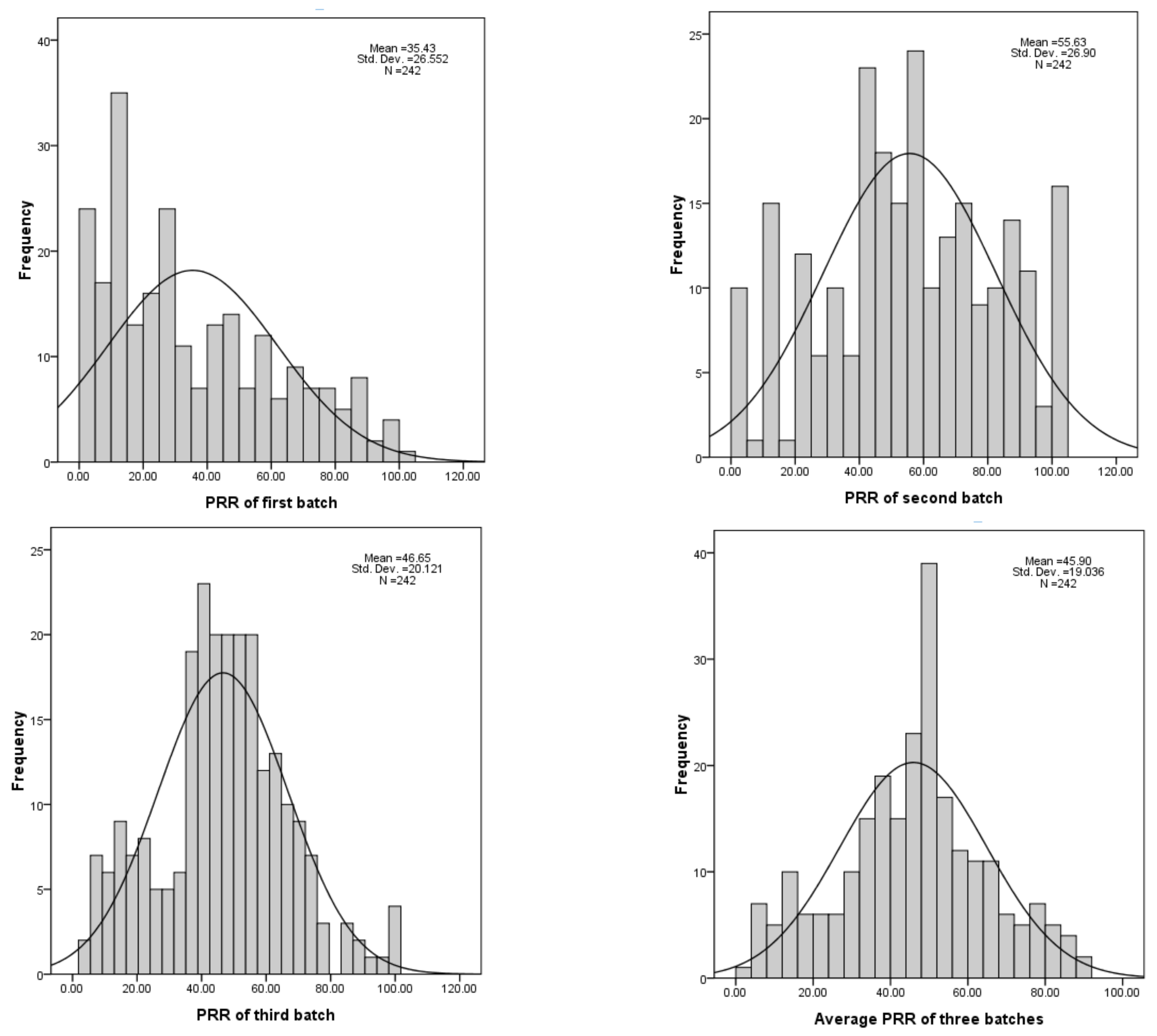
3.1. Variation in Resistant Levels among G. soja Accessions
3.2. SNP Data
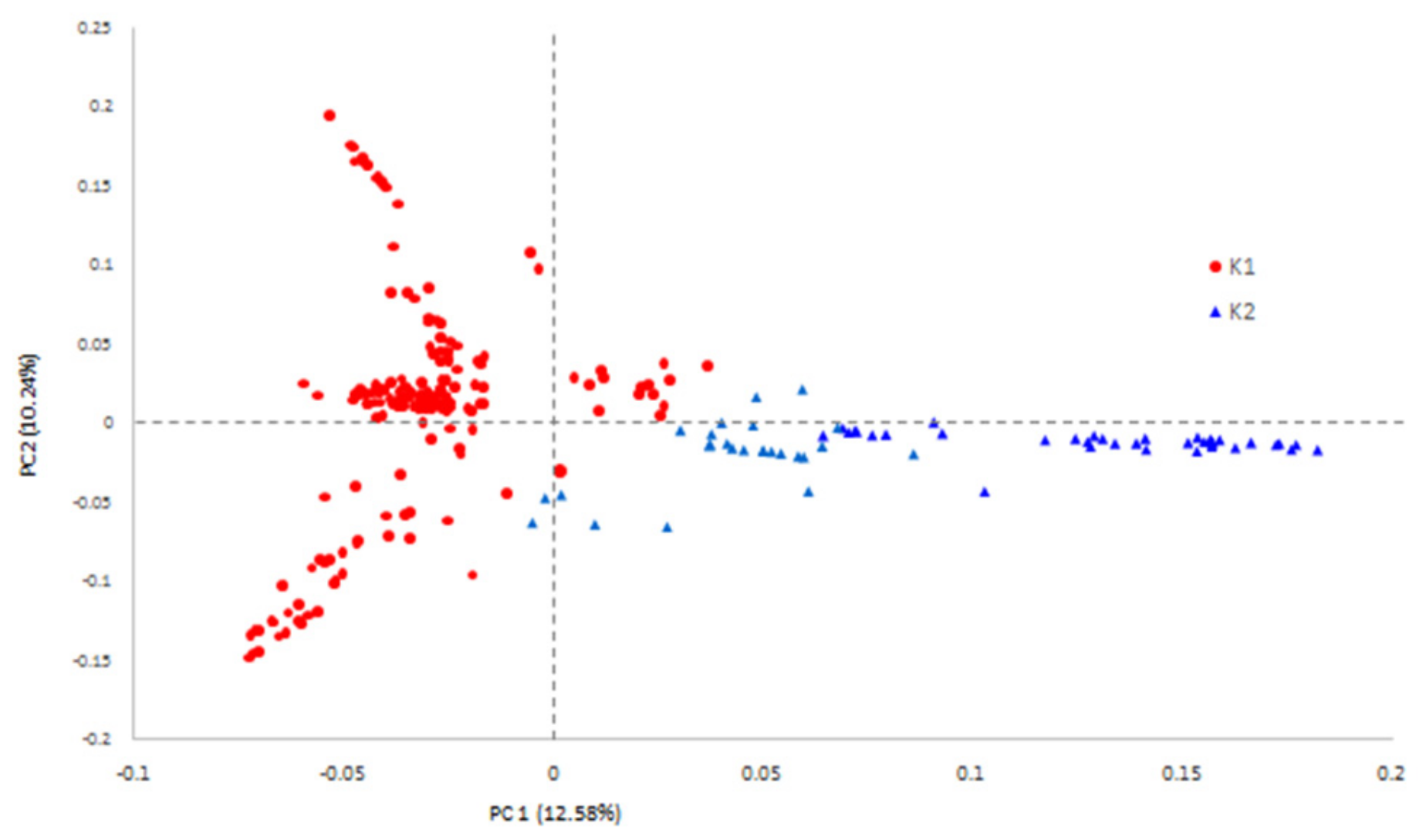
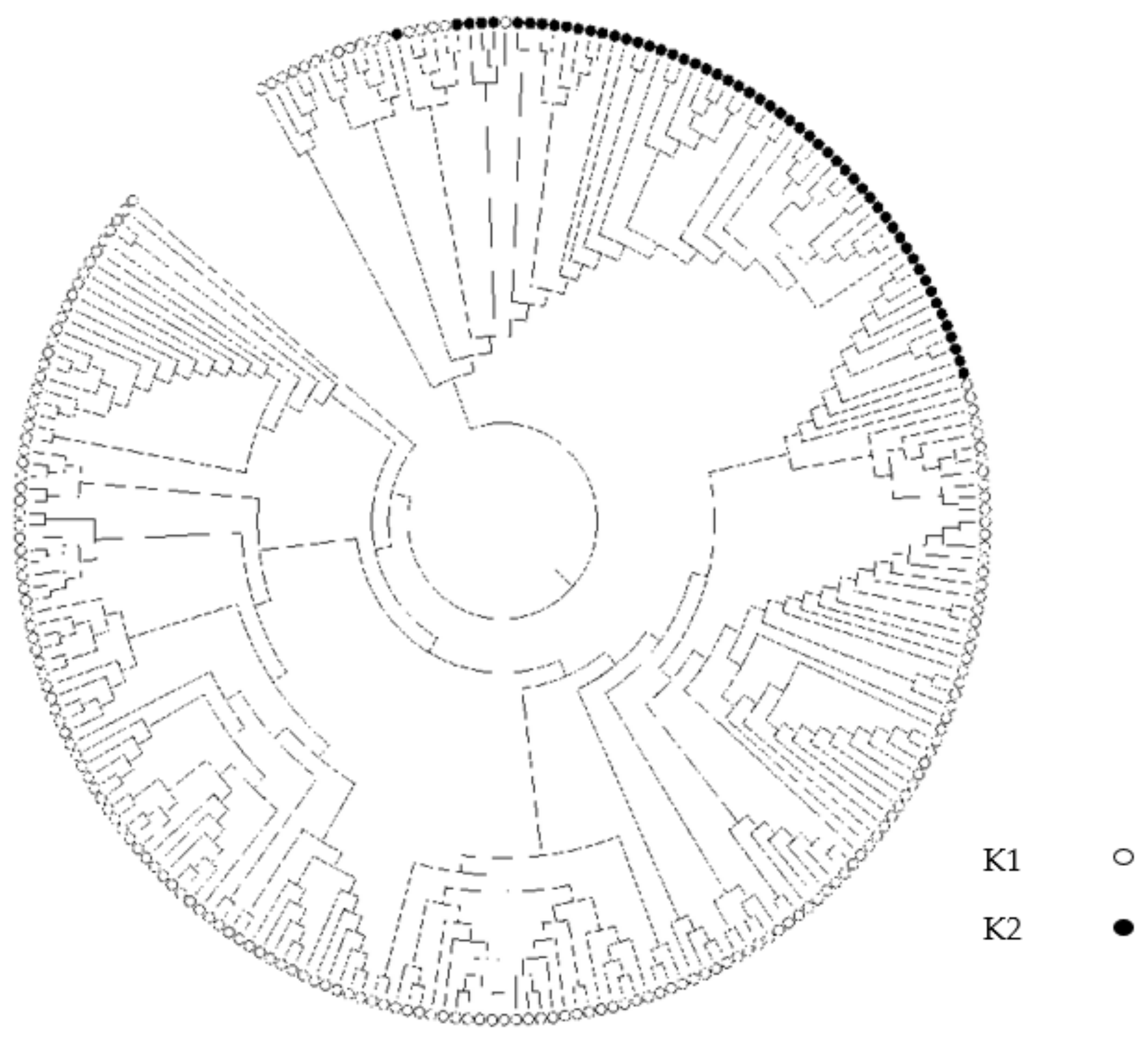
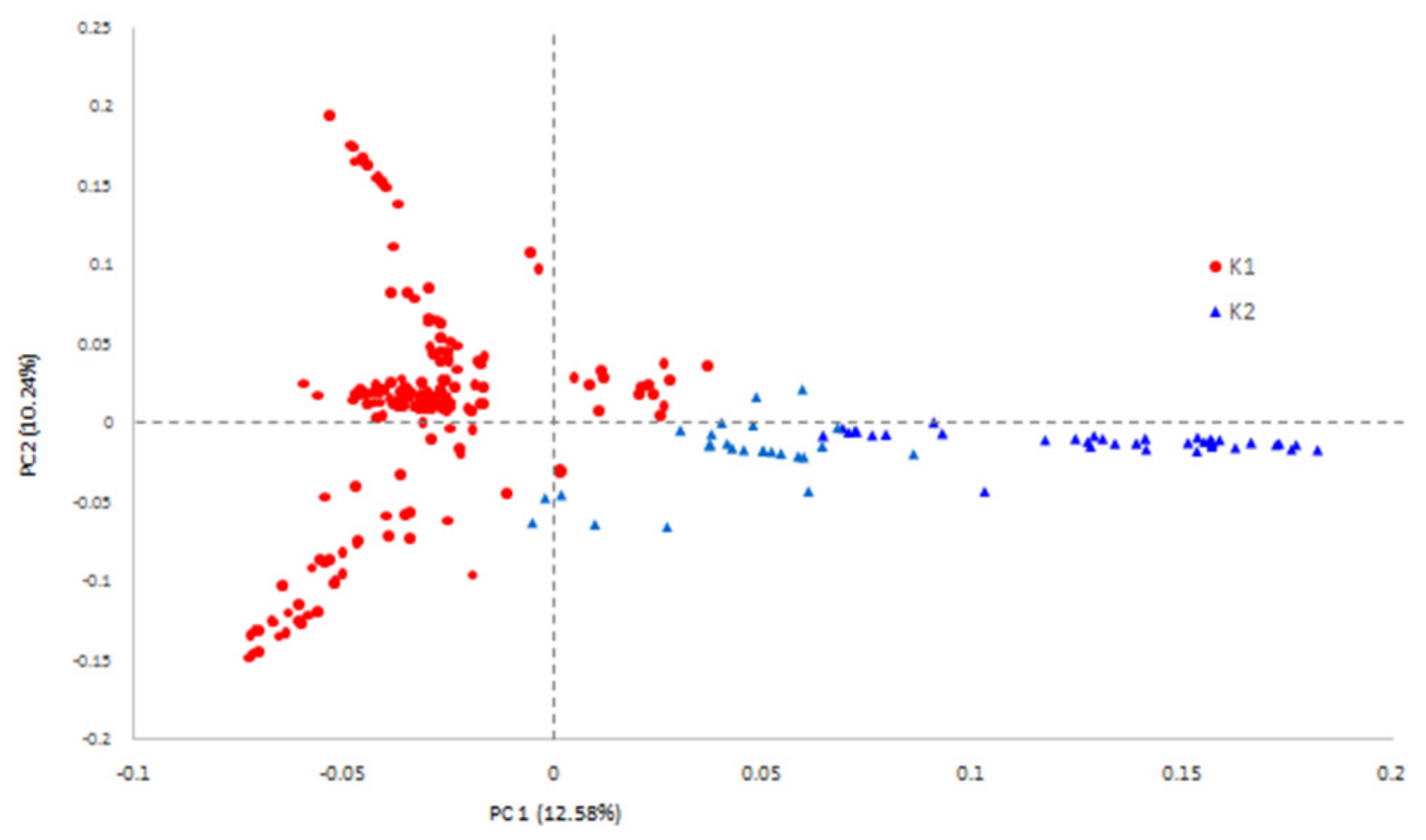
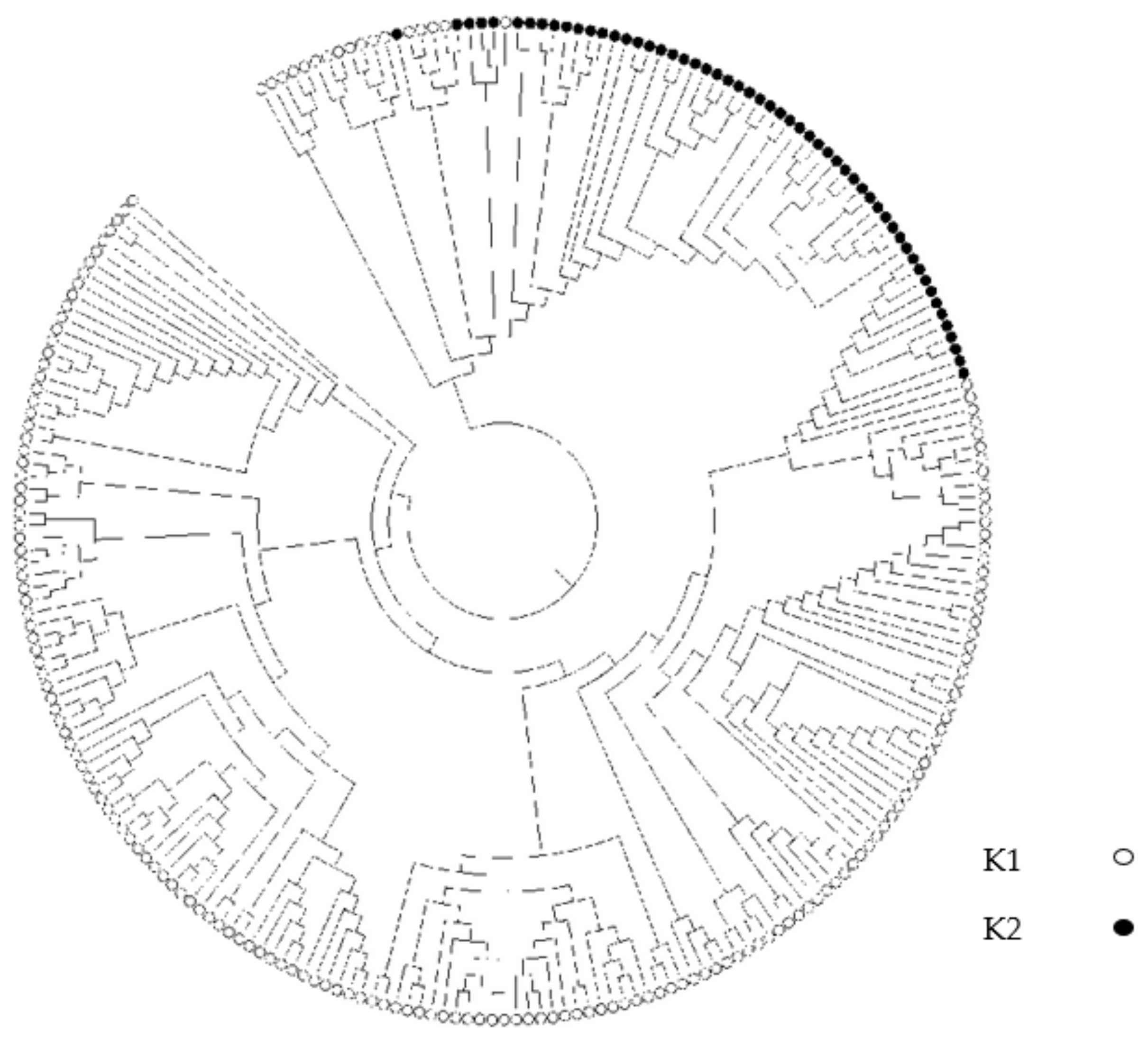
3.3. Population Structure
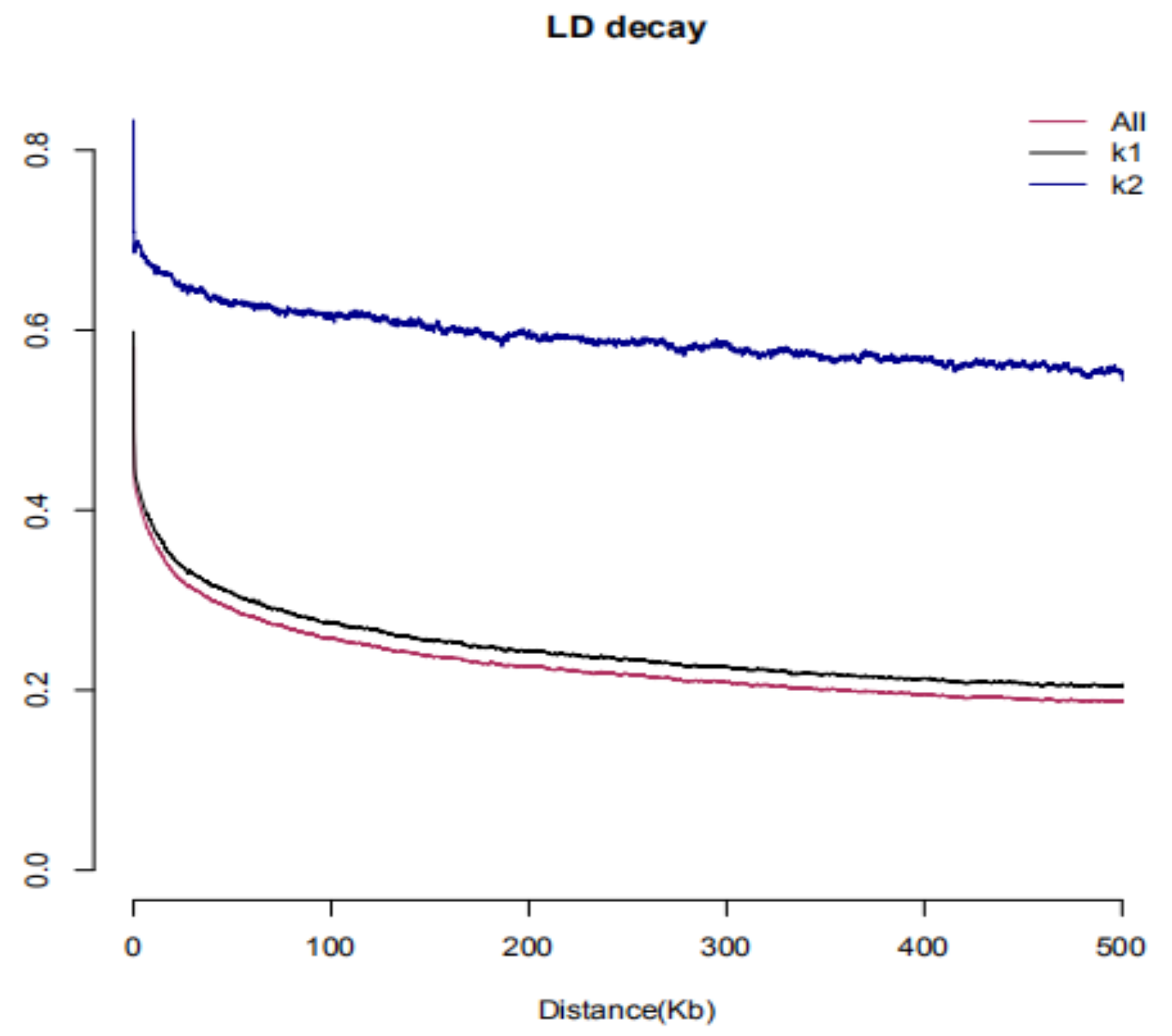
3.4. Linkage Disequilibrium
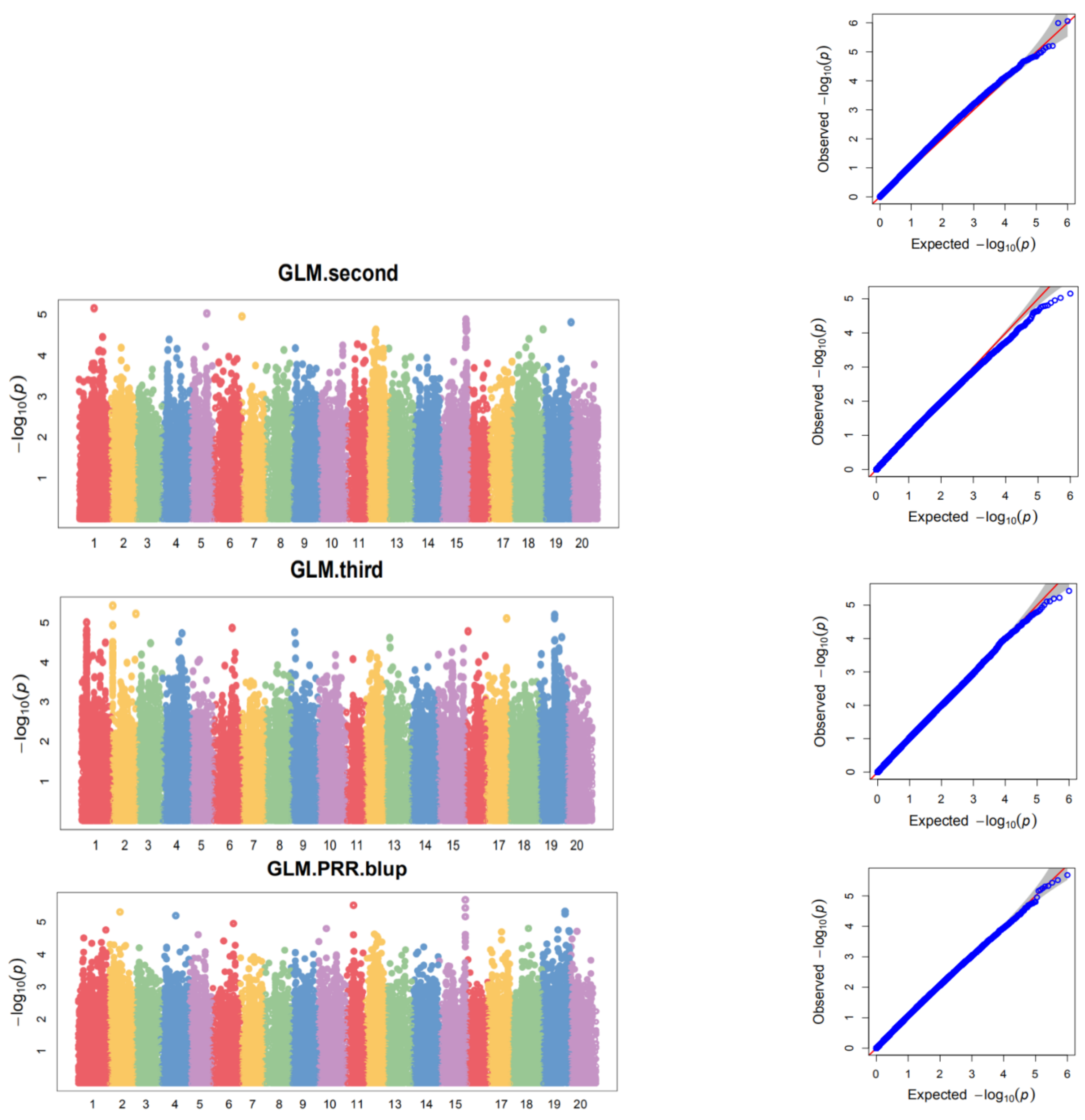
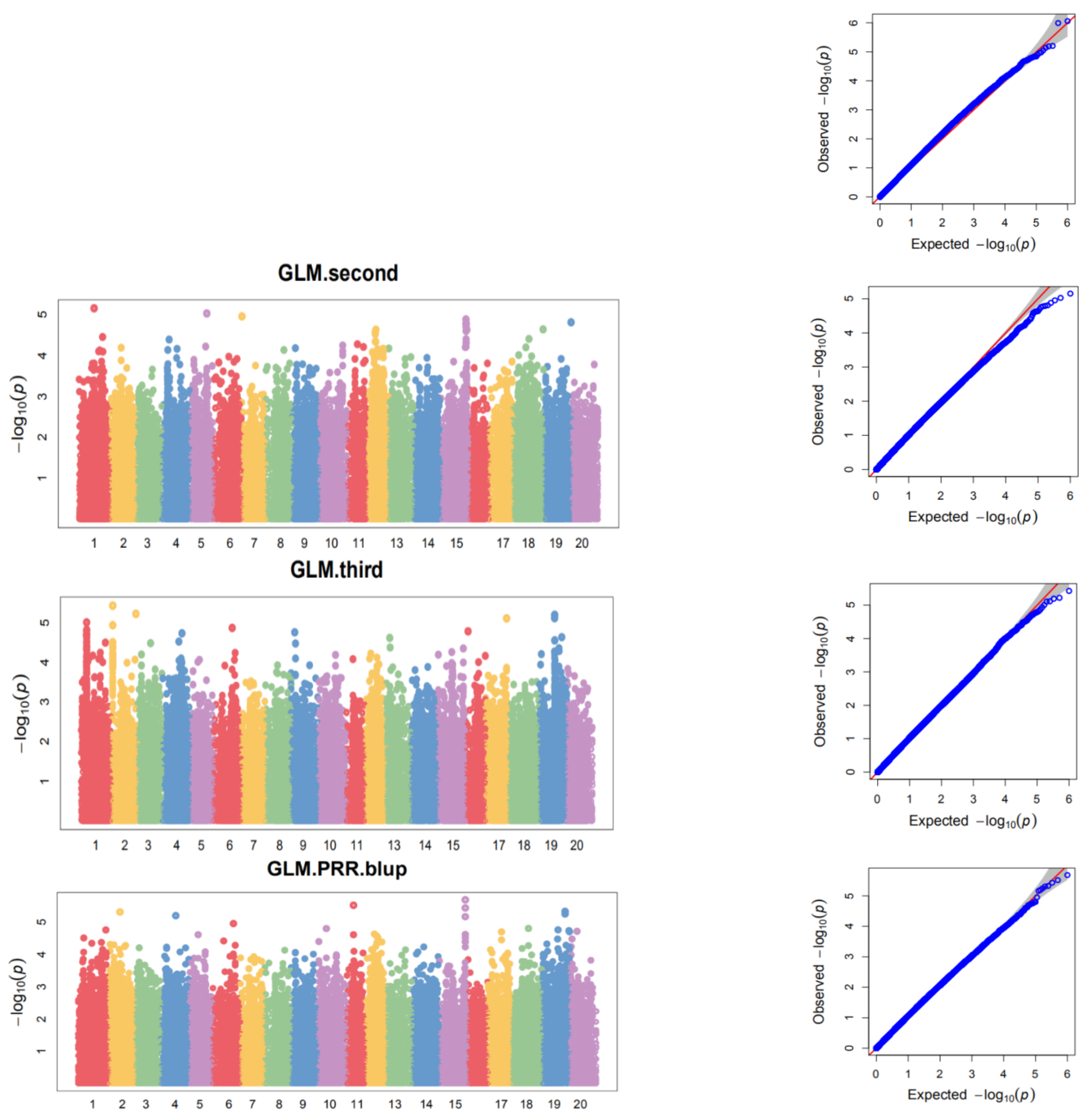
3.5. Genome-Wide Association Analysis
3.6. Prediction of Candidate Genes for PRR Resistance
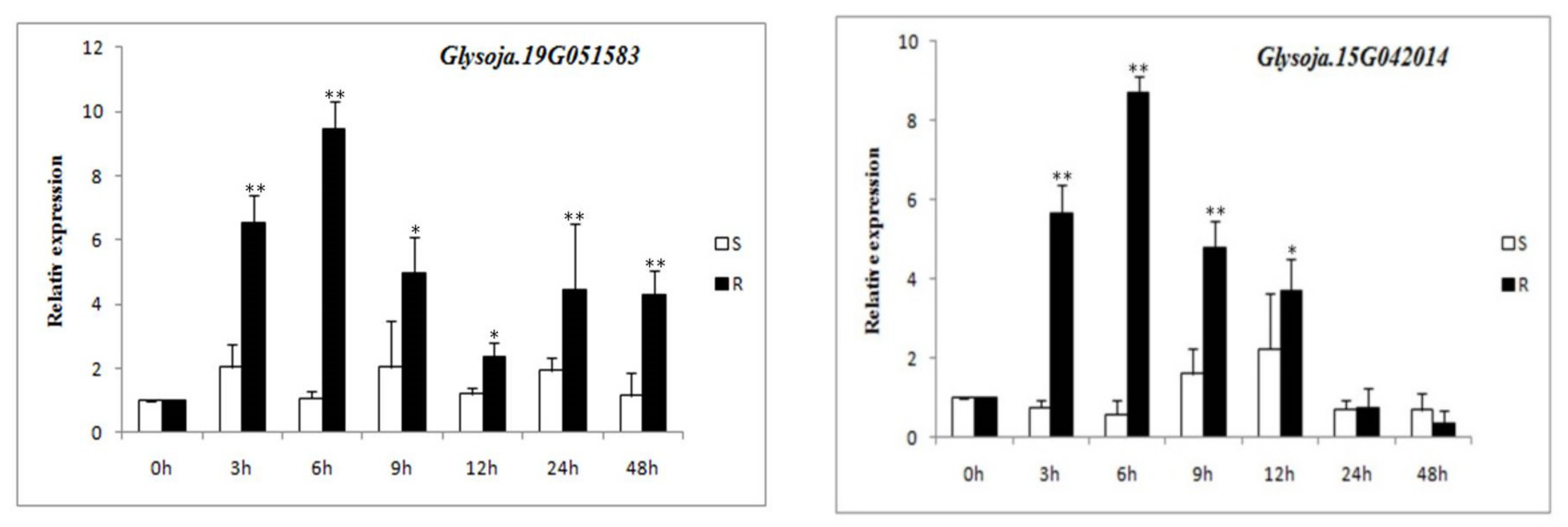
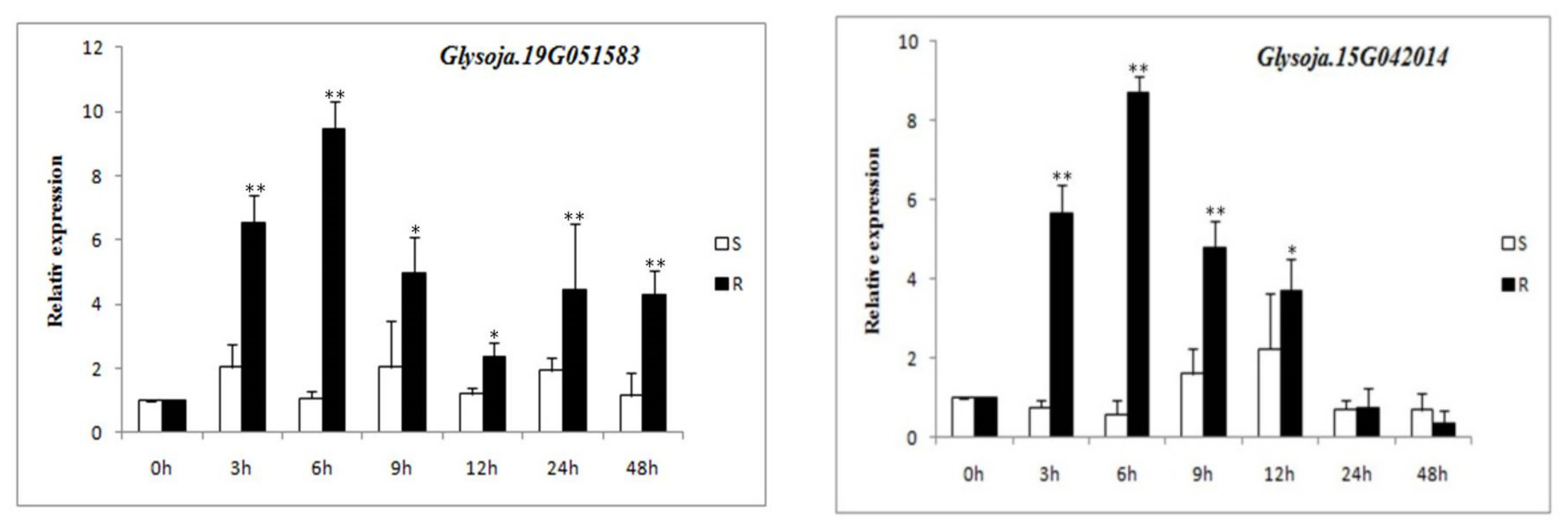
3.7. Expression of Candidate Gene’s Response to P. sojae Infection in Resistant and Susceptible Germplasms
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tyler, B.M. Phytophthora sojae: Root rot pathogen of soybean and model oomycete. Mol. Plant Pathol. 2007, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.P. Mapping of Resistance Gene to Phytophthora Sojae in Soybean and Analysis of Candidate Genes; Nanjing Agricultural University: Nanjing, China, 2018; pp. 3–4. [Google Scholar]
- Zhong, C.; Li, Y.; Sun, S.; Duan, C.; Zhu, Z. Genetic Mapping and Molecular Characterization of a Broad-spectrum Phytophthora sojae Resistance Gene in Chinese Soybean. Int. J. Mol. Sci. 2019, 20, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, T.; Kato, M.; Yoshida, S.; Matsumoto, I.; Kobayashi, T.; Kaga, A.; Hajika, M.; Yamamoto, R.; Watanabe, K.; Aino, M.; et al. Pathogenic diversity of Phytophthora sojae and breeding strategies to develop Phytophthora-resistant soybeans. Breed. Sci. 2012, 61, 511–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Li, L.; Zhao, J.; Huang, J.; Yan, Q.; Xing, H.; Guo, N. Genetic analysis and fine mapping of RpsJS, a novel resistance gene to Phytophthora sojae in soybean [Glycine max (L.) Merr.]. Theor. Appl. Genet. 2014, 127, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Bernard, R.; Smith, P.; Kaufmann, M.; Schmitthenner, A.F. Inheritance of resistance to phytophthora root and stem rot in soybean. Agron. J. 1957, 49, 391. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Abeysekara, N.S.; Cianzio, S.R.; Robertson, A.E.; Bhattacharyya, M.K. A Novel Phytophthora sojae Resistance Rps12 Gene Mapped to a Genomic Region That Contains Several Rps Genes. PLoS ONE 2017, 12, e0169950. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.; Guo, N.; Sun, J.; Li, L.; Cao, Y.; Li, S.; Huang, J.; Zhao, J.; Zhao, T.; Xing, H. Fine Mapping of a Resistance Gene RpsHN that Controls Phytophthora sojae Using Recombinant Inbred Lines and Secondary Populations. Front. Plant Sci. 2017, 8, 538. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, S.; Zhong, C.; Wang, X.; Wu, X.; Zhu, Z. Genetic mapping and development of co-segregating markers of RpsQ, which provides resistance to Phytophthora sojae in soybean. Theor. Appl. Genet. 2017, 130, 1223–1233. [Google Scholar] [CrossRef]
