
In Silico Prediction of Tetrastatin-Derived Peptide Interactions with αvβ3 and α5β1 Integrins

 , and
, and
Abstract

1. Introduction
2. Results
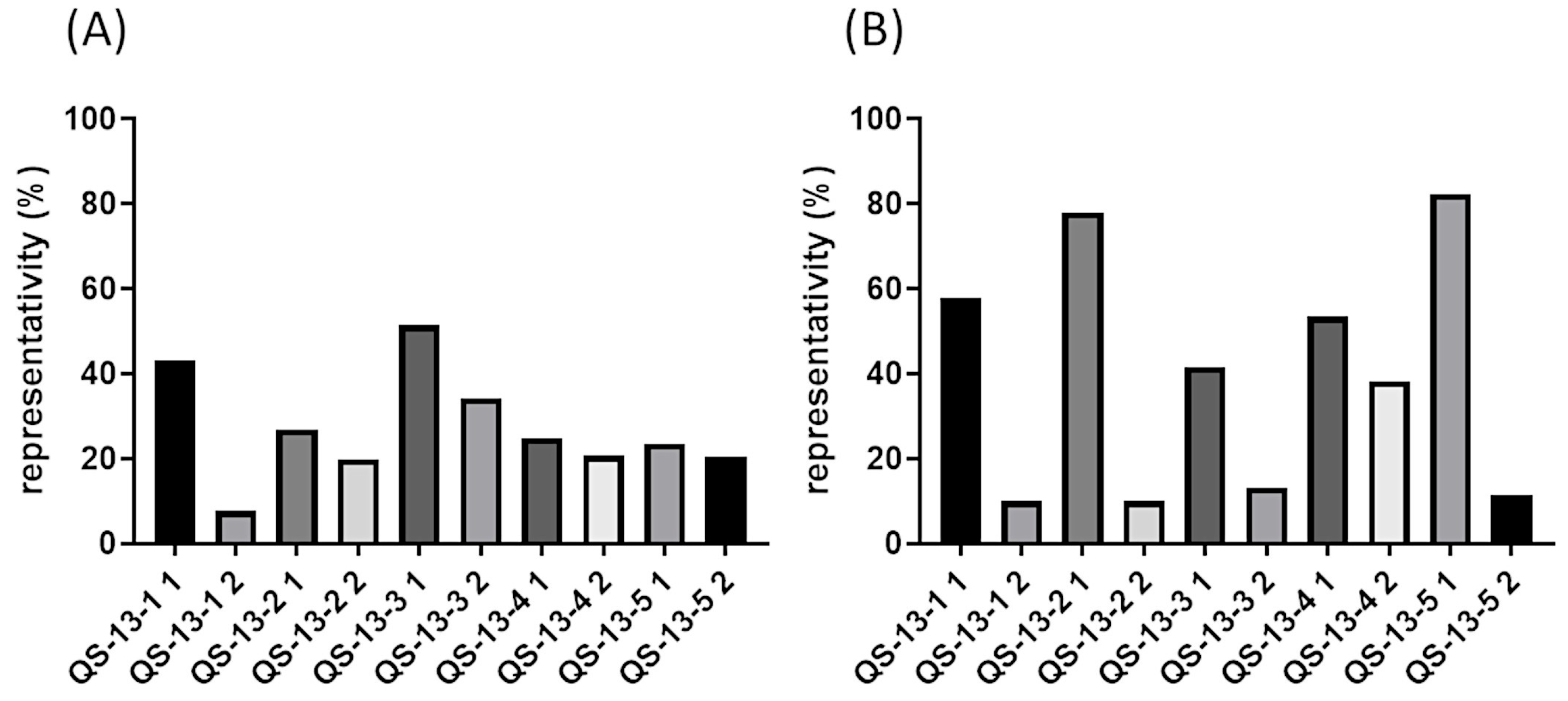
2.1. Substituted Peptide Conformations
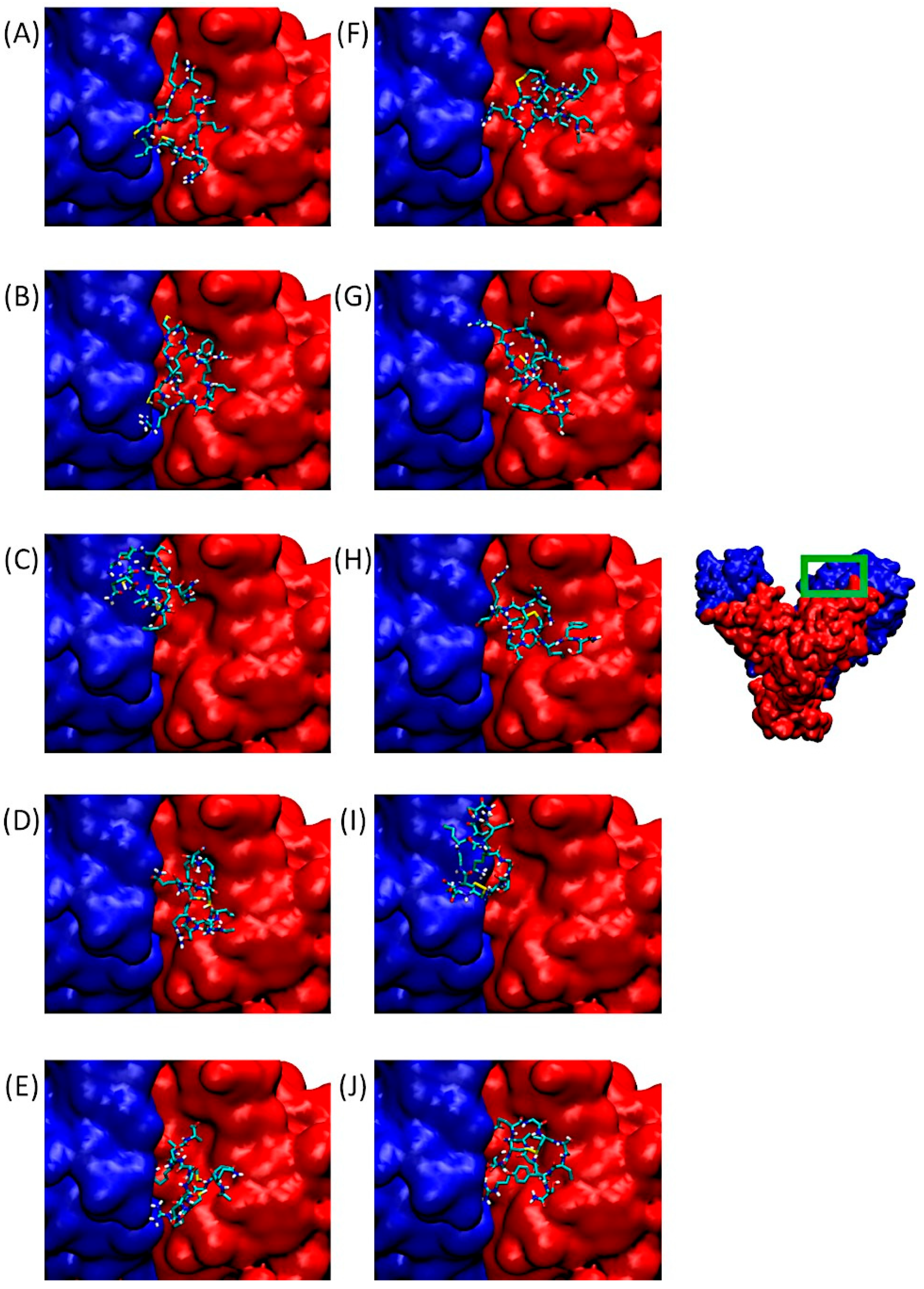
2.2. Docking of Substituted Peptides with αvβ3 Integrin
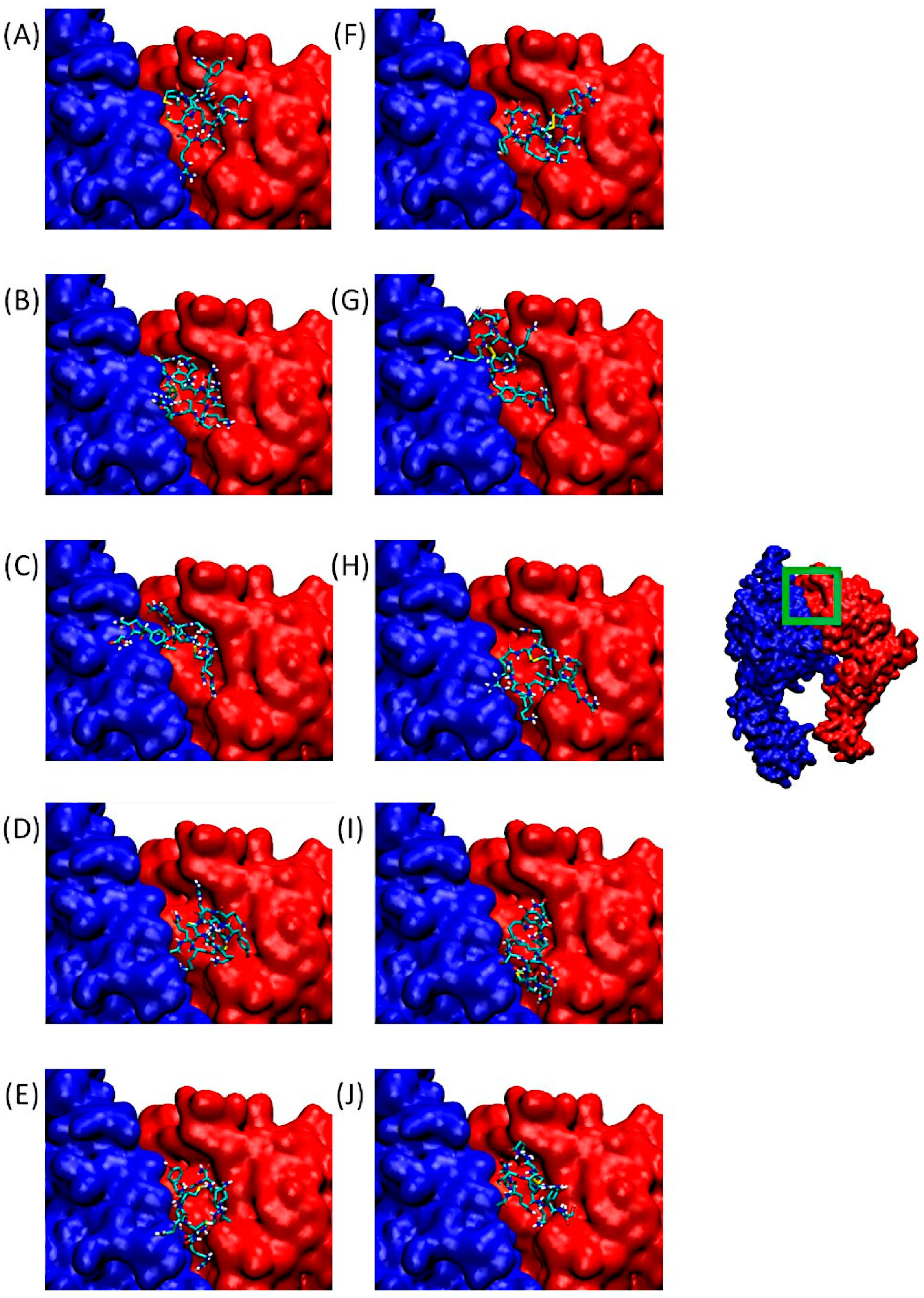
2.3. Docking of Substituted Peptides with α5β1 Integrin
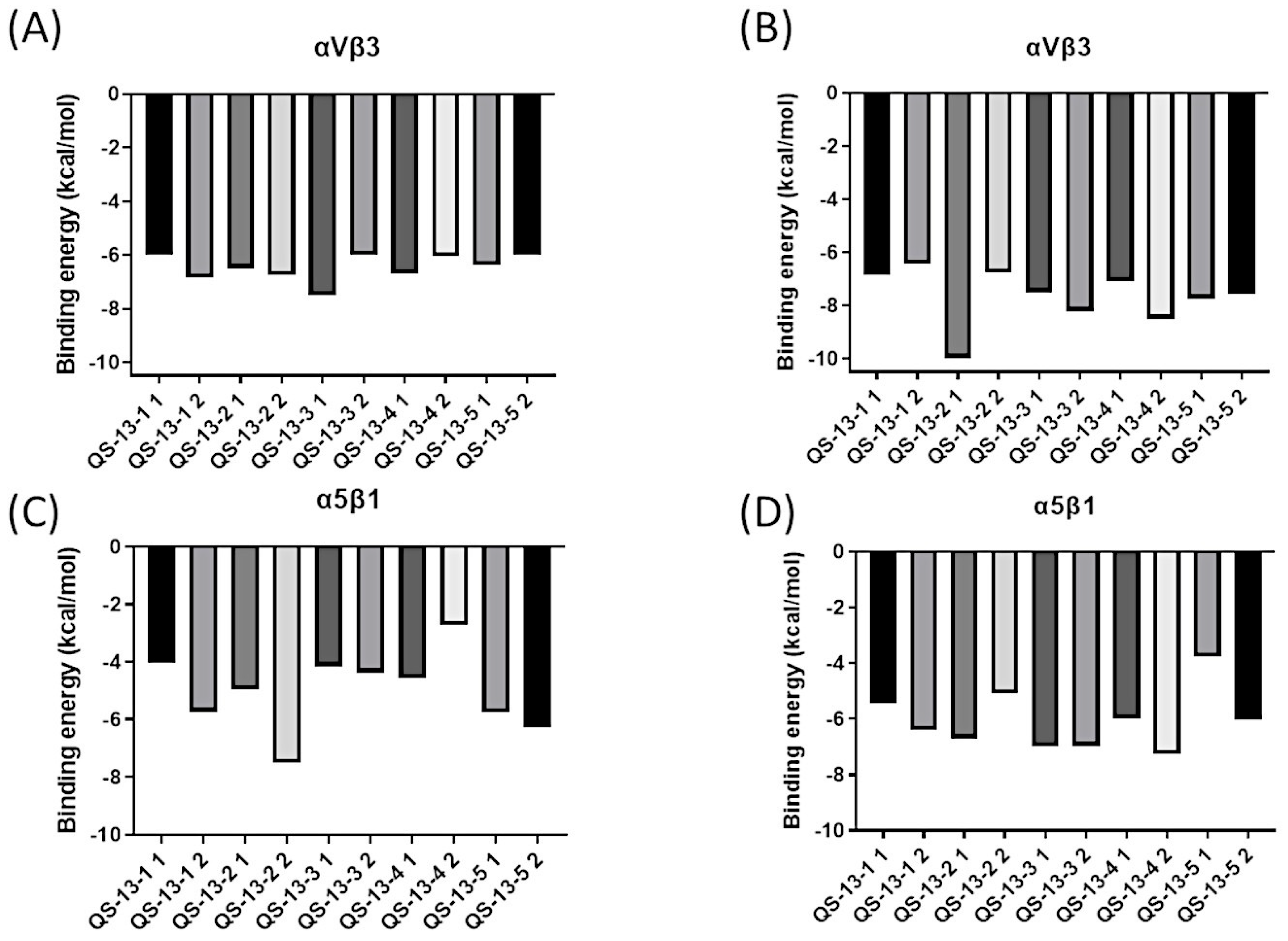
2.4. Binding Energy Analysis
- -
- For αvβ3: −5.70 kcal/mol and −7.40 kcal/mol, respectively, for conformations 1 and 2.
- -
- For α5β1: −5.71 kcal/mol and −7.25 kcal/mol, respectively, for conformations 1 and 2 [21].
2.5. Determination of the Amino Acids Involved in the Interactions
3. Discussion
4. Materials and Methods
4.1. Peptide Modeling and Target Preparation
4.2. Docking Experiments
4.3. Binding Free Energy and Network of Interaction
4.4. Docking Pose Representations
4.5. RMSD Calculations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ECM | Extracellular Matrix |
| EMT | Epithelial–Mesenchymal Transition |
| FN | Fibronectin |
| GROMACS | GROningen MAchine for Chemical Simulations |
| MD | Molecular Dynamics |
| MMPs | Matrix Metalloproteinases |
| NC1 | Non-Collagenous 1 |
| NVT | Number of particles, Volume, and Temperature |
| OPLSAA | Optimized Potentials for Liquid Simulations All-Atom |
| PDB | Protein Data Bank |
| PME | Particle Mesh Ewald |
| RMSD | Root Mean Square Deviation |
| VMD | Visual Molecular Dynamics |
References
- Li, J.; Jo, M.H.; Yan, J.; Hall, T.; Lee, J.; López-Sánchez, U.; Yan, S.; Ha, T.; Springer, T.A. Ligand binding initiates single-molecule integrin conformational activation. Cell 2024, 187, 2990–3005.e17. [Google Scholar] [CrossRef]
- Li, S.; Sampson, C.; Liu, C.; Piao, H.L.; Liu, H.X. Integrin signaling in cancer: Bidirectional mechanisms and therapeutic opportunities. Cell Commun. Signal 2023, 21, 266. [Google Scholar] [CrossRef] [PubMed]
- Han, S.B.; Lee, G.; Kim, D.; Kim, J.K.; Kim, I.S.; Kim, H.W.; Kim, D.H. Selective Suppression of Integrin-Ligand Binding by Single Molecular Tension Probes Mediates Directional Cell Migration. Adv. Sci. 2024, 11, e2306497. [Google Scholar] [CrossRef] [PubMed]
- Kechagia, J.Z.; Ivaska, J.; Roca-Cusachs, P. Integrins as biomechanical sensors of the microenvironment. Nat. Rev. Mol. Cell Biol. 2019, 20, 457–473. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, P.; Li, Q.; Yang, B.; Li, N.; Luo, Z.; Zhang, F. Interaction of integrin αvβ3 and fibronectin under fluid shear forces: Implications for tumor cell adhesion and migration. Front. Cell Dev. Biol. 2025, 13, 1512672. [Google Scholar] [CrossRef]
- Takada, Y.K.; Yu, J.; Ye, X.; Wu, C.Y.; Felding, B.H.; Fujita, M.; Takada, Y. The heparin-binding domain of VEGF165 directly binds to integrin αvβ3 and VEGFR2/KDR D1: A potential mechanism of negative regulation of VEGF165 signaling by αvβ3. Front. Cell Dev. Biol. 2024, 12, 1347616. [Google Scholar] [CrossRef]
