
Propofol and AZD3043 Inhibit Adult Muscle and Neuronal Nicotinic Acetylcholine Receptors Expressed in Xenopus Oocytes


Abstract
:1. Introduction
2. Results
2.1. Basic Nicotinic Acetylcholine Receptor Receptor Pharmacology
2.1.1. Effect of Propofol and AZD3043 on the Adult Human α1β1δε Nicotinic Acetylcholine Receptor Subtype
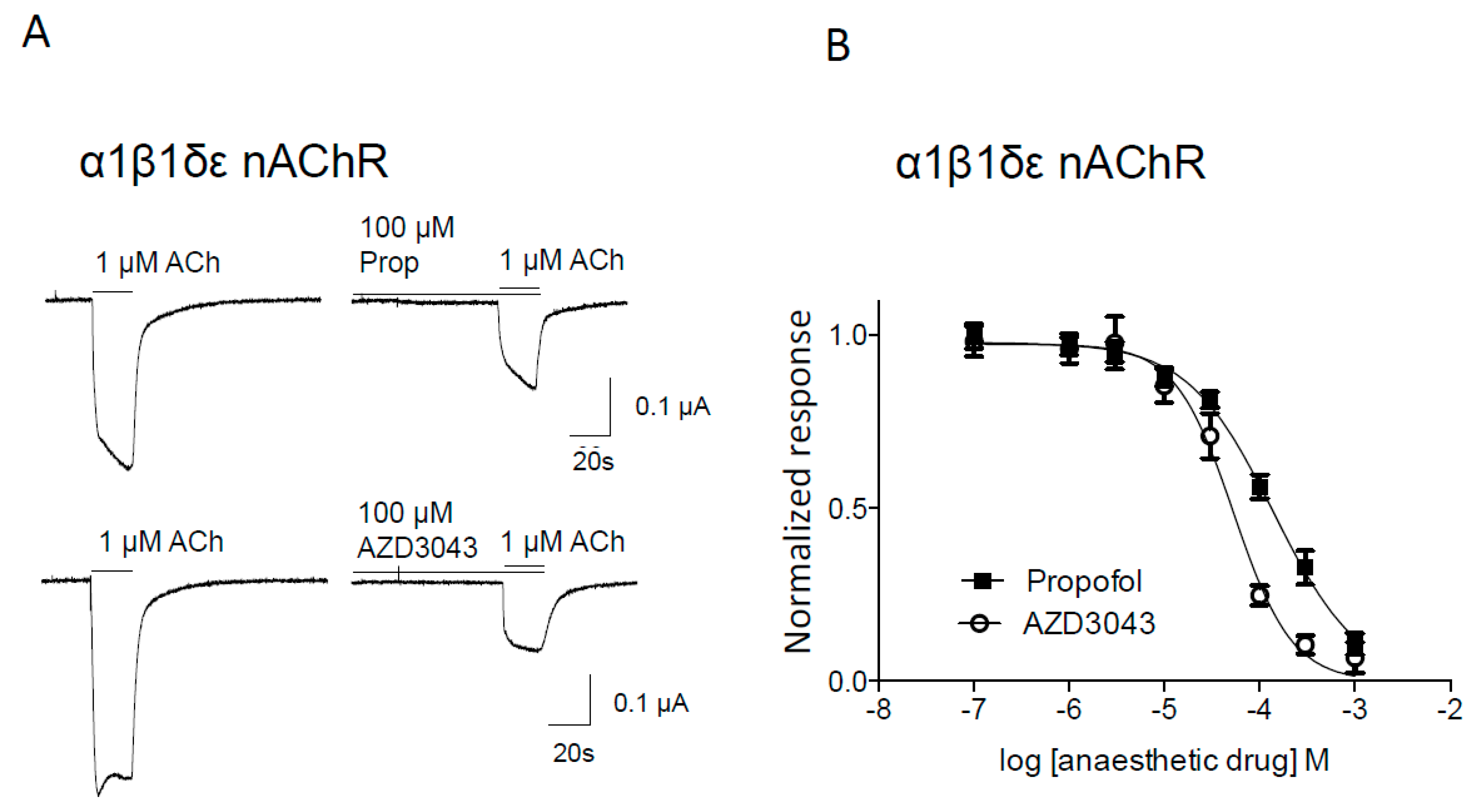

{kind=link}
{kind=link}
| nAChR Subtype | Drug | IC50 (95% C.I.) (μM) | nH (± SEM) | n/N |
|---|---|---|---|---|
| Muscle α1β1δε | Propofol | 139 (112–172) ** | 0.97 ± 0.09 | 8/2 |
| AZD3043 | 53 (41–69) | 1.41 ± 0.21 | 13/2 | |
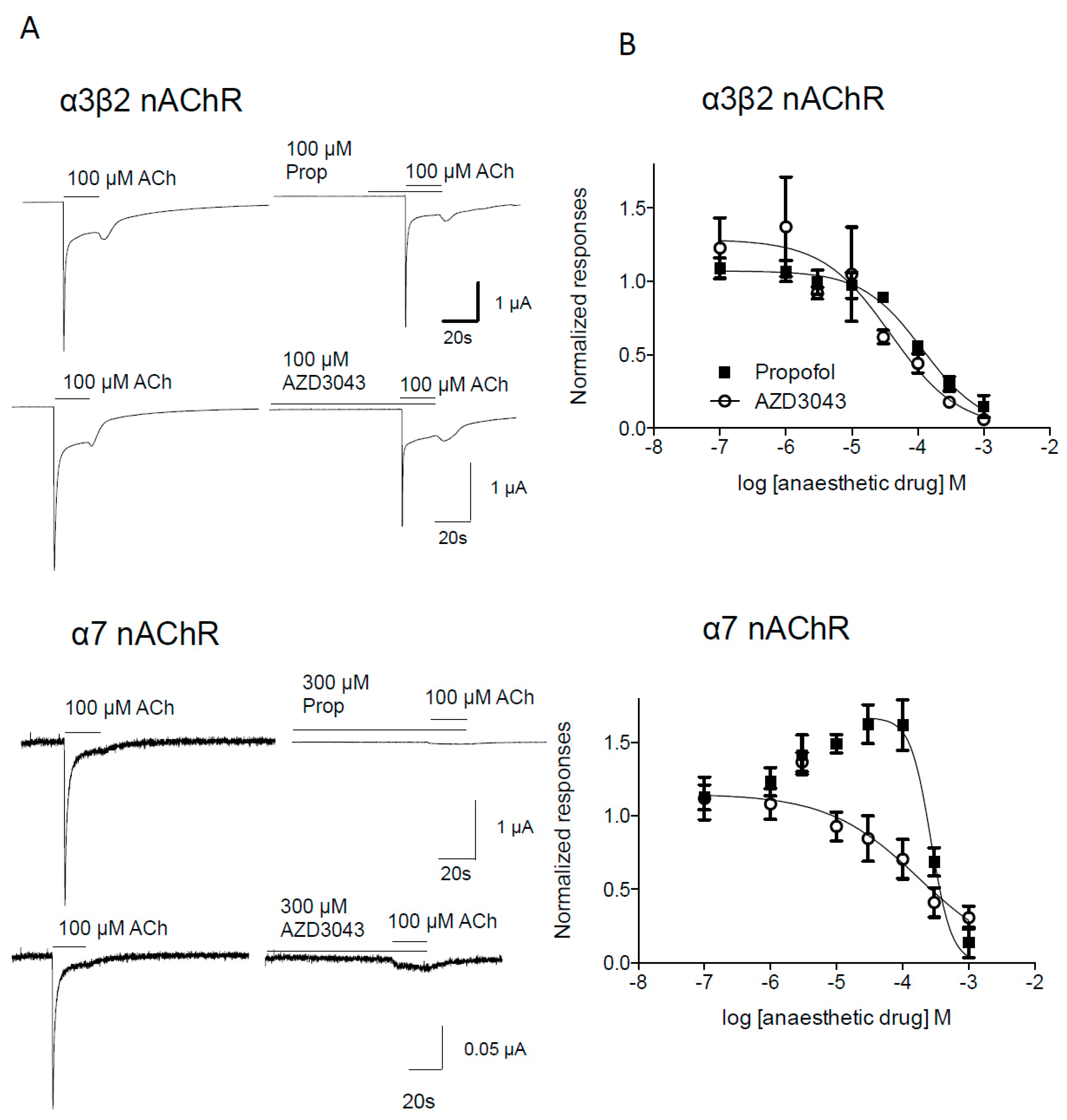
| α7 | Propofol | 269 (200–362) | 2.80 ± 1.49 | 8/2 |
| AZD3043 | 176 (59–527) | 0.63 ± 0.23 | 5/2 | |
| Presynaptic α3β2 | Propofol | 118 (80–175) | 0.95 ± 0.16 | 4/1 |
| AZD3043 | 38 (10–146) | 0.84 ± 0.38 | 5/2 |
2.1.2. Effect of Propofol and AZD3043 on the Human Neuronal α3β2 and α7 Nicotinic Acetylcholine Receptor Subtypes
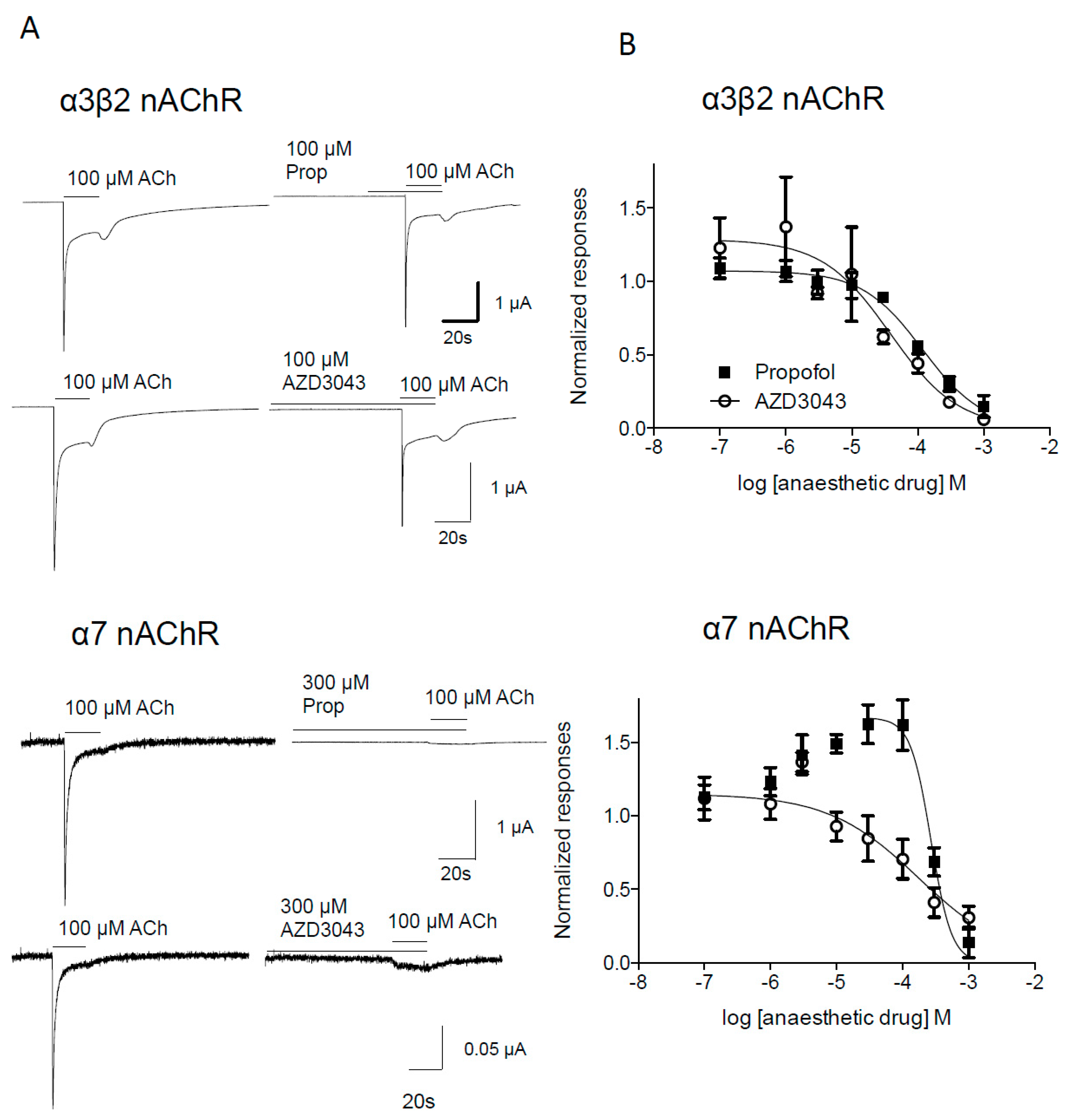
3. Discussion
4. Materials and Methods
4.1. Clones
4.2. Xenopus Oocyte Injection
4.3. Electrophysiological Recordings
4.4. Protocol
4.5. Drugs
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| nAChR | Nicotinic acetylcholine receptor |
| NMJ | Neuromuscular junction |
References
- Franks, N.P. General anaesthesia: From molecular targets to neuronal pathways of sleep and arousal. Nat. Rev. Neurosci. 2008, 9, 370–386. [Google Scholar] [CrossRef] [PubMed]
- Sieghart, W. Structure, pharmacology, and function of GABAA receptor subtypes. Adv. Pharmacol. 2006, 54, 231–263. [Google Scholar] [PubMed]
