
Effects of Subchronic Buspirone Treatment on Depressive Profile in Socially Isolated Rats: Implication of Early Life Experience on 5-HT1A Receptor-Related Depression
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
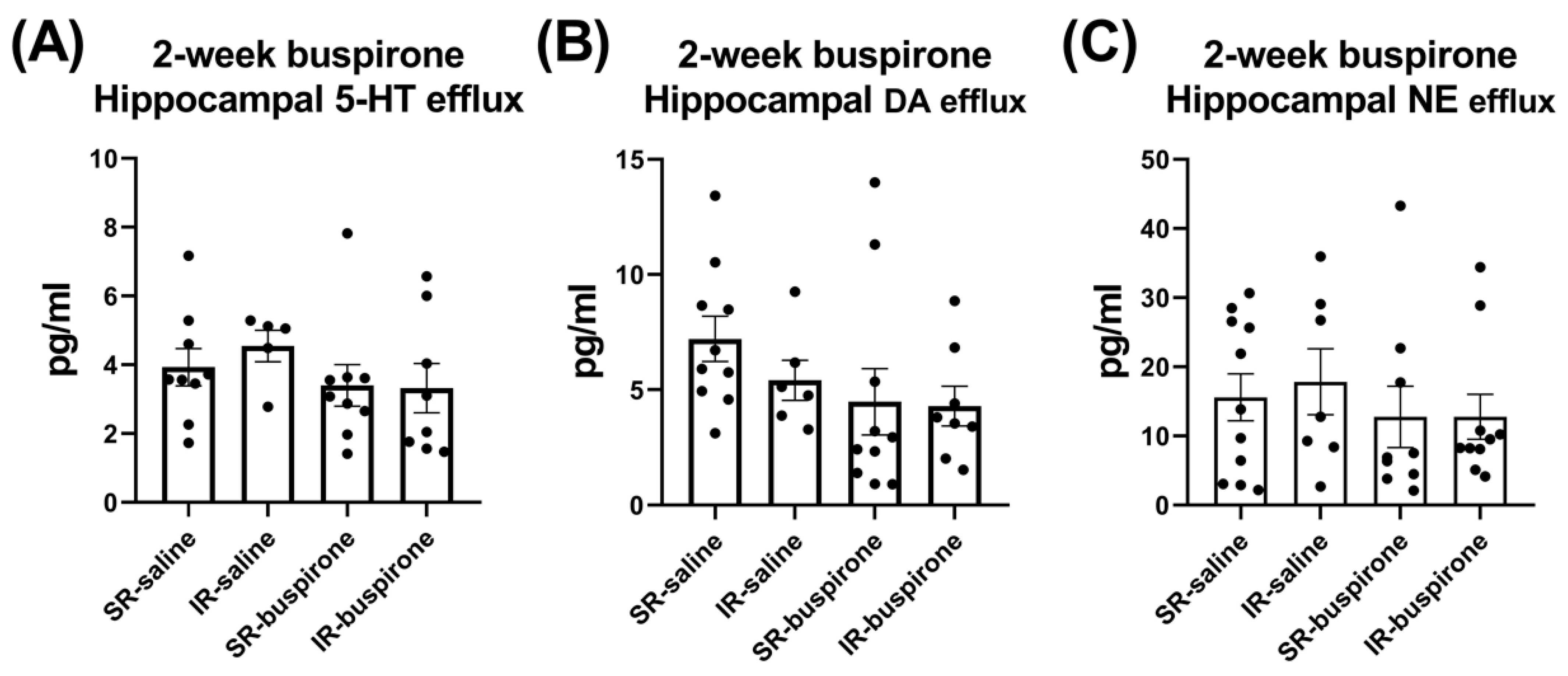
2. Results
3. Discussion
4. Material and Methods
4.1. Animals
4.2. IR Procedure
4.3. Drugs
4.4. Locomotor Activity
4.5. Forced Swim Test (FST)
4.6. Extracellular Levels of 5-HT, DA, and NE in the Hippocampus
4.7. Tissue Levels of 5-HT and 5-HIAA in Hippocampus, Amygdala, and Prefrontal Cortex
4.8. Data Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berman, R.M.; Narasimhan, M.; Charney, D.S. Treatment-refractory depression: Definitions and characteristics. Depress. Anxiety 1997, 5, 154–164. [Google Scholar] [CrossRef]
- Pilon, D.; Joshi, K.; Sheehan, J.J.; Zichlin, M.L.; Zuckerman, P.; Lefebvre, P.; Greenberg, P.E. Burden of treatment-resistant depression in Medicare: A retrospective claims database analysis. PLoS ONE 2019, 14, e0223255. [Google Scholar] [CrossRef] [PubMed]
- Santarelli, S.; Zimmermann, C.; Kalideris, G.; Lesuis, S.L.; Arloth, J.; Uribe, A.; Dournes, C.; Balsevich, G.; Hartmann, J.; Masana, M.; et al. An adverse early life environment can enhance stress resilience in adulthood. Psychoneuroendocrinology 2017, 78, 213–221. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.M.; Green, M.R. From the stressed adolescent to the anxious and depressed adult: Investigations in rodent models. Neuroscience 2013, 249, 242–257. [Google Scholar] [CrossRef] [PubMed]
- Aswar, U.; Shende, H.; Aswar, M. Buspirone, a 5-HT1A agonist attenuates social isolation-induced behavior deficits in rats: A comparative study with fluoxetine. Behav. Pharmacol. 2022, 33, 309–321. [Google Scholar] [CrossRef]
- Burke, A.R.; McCormick, C.M.; Pellis, S.M.; Lukkes, J.L. Impact of adolescent social experiences on behavior and neural circuits implicated in mental illnesses. Neurosci. Biobehav. Rev. 2017, 76, 280–300. [Google Scholar] [PubMed]
- Liu, Y.P.; Kao, Y.C.; Tung, C.S. Critical period exists in the effects of isolation rearing on sensorimotor gating function but not locomotor activity in rat. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Dimonte, S.; Sikora, V.; Bove, M.; Morgese, M.G.; Tucci, P.; Schiavone, S.; Trabace, L. Social isolation from early life induces anxiety-like behaviors in adult rats: Relation to neuroendocrine and neurochemical dysfunctions. Biomed. Pharmacother. 2023, 158, 31. [Google Scholar] [CrossRef] [PubMed]
- Carver, C.S.; Johnson, S.L.; Joormann, J.; Kim, Y.; Nam, J.Y. Serotonin transporter polymorphism interacts with childhood adversity to predict aspects of impulsivity. Psychol. Sci. 2011, 22, 589–595. [Google Scholar] [CrossRef]
- Phillips, B.U.; Robbins, T.W. Chapter 31—The role of central serotonin in impulsivity, compulsivity, and decision-making: Comparative studies in experimental animals and humans. In Handbook of Behavioral Neuroscience; Müller, C.P., Cunningham, K.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 31, pp. 531–548. [Google Scholar]