- Honda, S.; Kashiwagi, H.; Kiyoi, T.; Kato, H.; Kosugi, S.; Shiraga, M.; Tomiyama, Y. Amino acid mutagenesis within ligand-binding loops in αv confers loss-of-function or gain-of-function phenotype on integrin αvβ3. Thromb. Haemost. 2004, 92, 1092–1098. [Google Scholar] [CrossRef]
- Mori, S.; Hatori, N.; Kawaguchi, N.; Hamada, Y.; Shih, T.C.; Wu, C.Y.; Takada, Y. The integrin-binding defective FGF2 mutants potently suppress FGF2 signalling and angiogenesis. Biosci. Rep. 2017, 37, BSR20170173. [Google Scholar] [CrossRef]
- Pang, X.; He, X.; Qiu, Z.; Zhang, H.; Xie, R.; Liu, Z.; Cui, Y. Targeting integrin pathways: Mechanisms and advances in therapy. Targeting integrin pathways: Mechanisms and advances in therapy. Signal Transduct. Target. Ther. 2023, 8, 1. [Google Scholar] [CrossRef]
- Gu, Y.; Dong, B.; He, X.; Qiu, Z.; Zhang, J.; Zhang, M.; Liu, H.; Pang, X.; Cui, Y. The challenges and opportunities of αvβ3-based therapeutics in cancer: From bench to clinical trials. Pharmacol. Res. 2023, 189, 106694. [Google Scholar] [CrossRef]
- Mould, A.P.; Koper, E.J.; Byron, A.; Zahn, G.; Humphries, M.J. Mapping the ligand-binding pocket of integrin α5β1 using a gain-of-function approach. Biochem. J. 2009, 424, 179–189. [Google Scholar] [CrossRef]
- Puzon-McLaughlin, W.; Takada, Y. Critical Residues for Ligand Binding in an I Domain-like Structure of the Integrin β1 Subunit*. J. Biol. Chem. 1996, 271, 20438–20443. [Google Scholar] [CrossRef]
- Schumacher, S.; Dedden, D.; Nunez, R.V.; Matoba, K.; Takagi, J.; Biertümpfel, C.; Mizuno, N. Structural insights into integrin α5β1 opening by fibronectin ligand. Sci. Adv. 2021, 7, eabe9716. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, B.S.; Kessler, H.; Kossatz, S.; Reuning, U. RGD-Binding Integrins Revisited: How Recently Discovered Functions and Novel Synthetic Ligands (Re-)Shape an Ever-Evolving Field. Cancers 2021, 13, 1711. [Google Scholar] [CrossRef]
- Schaffner, F.; Ray, A.M.; Dontenwill, M. Integrin α5β1, the Fibronectin Receptor, as a Pertinent Therapeutic Target in Solid Tumors. Cancers 2013, 5, 27–47. [Google Scholar] [CrossRef]
- Kapp, T.; Rechenmacher, F.; Neubauer, S.; Maltsev, O.V.; Cavalcanti-Adam, E.A.; Zarka, R.; Reuning, U.; Notni, J.; Wester, H.J.; Mas-Moruno, C.; et al. A Comprehensive Evaluation of the Activity and Selectivity Profile of Ligands for RGD-binding Integrins. Sci. Rep. 2017, 7, 39805. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M.E.; Gorlia, T.; Erridge, S.C.; Perry, J.; Hong, Y.K.; Aldape, K.D.; Lhermitte, B.; Pietsch, T.; Grujicic, D.; et al. Cilengitide combined with standard treatment for patients with newly diagnosed glioblastoma with methylated MGMT promoter (CENTRIC EORTC 26071-22072 study): A multicentre, randomised, open-label, phase 3 trial. Lancet Oncol. 2014, 15, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Bhatwadekar, A.D.; Kansara, V.; Luo, Q.; Ciulla, T. Anti-integrin therapy for retinovascular diseases. Expert. Opin. Investig. Drugs 2020, 29, 935–945. [Google Scholar] [CrossRef]