- Jiang, B.; Cheng, Y.; Cai, Z.; Li, M.; Jiang, Z.; Ma, R.; Yuan, Y.; Xia, Q.; Nian, H. Fine mapping of a Phytophthora-resistance locus RpsGZ in soybean using genotyping-by-sequencing. BMC Genom. 2020, 21, 280. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, W.; Ping, J.; Fitzgerald, J.C.; Cai, G.; Clark, C.B.; Aggarwal, R.; Ma, J. Identification and molecular mapping of Rps14, a gene conferring broad-spectrum resistance to Phytophthora sojae in soybean. Theor. Appl. Genet. 2021, 134, 3863–3872. [Google Scholar] [CrossRef]
- Van, K.; Rolling, W.; Biyashev, R.M.; Matthiesen, R.L.; Abeysekara, N.S.; Robertson, A.E.; Veney, D.J.; Dorrance, A.E.; McHale, L.K.; Maroof, M.A.S. Mining germplasm panels and phenotypic datasets to identify loci for resistance to Phytophthora sojae in soybean. Plant Genome 2020, 14, e20063. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, L.; Fengler, K.; Bolar, J.; Llaca, V.; Wang, X.; Clark, C.B.; Fleury, T.J.; Myrvold, J.; Oneal, D.; et al. A giant NLR gene confers broad-spectrum resistance to Phytophthora sojae in soybean. Nat. Commun. 2021, 12, 6263. [Google Scholar] [CrossRef] [PubMed]
- Ping, J.; Fitzgerald, J.C.; Zhang, C.; Lin, F.; Bai, Y.; Wang, D.; Aggarwal, R.; Rehman, M.; Crasta, O.; Ma, J. Identification and molecular mapping of Rps11, a novel gene conferring resistance to Phytophthora sojae in soybean. Theor. Appl. Genet. 2016, 129, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Ma, Q.; Ren, H.; Xia, Q.; Song, E.; Tan, Z.; Li, S.; Zhang, G.; Nian, H. Fine mapping of a Phytophthora-resistance gene RpsWY in soybean (Glycine max L.) by high-throughput genome-wide sequencing. Theor. Appl. Genet. 2017, 130, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Bao, D.; Wang, W.; Zhang, C.; Jing, Y.; Jiang, H.; Qiu, L.; Li, W.; Han, Y. Loci and candidate gene identification for soybean resistance to Phytophthora root rot race 1 in combination with association and linkage mapping. Mol. Breed. 2020, 40, 100. [Google Scholar] [CrossRef]
- Zhong, C.; Sun, S.; Yao, L.; Ding, J.; Duan, C.; Zhu, Z. Fine Mapping and Identification of a Novel Phytophthora Root Rot Resistance Locus RpsZS18 on Chromosome 2 in Soybean. Front. Plant Sci. 2018, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Sun, S.; Zhang, X.; Duan, C.; Zhu, Z. Fine Mapping, Candidate Gene Identification and Co-segregating Marker Development for the Phytophthora Root Rot Resistance Gene RpsYD25. Front. Genet. 2020, 11, 799. [Google Scholar] [CrossRef] [PubMed]
- Tooley, P.W. Identification and quantitative characterization of rate-reducing resistance to phytophthora megasperma f. sp. glycinea in soybean seedlings. Phytopathology 1982, 72, 727–733. [Google Scholar] [CrossRef]
