
Osteogenic Potential of Simvastatin and Fluvastatin in an Organotypic Bone Model


 ,
, 
Abstract
1. Introduction
2. Results
2.1. Microtomographic Analysis
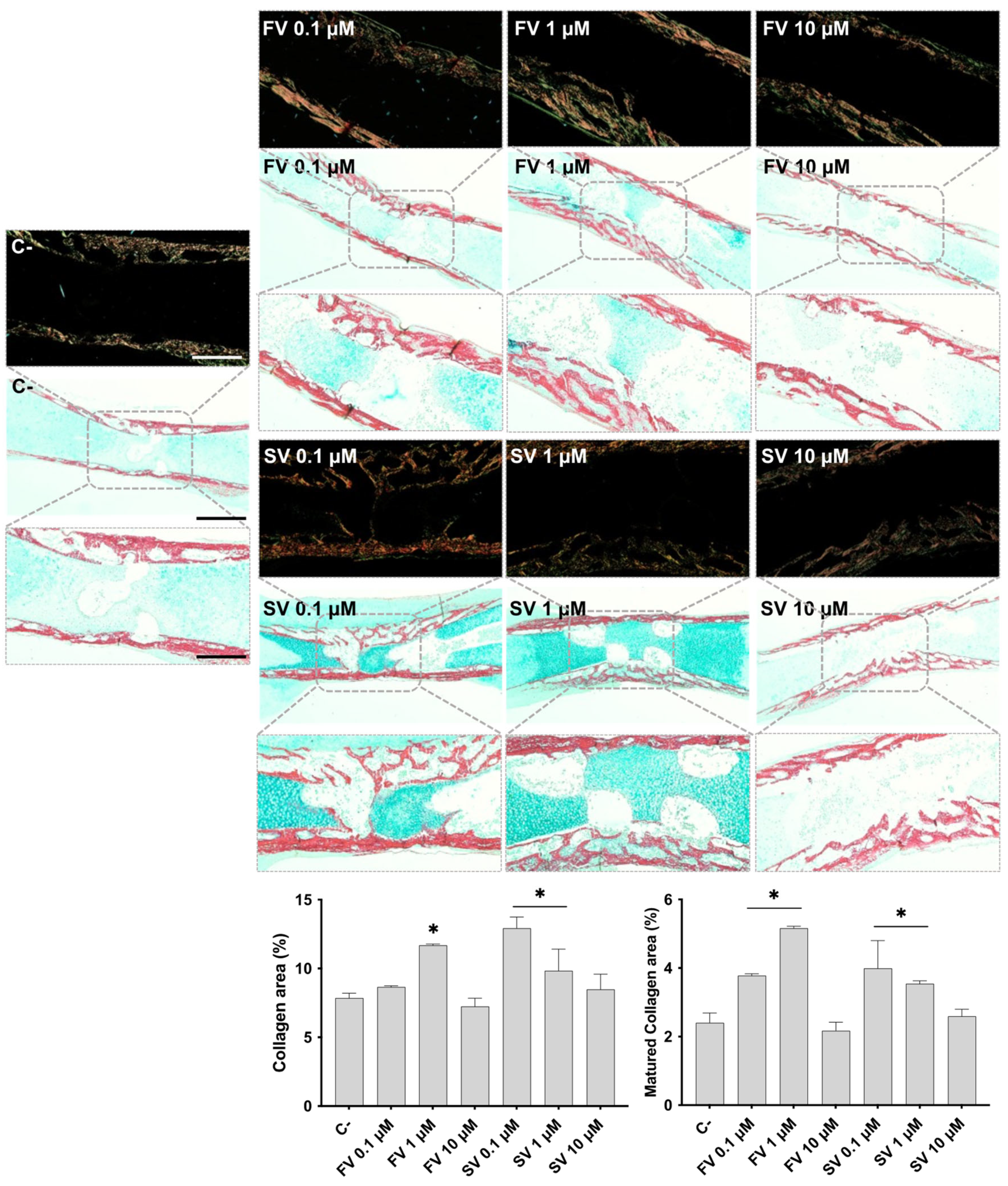
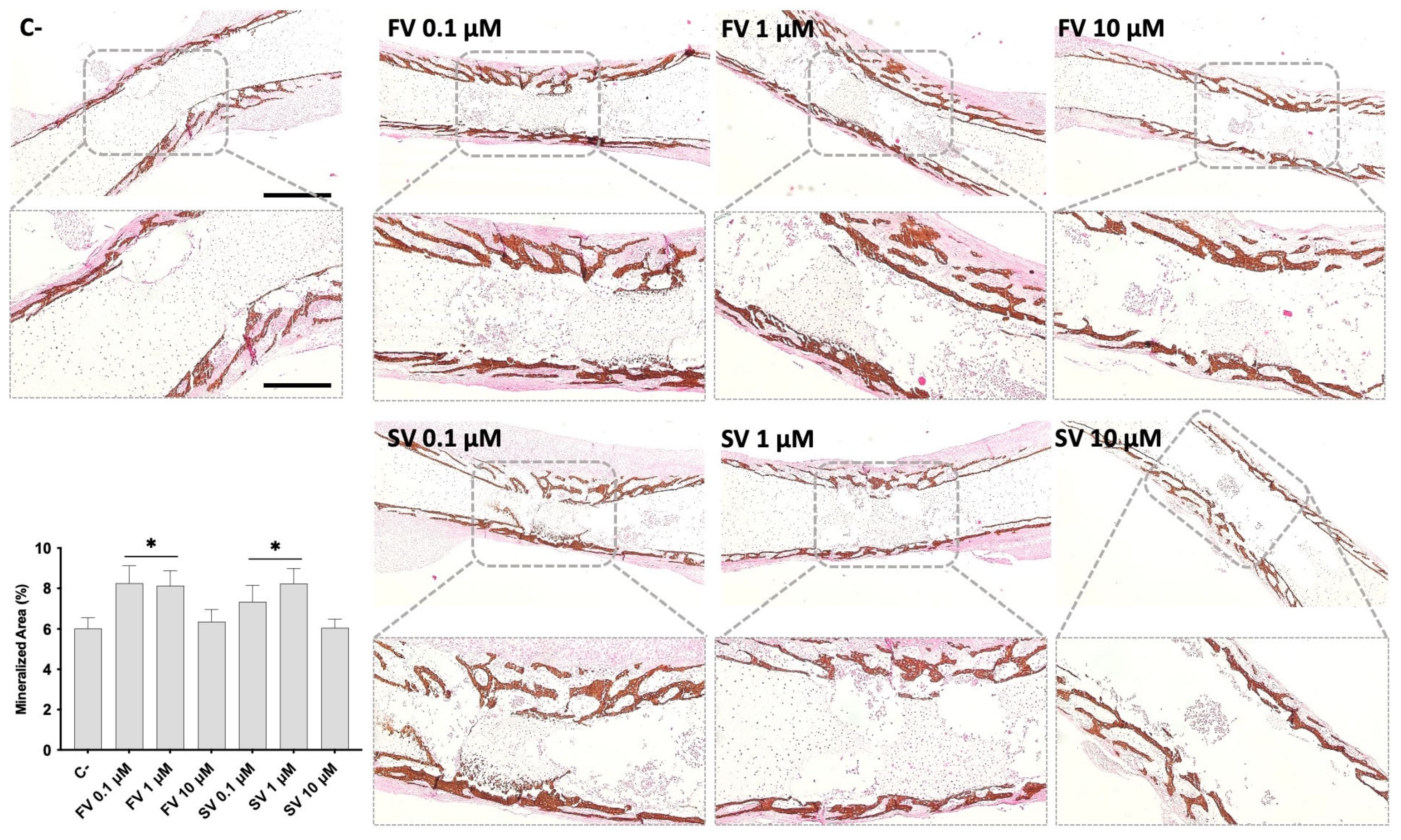
2.2. Histological Evaluation
2.3. Gene Expression Analysis
3. Discussion
4. Material and Methods
4.1. Organotypic Culture of Embryonic Chick Femora with Statins
4.2. Microtomographic Analysis
4.3. Histological Preparation and Histochemical Staining
4.4. Gene Expression Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Granat, M.M.; Eifler-Zydel, J.; Kolmas, J. Statins-Their Role in Bone Tissue Metabolism and Local Applications with Different Carriers. Int. J. Mol. Sci. 2024, 25, 2378. [Google Scholar] [CrossRef]
- Mundy, G.; Garrett, R.; Harris, S.; Chan, J.; Chen, D.; Rossini, G.; Boyce, B.; Zhao, M.; Gutierrez, G. Stimulation of bone formation in vitro and in rodents by statins. Science 1999, 286, 1946–1949. [Google Scholar] [CrossRef]
- Climent, E.; Benaiges, D.; Pedro-Botet, J. Hydrophilic or Lipophilic Statins? Front. Cardiovasc. Med. 2021, 8, 687585. [Google Scholar] [CrossRef]
- Salami, J.A.; Warraich, H.; Valero-Elizondo, J.; Spatz, E.S.; Desai, N.R.; Rana, J.S.; Virani, S.S.; Blankstein, R.; Khera, A.; Blaha, M.J.; et al. National Trends in Statin Use and Expenditures in the US Adult Population From 2002 to 2013: Insights From the Medical Expenditure Panel Survey. JAMA Cardiol. 2017, 2, 56–65. [Google Scholar] [CrossRef]
- Matyori, A.; Brown, C.P.; Ali, A.; Sherbeny, F. Statins utilization trends and expenditures in the U.S. before and after the implementation of the 2013 ACC/AHA guidelines. Saudi Pharm. J. 2023, 31, 795–800. [Google Scholar] [CrossRef]
- Chen, P.Y.; Sun, J.S.; Tsuang, Y.H.; Chen, M.H.; Weng, P.W.; Lin, F.H. Simvastatin promotes osteoblast viability and differentiation via Ras/Smad/Erk/BMP-2 signaling pathway. Nutr. Res. 2010, 30, 191–199. [Google Scholar] [CrossRef]
