

The Bioactive Compounds of Epimedium and Their Potential Mechanism of Action in Treating Osteoporosis: A Network Pharmacology and Experimental Validation Study
 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Identification of Potential Anti-Osteoporosis Compounds from Epimedium and Pathway Enrichment by Network Pharmacology Analysis
2.1.1. Databases of Epimedium Chemical Components and Known Anti-Osteoporosis Compounds
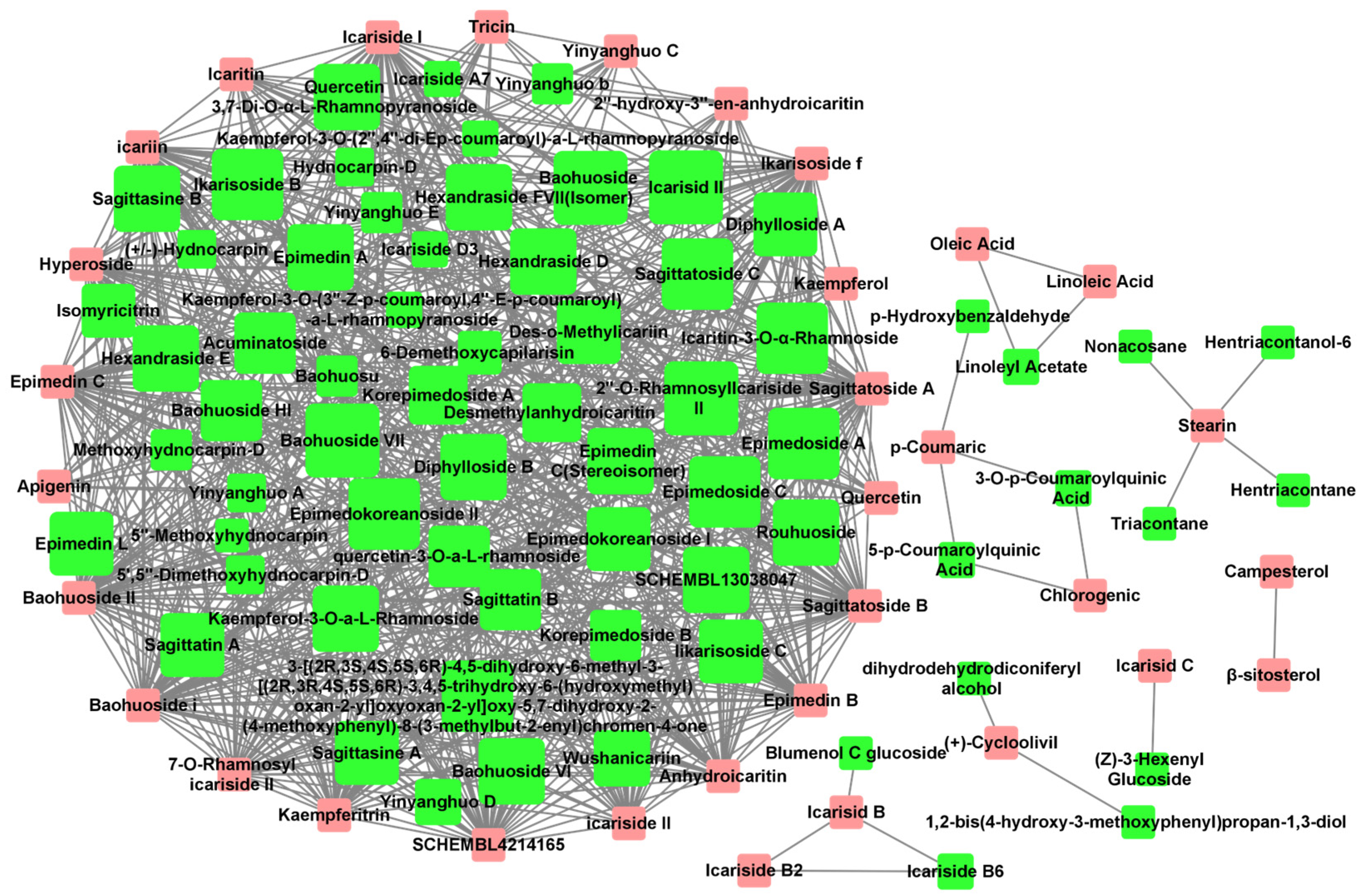
2.1.2. Potentially Active Compounds Prediction through the Construction of a Compound-Compound Interaction Network
2.1.3. Selection of Potential Active Compounds for Experimental Validation
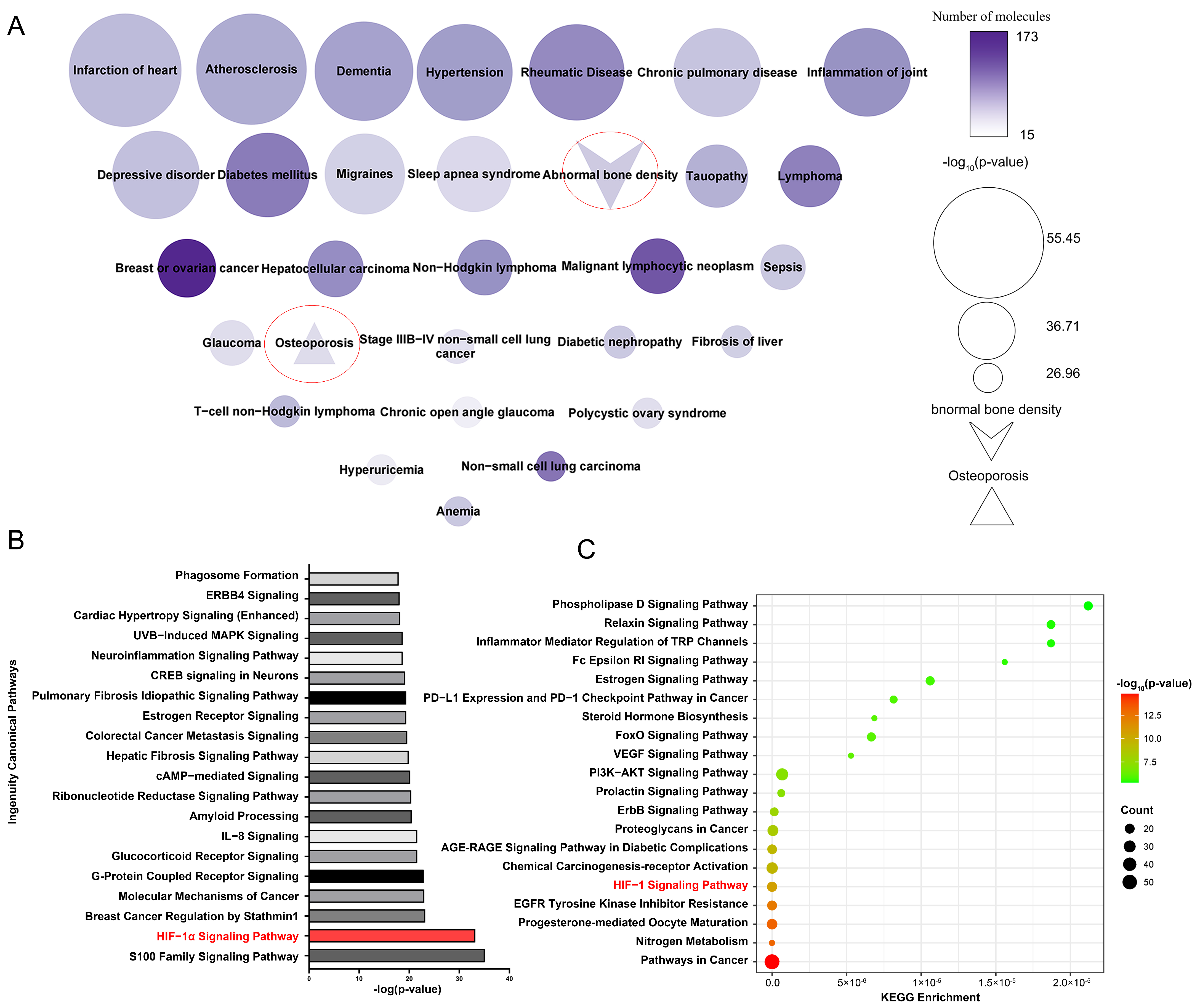
2.1.4. Selection of HIF-1α as Potential Target Protein for Further Experimental Validation
2.2. Identification of 2″-O-RhamnosylIcariside II as a Key Active Compound from Epimedium for Osteoporosis Treatment by Targeted Inhibition of HIF-1α: In Vitro Experimental Validation
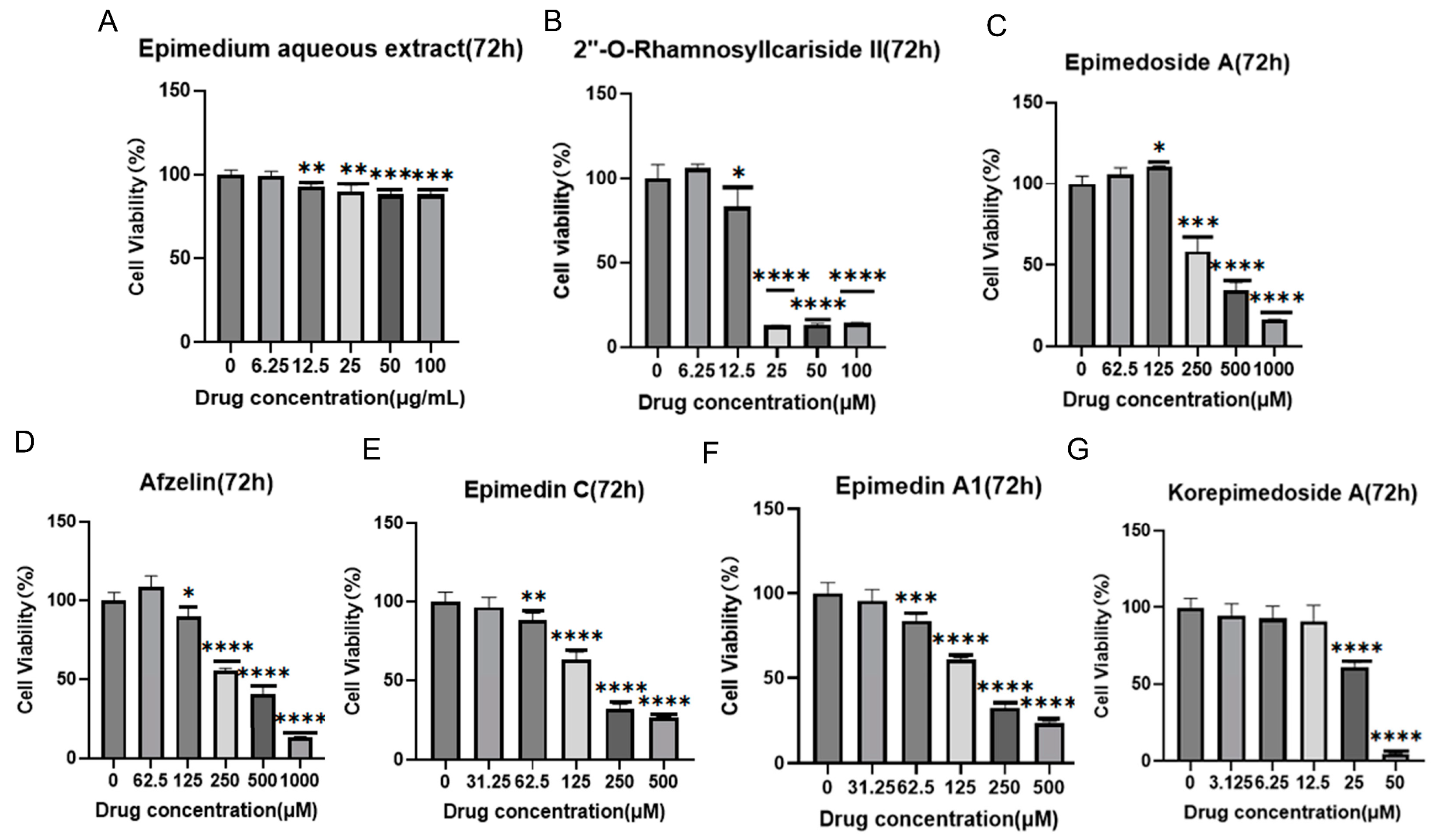
2.2.1. Identification of Active Compounds’ Non-Toxic Concentration from Epimedium
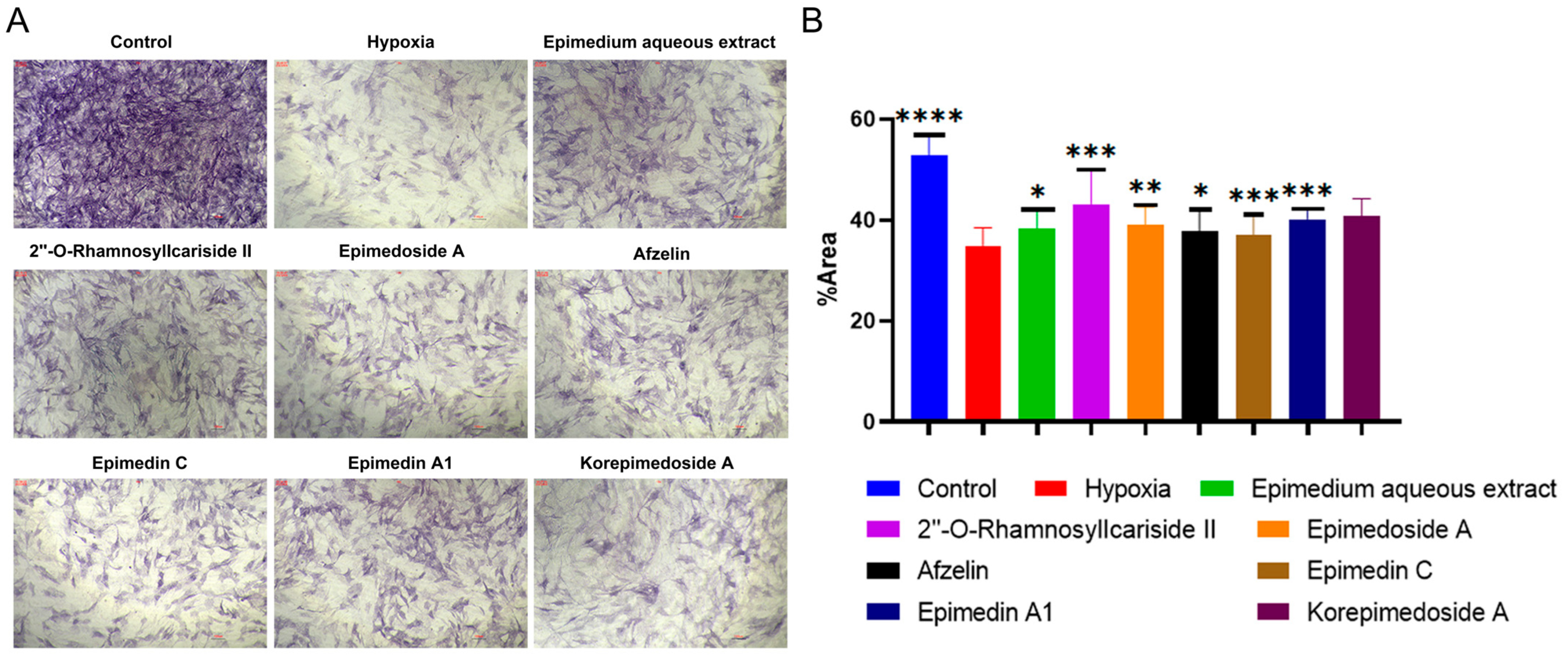