- de Ronne, M.; Santhanam, P.; Cinget, B.; Labbé, C.; Lebreton, A.; Ye, H.; Vuong, T.D.; Hu, H.; Valliyodan, B.; Edwards, D.; et al. Mapping of partial resistance to Phytophthora sojae in soybean PIs using whole-genome sequencing reveals a major QTL. Plant Genome 2021, 15, e20184. [Google Scholar] [CrossRef]
- Li, L.H.; Guo, N.; Niu, J.P.; Wang, Z.; Cui, X.; Sun, J.; Zhao, T.; Xing, H. Loci and Candidate Gene Identification for Resistance to Phytophthora sojae via Association Analysis in Soybean [Glycine Max (L.) Merr.]. Mol. Genet. Genom. 2016, 291, 1095–1103. [Google Scholar] [CrossRef]
- Karhoff, S.; Lee, S.; Mian, M.A.R.; Ralston, T.I.; Niblack, T.L.; Dorrance, A.E.; McHale, L.K. Phenotypic Characterization of a Major Quantitative Disease Resistance Locus for Partial Resistance to Phytophthora sojae. Crop Sci. 2019, 59, 968–980. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.; Guo, N.; Zhang, Z.; Wang, Z.; Huang, J.; Zhao, J.; Chang, F.; Wang, H.; Zhao, T.; Xing, H. Genome-wide SNP-based association mapping of resistance to Phytophthora sojae in soybean (Glycine max (L.) Merr.). Euphytica 2018, 214, 187. [Google Scholar] [CrossRef]
- Qin, J.; Song, Q.; Shi, A.; Li, S.; Zhang, M.; Zhang, B. Genome-wide association mapping of resistance to Phytophthora sojae in a soybean [Glycine max (L.) Merr.] germplasm panel from maturity groups IV and V. PLoS ONE 2017, 12, e0184613. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Guo, N.; Lei, J.; Li, L.; Hu, G.; Xing, H. Association mapping for partial resistance to Phytophthora sojae in soybean (Glycine max (L.) Merr.). J. Genet. 2014, 93, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zhao, M.; Ping, J.; Johnson, A.; Zhang, B.; Abney, T.S.; Hughes, T.J.; Ma, J. Molecular mapping of two genes conferring resistance to Phytophthora sojae in a soybean landrace PI 567139B. Theor. Appl. Genet. 2013, 126, 2177–2185. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Sharma, D.; Banyal, S.K. Potential, Challenges and Strategies Involved in Gene Introgression from Wild Relatives of Vegetable Crops: A Review. Agric. Rev. 2021, 4, 390–397. [Google Scholar] [CrossRef]
- Francis, D.; Bernal, E.; Orchard, C.; Subode, S. New approaches in the discovery and introgression of disease resistance genes from wild tomato. Acta Hortic. 2021, 1316, 23–34. [Google Scholar] [CrossRef]
- Jin, Y.; He, T.; Lu, B. Fine scale genetic structure in a wild soybean (Glycine soja) population and the implications for conservation. New Phytol. 2003, 159, 513–519. [Google Scholar] [CrossRef]
- Li, W.; Peng, M.; Wang, Z.; Bi, Y.; Liu, M.; Wang, L.; Di, S.; Liu, J.; Fan, C.; Yang, G.; et al. The Evaluation of Agronomic Traits of Wild Soybean Accessions (Glycine soja Sieb. and Zucc.) in Heilongjiang Province, China. Agronomy 2021, 11, 586. [Google Scholar] [CrossRef]