- Vukelic, S.; Stojadinovic, O.; Pastar, I.; Vouthounis, C.; Krzyzanowska, A.; Das, S.; Samuels, H.H.; Tomic-Canic, M. Farnesyl pyrophosphate inhibits epithelialization and wound healing through the glucocorticoid receptor. J. Biol. Chem. 2010, 285, 1980–1988. [Google Scholar] [CrossRef]
- Nes, W.D. Biosynthesis of cholesterol and other sterols. Chem. Rev. 2011, 111, 6423–6451. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, E.Y.; Lee, E.B.; Lee, Y.J.; Yoo, H.J.; Choi, J.; Song, Y.W. Atorvastatin inhibits osteoclastogenesis by decreasing the expression of RANKL in the synoviocytes of rheumatoid arthritis. Arthritis Res. Ther. 2012, 14, R187. [Google Scholar] [CrossRef]
- Sharif, P.S.; Abdollahi, M. A Systematic Review on the Relation between use of Statins and Osteoporosis. Int. J. Pharmacol. 2011, 7, 180–188. [Google Scholar] [CrossRef]
- Zhao, H.; Tang, Y.; Zhen, Y.; Qi, C.; Chen, S. The effect of statins on bone turnover biomarkers: A systematic review and meta-analysis of randomized controlled trials. Endocr. J. 2023, 70, 473–480. [Google Scholar] [CrossRef]
- Bone, H.G.; Kiel, D.P.; Lindsay, R.S.; Lewiecki, E.M.; Bolognese, M.A.; Leary, E.T.; Lowe, W.; McClung, M.R. Effects of atorvastatin on bone in postmenopausal women with dyslipidemia: A double-blind, placebo-controlled, dose-ranging trial. J. Clin. Endocrinol. Metab. 2007, 92, 4671–4677. [Google Scholar] [CrossRef]
- Di Spirito, F.; Schiavo, L.; Pilone, V.; Lanza, A.; Sbordone, L.; D’Ambrosio, F. Periodontal and Peri-Implant Diseases and Systemically Administered Statins: A Systematic Review. Dent. J. 2021, 9, 100. [Google Scholar] [CrossRef]
- Pruthi, G.; Mahajan, R.; Gupta, A.; Patil, A.N.; Paramasivam, V.; Kaundal, S. The Effects of Statins on Bone Formation Around Implants Placed in Animal Bones: A Systematic Review and Meta-Analysis. J. Maxillofac. Oral Surg. 2023, 22, 265–286. [Google Scholar] [CrossRef]