2.2.2. Identification of Active Compounds from Epimedium That Influence Osteoblast Differentiation of MC3T3-E1 Cells under Hypoxic Conditions
2.2.3. Identification of Active Compounds from Epimedium That Inhibit HIF-1α Gene Expression
2.2.4. Confirmation of 2″-O-RhamnosylIcariside II Existing in Epimedium Aqueous Extract Based on Mass Spectrometry Analysis
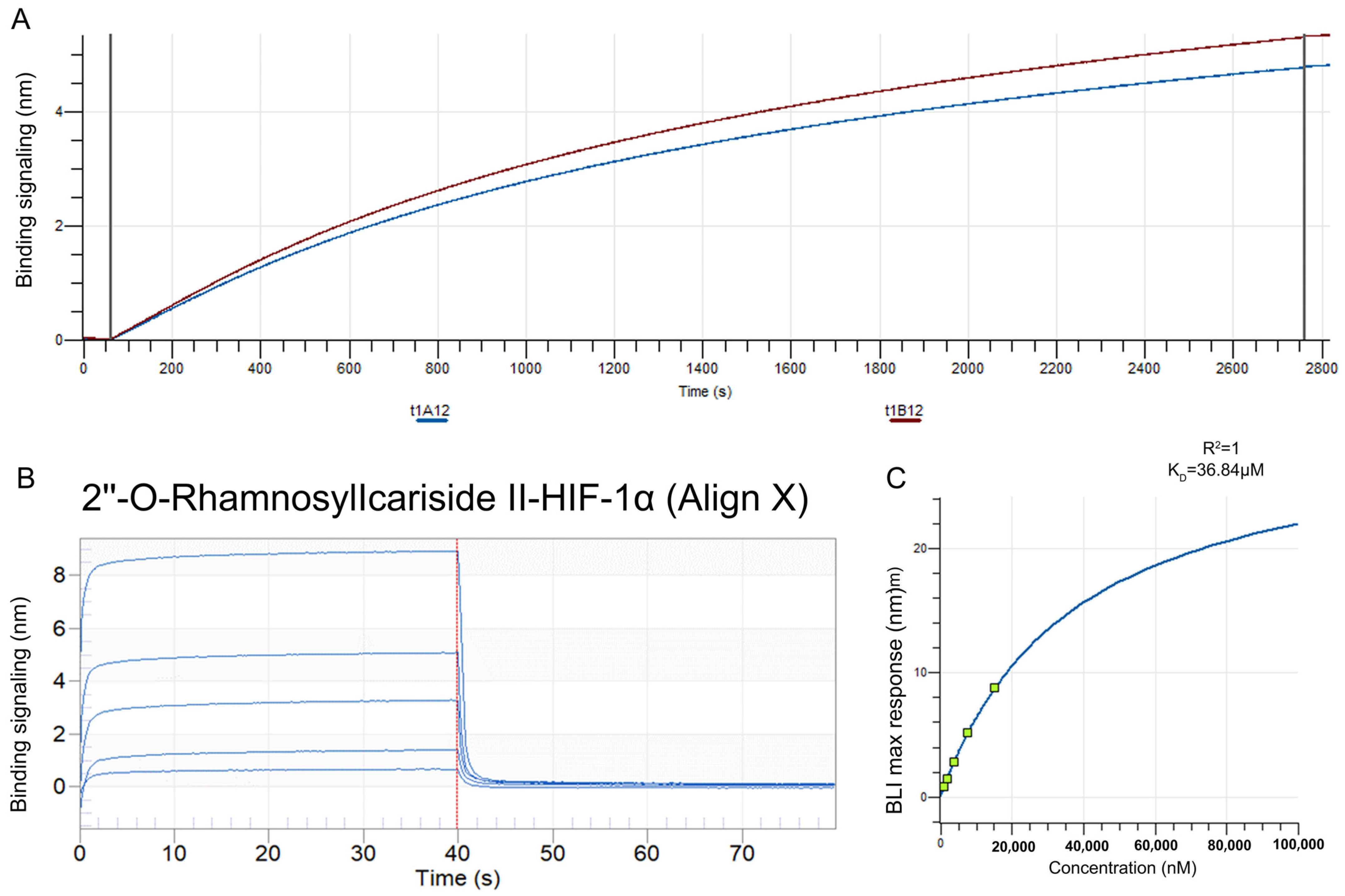
2.2.5. Validation of HIF-1α as a Target Protein for 2″-O-Rhamnosylicariside II Using Bio-Layer Interferometry (BLI)
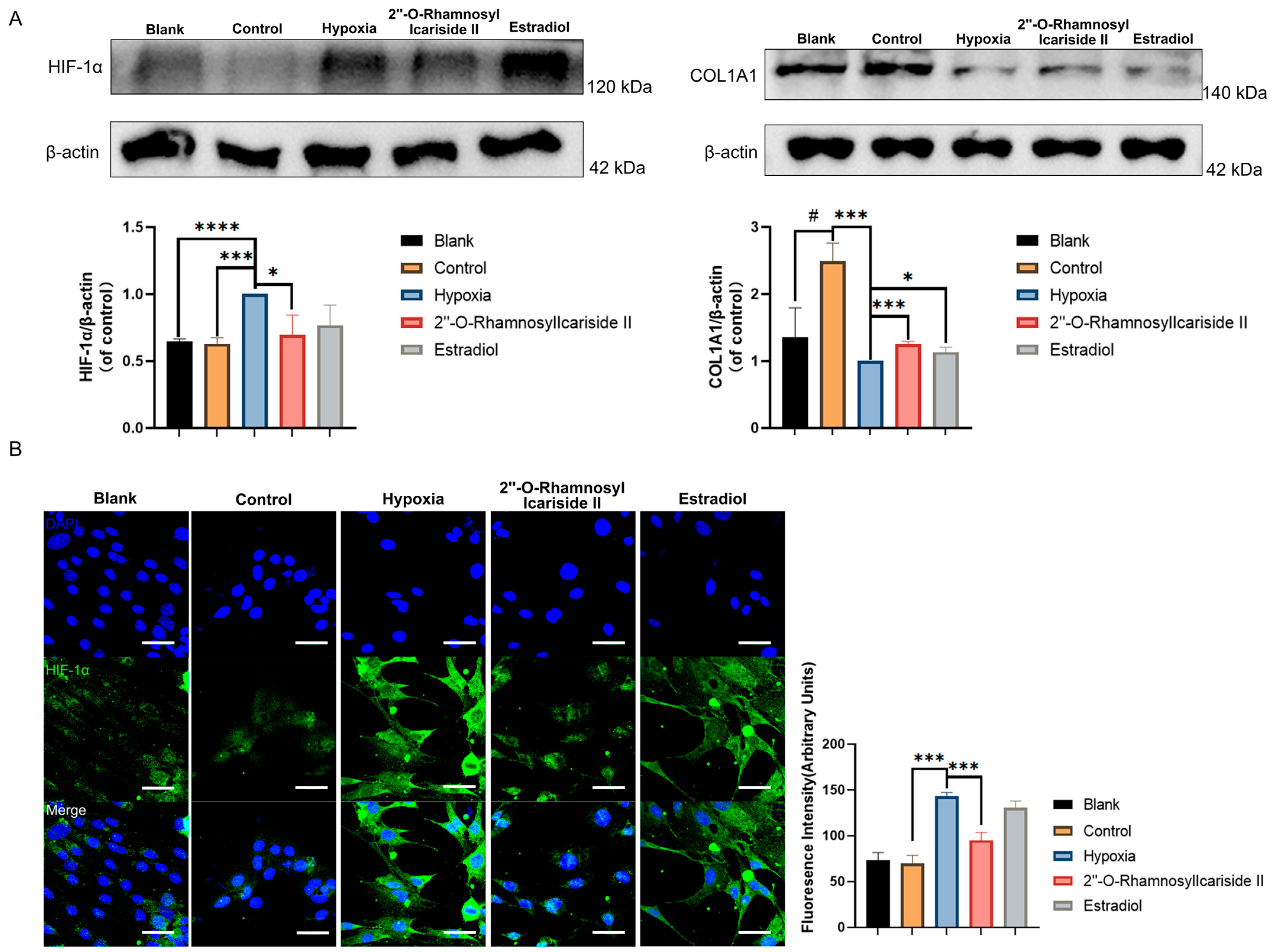
2.2.6. 2″-O-RhamnosylIcariside II Can Inhibit the HIF-1α and Increase COL1A1 Protein Expression in MC3T3-E1 Cells Differentiation
2.3. Validating 2″-O-RhamnosylIcariside II as an HIF-1α Inhibitor for Osteoporosis Treatment: In Vivo Experiments
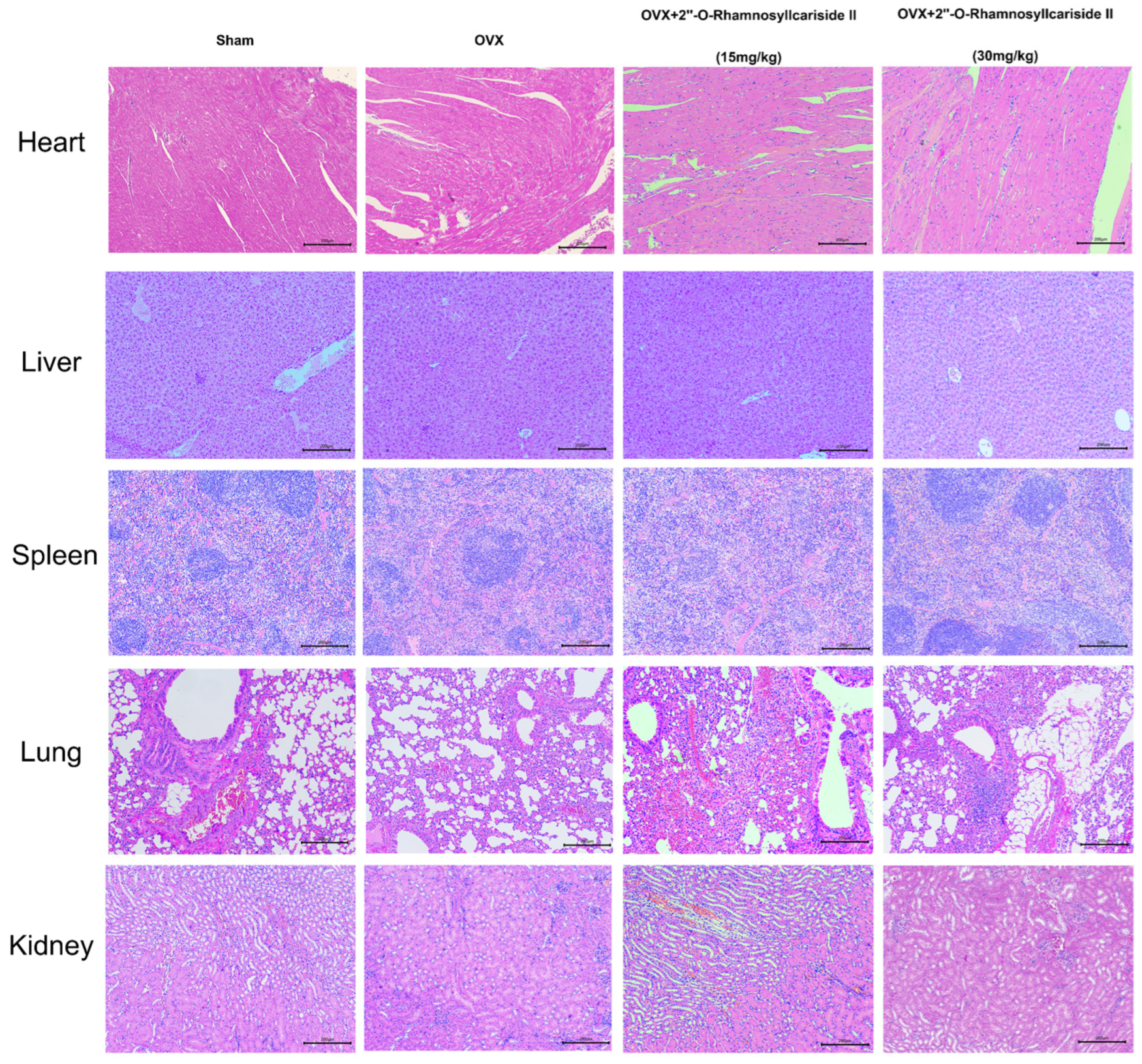
2.3.1. Assessment of In Vivo Safety of 2″-O-RhamnosylIcariside II
2.3.2. 2″-O-RhamnosylIcariside II Can Alleviate Bone Loss of OVX Model
2.3.3. 2″-O-RhamnosylIcariside II Alleviates Weight Gain in OVX Mice
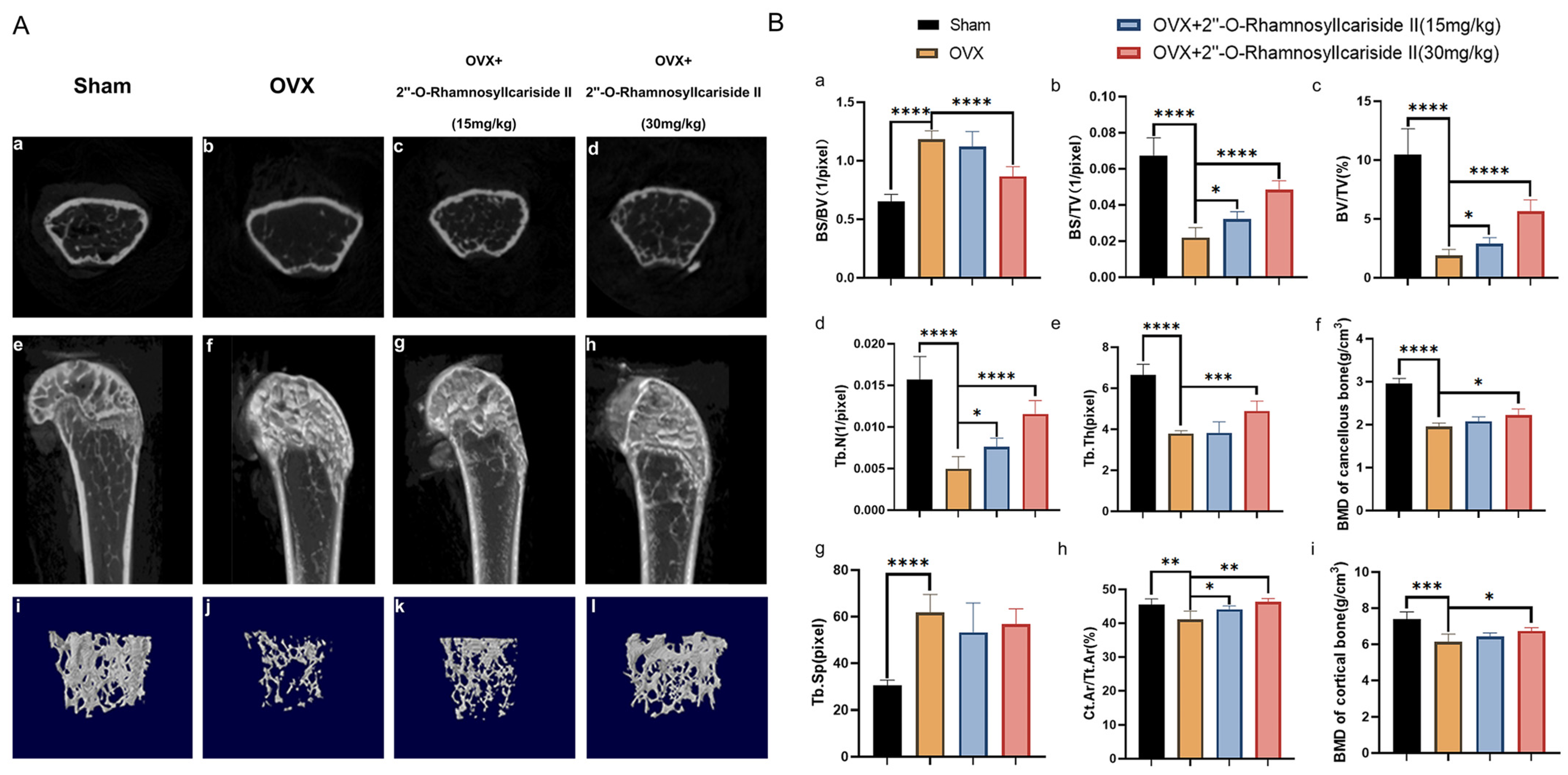
2.3.4. 2″-O-RhamnosylIcariside II Improved Bone Microstructure and Reduced Bone Marrow Adipose Tissues in OVX Mice
2.3.5. 2″-O-RhamnosylIcariside II Can Promote Bone Formation In Vivo