- Lambert, E.; Fuselier, E.; Ramont, L.; Brassart, B.; Dukic, S.; Oudart, J.B.; Dupont-Deshorgue, A.; Sellier, C.; Machado, C.; Dauchez, M.; et al. Conformation-dependent binding of a Tetrastatin peptide to αvβ3 integrin decreases melanoma progression through FAK/PI3K/Akt pathway inhibition. Sci. Rep. 2018, 8, 9837. [Google Scholar] [CrossRef]
- Brassart-Pasco, S.; Sénéchal, K.; Thevenard, J.; Ramont, L.; Devy, J.; Di Stefano, L.; Dupont-Deshorgue, A.; Brézillon, S.; Feru, J.; Jazeron, J.F.; et al. Tetrastatin, the NC1 domain of the α4(IV) collagen chain: A novel potent anti-tumor matrikine. PLoS ONE 2012, 7, e29587. [Google Scholar] [CrossRef]
- Vautrin-Glabik, A.; Devy, J.; Bour, C.; Baud, S.; Choulier, L.; Hoarau, A.; Dupont-Deshorgue, A.; Sellier, C.; Brassart, B.; Oudart, J.B.; et al. Angiogenesis Inhibition by a Short 13 Amino Acid Peptide Sequence of Tetrastatin, the α4(IV) NC1 Domain of Collagen IV. Front. Cell Dev. Biol. 2020, 8, 775. [Google Scholar] [CrossRef]
- Paturel, V.; Baud, S.; Schneider, C.; Brassart-Pasco, S. In Silico Modeling and Analysis of Physicochemical Properties of Antitumoral QS-13 Peptide and its Derivatives in Water and Dimethyl Sulfoxide. Pept. Sci. 2024, 116, e24352. [Google Scholar] [CrossRef]
- Xiong, J.P.; Stehle, T.; Zhang, R.; Joachimiak, A.; Frech, M.; Goodman, S.L.; Arnaout, M.A. Crystal structure of the extracellular segment of integrin αVβ3 in complex with an Arg-Gly-Asp ligand. Science 2002, 296, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Takagi, J.; Petre, B.M.; Walz, T.; Springer, T.A. Global conformational rearrangements in integrin extracellular domains in outside-in and inside-out signaling. Cell 2002, 110, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Takagi, J.; Strokovich, K.; Springer, T.A.; Walz, T. Structure of integrin α5β1 in complex with fibronectin. EMBO J. 2003, 22, 4607–4615. [Google Scholar] [CrossRef]
- Friedlander, M.; Brooks, P.C.; Shaffer, R.W.; Kincaid, C.M.; Varner, J.A.; Cheresh, D.A. Definition of Two Angiogenic Pathways by Distinct αv Integrins. Science 1995, 270, 1500–1502. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.R.; Zhao, J.T.; Xie, Z.Z. Integrin-mediated cancer progression as a specific target in clinical therapy. Biomed. Pharmacother. 2022, 155, 113745. [Google Scholar] [CrossRef]
- Trabocchi, A.; Menchi, G.; Cini, N.; Bianchini, F.; Raspanti, S.; Bottoncetti, A.; Pupi, A.; Calorini, L.; Guarna, A. Click-chemistry-derived triazole ligands of arginine-glycine-aspartate (RGD) integrins with a broad capacity to inhibit adhesion of melanoma cells and both in vitro and in vivo angiogenesis. J. Med. Chem. 2010, 53, 7119–7128. [Google Scholar] [CrossRef]