- Tahamtan, S.; Shirban, F.; Bagherniya, M.; Johnston, T.P.; Sahebkar, A. The effects of statins on dental and oral health: A review of preclinical and clinical studies. J. Transl. Med. 2020, 18, 155. [Google Scholar] [CrossRef]
- Lebeaux, D.; Chauhan, A.; Rendueles, O.; Beloin, C. From in vitro to in vivo Models of Bacterial Biofilm-Related Infections. Pathogens 2013, 2, 288–356. [Google Scholar] [CrossRef]
- Mariano, L.C.; Grenho, L.; Fernandes, M.H.; de Sousa Gomes, P. Integrative tissue, cellular and molecular responsiveness of an innovative ex vivo model of the Staphylococcus aureus-mediated bone infection. FASEB J. 2023, 37, e23166. [Google Scholar] [CrossRef]
- Groff, K.; Evans, S.J.; Doak, S.H.; Pfuhler, S.; Corvi, R.; Saunders, S.; Stoddart, G. In vitro and integrated in vivo strategies to reduce animal use in genotoxicity testing. Mutagenesis 2021, 36, 389–400. [Google Scholar] [CrossRef]
- Furtado, G.S.; Martin, V.; Araújo, R.; Gomes, P.S.; Lago, A.D.N. Osteoinductive activity of photobiomodulation in an organotypic bone model. Photodiagnosis Photodyn. Ther. 2024, 45, 103936. [Google Scholar] [CrossRef]
- Araújo, R.; Martin, V.; Ferreira, R.; Fernandes, M.H.; Gomes, P.S. A new ex vivo model of the bone tissue response to the hyperglycemic environment—The embryonic chicken femur organotypic culture in high glucose conditions. Bone 2022, 158, 116355. [Google Scholar] [CrossRef]
- Cramer, E.E.A.; Ito, K.; Hofmann, S. Ex vivo Bone Models and Their Potential in Preclinical Evaluation. Curr. Osteoporos. Rep. 2021, 19, 75–87. [Google Scholar] [CrossRef]
- Smith, E.L.; Kanczler, J.M.; Gothard, D.; Roberts, C.A.; Wells, J.A.; White, L.J.; Qutachi, Q.; Sawkins, M.J.; Peto, H.; Rashidi, H.; et al. Evaluation of skeletal tissue repair, part 2: Enhancement of skeletal tissue repair through dual-growth-factor-releasing hydrogels within an ex vivo chick femur defect model. Acta Biomater. 2014, 10, 4197–4205. [Google Scholar] [CrossRef]