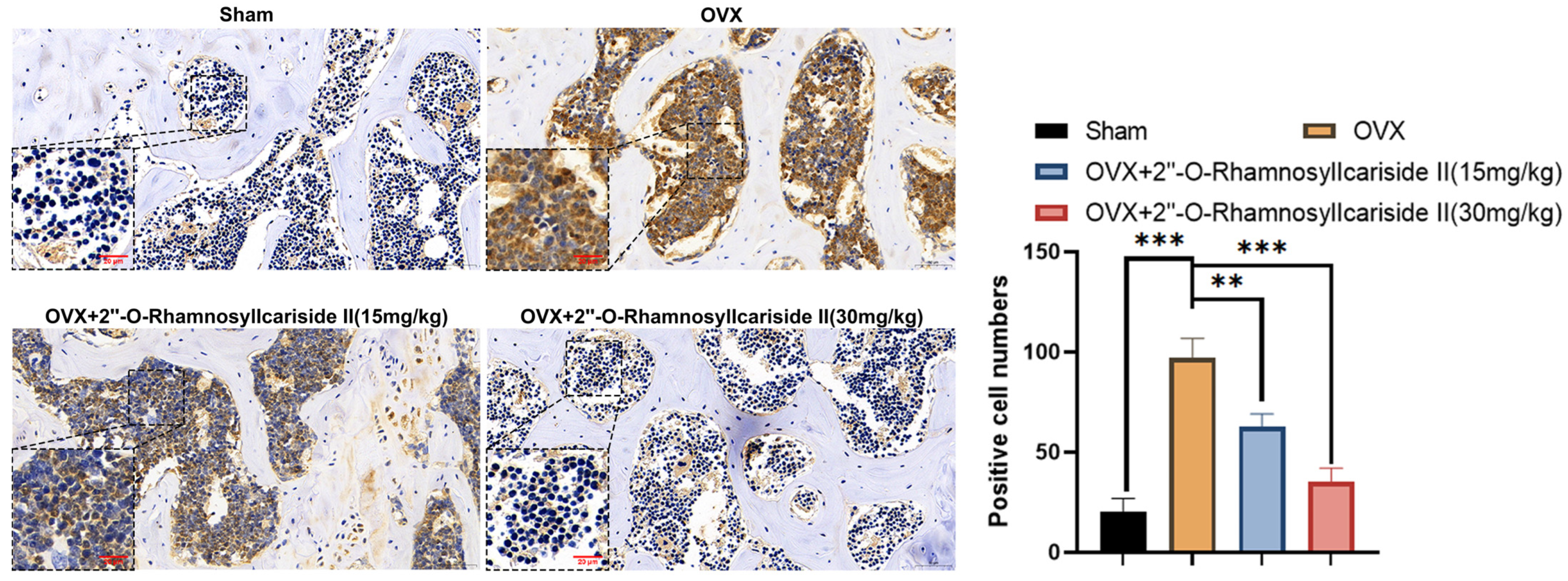
2.3.6. 2″-O-RhamnosylIcariside II Can Inhibit HIF-1α Protein Expression in Femur
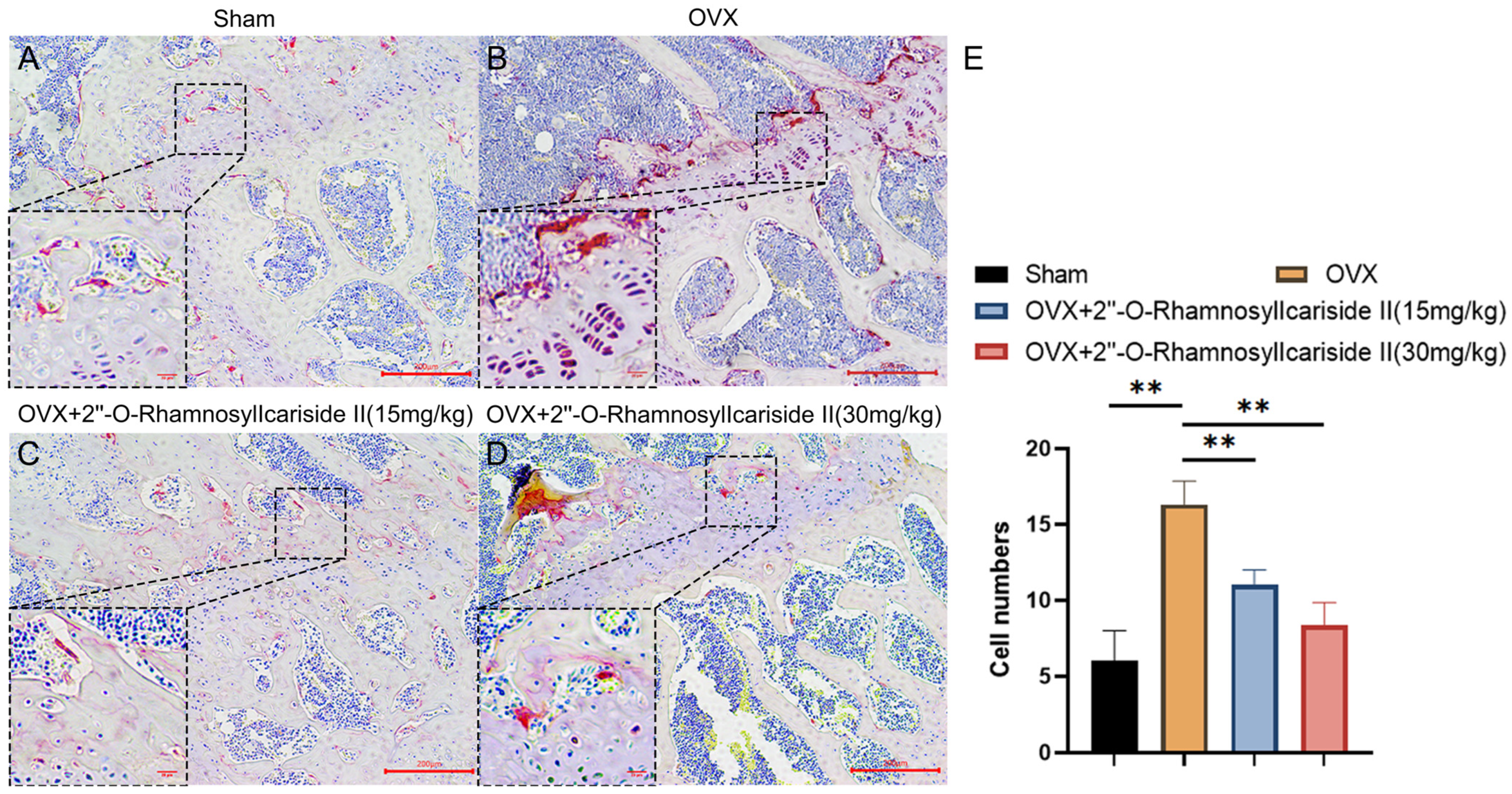
2.3.7. 2″-O-RhamnosylIcariside II Can Decrease the Number of Osteoclasts in the Femur of OVX Model
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Construction of the Epimedium Chemical Database
4.3. Construction of a Database with Known Anti-Osteoporosis Compounds in Epimedium
4.4. Construction of a Potential Active Compound Interaction Network Based on Molecular Similarity Calculations
4.5. Prediction of Potential Target Proteins and Pathway Enrichment Analysis
4.6. Preparation of the Aqueous Epimedium Extract
4.7. Cell Culture and Construction of Cellular Hypoxia Model
4.8. Cell Viability Experiments
4.9. ALP Staining
4.10. RT-qPCR Experiment
4.11. BLI Technology
4.12. Quality Control of 2″-O-RhamnosylIcariside II
4.13. Western Blot Analysis
4.14. Immunofluorescence Analysis
4.15. Animal Experiments
4.16. Histopathologic Analysis
4.17. Micro-CT and Histomorphometric Analysis
4.18. Statistical Analysis