- Huang, J.; Guo, N.; Li, Y.; Sun, J.; Hu, G.; Zhang, H.; Li, Y.; Zhang, X.; Zhao, J.; Xing, H.; et al. Phenotypic evaluation and genetic dissection of resistance to Phytophthora sojae in the Chinese soybean mini core collection. BMC Genet. 2016, 17, 85. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.-J.; Sugano, S.; Kaga, A.; Lee, S.S.; Sugimoto, T.; Takahashi, M.; Ishimoto, M. Evaluation of Resistance to Phytophthora sojae in Soybean Mini Core Collections Using an Improved Assay System. Phytopathology 2017, 107, 216–223. [Google Scholar] [CrossRef]
- Yang, J.; Ye, W.; Wang, X.; Ren, L.; Yao, Y.; Wang, X.; Wang, Y.; Dong, S.; Zheng, X.; Wang, Y. An Improved Method for the Identification of Soybean Resistance to Phytophthora sojae Applied to Germplasm Resources from the Huanghuaihai and Dongbei Regions of China. Plant Dis. 2020, 104, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D.; Glazebrook, J.; Glazebrook, W. Arabidopsis a Laboratory Manual; CSHL Press: Cold Spring Harbor, NY, USA, 2002. [Google Scholar]
- Yang, X.B. Races of Phytophthora sojae in Iowa soybean fields. Plant Dis. 1996, 80, 14180–14201. [Google Scholar] [CrossRef]
- Lam, H.-M.; Xu, X.; Liu, X.; Chen, W.; Yang, G.; Wong, F.-L.; Li, M.-W.; He, W.; Qin, N.; Wang, B.; et al. Resequencing of 31 wild and cultivated soybean genomes identifies patterns of genetic diversity and selection. Nat. Genet. 2010, 42, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; Stephens, M.; Pritchard, J.K. fastSTRUCTURE: Variational Inference of Population Structure in Large SNP Data Sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanRaden, P. Efficient Methods to Compute Genomic Predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Wang, X.M.; Ye, W.W.; Zheng, X.; Wang, Y. Identification of soybean resistance to Phytophthora sojae in the germplasm resources from Huanghuaihai Region of China. Soybean Sci. 2020, 1, 12–22. [Google Scholar] [CrossRef]
- Jin, L.M.; Xu, P.F.; Wu, J.J.; Li, W.; Qiu, L.; Chang, R.; Xu, P.; Zhang, S. Identification the resistance of wild soybean germplasm to Phytophthora sojae. Soybean Sci. 2007, 26, 300–304. [Google Scholar]
- Schneider, R.; Rolling, W.; Song, Q.; Cregan, P.; Dorrance, A.E.; McHale, L.K. Genome-wide association mapping of partial resistance to Phytophthora sojae in soybean plant introductions from the Republic of Korea. BMC Genom. 2016, 17, 607. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; He, J.; Xiao, Y.; Zhang, Y.; Liu, Y.; Wan, S.; Liu, L.; Dong, Y.; Liu, H.; Yu, Y. CsGSTU8, a Glutathione S-Transferase from Camellia sinensis, Is Regulated by CsWRKY48 and Plays a Positive Role in Drought Tolerance. Front. Plant Sci. 2021, 12, 795919. [Google Scholar] [CrossRef]