- Smith, E.L.; Kanczler, J.M.; Oreffo, R.O.C. A new take on an old story: Chick limb organ culture for skeletal niche development and regenerative medicine evaluation. Eur. Cells Mater. 2013, 26, 91–106. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Hayslip, J.; Leggas, M. Pharmacokinetics of high-dose simvastatin in refractory and relapsed chronic lymphocytic leukemia patients. Cancer Chemother. Pharmacol. 2013, 72, 1369–1374. [Google Scholar] [CrossRef]
- Sabandal, M.M.I.; Schäfer, E.; Imper, J.; Jung, S.; Kleinheinz, J.; Sielker, S. Simvastatin induces in vitro mineralization effects of primary human odontoblast-like cells. Materials 2020, 13, 4679. [Google Scholar] [CrossRef]
- Zhang, M.; Bian, Y.Q.; Tao, H.M.; Yang, X.F.; Mu, W.D. Simvastatin induces osteogenic differentiation of MSCs via Wnt/β-catenin pathway to promote fracture healing. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2896–2905. [Google Scholar]
- Baek, K.H.; Lee, W.Y.; Oh, K.W.; Tae, H.J.; Lee, J.M.; Lee, E.J.; Han, J.H.; Kang, M.I.; Cha, B.Y.; Lee, K.W.; et al. The effect of simvastatin on the proliferation and differentiation of human bone marrow stromal cells. J. Korean Med. Sci. 2005, 20, 438–444. [Google Scholar] [CrossRef]
- Jia, W.; Zhao, Y.; Yang, J.; Wang, W.; Wang, X.; Ling, L.; Ge, L. Simvastatin Promotes Dental Pulp Stem Cell-induced Coronal Pulp Regeneration in Pulpotomized Teeth. J. Endod. 2016, 42, 1049–1054. [Google Scholar] [CrossRef]
- Gorabi, A.M.; Kiaie, N.; Pirro, M.; Bianconi, V.; Jamialahmadi, T.; Sahebkar, A. Effects of statins on the biological features of mesenchymal stem cells and therapeutic implications. Heart Fail. Rev. 2021, 26, 1259–1272. [Google Scholar] [CrossRef]