4.19. Ethics Statement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ensrud, K.E.; Crandall, C.J. Osteoporosis. Ann. Intern. Med. 2017, 167, ITC17–ITC32. [Google Scholar] [CrossRef] [PubMed]
- Johnell, O.; Kanis, J.A. An Estimate of the Worldwide Prevalence and Disability Associated with Osteoporotic Fractures. Osteoporos. Int. 2006, 17, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Tella, S.H.; Gallagher, J.C. Prevention and Treatment of Postmenopausal Osteoporosis. J. Steroid Biochem. Mol. Biol. 2014, 142, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.B.; Dagar, M. Osteoporosis in Older Adults. Med. Clin. N. Am. 2020, 104, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Shieh, A.; Ishii, S.; Greendale, G.A.; Cauley, J.A.; Lo, J.C.; Karlamangla, A.S. Urinary N-Telopeptide and Rate of Bone Loss Over the Menopause Transition and Early Postmenopause. J. Bone Miner. Res. 2016, 31, 2057–2064. [Google Scholar] [CrossRef] [PubMed]
- Gosset, A.; Pouillès, J.-M.; Trémollieres, F. Menopausal Hormone Therapy for the Management of Osteoporosis. Best. Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101551. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Hofbauer, L.C. Osteoporosis Treatment: Recent Developments and Ongoing Challenges. Lancet Diabetes Endocrinol. 