
Integrated Omics Reveal Dendrobium nobile Lindl.’s Anti-Diabetic Mechanisms via Arginine/Proline and Glycerophospholipid Pathways

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
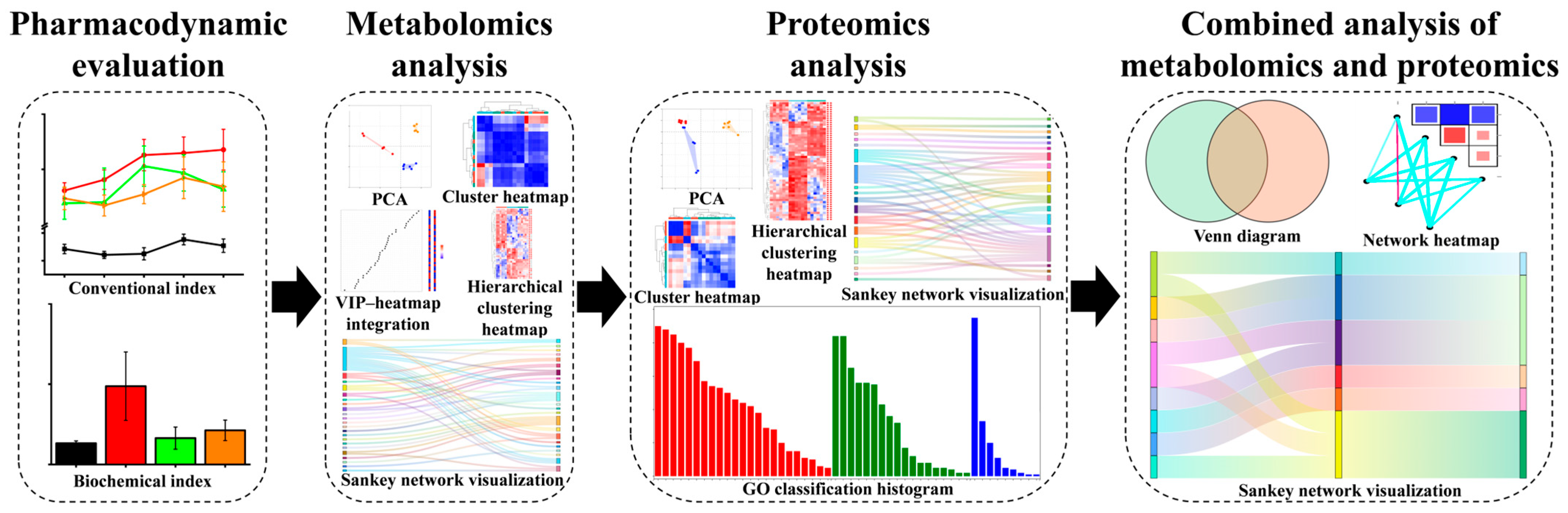
1. Introduction
2. Results
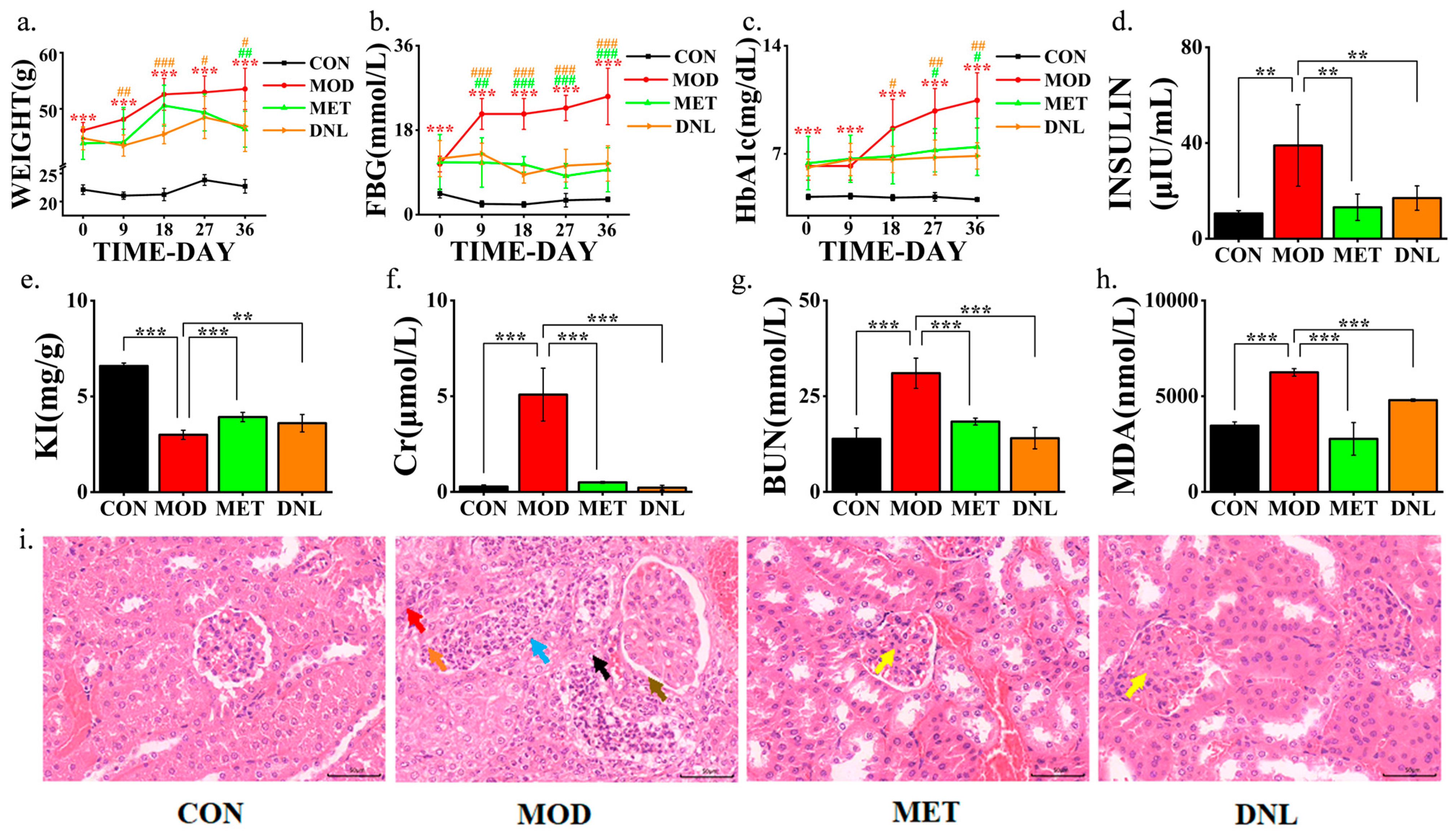
2.1. DNL Significantly Attenuated the T2DM Disease Phenotype in db/db Mice
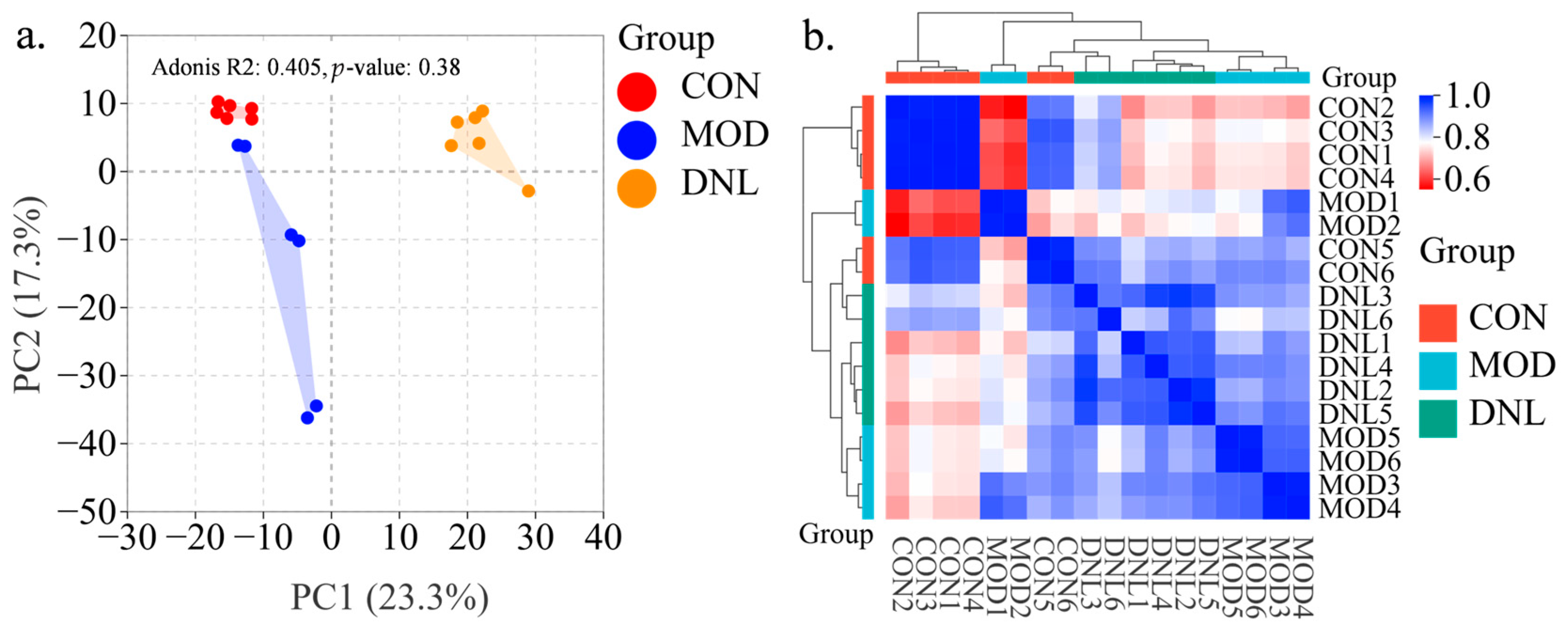
2.2. Metabolic Effects of DNL on Serum Profiles of db/db Mice
2.3. DNL Exhibited Effects on db/db Mice with T2DM Related Various Metabolites
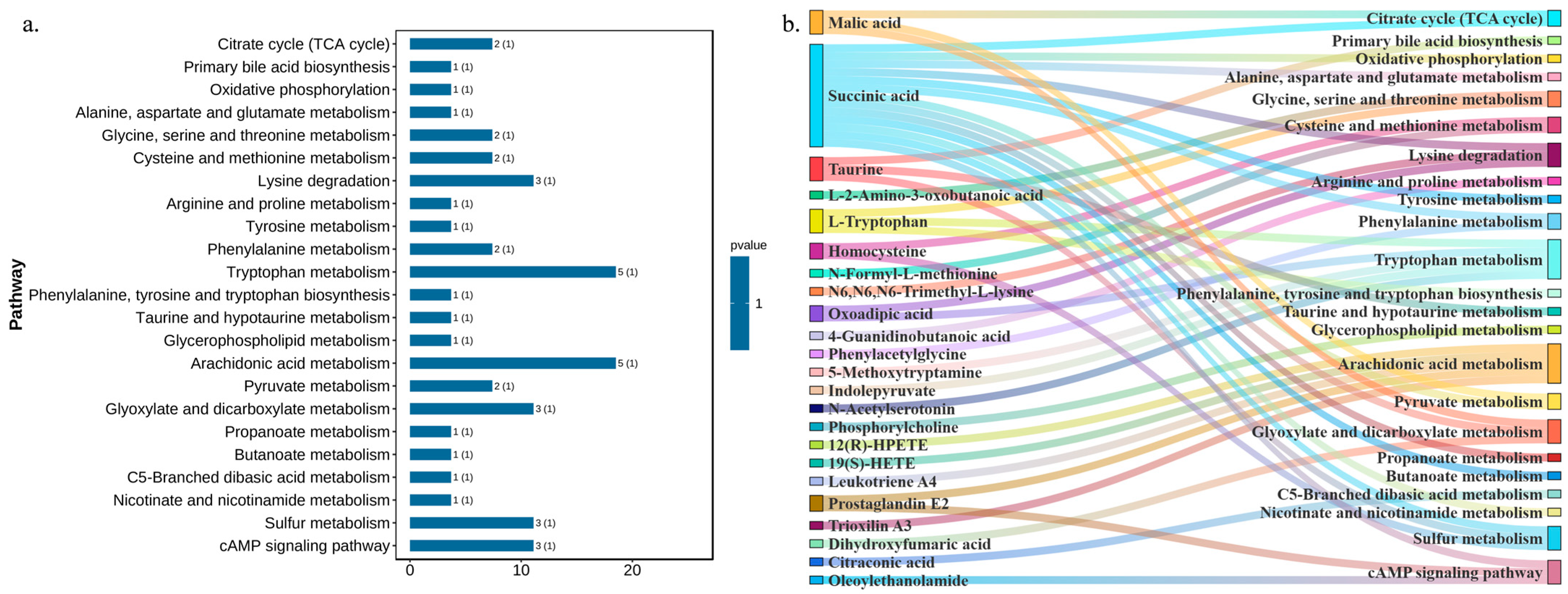
2.4. DNL Exerted Influences on db/db Mice with T2DM-Related Metabolic Pathways
2.5. DNL Showed Effects on Serum Proteinic Profiles in db/db Mice with T2DM
2.6. Proteins Changes of DNL on db/db Mice with T2DM
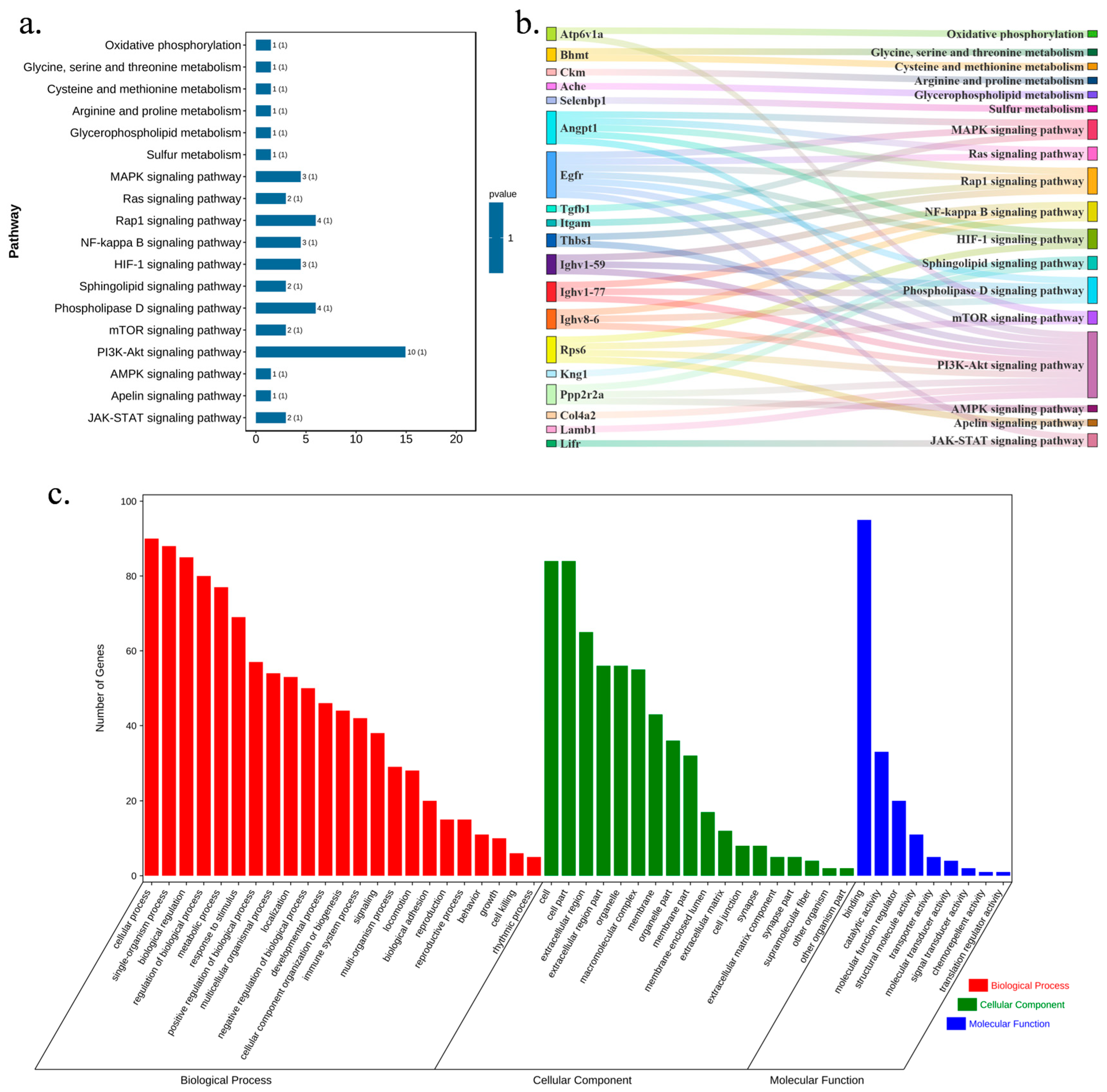
2.7. DNL Exerted Influences on db/db Mice with T2DM-Related Pathways
2.8. Metabolomics with Proteomics Revealed Mechanism of DNL Intervention in db/db Mice
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. DNL Preparation
4.3. General Status Observation and Monitoring of Indicators
4.4. Histopathological Examination
4.5. Serum Biochemical Analysis
4.6. Sample Preparation for Metabolomics Analysis
4.7. Liquid Chromatographic and Mass Spectrometry Conditions
4.8. Metabolomics Data Processing
4.9. Proteomics Analysis of Sample Preparation
4.10. Chromatographic and Mass Spectrometry Conditions
4.11. Proteomics Data Processing