- Wood, W.G.; Igbavboa, U.; Muller, W.E.; Eckert, G.P. Statins, Bcl-2, and apoptosis: Cell death or cell protection? Mol. Neurobiol. 2013, 48, 308–314. [Google Scholar] [CrossRef]
- Malik, S.; Sharma, A.K.; Bharti, S.; Nepal, S.; Bhatia, J.; Nag, T.C.; Narang, R.; Arya, D.S. In vivo cardioprotection by pitavastatin from ischemic-reperfusion injury through suppression of IKK/NF-kappaB and upregulation of pAkt-e-NOS. J. Cardiovasc. Pharmacol. 2011, 58, 199–206. [Google Scholar] [CrossRef]
- Qin, W.; Lu, Y.; Zhan, C.; Shen, T.; Dou, L.; Man, Y.; Wang, S.; Xiao, C.; Bian, Y.; Li, J. Simvastatin suppresses apoptosis in vulnerable atherosclerotic plaques through regulating the expression of p(53), Bcl-2 and Bcl-xL. Cardiovasc. Drugs Ther. 2012, 26, 23–30. [Google Scholar] [CrossRef]
- Moriishi, T.; Maruyama, Z.; Fukuyama, R.; Ito, M.; Miyazaki, T.; Kitaura, H.; Ohnishi, H.; Furuichi, T.; Kawai, Y.; Masuyama, R.; et al. Overexpression of Bcl2 in osteoblasts inhibits osteoblast differentiation and induces osteocyte apoptosis. PLoS ONE 2011, 6, e27487. [Google Scholar] [CrossRef]
- Lai, M.; Yan, X.; Jin, Z. The response of bone cells to titanium surfaces modified by simvastatin-loaded multilayered films. J. Biomater. Sci. Polym. Ed. 2018, 29, 1895–1908. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.; Zhang, Z. Porous Chitosan/Nano-Hydroxyapatite Composite Scaffolds Incorporating Simvastatin-Loaded PLGA Microspheres for Bone Repair. Cells Tissues Organs 2018, 205, 20–31. [Google Scholar] [CrossRef]
- Zhang, H.X.; Xiao, G.Y.; Wang, X.; Dong, Z.G.; Ma, Z.Y.; Li, L.; Li, Y.H.; Pan, X.; Nie, L. Biocompatibility and osteogenesis of calcium phosphate composite scaffolds containing simvastatin-loaded PLGA microspheres for bone tissue engineering. J. Biomed. Mater. Res. A 2015, 103, 3250–3258. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, R.; Li, Y.; He, G.; Zhang, D.; Zhang, F. Simvastatin augments the efficacy of therapeutic angiogenesis induced by bone marrow-derived mesenchymal stem cells in a murine model of hindlimb ischemia. Mol. Biol. Rep. 2012, 39, 285–293. [Google Scholar] [CrossRef]