- Rudolph, U.; Antkowiak, B. Molecular and neuronal substrates for general anaesthetics. Nat. Rev. Neurosci. 2004, 5, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Furuya, R.; Oka, K.; Watanabe, I.; Kamiya, Y.; Itoh, H.; Andoh, T. The effects of ketamine and propofol on neuronal nicotinic acetylcholine receptors and P2x purinoceptors in PC12 cells. Anesth. Analg. 1999, 88, 174–180. [Google Scholar] [PubMed]
- Violet, J.M.; Downie, D.L.; Nakisa, R.C.; Lieb, W.R.; Franks, N.P. Differential sensitivities of mammalian neuronal and muscle nicotinic acetylcholine receptors to general anesthetics. Anesthesiology 1997, 86, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Hurst, R.; Rollema, H.; Bertrand, D. Nicotinic acetylcholine receptors: From basic science to therapeutics. Pharmacol. Ther. 2013, 137, 22–54. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, M.J.; Eriksson, L.I. Current concepts in neuromuscular transmission. Br. J. Anaesth. 2009, 103, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Martyn, J.A.; Richtsfeld, M. Succinylcholine-induced hyperkalemia in acquired pathologic states: Etiologic factors and molecular mechanisms. Anesthesiology 2006, 104, 158–169. [Google Scholar] [CrossRef] [PubMed]
- McKeating, K.; Bali, I.M.; Dundee, J.W. The effects of thiopentone and propofol on upper airway integrity. Anaesthesia 1988, 43, 638–640. [Google Scholar] [CrossRef] [PubMed]
- Barker, P.; Langton, J.A.; Wilson, I.G.; Smith, G. Movements of the vocal cords on induction of anaesthesia with thiopentone or propofol. Br. J. Anaesth. 1992, 69, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Taha, S.; Siddik-Sayyid, S.; Alameddine, M.; Wakim, C.; Dahabra, C.; Moussa, A.; Khatib, M.; Baraka, A. Propofol is superior to thiopental for intubation without muscle relaxants. Can. J. Anaesth. 