- Van Agthoven, J.F.; Xiong, J.P.; Alonso, J.L.; Rui, X.; Adair, B.D.; Goodman, S.L.; Arnaout, M.A. Structural basis for pure antagonism of integrin αVβ3 by a high affinity form of fibronectin. Nat. Struct. Mol. Biol. 2014, 21, 383–388. [Google Scholar] [CrossRef]
- Mori, S.; Wu, C.Y.; Yamaji, S.; Saegusa, J.; Shi, B.; Ma, Z.; Kuwabara, Y.; Lam, K.S.; Isseroff, R.R.; Takada, Y.K.; et al. Direct binding of integrin αv5β3 to FGF1 plays a role in FGF1 signaling. J. Biol. Chem. 2008, 283, 18066–18075. [Google Scholar] [CrossRef]
- Humphries, J.D.; Askari, J.A.; Zhang, X.P.; Takada, Y.; Humphries, M.J.; Mould, A.P. Molecular basis of ligand recognition by integrin α5β1. II. Specificity of arg-gly-Asp binding is determined by Trp157 of the α subunit. J. Biol. Chem. 2000, 275, 20337–20345. [Google Scholar] [CrossRef] [PubMed]
- Mould, A.P.; Burrows, L.; Humphries, M.J. Identification of amino acid residues that form part of the ligand-binding pocket of integrin α5β1. J. Biol. Chem. 1998, 273, 25664–25672. [Google Scholar] [CrossRef]
- Finney, A.C.; Stokes, K.Y.; Pattillo, C.B.; Orr, A.W. Integrin signaling in atherosclerosis. Cell Mol. Life Sci. 2017, 74, 2263–2282. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.C.; Yeh, S.I.; Chen, S.L.; Tsao, Y.P. The Psoriasis Therapeutic Potential of a Novel Short Laminin Peptide C16. Int. J. Mol. Sci. 2019, 20, 3144. [Google Scholar] [CrossRef] [PubMed]
- Morshed, A.; Abbas, A.B.; Hu, J.; Xu, H. Shedding New Light on The Role of ανβ3 and α5β1 Integrins in Rheumatoid Arthritis. Molecules 2019, 24, 1537. [Google Scholar] [CrossRef]
- Lin, T.H.; Yang, R.S.; Tu, H.J.; Liou, H.C.; Lin, Y.M.; Chuang, W.J.; Fu, W.M. Inhibition of osteoporosis by the αvβ3 integrin antagonist of rhodostomin variants. Eur. J. Pharmacol. 2017, 804, 94–101. [Google Scholar] [CrossRef]
- Jin, D.K.; Fish, A.J.; Wayner, E.A.; Mauer, M.; Setty, S.; Tsilibary, E.; Kim, Y. Distribution of integrin subunits in human diabetic kidneys. J. Am. Soc. Nephrol. 1996, 7, 2636–2645. [Google Scholar] [CrossRef]
- Friedlander, M.; Theesfeld, C.L.; Sugita, M.; Fruttiger, M.; Thomas, M.A.; Chang, S.; Cheresh, D.A. Involvement of integrins αvβ3 and αvβ5 in ocular neovascular diseases. Proc. Natl. Acad. Sci. USA 1996, 93, 9764–9769. [Google Scholar] [CrossRef]
- Slack, R.J.; Macdonald, S.J.F.; Roper, J.A.; Jenkins, R.G.; Hatley, R.J.D. Emerging therapeutic opportunities for integrin inhibitors. Nat. Rev. Drug Discov. 2022, 21, 60–78. [Google Scholar] [CrossRef]
- Lima ESilva, R.; Mirando, A.C.; Tzeng, S.Y.; Green, J.J.; Popel, A.S.; Pandey, N.B.; Campochiaro, P.A. Anti-angiogenic collagen IV-derived peptide target engagement with αvβ3 and α5β1 in ocular neovascularization models. iScience 2023, 26, 106078. [Google Scholar] [CrossRef]