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Diabetes Association Professional Practice Committee. 2. Diagnosis and Classification of Diabetes: Standards of Care in Diabetes-2024. Diabetes Care 2024, 47 (Suppl. S1), S20–S42. [Google Scholar] [CrossRef] [PubMed]
- Mulder, H. Transcribing β-cell mitochondria in health and disease. Mol. Metab. 2017, 6, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Festa, A.; D’Agostino, R., Jr.; Howard, G.; Mykkänen, L.; Tracy, R.P.; Haffner, S.M. Chronic subclinical inflammation as part of the insulin resistance syndrome: The Insulin Resistance Atherosclerosis Study (IRAS). Circulation 2000, 102, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Chen, Y.; Li, W.; Lu, Y. Anti-aging properties of Dendrobium nobile Lindl.: From molecular mechanisms to potential treatments. J. Ethnopharmacol. 2020, 257, 112839. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.Z.; Dan, Y.; Liu, Y.Z. Pharmacopoeia of the People’s Republic of China (2010 Edition): A milestone in development of China’s healthcare. Chin. Herb. Med. 2010, 2, 157–160. [Google Scholar]
- Fan, C.; Sun, X.; Wang, X.; Yu, H. Therapeutic potential of the chemical composition of Dendrobium nobile Lindl. Front Pharmacol. Front. Pharmacol. 2023, 14, 1163830. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, Y.; Yang, L.; Wang, B.; Fan, Y.; Ruan, B.; Zhang, X.; Dai, H.; Mei, W.; Jie, W.; et al. Integrated analysis of Dendrobium nobile extract Dendrobin A against pancreatic ductal adenocarcinoma based on network pharmacology, bioinformatics, and validation experiments. Front. Pharmacol. 2023, 14, 1079539. [Google Scholar] [CrossRef]