- Zhao, D.; Luan, Y.; Shi, W.; Zhang, X.; Meng, J.; Tao, J. A Paeonia ostii caffeoyl-CoA O-methyltransferase confers drought stress tolerance by promoting lignin synthesis and ROS scavenging. Plant Sci. 2021, 303, 110765. [Google Scholar] [CrossRef] [PubMed]
- Zambounis, A.; Psomopoulos, F.E.; Ganopoulos, I.; Avramidou, E.; Aravanopoulos, F.; Tsaftaris, A.; Madesis, P. In silico analysis of the LRR receptor-like serine threonine kinases subfamily in Morus notabilis. Plant Omics 2016, 9, 319–326. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, X.; Gao, Y.; Jiao, J.; Sun, Y.; Zhu, D.; Yang, J.; Wu, F.; Su, H. Genome-wide analysis of lectin receptor-like kinases (LecRLKs) in sweet cherry (Prunus avium) and reveals PaLectinL16 enhances sweet cherry resistance with salt stress. Environ. Exp. Bot. 2022, 194, 104751. [Google Scholar] [CrossRef]
- Cui, Y.W.; Lu, X.T.; Gou, X.P. Receptor-like protein kinases in plant reproduction: Current understanding and future perspec-tives. Plant Commun. 2021, 3, 100273. [Google Scholar] [CrossRef]
- Mondal, R.; Biswas, S.; Srivastava, A.; Basu, S.; Trivedi, M.; Singh, S.K.; Mishra, Y. In silico analysis and expression profiling of S-domain receptor-like kinases (SD-RLKs) under different abiotic stresses in Arabidopsis thaliana. BMC Genom. 2021, 22, 817. [Google Scholar] [CrossRef] [PubMed]
- Jing, M.; Ma, H.; Li, H.; Guo, B.; Zhang, X.; Ye, W.; Wang, H.; Wang, Q.; Wang, Y. Differential regulation of defense-related proteins in soybean during compatible and incompatible interactions between Phytophthora sojae and soybean by comparative proteomic analysis. Plant Cell Rep. 2015, 34, 1263–1280. [Google Scholar] [CrossRef]
- Zeng, M.; Wan, B.; Wang, L.; Chen, Z.; Lin, Y.; Ye, W.; Wang, Y.; Wang, Y. Identification and characterization of L-type lectin receptor-like kinases involved in Glycine max–Phytophthora sojae interaction. Planta 2021, 254, 128. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hanikenne, M.; Clemens, S. GmSnRK1.1, a Sucrose Non-fermenting-1(SNF1)-Related Protein Kinase, Promotes Soybean Resistance to Phytophthora sojae. Front. Plant Sci. 2019, 10, 996. [Google Scholar] [CrossRef] [Green Version]
- Jang, I.-H.; Kang, I.J.; Kim, J.-M.; Kang, S.-T.; Jang, Y.E.; Lee, S. Genetic Mapping of a Resistance Locus to Phytophthora sojae in the Korean Soybean Cultivar Daewon. Plant Pathol. J. 2020, 36, 591–599. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction | Standard of Identification | Susceptible Rate (%) |
|---|---|---|
| R | Yellowing, browning, or chlorosis of leaves | <30 |
| I | 30–70 | |
| S | >70 |
| Reaction | Numbers | Percent |
|---|---|---|
| S | 47 | 19.50 |
| I | 167 | 69.29 |
| R | 27 | 11.20 |
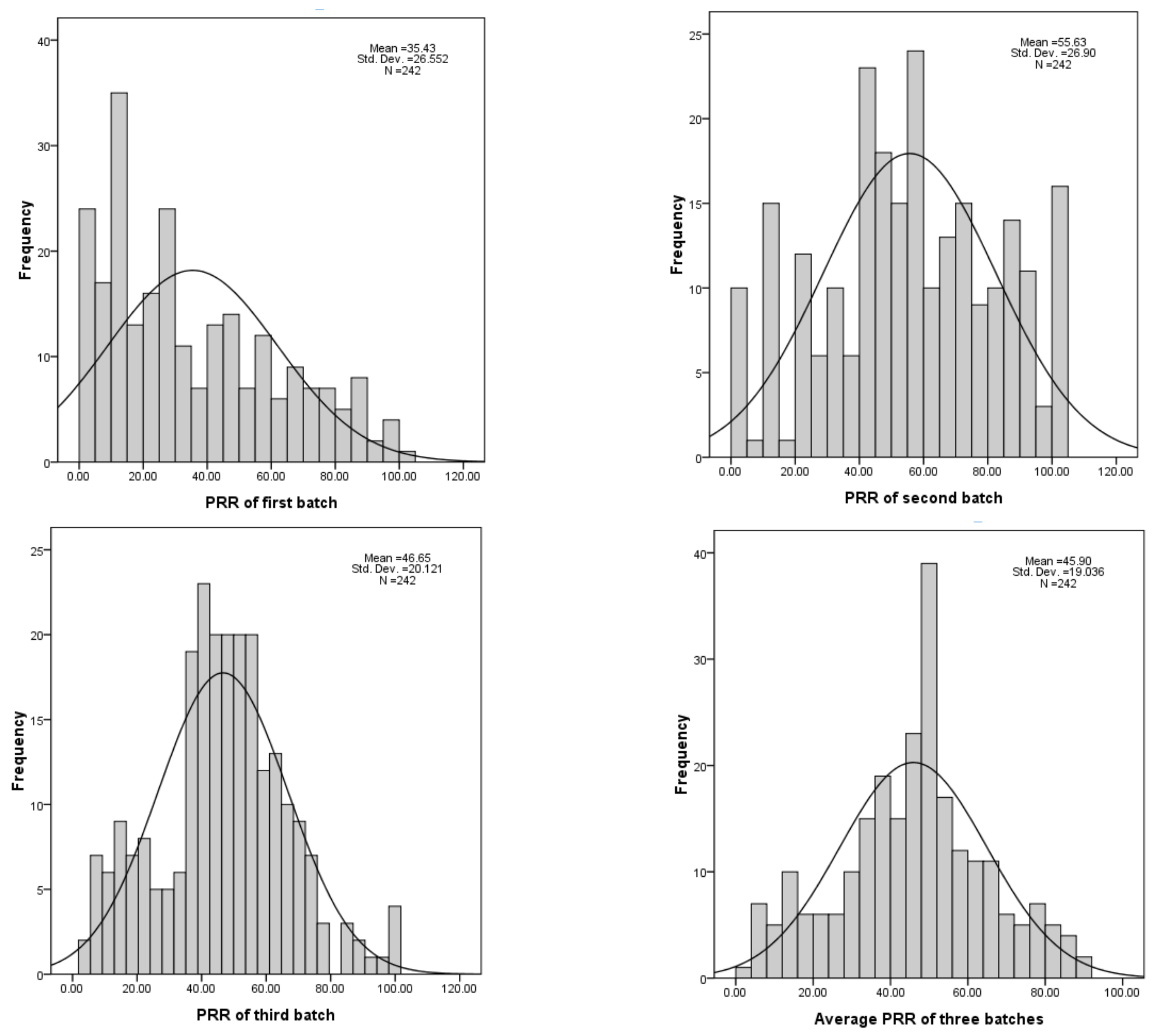
| Batch | Skew | Kurtosis | Heritability |