- Bretaudeau, C.; Baud, S.; Dupont-Deshorgue, A.; Cousin, R.; Brassart, B.; Brassart-Pasco, S. AG-9, an Elastin-Derived Peptide, Increases In Vitro Oral Tongue Carcinoma Cell Invasion, through an Increase in MMP-2 Secretion and MT1-MMP Expression, in a RPSA-Dependent Manner. Biomolecules 2020, 11, 39. [Google Scholar] [CrossRef]
- Zhang, Y.; Sanner, M.F. AutoDock CrankPep: Combining folding and docking to predict protein-peptide complexes. Bioinformatics 2019, 35, 5121–5127. [Google Scholar] [CrossRef] [PubMed]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- GROMACS 2023.3 Manual. Available online: https://zenodo.org/records/10017699 (accessed on 1 September 2024).
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Madura, J.D. Quantum and Statistical Mechanical Studies of Liquids. 25. Solvation and Conformation of Methanol in Water. J. Am. Chem. Soc. 1983, 105, 1407–1413. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Maxwell, D.S.; Tirado-Rives, J. Development and Testing of the OPLS All-Atom Force Field on Conformational Energetics and Properties of Organic Liquids. J. Am. Chem. Soc. 1996, 118, 11225–11236. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A Linear Constraint Solver for Molecular Simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical Sampling Through Velocity Rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [PubMed]
- Essmann, U.; Perera, L.; Berkowitz, M.; Darden, T.; Lee, H.; Pedersen, L. A Smooth Particle Mesh Ewald Method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Parrinello, M.; Rahman, A. Polymorphic Transitions in Single Crystals: A New Molecular Dynamics Method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Brigida Bochicchio, B.; Pepe, A.; Crudele, M.; Belloy, N.; Baud, S.; Dauchez, M. Tuning Self-Assembly in Elastin-Derived Peptides. Soft Matter. 2015, 11, 3385–3395. [Google Scholar] [CrossRef] [PubMed]
- Oudart, J.; Doué, M.; Vautrin, A.; Brassart, B.; Sellier, C.; Dupont-Deshorgue, A.; Monboisse, J.C.; Maquart, F.X.; Brassart-Pasco, S.; Ramont, L. The anti-tumor NC1 domain of collagen XIX inhibits the FAK/ PI3K/Akt/mTOR signaling pathway through αvβ3 integrin interaction. Oncotarget 2016, 7, 1516–1528. [Google Scholar] [CrossRef]
- Dauvé, J.; Belloy, N.; Rivet, R.; Etique, N.; Nizet, P.; Pietraszek-Gremplewicz, K.; Karamanou, K.; Dauchez, M.; Ramont, L.; Brézillon, S.; et al. Differential MMP-14 Targeting by Lumican-Derived Peptides Unraveled by In Silico Approach. Cancers 2021, 13, 4930. [Google Scholar] [CrossRef]
- Bank, R.P.D. RCSB PDB—4G1M: Re-Refinement of Alpha V Beta 3 Structure. Available online: https://www.rcsb.org/structure/4g1m (accessed on 1 September 2024).
- Bank, R.P.D. RCSB PDB—3VI3: Crystal Structure of Alpha5beta1 Integrin Headpiece (Ligand-Free Form). Available online: https://www.rcsb.org/structure/3vi3 (accessed on 1 September 2024).