- Lei, X.; Huo, P.; Xie, Y.J.; Wang, Y.; Liu, G.; Tu, H.; Shi, Q.; Mo, Z.C.; Zhang, S. Dendrobium nobile Lindl polysaccharides improve testicular spermatogenic function in streptozotocin-induced diabetic rats. Mol. Reprod. Dev. 2022, 89, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, X.; Wang, H.; Fang, J.; Zhang, Z.; Zhu, X.; Qi, Z.; Guo, Y.; Liu, R.; Li, D. Dendrobium officinale polysaccharide prevents neuronal apoptosis via TET2-dependent DNA demethylation in high-fat diet-induced diabetic mice. Int. J. Biol. Macromol. 2023, 233, 123288. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yang, D.; Xu, Y.; Wu, D.; Tan, D.; Qin, L.; Wu, X.; Lu, Y.; He, Y. Hypoglycemic Effects and Quality Marker Screening of Dendrobium nobile Lindl. at Different Growth Years. Molecules 2024, 29, 699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, M.; Zhang, C.; Liu, Z.; Zhou, S. Clinical study of Dendrobium nobile Lindl intervention on patients with metabolic syndrome. Medicine 2021, 100, e24574. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Cai, Y.; Yao, H.; Lin, C.; Xie, Y.; Tang, S.; Zhang, A. Small molecule metabolites: Discovery of biomarkers and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Guo, S.; Cai, Y.; Yang, Q.; Wang, Y.; Yu, X.; Sun, W.; Qiu, S.; Li, X.; Guo, Y.; et al. Decoding active compounds and molecular targets of herbal medicine by high-throughput metabolomics technology: A systematic review. Bioorg. Chem. 2024, 144, 107090. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Wang, Y.; Guo, S.; Wang, Z.; Chen, H.; Wang, X.; Xie, D.; Cai, Y.; Wang, S.; Hu, Z.; et al. Proteomics exploration of metformin hydrochloride for diabetic kidney disease treatment via the butanoate metabolism pathway. J. Pharm. Biomed. Anal. 2025, 254, 116584. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Wang, Z.; Guo, S.; Xie, D.; Cai, Y.; Wang, X.; Lin, C.; Tang, S.; Xie, Y.; Zhang, A. Single cell sequencing and spatial multiomics of diabetic kidney segmentation insights zonation-specific therapeutic metabolic pathways. Cell Insight 2025, 4, 100252. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Xie, D.; Guo, S.; Wang, Z.; Wang, X.; Cai, Y.; Lin, C.; Yao, H.; Guan, Y.; Zhao, Q. Spatially segregated multiomics decodes metformin-mediated functional-specific metabolic characteristics in diabetic kidney disease. Life Metab. 2025, loaf019. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.; Wu, N.; Lv, C.; Wang, Y. P4HB regulates the TGFβ/SMAD3 signaling pathway through PRMT1 to participate in high glucose-induced epithelial-mesenchymal transition and fibrosis of renal tubular epithelial cells. BMC Nephrol. 2024, 25, 297. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, A.; Yao, L.; Xu, Z.; Toque, H.A.; Chen, J.; Atawia, R.T.; Fouda, A.Y.; Bagi, Z.; Lucas, R.; Caldwell, R.B.; et al. Obesity-induced vascular dysfunction and arterial stiffening requires endothelial cell arginase 1. Cardiovasc. Res. 2017, 113, 1664–1676. [Google Scholar] [CrossRef]