|---|---|---|---|
| First | 0.62 | −0.67 | 65.82 |
| Second | −0.17 | −0.73 | |
| Third | 0.08 | 0.07 |
| Batch | SNP | Chr | Position | SNP | Chr | Position |
|---|---|---|---|---|---|---|
| First | rs64989 | 1 | 51,714,188 | rs494319 | 11 | 15,826,009 |
| rs70530 | 2 | 9,109,207 | rs494372 | 11 | 15,827,702 | |
| rs70550 | 2 | 9,110,030 | rs497213 | 11 | 17,412,064 | |
| rs70646 | 2 | 9,120,078 | rs498507 | 11 | 17,862,252 | |
| rs264115 | 6 | 21,032,877 | rs570096 | 13 | 1,285,457 | |
| rs434454 | 10 | 8,904,675 | rs779074 | 17 | 24,607,598 | |
| rs444818 | 10 | 14,862,256 | rs885444 | 19 | 8,001,494 | |
| Second | rs32673 | 1 | 27,448,011 | rs717476 | 15 | 46,519,729 |
| rs248180 | 5 | 30,006,501 | rs717478 | 15 | 46,519,754 | |
| rs299633 | 7 | 625,039 | rs717479 | 15 | 46,519,777 | |
| rs530881 | 12 | 15,479,974 | rs717480 | 15 | 46,519,780 | |
| rs530911 | 12 | 15,481,110 | rs718743 | 15 | 47,944,309 | |
| rs532502 | 12 | 16,557,015 | rs875366 | 18 | 56,051,504 | |
| rs717197 | 15 | 46,275,173 | rs940996 | 19 | 49,218,250 | |
| rs717475 | 15 | 46,519,716 | ||||
| Third | rs9312 | 1 | 10,161,448 | rs193415 | 4 | 30,457,058 |
| rs9748 | 1 | 10,439,969 | rs200863 | 4 | 36,132,462 | |
| rs10026 | 1 | 10,532,649 | rs281244 | 6 | 34,683,089 | |
| rs10216 | 1 | 10,586,571 | rs371411 | 9 | 6,767,103 | |
| rs10270 | 1 | 10,593,132 | rs576319 | 13 | 6,313,762 | |
| rs10380 | 1 | 10,641,555 | rs721959 | 16 | 5,055,148 | |
| rs10436 | 1 | 10,669,494 | rs801645 | 17 | 38,595,915 | |
| rs10641 | 1 | 10,725,370 | rs921647 | 19 | 27,986,596 | |
| rs67599 | 2 | 2,186,517 | rs921800 | 19 | 28,122,057 | |
| rs67717 | 2 | 2,190,064 | rs921801 | 19 | 28,122,124 | |
| rs67718 | 2 | 2,190,067 | rs922217 | 19 | 28,376,899 | |
| rs67720 | 2 | 2,190,157 | rs938638 | 19 | 41,267,399 | |
| rs116637 | 2 | 45,054,777 | ||||
| BLUP | rs10641 | 1 | 10,725,370 | rs718675 | 15 | 47,928,197 |
| rs64791 | 1 | 51,140,109 | rs718693 | 15 | 47,938,751 | |
| rs85323 | 2 | 20,208,766 | rs718743 | 15 | 47,944,309 | |
| rs190201 | 4 | 27,955,786 | rs718756 | 15 | 47,944,902 | |
| rs230678 | 5 | 16,483,623 | rs779074 | 17 | 24,607,598 | |
| rs286790 | 6 | 38,875,969 | rs844904 | 18 | 32,247,141 | |
| rs444818 | 10 | 14,862,256 | rs922217 | 19 | 28,376,899 | |
| rs490370 | 11 | 12,974,021 | rs938637 | 19 | 41,267,263 | |
| rs490521 | 11 | 13,035,152 | rs938638 | 19 | 41,267,399 | |
| rs532502 | 12 | 16,557,015 | rs940814 | 19 | 48,382,478 | |
| rs539755 | 12 | 21,078,905 | rs940996 | 19 | 49,218,250 | |
| rs545454 | 12 | 22,657,211 | rs958957 | 20 | 12,404,540 | |
| rs718653 | 15 | 47,921,059 |
| Chr | SNP | Batch | Position | p Value | −log10(p) | Maf | Phenotypic Variation (%) |
|---|---|---|---|---|---|---|---|
| 1 | rs10641 | Third | 10,725,370 | 1.98 × 10−5 | 4.70 | 0.13 | 8.21 |