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph Model 1999, 17, 57–61. [Google Scholar] [PubMed]
- Bitencourt-Ferreira, G.; Pintro, V.O.; de Azevedo, W.F. Docking with AutoDock4. Methods Mol. Biol. 2019, 2053, 125–148. [Google Scholar] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- Durrant, J.D.; McCammon, J.A. BINANA: A novel algorithm for ligand-binding characterization. J. Mol. Graph Model 2011, 29, 888–893. [Google Scholar] [CrossRef]
- Bank, R.P.D. RCSB PDB—3VI4: Crystal Structure of Alpha5beta1 Integrin Headpiece in Complex with RGD Peptide. Available online: https://www.rcsb.org/structure/3vi4 (accessed on 1 September 2024).
- Bank, R.P.D. RCSB PDB—1L5G: Crystal Structure of the Extracellular Segment of Integrin AVB3 in Complex with an ARG-GLY-ASP Ligand. Available online: https://www.rcsb.org/structure/1L5G (accessed on 1 September 2024).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB ID | Source | Name of the Protein and Experimental Technics | 3D Structure |
|---|---|---|---|
| 4G1M | Homo sapiens | αvβ3 integrin, X-ray diffraction (2.90 Å resolution) | 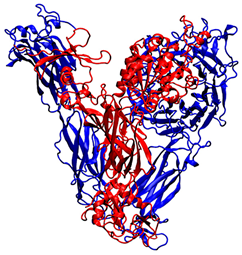 |
| 3VI3 | Homo sapiens | α5β1 integrin, X-ray diffraction (2.90 Å resolution) | 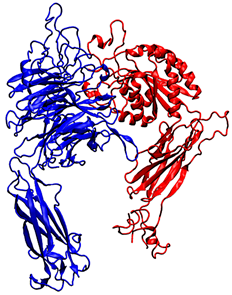 |
| Peptide | Sequence |
| QS-13 | QKISRCQVCVKYS |
| QS-13-1 | QKSSRCQVCVKYS |
| QS-13-2 | QKSSRCQGCGKYS |
| QS-13-3 | QKGSRCQGCVKYS |
| QS-13-4 | QKGSRCQGCGKYS |
| QS-13-5 | QKGSRCQGCSKYS |
| Most Representative Conformation | Integrin α5β1 | Integrin αvβ3 | ||
| With Disulfide Bridge | Without Disulfide Bridge | With Disulfide Bridge | Without Disulfide Bridge | |
| QS-13-1 | −5.44 | −4.05 | −6.84 | −5.97 |
| −5.25 | −3.56 | −6.67 | −5.90 | |
| −4.71 | −2.89 | −6.58 | −5.45 | |
| QS-13-2 | −6.70 | −4.97 | −9.98 | −6.50 |
| −6.58 | −4.92 | −9.65 | −6.19 | |
| −6.12 | −4.65 | −9.48 | −5.97 | |
| QS-13-3 | −6.98 | −4.16 | −7.49 | −7.52 |
| −6.40 | −2.37 | −7.49 | −5.72 | |
| −6.38 | −2.17 | −7.21 | −5.51 | |
| QS-13-4 | −5.99 | −4.57 | −7.11 | −6.71 |
| −5.42 | −4.48 | −6.10 | −5.95 | |
| −4.93 | −4.08 | −5.92 | −5.75 | |
| QS-13-5 | −3.76 | −5.76 | −7.74 | −5.79 |
| −3.72 | −5.12 | −6.40 | −5.64 | |
| −3.60 | −4.12 | −6.27 | −5.62 | |
| Second Most Representative Conformation | Integrin α5β1 | Integrin αVβ3 | ||
| With Disulfide Bridge | Without Disulfide Bridge | With Disulfide Bridge | Without Disulfide Bridge | |
| QS-13-1 | −6.39 | −5.73 | −6.43 | −6.86 |
| −4.19 | −5.17 | −6.10 | −6.36 | |
| −3.66 | −3.88 | −5.82 | −5.78 | |
| QS-13-2 | −5.09 | −7.52 | −6.75 | −6.77 |
| −5.03 | −4.31 | −6.38 | −6.63 | |
| −4.92 | −4.00 | −6.30 | −5.90 | |
| QS-13-3 | −6.96 | −4.38 | −8.22 | −6.00 |
| −5.77 | −4.02 | −7.95 | −5.89 | |
| −5.58 | −3.79 | −7.78 | −5.87 | |
| QS-13-4 | −7.27 | −2.74 | −8.50 | −6.03 |
| −6.78 | −2.62 | −8.15 | −5.18 | |
| −6.55 | −2.45 | −7.78 | −4.86 | |
| QS-13-5 | −6.01 | −6.26 | −7.56 | −7.84 |
| −6.01 | −6.22 | −7.39 | −7.71 | |
| −5.71 | −5.46 | −7.35 | −7.09 | |
| Peptides Without Disulfide Bond—Most Representative Conformation | αv | β3 |
|---|---|---|
| QS-13-1 | Hydrophobic contact | |
| Cys9—Ala215 | ||
| Hydrogen bond | ||
| Val8 (C=O)—Ala215 (N) | ||
| Val8 (C=O)—Ile216 (N) | ||
| QS-13-2 | Hydrophobic contact | |
| Arg5—Ile216 | Cys9—Arg214 | |
| Cys6—Ala215 | ||
| Gly8—Tyr178 | ||
| Cys9—Ty178 | ||
| Hydrogen bond | ||
| Cys9 (C=0)—Arg214 (N sidechain) | ||
| QS-13-3 | Hydrophobic contact | |
| Val10—Trp179 | Ser13—Ala218 | |
| Lys11—Tyr178 | ||
| Lys11—Trp179 | ||
| Tyr12—Tyr178 | ||
| Tyr12—Asp219 | ||
| Hydrogen bond | ||
| Ser13 (C=O)—Ala 218 (N) | ||
| QS-13-4 | Hydrophobic contact | |
| Nter—Met335 | ||
| Gly3—Asp251 | ||
| Ser4—Asn313 | ||
| Lys11—Tyr122 | ||
| Tyr12—Arg214 | ||
| Cter—Met180 | ||
| Hydrogen bond | ||
| Lys11 (C=O)—Arg214 (N sidechain) | ||
| QS-13-5 | Hydrophobic contact | |
| Nter—Met335 | ||
| Gly3—Asn313 | ||
| Ser4—Asn313 | ||
| Arg5—Asn313 | ||
| Ser13—Arg214 | ||
| Ser13—Asn215 | ||
| Hydrogen bond | ||
| Cter (N)—Asn215 (O sidechain) | ||
| Cter (N)—Arg216 (O sidechain) | ||