2005, 52, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, P.R.; Platt, P.R.; Shepherd, K.; Maddison, K.; Hillman, D.R. Collapsibility of the upper airway at different concentrations of propofol anesthesia. Anesthesiology 2005, 103, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Jonsson Fagerlund, M.; Sjodin, J.; Dabrowski, M.A.; Krupp, J. Reduced efficacy of the intravenous anesthetic agent AZD3043 at GABAA receptors with β2 (N289M) and β3 (N290M) point-mutations. Eur. J. Pharmacol. 2012, 694, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Egan, T.D.; Obara, S.; Jenkins, T.E.; Jaw-Tsai, S.S.; Amagasu, S.; Cook, D.R.; Steffensen, S.C.; Beattie, D.T. AZD-3043: A novel, metabolically labile sedative-hypnotic agent with rapid and predictable emergence from hypnosis. Anesthesiology 2012, 116, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Jonsson Fagerlund, M.; Dabrowski, M.; Eriksson, L.I. Pharmacological characteristics of the inhibition of nondepolarizing neuromuscular blocking agents at human adult muscle nicotinic acetylcholine receptor. Anesthesiology 2009, 110, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Dabrowski, M.; Gurley, D.A.; Larsson, O.; Johnson, E.C.; Fredholm, B.B.; Eriksson, L.I. Activation and inhibition of human muscular and neuronal nicotinic acetylcholine receptors by succinylcholine. Anesthesiology 2006, 104, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Gurley, D.; Dabrowski, M.; Larsson, O.; Johnson, E.C.; Eriksson, L.I. Distinct pharmacological properties of neuromuscular blocking agents on human neuronal nicotinic acetylcholine receptors-a possible mechanism behind the train-of-four fade. Anesthesiology 2006, 105, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Jayakar, S.S.; Dailey, W.P.; Eckenhoff, R.G.; Cohen, J.B. Identification of propofol binding sites in a nicotinic acetylcholine receptor with a photoreactive propofol analog. J. Biol. Chem. 2013, 288, 6178–6189. [Google Scholar] [CrossRef] [PubMed]