2017, 5, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Quattrocchi, E.; Kourlas, H. Teriparatide: A Review. Clin. Ther. 2004, 26, 841–854. [Google Scholar] [CrossRef] [PubMed]
- Lamy, O.; Stoll, D.; Aubry-Rozier, B.; Rodriguez, E.G. Stopping Denosumab. Curr. Osteoporos. Rep. 2019, 17, 8–15. [Google Scholar] [CrossRef]
- Zhuo, Y.; Li, M.; Jiang, Q.; Ke, H.; Liang, Q.; Zeng, L.-F.; Fang, J. Evolving Roles of Natural Terpenoids From Traditional Chinese Medicine in the Treatment of Osteoporosis. Front. Endocrinol. 2022, 13, 901545. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, S.; Zhang, H.; Gu, Y.; Chen, H.; Huang, Z.; Yang, F.; Li, W.; Chen, C.; Men, L.; et al. The Triangular Relationship between Traditional Chinese Medicines, Intestinal Flora, and Colorectal Cancer. Med. Res. Rev. 2024, 44, 539–567. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, J.; Wang, J.; Meng, T.; Yang, J.; Zhou, Y. The Role of Ferroptosis in Diabetic Cardiovascular Diseases and the Intervention of Active Ingredients of Traditional Chinese Medicine. Front. Pharmacol. 2023, 14, 1286718. [Google Scholar] [CrossRef]