- Hong, H.; Fill, T.; Leadlay, P.F. A common origin for guanidinobutanoate starter units in antifungal natural products. Angew. Chem Int. Ed. Engl. 2013, 52, 13096–13099. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wei, D.; Wang, Q.; Guo, J.; Zhu, Y.; Shang, E.; Duan, J.A. Luteoloside mitigates premature age-related macular degeneration by suppressing p53-p21-Rb1 axis: Insights from transcriptomic analysis, serum metabolomics and gut microbiota analysis. J. Pharm. Biomed. Anal. 2024, 248, 116296. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, F.; Murakami, C.; Sakai, H.; Satoh, M.; Sakane, F. Creatine kinase muscle type specifically interacts with saturated fatty acid- and/or monounsaturated fatty acid-containing phosphatidic acids. Biochem. Biophys. Res. Commun. 2019, 513, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Hatch, G.M.; Wang, Y.; Yu, F.; Wang, M. The relationship between phospholipids and insulin resistance: From clinical to experimental studies. J. Cell. Mol. Med. 2019, 23, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Wuttke, A. Lipid signalling dynamics at the β-cell plasma membrane. Basic Clin. Pharmacol. Toxicol. 2015, 116, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Frostegård, J. Antibodies against Phosphorylcholine-Implications for Chronic Inflammatory Diseases. Metabolites 2023, 13, 720. [Google Scholar]
- He, M.; Li, Z.; Tung, V.S.K.; Pan, M.; Han, X.; Evgrafov, O.; Jiang, X.C. Inhibiting Phosphatidylcholine Remodeling in Adipose Tissue Increases Insulin Sensitivity. Diabetes 2023, 72, 1547–1559. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.F.; Aristizabal-Henao, J.J.; Marvyn, P.M.; M’Hiri, I.; Wiens, M.A.; Hoang, M.; Sebastian, M.; Nachbar, R.; St-Pierre, P.; Diaguarachchige De Silva, K.; et al. Renal tubule-specific Atgl deletion links kidney lipid metabolism to glucagon-like peptide 1 and insulin secretion independent of renal inflammation or lipotoxicity. Mol. Metab. 2024, 81, 101887. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.N.; Morton, N.M. Cysteine and hydrogen sulphide in the regulation of metabolism: Insights from genetics and pharmacology. J. Pathol. 2016, 238, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Prajjwal, P.; Asharaf, S.; Makhanasa, D.; Yamparala, A.; Tariq, H.; Aleti, S.; Gadam, S.; Vora, N. Association of Alzheimer’s dementia with oral bacteria, vitamin B12, folate, homocysteine levels, and insulin resistance along with its pathophysiology, genetics, imaging, and biomarkers. Dis. Mon. 2023, 69, 101546. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qu, Y.Y.; Liu, L.; Qiao, Y.N.; Geng, H.R.; Lin, Y.; Xu, W.; Cao, J.; Zhao, J.Y. Homocysteine inhibits pro-insulin receptor cleavage and causes insulin resistance via protein cysteine-homocysteinylation. Cell Rep. 2021, 37, 109821. [Google Scholar] [CrossRef] [PubMed]
- Sastre, J.; Pérez, S.; Sabater, L.; Rius-Pérez, S. Redox signaling in the pancreas in health and disease. Physiol Rev. 2025, 105, 593–650. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Li, Y.; Lv, Y.; Sun, L.; Zhang, S.; Li, Y.; Wang, Y.; Liu, G.; Xu, M.J.; Wang, X.; et al. Intermedin restores hyperhomocysteinemia-induced macrophage polarization and improves insulin resistance in mice. J. Biol. Chem. 2016, 291, 12336–12345. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.C.; Guo, Y.; Xu, F.; Saxena, D.; Li, X. A novel SUCNR1 inhibitor alleviates dysbiosis through inhibition of host responses without direct interaction with host microbiota. Mol. Oral. Microbiol. 2024, 39, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Rawal, S.; Randhawa, V.; Rizvi, S.H.M.; Sachan, M.; Wara, A.K.; Pérez-Cremades, D.; Weisbrod, R.M.; Hamburg, N.M.; Feinberg, M.W. miR-369-3p ameliorates diabetes-associated atherosclerosis by regulating macrophage succinate-GPR91 signalling. Cardiovasc. Res. 2024, 120, 1693–1712. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; An, N.; Zhang, Q.; Liu, Y.; Gu, P.; Zhao, J.; Pan, W.; Pu, Q.; Wen, Z. SLC34A2 inhibits tumorigenesis and progression via upregulating LRRK2/TTF-1/SELENBP1 axis in lung adenocarcinoma. Cancer Gene Ther. 2025. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, K.; Xia, J.; Qian, D.; Guo, J.; Zhong, L.; Tang, D.; Chen, X.; Peng, W.; Chen, Y.; et al. Natural exosomes-like nanoparticles in mung bean sprouts possesses anti-diabetic effects via activation of PI3K/Akt/GLUT4/GSK-3β signaling pathway. J. Nanobiotechnol. 2023, 21, 349. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, B.; Du, R.; Zhong, M.; Zhang, C.; Jin, X.; Cui, X.; Zhou, Y.; Kang, Q.; Xu, H.; et al. An herbal formulation “Shugan Xiaozhi decoction” ameliorates methionine/choline deficiency-induced nonalcoholic steatohepatitis through regulating inflammation and apoptosis-related pathways. J. Ethnopharmacol. 2024, 329, 118127. [Google Scholar] [CrossRef] [PubMed]