- Fletcher, P.J. Effects of combined or separate 5,7-dihydroxytryptamine lesions of the dorsal and median raphe nuclei on responding maintained by a DRL 20s schedule of food reinforcement. Brain Res. 1995, 675, 45–54. [Google Scholar] [CrossRef]
- Ago, Y.; Takuma, K.; Matsuda, T. Potential role of serotonin1A receptors in post-weaning social isolation-induced abnormal behaviors in rodents. J. Pharmacol. Sci. 2014, 125, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Sikich, L.; Hickok, J.M.; Todd, R.D. 5-HT1A receptors control neurite branching during development. Brain Res. Dev. Brain Res. 1990, 56, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Tung, C.S.; Lin, Y.W.; Lin, C.C.; Liu, Y.P. Time-Dependent Effects of Buspirone versus Desipramine on the 5-Choice Serial Reaction Time Task in Rats Reared in Social Isolation: Implication of Early Life Experience and Motoric Impulsivity. J. Integr. Neurosci. 2023, 22, 80. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.J.; Chen, Y.J.; Chung, J.Y.; Lin, C.C.; Liu, Y.P. Effects of 5HT1A Activation on Depression Profile Following 5-HT Depletion in Rats Lacking Social Attachment Since Weanling. Psychiatry Investig. 2018, 15, 1000–1006. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.M.; Ko, C.Y.; Lin, C.C.; Wan, Y.Y.; Chung, J.Y.; Liu, Y.P. Effects of 5HT1A Activation on Gating Profile Following 5HT Depletion in Rats Lacking Social Attachment Since Weanling. Psychiatry Investig. 2018, 15, 193–199. [Google Scholar] [CrossRef]
- Sprouse, J.S.; Aghajanian, G.K. Electrophysiological responses of serotoninergic dorsal raphe neurons to 5-HT1A and 5-HT1B agonists. Synapse 1987, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Celada, P.; Bortolozzi, A.; Artigas, F. Serotonin 5-HT1A receptors as targets for agents to treat psychiatric disorders: Rationale and current status of research. CNS Drugs 2013, 27, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, S.; Carlsson, A. Buspirone: Effects on central monoaminergic transmission--possible relevance to animal experimental and clinical findings. Eur. J. Pharmacol. 1982, 83, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Frances, H.; Lienard, C. Isolation-induced social behavioral deficit test: Effect of tranquilizing drugs. Pharmacol. Biochem. Behav. 1989, 34, 293–296. [Google Scholar] [CrossRef]
- Brewin, C.R.; Andrews, B.; Valentine, J.D. Meta-analysis of risk factors for posttraumatic stress disorder in trauma-exposed adults. J. Consult. Clin. Psychol. 2000, 68, 748–766. [Google Scholar] [CrossRef]
- Koenen, K.C.; Moffitt, T.E.; Poulton, R.; Martin, J.; Caspi, A. Early childhood factors associated with the development of post-traumatic stress disorder: Results from a longitudinal birth cohort. Psychol. Med. 2007, 37, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Liu, Y.P. Early life social experience and adulthood impulsivity-Implications of central serotonergic system during development. Med. Hypotheses 2020, 143, 110032. [Google Scholar] [CrossRef] [PubMed]