- Liu, Z.-Q.; Sun, X.; Liu, Z.-B.; Zhang, T.; Zhang, L.-L.; Wu, C.-J. Phytochemicals in Traditional Chinese Medicine Can Treat Gout by Regulating Intestinal Flora through Inactivating NLRP3 and Inhibiting XOD Activity. J. Pharm. Pharmacol. 2022, 74, 919–929. [Google Scholar] [CrossRef]
- Guo, C.; He, J.; Song, X.; Tan, L.; Wang, M.; Jiang, P.; Li, Y.; Cao, Z.; Peng, C. Pharmacological Properties and Derivatives of Shikonin-A Review in Recent Years. Pharmacol. Res. 2019, 149, 104463. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Xiao, P. Comment on main species of herba epimedii. Zhongguo Zhong Yao Za Zhi 2003, 28, 303–307. [Google Scholar] [PubMed]
- Wang, Z.; Lou, Y. Proliferation-Stimulating Effects of Icaritin and Desmethylicaritin in MCF-7 Cells. Eur. J. Pharmacol. 2004, 504, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; He, X.; Yang, Y.; Li, M.; Hao, D.; Jia, Z. The Genus Epimedium: An Ethnopharmacological and Phytochemical Review. J. Ethnopharmacol. 2011, 134, 519–541. [Google Scholar] [CrossRef] [PubMed]
- Indran, I.R.; Liang, R.L.Z.; Min, T.E.; Yong, E.