- Pirzad Jahromi, G.; Shabanzadeh, A.P.; Mokhtari Hashtjini, M.; Sadr, S.S.; Rasouli Vani, J.; Raouf Sarshoori, J.; Charish, J. Bone marrow-derived mesenchymal stem cell and simvastatin treatment leads to improved functional recovery and modified c-Fos expression levels in the brain following ischemic stroke. Iran. J. Basic Med. Sci. 2018, 21, 1004–1012. [Google Scholar]
- Cai, J.; Yu, X.; Zhang, B.; Zhang, H.; Fang, Y.; Liu, S.; Liu, T.; Ding, X. Atorvastatin improves survival of implanted stem cells in a rat model of renal ischemia-reperfusion injury. Am. J. Nephrol. 2014, 39, 466–475. [Google Scholar] [CrossRef]
- Dai, B.; Li, X.; Xu, J.; Zhu, Y.; Huang, L.; Tong, W.; Yao, H.; Chow, D.H.-K.; Qin, L. Synergistic effects of magnesium ions and simvastatin on attenuation of high-fat diet-induced bone loss. Bioact. Mater. 2021, 6, 2511–2522. [Google Scholar] [CrossRef]
- Feng, C.; Xiao, L.; Yu, J.C.; Li, D.Y.; Tang, T.Y.; Liao, W.; Wang, Z.R.; Lu, A.Q. Simvastatin promotes osteogenic differentiation of mesenchymal stem cells in rat model of osteoporosis through BMP-2/Smads signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 434–443. [Google Scholar]
- Long, F. Building strong bones: Molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell. Biol. 2011, 13, 27–38. [Google Scholar] [CrossRef]
- Komori, T. Whole Aspect of Runx2 Functions in Skeletal Development. Int. J. Mol. Sci. 2022, 23, 5776. [Google Scholar] [CrossRef]
- Li, X.; Cui, Q.; Kao, C.; Wang, G.J.; Balian, G. Lovastatin inhibits adipogenic and stimulates osteogenic differentiation by suppressing PPARgamma2 and increasing Cbfa1/Runx2 expression in bone marrow mesenchymal cell cultures. Bone 2003, 33, 652–659. [Google Scholar] [CrossRef]