- Flood, P.; Ramirez-Latorre, J.; Role, L. α4β2 neuronal nicotinic acetylcholine receptors in the central nervous system are inhibited by isoflurane and propofol, but α7-type nicotinic acetylcholine receptors are unaffected. Anesthesiology 1997, 86, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Dueck, M.H.; Oberthuer, A.; Wedekind, C.; Paul, M.; Boerner, U. Propofol impairs the central but not the peripheral part of the motor system. Anesth. Analg. 2003, 96, 449–455. [Google Scholar] [PubMed]
- Haeseler, G.; Stormer, M.; Bufler, J.; Dengler, R.; Hecker, H.; Piepenbrock, S.; Leuwer, M. Propofol blocks human skeletal muscle sodium channels in a voltage-dependent manner. Anesth. Analg. 2001, 92, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- Ginz, H.F.; Zorzato, F.; Iaizzo, P.A.; Urwyler, A. Effect of three anaesthetic techniques on isometric skeletal muscle strength. Br. J. Anaesth. 2004, 92, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; McEwan, A.I.; Jhaveri, R.; Wilkinson, M.; Goodman, D.; Smith, L.R.; Canada, A.T.; Glass, P.S. The interaction of fentanyl on the cp50 of propofol for loss of consciousness and skin incision. Anesthesiology 1994, 81, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Jonsson Fagerlund, M.; Sjodin, J.; Krupp, J.; Dabrowski, M.A. Reduced effect of propofol at human α1β2N289Mγ2 and α2β3N290Mγ2 mutant GABAA receptors. Br. J. Pharmacol. 2010, 104, 472–481. [Google Scholar]