| BLUP | 3.13 × 10−5 | 4.50 | 7.43 | ||||
| 10 | rs444818 | First | 14,862,256 | 1.46 × 10−5 | 4.84 | 0.12 | 7.75 |
| BLUP | 1.60 × 10−5 | 4.80 | 8 | ||||
| 12 | rs532502 | Second | 16,557,015 | 2.35 × 10−5 | 4.63 | 0.33 | 7.63 |
| BLUP | 2.37 × 10−5 | 4.63 | 7.67 | ||||
| 15 | rs718743 | Second | 47,944,309 | 2.36 × 10−5 | 4.63 | 0.30 | 7.63 |
| BLUP | 2.07 × 10−6 | 5.68 | 9.77 | ||||
| 17 | rs779074 | First | 24,607,598 | 1.27 × 10−5 | 4.90 | 0.17 | 7.59 |
| BLUP | 2.03 × 10−5 | 4.69 | 7.8 | ||||
| 19 | rs922217 | Third | 28,376,899 | 2.79 × 10−5 | 4.55 | 0.19 | 7.91 |
| BLUP | 1.75 × 10−5 | 4.76 | 7.92 | ||||
| rs938638 | Third | 41,267,399 | 2.33 × 10−5 | 4.63 | 0.36 | 8.07 | |
| BLUP | 4.71 × 10−6 | 5.33 | 9.05 | ||||
| rs940996 | Second | 49,218,250 | 1.55 × 10−5 | 4.81 | 0.42 | 7.98 | |
| BLUP | 2.00 × 10−5 | 4.70 | 7.81 | ||||
| 20 | rs958957 | First | 12,404,540 | 5.29 × 10−6 | 5.28 | 0.41 | 10.05 |
| BLUP | 1.95 × 10−5 | 4.71 | 7.83 |
| SNP | Chr | Position | Allele | Mean of Susceptible Rate | Allele | Mean of Susceptible Rate | t-Test | Significant Material |
|---|---|---|---|---|---|---|---|---|
| rs10641 | Chr01 | 10,725,370 | C | 48.45 | T | 37.04 | 0.0038 | HAAS_077 |
| rs532502 | Chr12 | 16,557,015 | C | 59.4 | T | 48.78 | 0.0034 | HAAS_264 |
| rs718743 | Chr15 | 47,944,309 | A | 63.98 | C | 51.14 | 0.0010 | HAAS_264 |
| rs922217 | Chr19 | 28,376,899 | A | 54.22 | G | 44.49 | 0.0089 | HAAS_077 |
| rs938638 | 41,267,399 | T | 52.31 | G | 43.92 | 0.0018 | HAAS_077 | |
| rs940996 | 49,218,250 | C | 47.83 | G | 60.20 | 0.0002 | HAAS_077 | |
| rs958957 | Chr20 | 12,404,540 | C | 48.44 | T | 37.96 | 0.0003 | HAAS_077 |
| SNP | GENE ID | Annotations |
|---|---|---|
| Chr15 rs718743 | Glysoja.15G042021 | Putative glutathione S-transferase parC |
| Glysoja.15G042020 | Putative glutathione S-transferase | |
| Glysoja.15G042019 | Putative glutathione S-transferase | |
| Glysoja.15G042017 | Nicotianamine synthase | |
| Glysoja.15G042016 | Ubiquitin carboxyl-terminal hydrolase 25 | |
| Glysoja.15G042015 | Putative sugar phosphate/phosphate translocator | |
| Glysoja.15G042014 | Putative caffeoyl-CoA O-methyltransferase 1 | |
| Glysoja.15G042012 | (S)-2-hydroxy-acid oxidase GLO1(s) | |
| Chr19 rs922217 | Glysoja.19G050845 | Elongation factor 1-alpha |
| Chr19 rs938638 | Glysoja.19G051587 | Protein resistance to Phytophthora 1, chloroplastic-like |
| Glysoja.19G051585 | Sugar transporter ERD6-like 7 | |
| Glysoja.19G051583 | Putative LRR receptor-like serine/threonine protein kinase RKF3 | |
| Glysoja.19G051582 | Putative LRR receptor-like serine/threonine protein kinase RKF3 | |
| Glysoja.19G051581 | Receptor-like cytosolic serine/threonine protein kinase RBK2 | |
| Glysoja.19G051580 | Autophagy-related protein 18 g | |
| Glysoja.19G051579 | Histidine-containing phosphotransfer protein AHP1 | |