- Qiao, L.J.; Kang, K.L.; Heo, J.S. Simvastatin promotes osteogenic differentiation of mouse embryonic stem cells via canonical Wnt/beta-catenin signaling. Mol. Cells 2011, 32, 437–444. [Google Scholar] [CrossRef]
- Alam, S.; Ueki, K.; Nakagawa, K.; Marukawa, K.; Hashiba, Y.; Yamamoto, E.; Sakulsak, N.; Iseki, S. Statin-induced bone morphogenetic protein (BMP) 2 expression during bone regeneration: An immunohistochemical study. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2009, 107, 22–29. [Google Scholar] [CrossRef]
- Maciel-Oliveira, N.; Bradaschia-Correa, V.; Arana-Chavez, V.E. Early alveolar bone regeneration in rats after topical administration of simvastatin. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2011, 112, 170–179. [Google Scholar] [CrossRef]
- Ruiz-Gaspa, S.; Nogues, X.; Enjuanes, A.; Monllau, J.C.; Blanch, J.; Carreras, R.; Mellibovsky, L.; Grinberg, D.; Balcells, S.; Díez-Perez, A.; et al. Simvastatin and atorvastatin enhance gene expression of collagen type 1 and osteocalcin in primary human osteoblasts and MG-63 cultures. J. Cell. Biochem. 2007, 101, 1430–1438. [Google Scholar] [CrossRef]
- Mansour, G.; Al Ashwah, A.; Koura, A. Evaluation of simvastatin grafting around immediate dental implants in dogs. Implant. Dent. 2014, 23, 195–199. [Google Scholar] [CrossRef]
- Selvaraj, V.; Sekaran, S.; Dhanasekaran, A.; Warrier, S. Type 1 collagen: Synthesis, structure and key functions in bone mineralization. Differentiation 2024, 136, 100757. [Google Scholar] [CrossRef]
- Dolkart, O.; Pritsch, T.; Sharfman, Z.T.; Somjen, D.; Salai, M.; Maman, E.; Steinberg, E.L. The Effects of Lipophilic and Hydrophilic Statins on Bone Tissue Mineralization in Saos2 Human Bone Cell Line?In vitro Comparative Study. Pharm. Anal. Acta 2015, 6, 1000363. [Google Scholar]