- Bickerdike, M.J.; Wright, I.K.; Marsden, C.A. Social isolation attenuates rat forebrain 5-HT release induced by KCI stimulation and exposure to a novel environment. Behav. Pharmacol. 1993, 4, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.K.; Ismail, H.; Upton, N.; Marsden, C.A. Effect of isolation rearing on 5-HT agonist-induced responses in the rat. Psychopharmacology 1991, 105, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Whitaker-Azmitia, P.; Zhou, F.; Hobin, J.; Borella, A. Isolation-rearing of rats produces deficits as adults in the serotonergic innervation of hippocampus. Peptides 2000, 21, 1755–1759. [Google Scholar] [CrossRef] [PubMed]
- Mahar, I.; Bambico, F.R.; Mechawar, N.; Nobrega, J.N. Stress, serotonin, and hippocampal neurogenesis in relation to depression and antidepressant effects. Neurosci. Biobehav. Rev. 2014, 38, 173–192. [Google Scholar] [CrossRef] [PubMed]
- Przegaliński, E.; Moryl, E.; Papp, M. The effect of 5-HT1A receptor ligands in a chronic mild stress model of depression. Neuropharmacology 1995, 34, 1305–1310. [Google Scholar] [CrossRef] [PubMed]
- Chekroud, A.M.; Gueorguieva, R.; Krumholz, H.M.; Trivedi, M.H.; Krystal, J.H.; McCarthy, G. Reevaluating the Efficacy and Predictability of Antidepressant Treatments: A Symptom Clustering Approach. JAMA Psychiatry 2017, 74, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Appelberg, B.G.; Syvälahti, E.K.; Koskinen, T.E.; Mehtonen, O.P.; Muhonen, T.T.; Naukkarinen, H.H. Patients with severe depression may benefit from buspirone augmentation of selective serotonin reuptake inhibitors: Results from a placebo-controlled, randomized, double-blind, placebo wash-in study. J. Clin. Psychiatry 2001, 62, 448–452. [Google Scholar] [CrossRef]
- Blier, P.; Ward, N.M. Is there a role for 5-HT1A agonists in the treatment of depression? Biol. Psychiatry 2003, 53, 193–203. [Google Scholar] [CrossRef]
- Blier, P.; Bergeron, R.; de Montigny, C. Selective activation of postsynaptic 5-HT1A receptors induces rapid antidepressant response. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 1997, 16, 333–338. [Google Scholar] [CrossRef]
- Sprouse, J.S.; Aghajanian, G.K. Responses of hippocampal pyramidal cells to putative serotonin 5-HT1A and 5-HT1B agonists: A comparative study with dorsal raphe neurons. Neuropharmacology 1988, 27, 707–715. [Google Scholar] [CrossRef]
- Kokras, N.; Pastromas, N.; Papasava, D.; de Bournonville, C.; Cornil, C.A.; Dalla, C. Sex differences in behavioral and neurochemical effects of gonadectomy and aromatase inhibition in rats. Psychoneuroendocrinology 2018, 87, 93–107. [Google Scholar] [CrossRef]
- Bergmann, B.M.; Seiden, L.S.; Landis, C.A.; Gilliland, M.A.; Rechtschaffen, A. Sleep deprivation in the rat: XVIII. Regional brain levels of monoamines and their metabolites. Sleep 1994, 17, 583–589. [Google Scholar] [CrossRef]
- Lalonde, R.; Strazielle, C. Regional variations of 5HT concentrations in Rorasg (staggerer) mutants. Neurochem. Res. 2006, 31, 921–924. [Google Scholar] [CrossRef]