| Peptides Without Disulfide Bond—Second Most Representative Conformation | αv | β3 |
|---|---|---|
| QS-13-1 | Hydrophobic contact | |
| Arg5—Ala218 | ||
| Cys6—Ala215 | ||
| Gln7—Tyr178 | ||
| Val8—Tyr178 | ||
| Tyr12—Asp148 | ||
| Hydrogen bond | ||
| Cter (N)—Arg216 (N sidechain) | ||
| QS-13-2 | Hydrophobic contact | |
| Nter—Tyr178 | ||
| Gln1—Ala215 | ||
| Lys2—Tyr178 | ||
| Hydrogen bond | ||
| Gln1 (N)—Asp218 (O sidechain) | ||
| QS-13-3 | Hydrophobic contact | |
| Nter—Ser123 | ||
| Nter—Asp251 | ||
| Ser4—Ser123 | ||
| Arg5—Ser123 | ||
| Val10—Met180 | ||
| Ser13—Asn215 | ||
| Ser13—Arg216 | ||
| Ser13—Asp217 | ||
| Ser13—Ala218 | ||
| Hydrogen bond | ||
| Gln1 (N)—Asp251 (C=O) | ||
| QS-13-4 | Hydrophobic contact | |
| Nter—Ala215 | ||
| Nter—Asp218 | ||
| QS-13-5 | Hydrophobic contact | |
| Nter—Met335 | ||
| Gln1—Asp251 | ||
| Ser13—Asn313 | ||
| Cter—Asn313 | ||
| Hydrogen bond | ||
| Nter (C=O)—Thr311 (O sidechain) | ||
| Peptides with Disulfide Bond—Most Representative Conformation | αv | β3 |
|---|---|---|
| QS-13-1 | Hydrophobic contact | |
| Gln7—Ser123 | ||
| Val8—Tyr122 | ||
| Cys9—Met180 | ||
| QS-13-2 | Hydrophobic contact | |
| Arg5—Tyr178 | ||
| QS-13-3 | Hydrophobic contact | |
| Ser4—Tyr178 | ||
| Arg5—Tyr178 | ||
| QS-13-4 | Hydrophobic contact | |
| Cys6—Tyr178 | ||
| Gln7—Ala215 | ||
| Lys11—Asp150 | ||
| Tyr12—Asp148 | ||
| Hydrogen bond | ||
| Ser13 (C=O)—Lys119 (N sidechain) | ||
| QS-13-5 | Hydrophobic contact | |
| Gly3—Tyr178 | Gln7—Arg214 | |
| Hydrogen bond | ||
| Gln7 (C=O)—Arg214 (N sidechain) | ||
| Peptides with Disulfide Bond—Second Most Representative Conformation | αv | β3 |
|---|---|---|
| QS-13-1 | Hydrophobic contact | |
| Lys11—Ala215 | ||
| Ser13—Thr212 | ||
| Ser13—Ala215 | ||
| Hydrogen bond | ||
| Cter (N)—Thr212 (O sidechain) | ||
| Cter (N)—Ala213 (C=O) | ||
| QS-13-2 | Hydrophobic contact | |
| Nter—Tyr122 | ||
| Gln1—Ser123 | ||
| Lys2—Ser123 | ||
| Ser13—Asp126 | ||
| Ser13—Met335 | ||
| Tyr12—Asp126 | ||
| Tyr12—Met335 | ||
| Cter—Asp336 | ||
| QS-13-3 | Hydrophobic contact | |
| Nter—Tyr178 | ||
| Nter—Ala215 | ||
| Gln1—Ala215 | ||
| Hydrogen bond | ||
| Gln1 (N)—Asp218 (O sidechain) | ||
| QS-13-4 | Hydrophobic contact | |
| Gly3—Met335 | ||
| Ser4—Met180 | ||
| Ser4—Asn313 | ||
| Ser4—Met335 | ||
| Gly8—Met180 | ||
| Cys9—Tyr122 | ||
| Cys9—Ser123 | ||
| Gly10—Arg214 | ||
| Lys11—Asn215 | ||
| Tyr12—Ala218 | ||
| Hydrogen bond | ||
| Gly10 (C=O)—Asn215 (N) | ||
| QS-13-5 | Hydrophobic contact | |
| Gly8—Tyr178 | ||
| Cys9—Tyr178 | ||
| Peptides Without Disulfide Bond—Most Representative Conformation | α5 | β1 |
|---|---|---|
| QS-13-1 | Hydrophobic contact | |
| Ser3—Phe187 | Arg5—Gln191 | |
| Val8—Ser134 | ||
| Hydrogen bond | ||
| Ser4 (C=O)—Ser134 (N) | ||
| Val10 (C=O)—Lys182 (N sidechain) | ||
| Val10 (C=O)—Gln191 (N sidechain) | ||
| QS-13-2 | Hydrophobic contact | |
| Gly6—Trp157 | Cys9—Tyr133 | |
| Gln7—Phe187 | ||
| Hydrogen bond | ||
| Cys9 (C=O)—Lys182 (N sidechain) | ||
| Ser13 (C=O)—Ser132 (O sidechain) | ||
| Ser13 (C=O)—Tyr133 (N) | ||
| Ser13 (C=O)—Ser134 (N) | ||
| Ser13 (C=O)—Ser134 (O sidechain) | ||
| QS-13-3 | Hydrophobic contact | |
| Cys9—Trp157 | ||
| Val10—Trp157 | ||
| Lys11—Trp157 | ||
| Lys11—Ala159 | ||
| Hydrogen bond | ||
| Gln7 (C=O)—Tyr133 (N) | ||
| QS-13-4 | Hydrophobic contact | |
| Nter—Phe187 | Gly3—Ser134 | |
| Nter—Asp227 | Gly3—Ser227 | |
| Hydrogen bond | ||
| Nter (C=O)—Ser227 (O sidechain) | ||
| Gly3—(C=O)—Ser134 (O sidechain) | ||
| QS-13-5 | Hydrophobic contact | |
| Ser4—Asp227 | Cter—Asn224 | |
| Arg5—Ale225 | Cter—Ser227 | |
| Lys11—Phe187 | ||
| Hydrogen bond | ||
| Cys6 (C=O)—Ser224 (O sidechain) | Cter (N)—Asn224 (C=O) | |
| Cter (N)—Asn224 (N) | ||
| Gly3 (C=O)—Ser227 (O sidechain) | ||
| Ser13 (N)—Asn224 (C=O) | ||
| Peptides Without Disulfide Bond—Second Most Representative Conformation | α5 | β1 |
|---|---|---|
| QS-13-1 | Hydrophobic contact | |
| Nter—Gly255 | ||
| Nter—Asn256 | ||
| Nter—Leu257 | ||
| Gln1—Ile225 | ||
| Gln1—Leu257 | ||
| QS-13-2 | Hydrophobic contact | |
| Nter—Trp157 | Nter—Ser177 | |
| Gln1—Pro186 | ||
| Gln1—Ser177 | ||