- Moriyama, Y.; Ayukawa, Y.; Ogino, Y.; Atsuta, I.; Koyano, K. Topical application of statin affects bone healing around implants. Clin. Oral Implant. Res. 2008, 19, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, Y.; Ayukawa, Y.; Ogino, Y.; Atsuta, I.; Todo, M.; Takao, Y.; Koyano, K. Local application of fluvastatin improves peri-implant bone quantity and mechanical properties: A rodent study. Acta Biomater. 2010, 6, 1610–1618. [Google Scholar] [CrossRef] [PubMed]
- Gruber, H.E.; Ingram, J.; Hanley, E.N., Jr. An improved staining method for intervertebral disc tissue. Biotech. Histochem. 2002, 77, 81–83. [Google Scholar] [CrossRef]

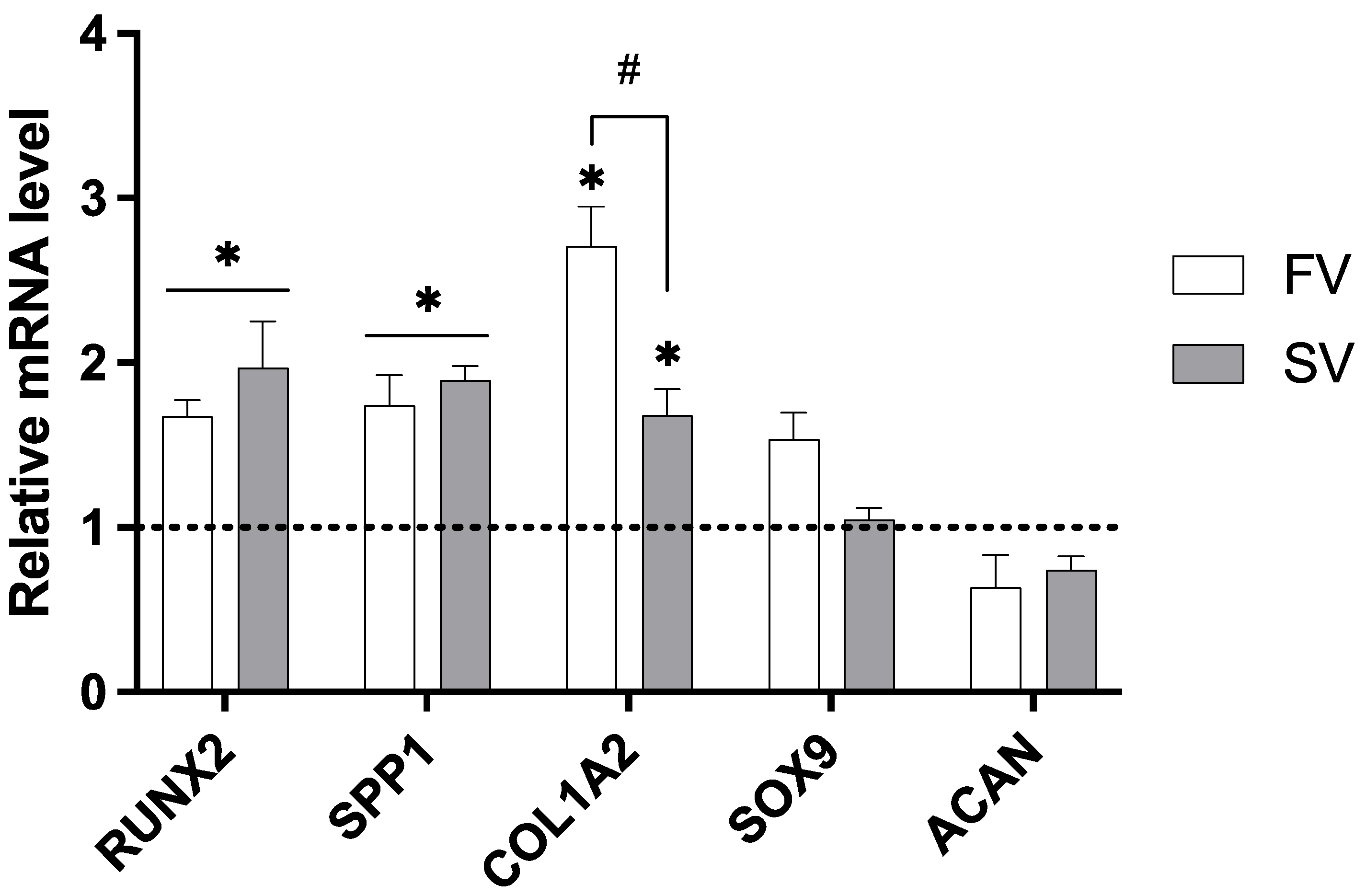

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | |||
|---|---|---|---|
| GAPDH | qGgaCED0029996 | COL1A2 | qGgaCED0025365 |
| RUNX2 | qGgaCID0019198 | ACAN | qGgaCID0030890 |
| SPP1 | qGgaCED0023869 | SOX9 | qGgaCED0029640 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poskevicius, L.; Martin, V.; Costa, G.; Juodžbalys, G.; Sousa Gomes, P. Osteogenic Potential of Simvastatin and Fluvastatin in an Organotypic Bone Model. Pharmaceuticals 2025, 18, 939. https://doi.org/10.3390/ph18070939
Poskevicius L, Martin V, Costa G, Juodžbalys G, Sousa Gomes P. Osteogenic Potential of Simvastatin and Fluvastatin in an Organotypic Bone Model. Pharmaceuticals. 2025; 18(7):939. https://doi.org/10.3390/ph18070939
Chicago/Turabian StylePoskevicius, Lukas, Victor Martin, Guilherme Costa, Gintaras Juodžbalys, and Pedro Sousa Gomes. 2025. "Osteogenic Potential of Simvastatin and Fluvastatin in an Organotypic Bone Model" Pharmaceuticals 18, no. 7: 939. https://doi.org/10.3390/ph18070939
APA StylePoskevicius, L., Martin, V., Costa, G., Juodžbalys, G., & Sousa Gomes, P. (2025). Osteogenic Potential of Simvastatin and Fluvastatin in an Organotypic Bone Model. Pharmaceuticals, 18(7), 939. https://doi.org/10.3390/ph18070939