- Sprouse, J.; Braselton, J.; Reynolds, L. 5-HT(1A) agonist potential of pindolol: Electrophysiologic studies in the dorsal raphe nucleus and hippocampus. Biol. Psychiatry 2000, 47, 1050–1055. [Google Scholar] [CrossRef]
- Carrasco, G.A.; Barker, S.A.; Zhang, Y.; Damjanoska, K.J.; Sullivan, N.R.; Garcia, F.; D’Souza, D.N.; Muma, N.A.; van De Kar, L.D. Estrogen treatment increases the levels of regulator of G protein signaling-Z1 in the hypothalamic paraventricular nucleus: Possible role in desensitization of 5-hydroxytryptamine1A receptors. Neuroscience 2004, 127, 261–267. [Google Scholar] [CrossRef]
- Lea, J.W.; Stoner, S.C.; Lafollette, J. Agitation associated with aripiprazole initiation. Pharmacotherapy 2007, 27, 1339–1342. [Google Scholar] [CrossRef]
- Sato, H.; Skelin, I.; Diksic, M. Chronic buspirone treatment decreases 5-HT1B receptor densities and the serotonin transporter but increases the density of 5-HT2A receptors in the bulbectomized rat model of depression: An autoradiographic study. Brain Res. 2010, 23, 28–44. [Google Scholar] [CrossRef]
- Jarończyk, M.; Chilmonczyk, Z.; Mazurek, A.P.; Nowak, G.; Ravna, A.W.; Kristiansen, K.; Sylte, I. The molecular interactions of buspirone analogues with the serotonin transporter. Bioorg. Med. Chem. 2008, 16, 9283–9294. [Google Scholar] [CrossRef]
- Ko, C.Y.; Liu, Y.P. Disruptions of sensorimotor gating, cytokines, glycemia, monoamines, and genes in both sexes of rats reared in social isolation can be ameliorated by oral chronic quetiapine administration. Brain Behav. Immun. 2016, 51, 119–130. [Google Scholar] [CrossRef]
- Lin, C.C.; Chen, T.Y.; Cheng, P.Y.; Liu, Y.P. Early life social experience affects adulthood fear extinction deficit and associated dopamine profile abnormalities in a rat model of PTSD. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 101, 109914. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzeng, N.-S.; Chung, J.-Y.; Lin, C.-C.; Cheng, P.-Y.; Liu, Y.-P. Effects of Subchronic Buspirone Treatment on Depressive Profile in Socially Isolated Rats: Implication of Early Life Experience on 5-HT1A Receptor-Related Depression. Pharmaceuticals 2024, 17, 717. https://doi.org/10.3390/ph17060717
Tzeng N-S, Chung J-Y, Lin C-C, Cheng P-Y, Liu Y-P. Effects of Subchronic Buspirone Treatment on Depressive Profile in Socially Isolated Rats: Implication of Early Life Experience on 5-HT1A Receptor-Related Depression. Pharmaceuticals. 2024; 17(6):717. https://doi.org/10.3390/ph17060717
Chicago/Turabian StyleTzeng, Nian-Sheng, Jing-Yi Chung, Chen-Cheng Lin, Pao-Yun Cheng, and Yia-Ping Liu. 2024. "Effects of Subchronic Buspirone Treatment on Depressive Profile in Socially Isolated Rats: Implication of Early Life Experience on 5-HT1A Receptor-Related Depression" Pharmaceuticals 17, no. 6: 717. https://doi.org/10.3390/ph17060717
APA StyleTzeng, N.-S., Chung, J.-Y., Lin, C.-C., Cheng, P.-Y., & Liu, Y.-P. (2024). Effects of Subchronic Buspirone Treatment on Depressive Profile in Socially Isolated Rats: Implication of Early Life Experience on 5-HT1A Receptor-Related Depression. Pharmaceuticals, 17(6), 717. https://doi.org/10.3390/ph17060717