- Richardson, J.E.; Garcia, P.S.; O’Toole, K.K.; Derry, J.M.; Bell, S.V.; Jenkins, A. A conserved tyrosine in the β2 subunit m4 segment is a determinant of γ-aminobutyric acid type a receptor sensitivity to propofol. Anesthesiology 2007, 107, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Lam, D.W.; Reynolds, J.N. Modulatory and direct effects of propofol on recombinant GABAA receptors expressed in xenopus oocytes: Influence of α- and γ2-subunits. Brain Res. 1998, 784, 179–187. [Google Scholar] [CrossRef]
- Eaton, M.J.; Pagan, O.R.; Hann, R.M.; Eterovic, V.A. Differential effects of dimethyl sulfoxide on nicotinic acetylcholine receptors from mouse muscle and torpedo electrocytes. Neurosci. Lett. 1997, 230, 163–166. [Google Scholar] [CrossRef]
- Papke, R.L.; Porter Papke, J.K. Comparative pharmacology of rat and human α7 nAChR conducted with net charge analysis. Br. J. Pharmacol. 2002, 137, 49–61. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jonsson Fagerlund, M.; Krupp, J.; Dabrowski, M.A. Propofol and AZD3043 Inhibit Adult Muscle and Neuronal Nicotinic Acetylcholine Receptors Expressed in Xenopus Oocytes. Pharmaceuticals 2016, 9, 8. https://doi.org/10.3390/ph9010008
Jonsson Fagerlund M, Krupp J, Dabrowski MA. Propofol and AZD3043 Inhibit Adult Muscle and Neuronal Nicotinic Acetylcholine Receptors Expressed in Xenopus Oocytes. Pharmaceuticals. 2016; 9(1):8. https://doi.org/10.3390/ph9010008
Chicago/Turabian StyleJonsson Fagerlund, Malin, Johannes Krupp, and Michael A. Dabrowski. 2016. "Propofol and AZD3043 Inhibit Adult Muscle and Neuronal Nicotinic Acetylcholine Receptors Expressed in Xenopus Oocytes" Pharmaceuticals 9, no. 1: 8. https://doi.org/10.3390/ph9010008
APA StyleJonsson Fagerlund, M., Krupp, J., & Dabrowski, M. A. (2016). Propofol and AZD3043 Inhibit Adult Muscle and Neuronal Nicotinic Acetylcholine Receptors Expressed in Xenopus Oocytes. Pharmaceuticals, 9(1), 8. https://doi.org/10.3390/ph9010008