- National Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China: Volume I; China Pharmaceutical Science and Technology Press: Beijing, China, 2020; pp. 94–97. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Wang, X.; Guo, S.; Cai, Y.; Xie, D.; Wang, Y.; Zhang, A.; Dai, J.; Qiu, S. Integrated Omics Reveal Dendrobium nobile Lindl.’s Anti-Diabetic Mechanisms via Arginine/Proline and Glycerophospholipid Pathways. Pharmaceuticals 2025, 18, 1061. https://doi.org/10.3390/ph18071061
Wang Z, Wang X, Guo S, Cai Y, Xie D, Wang Y, Zhang A, Dai J, Qiu S. Integrated Omics Reveal Dendrobium nobile Lindl.’s Anti-Diabetic Mechanisms via Arginine/Proline and Glycerophospholipid Pathways. Pharmaceuticals. 2025; 18(7):1061. https://doi.org/10.3390/ph18071061
Chicago/Turabian StyleWang, Zhibo, Xian Wang, Sifan Guo, Ying Cai, Dandan Xie, Yujuan Wang, Aihua Zhang, Jun Dai, and Shi Qiu. 2025. "Integrated Omics Reveal Dendrobium nobile Lindl.’s Anti-Diabetic Mechanisms via Arginine/Proline and Glycerophospholipid Pathways" Pharmaceuticals 18, no. 7: 1061. https://doi.org/10.3390/ph18071061
APA StyleWang, Z., Wang, X., Guo, S., Cai, Y., Xie, D., Wang, Y., Zhang, A., Dai, J., & Qiu, S. (2025). Integrated Omics Reveal Dendrobium nobile Lindl.’s Anti-Diabetic Mechanisms via Arginine/Proline and Glycerophospholipid Pathways. Pharmaceuticals, 18(7), 1061. https://doi.org/10.3390/ph18071061