| Glysoja.19G051577 | Pentatricopeptide repeat-containing protein | |
| Glysoja.19G051576 | Gibberellin receptor GID1B | |
| Glysoja.19G051575 | Hypothetical protein | |
| Chr19 rs940996 | Glysoja.19G052510 | Receptor-like protein kinase ANXUR2 |
| Glysoja.19G052507 | Pathogenesis-related protein PR-4A | |
| Glysoja.19G052505 | Pro-hevein | |
| Glysoja.19G052504 | Auxin-responsive protein IAA16-like | |
| Glysoja.19G052503 | Mediator of RNA polymerase II transcription subunit 14RNA | |
| Glysoja.19G052502 | Sec-independent protein translocase protein TATA, chloroplastic | |
| Glysoja.19G052501 | Light-inducible protein CPRF2 | |
| Glysoja.19G052500 | GDP-mannose 3,5-epimerase 1 | |
| Glysoja.19G052499 | GDP-mannose 3,5-epimerase 1 | |
| Glysoja.19G052497 | Ammonium transporter 3 member 1 | |
| Glysoja.19G052496 | Calmodulin-like protein 8 |
| GENE ID | GENE Name | Position | SNP | SNP Location |
|---|---|---|---|---|
| Glysoja.15G042021 | PARC | 48,129,405–48,131,670 | rs718743 | 48,076,672 |
| Glysoja.15G042020 | GST | 48,126,966–48,129,356 | ||
| Glysoja.15G042019 | GST | 48,099,201–48,101,217 | ||
| Glysoja.15G042014 | Omt5 | 48,039,247–48,039,770 | ||
| Glysoja.19G051587 | PRR1 | 41,224,586–41,226,602 | rs938638 | 41,190,696 |
| Glysoja.19G051583 | RKF3 | 41,197,436–41,199,245 | ||
| Glysoja.19G051582 | RKF3 | 41,190,378–41,192,993 | ||
| Glysoja.19G051581 | RBK2 | 41,184,811–41,188,366 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Liu, M.; Lai, Y.-C.; Liu, J.-X.; Fan, C.; Yang, G.; Wang, L.; Liang, W.-W.; Di, S.-F.; Yu, D.-Y.; et al. Genome-Wide Association Study of Partial Resistance to P. sojae in Wild Soybeans from Heilongjiang Province, China. Curr. Issues Mol. Biol. 2022, 44, 3194-3207. https://doi.org/10.3390/cimb44070221
Li W, Liu M, Lai Y-C, Liu J-X, Fan C, Yang G, Wang L, Liang W-W, Di S-F, Yu D-Y, et al. Genome-Wide Association Study of Partial Resistance to P. sojae in Wild Soybeans from Heilongjiang Province, China. Current Issues in Molecular Biology. 2022; 44(7):3194-3207. https://doi.org/10.3390/cimb44070221
Chicago/Turabian StyleLi, Wei, Miao Liu, Yong-Cai Lai, Jian-Xin Liu, Chao Fan, Guang Yang, Ling Wang, Wen-Wei Liang, Shu-Feng Di, De-Yue Yu, and et al. 2022. "Genome-Wide Association Study of Partial Resistance to P. sojae in Wild Soybeans from Heilongjiang Province, China" Current Issues in Molecular Biology 44, no. 7: 3194-3207. https://doi.org/10.3390/cimb44070221
APA StyleLi, W., Liu, M., Lai, Y.-C., Liu, J.-X., Fan, C., Yang, G., Wang, L., Liang, W.-W., Di, S.-F., Yu, D.-Y., & Bi, Y.-D. (2022). Genome-Wide Association Study of Partial Resistance to P. sojae in Wild Soybeans from Heilongjiang Province, China. Current Issues in Molecular Biology, 44(7), 3194-3207. https://doi.org/10.3390/cimb44070221