| Hydrogen bond | ||
| Lys2 (C=O)—Trp157 (N sidechain) | Nter (C=O)—Ser177 (N) | |
| Gln1 (N)—Pro186 (C=O) | ||
| QS-13-3 | Hydrophobic contact | |
| Lys11—Phe187 | ||
| Hydrogen bond | ||
| Ser4 (C=O)—Gln191 (N sidechain) | ||
| Ser13 (C=O)—Tyr133 (N) | ||
| QS-13-4 | Hydrophobic contact | |
| Nter—Ile225 | ||
| Gln1—Asp227 | ||
| Hydrogen bond | ||
| Gln1 (N)—Ser224 (C=O) | ||
| QS-13-5 | Hydrophobic contact | |
| Cys6—Trp157 | Gln7—Leu225 | |
| Gly8—Trp157 | Ser10—Leu225 | |
| Hydrogen bond | ||
| Gln7 (C=O)—Lys182 (N sidechain) | ||
| Ser10 (N)—Leu225 (C=O) | ||
| Peptides with Disulfide Bond—Most Representative Conformation | α5 | β1 |
|---|---|---|
| QS-13-1 | Hydrophobic contact | |
| Tyr12—Phe187 | Ser4—Tyr133 | |
| Hydrogen bond | ||
| Ser4 (N)—Tyr133 (C=O) | ||
| QS-13-2 | Hydrophobic contact | |
| Nter—Phe187 | ||
| Gln1—Phe187 | ||
| Lys2—Leu128 | ||
| Gly10—Phe187 | ||
| Lys 11—Phe187 | ||
| Gln12—Trp157 | ||
| Hydrogen bond | ||
| Cys9 (C=O)—Trp157 (N sidechain) | Gln1 (C=O)—Lys182 (N sidechain) | |
| Ser4 (C=O)—Lys182 (N sidechain) | ||
| Ser13 (C=O)—Ser132 (O sidechain) | ||
| Ser13 (C=O)—Ser134 (O sidechain) | ||
| QS-13-3 | Hydrophobic contact | |
| Ser4—Phe187 | Nter—Ser134 | |
| Arg5—Phe187 | Nter—Ser227 | |
| —Asp227 | ||
| Hydrogen bond | ||
| Nter (C=O)—Ser224 (O sidechain) | Gly3 (C=O)—Ser227 (O sidechain) | |
| Gln7 (C=O)—Ser134 (N) | ||
| QS-13-4 | Hydrophobic contact | |
| Nter—Phe187 | Nter—Glu320 | |
| Cys6—Ser224 | ||
| Cys6—Ile225 | ||
| Cys9—Ser224 | ||
| Cys9—Ile225 | ||
| Cys9—Asp227 | ||
| Hydrogen bond | ||
| Nter (C=O)—Ser227 (O sidechain) | ||
| Gly8 (N)—Glu320 (O sidechain) | ||
| Lys11 (N))—Ser227 (O sidechain) | ||
| QS-13-5 | Hydrophobic contact | |
| Nter—Phe187 | ||
| Hydrogen bond | ||
| Ser10 (C=O)—Ser227 (O sidechain) | ||
| Lys11 (C=O)—Ser134 (O sidechain) | ||
| Peptides with Disulfide Bond—Second Most Representative Conformation | α5 | β1 |
|---|---|---|
| QS-13-1 | Hydrophobic contact | |
| Cys6—Ala159 | Gln1—Asn224 | |
| Val 8—Asp227 | ||
| Cys9—Phe187 | ||
| Cys9—Asp227 | ||
| Tyr12—Ser224 | ||
| Hydrogen bond | ||
| Lys11 (C=O)—Ser224 (O sidechain) | Gln1 (N)—Asn224 (C=O) | |
| QS-13-2 | Hydrophobic contact | |
| Cys6—Phe187 | ||
| Gly10—Ser224 | ||
| Lys11—Ser224 | ||
| Tyr12—Ser224 | ||
| Ser13—Ser224 | ||
| Hydrogen bond | ||
| Cter (N)—Ile (C=O) | ||
| Arg5 (C=O)—Trp157 (N sidechain) | ||
| QS-13-3 | Hydrophobic contact | |
| Lys2—Trp157 | Gln7—Lys182 | |
| Lys2—Asp227 | ||
| Gly3—Phe187 | ||
| Gly3—Ser224 | ||
| Gly3—Asp227 | ||
| Gly8—trp157 | ||
| Cys9—Trp157 | ||
| Cter—Phe187 | ||
| Cter—Ser224 | ||
| Hydrogen bond | ||
| Ser13 (C=O)—Gln221 (N sidechain) | Gln7 (C=O)—Lys182 ( N sidechain) | |
| QS-13-4 | Hydrophobic contact | |
| Tyr12—Phe187 | ||
| Hydrogen bond | ||
| Gln7 (C=O)—Gln191 (N sidechain) | ||
| Gly8 (C=O)—Tyr133 (N) | ||
| Cys9 (C=O)—Ser134 (O sidechain) | ||
| QS-13-5 | Hydrophobic contact | |
| Gly8—Ala159 | Ser13—Lys182 | |
| Ser10—Phe187 | ||
| Lys11—Phe187 | ||
| Tyr12—Phe187 | ||
| Ser13—Trp157 | ||
| Hydrogen bond | ||
| Ser13 (C=O)—Lys182 (N sidechain) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paturel, V.; Baud, S.; Schneider, C.; Brassart-Pasco, S. In Silico Prediction of Tetrastatin-Derived Peptide Interactions with αvβ3 and α5β1 Integrins. Pharmaceuticals 2025, 18, 940. https://doi.org/10.3390/ph18070940
Paturel V, Baud S, Schneider C, Brassart-Pasco S. In Silico Prediction of Tetrastatin-Derived Peptide Interactions with αvβ3 and α5β1 Integrins. Pharmaceuticals. 2025; 18(7):940. https://doi.org/10.3390/ph18070940
Chicago/Turabian StylePaturel, Vivien, Stéphanie Baud, Christophe Schneider, and Sylvie Brassart-Pasco. 2025. "In Silico Prediction of Tetrastatin-Derived Peptide Interactions with αvβ3 and α5β1 Integrins" Pharmaceuticals 18, no. 7: 940. https://doi.org/10.3390/ph18070940
APA StylePaturel, V., Baud, S., Schneider, C., & Brassart-Pasco, S. (2025). In Silico Prediction of Tetrastatin-Derived Peptide Interactions with αvβ3 and α5β1 Integrins. Pharmaceuticals, 18(7), 940. https://doi.org/10.3390/ph18070940