-L. Preclinical Studies and Clinical Evaluation of Compounds from the Genus Epimedium for Osteoporosis and Bone Health. Pharmacol. Ther. 2016, 162, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhou, S.; Qu, R.; Yang, Y.; Gong, X.; Hong, Y.; Jin, A.; Huang, X.; Dai, Q.; Jiang, L. Icariin Prevents Oestrogen Deficiency-Induced Alveolar Bone Loss through Promoting Osteogenesis via STAT3. Cell Prolif. 2020, 53, e12743. [Google Scholar] [CrossRef]
- Zhou, L.; Poon, C.C.-W.; Wong, K.-Y.; Cao, S.; Dong, X.; Zhang, Y.; Wong, M.-S. Icariin Ameliorates Estrogen-Deficiency Induced Bone Loss by Enhancing IGF-I Signaling via Its Crosstalk with Non-Genomic ERα Signaling. Phytomedicine 2021, 82, 153413. [Google Scholar] [CrossRef]
- Tsai, K.-C.; Huang, Y.-C.; Liaw, C.-C.; Tsai, C.-I.; Chiou, C.-T.; Lin, C.-J.; Wei, W.-C.; Lin, S.J.-S.; Tseng, Y.-H.; Yeh, K.-M.; et al. A Traditional Chinese Medicine Formula NRICM101 to Target COVID-19 through Multiple Pathways: A Bedside-to-Bench Study. Biomed. Pharmacother. 2021, 133, 111037. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Wang, Z.; Liu, W.; Pu, L.; Li, R.; Ai, C.; Xu, H.; Zhang, B.; Wang, T.; Zhang, X.; et al. Resveratrol Ameliorates High Altitude Hypoxia-Induced Osteoporosis by Suppressing the ROS/HIF Signaling Pathway. Molecules 2022, 27, 5538. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Lin, Z.; Cao, S.; Janowska, I.; Sonomoto, K.; Andreev, D.; Katharina, K.; Wen, J.; Knaup, K.X.; Wiesener, M.S.; et al. Estrogen-Mediated Downregulation of HIF-1α Signaling in B Lymphocytes Influences Postmenopausal Bone Loss. Bone Res. 2022, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Warbrick, I.; Rabkin, S.W. Hypoxia-Inducible Factor 1-Alpha (HIF-1α) as a Factor Mediating the Relationship between Obesity and Heart Failure with Preserved Ejection Fraction. Obes. Rev. 2019, 20, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.-Y.; Wang, X.-F.; Hua, F.-Z. HIF-1α in Osteoarthritis: From Pathogenesis to Therapeutic Implications. Front. Pharmacol. 2022, 13, 927126. [Google Scholar] [CrossRef] [PubMed]
- Qiao, S.; Zhang, X.; Chen, Z.; Zhao, Y.; Tzeng, C.-M. Alloferon-1 Ameliorates Estrogen Deficiency-Induced Osteoporosis through Dampening the NLRP3/Caspase-1/IL-1β/IL-18 Signaling Pathway. Int. Immunopharmacol. 2023, 124, 110954. [Google Scholar] [CrossRef] [PubMed]
- Knowles, H.J.; Cleton-Jansen, A.-M.; Korsching, E.; Athanasou, N.A. Hypoxia-Inducible Factor Regulates Osteoclast-Mediated Bone Resorption: Role of Angiopoietin-like 4. FASEB J. 2010, 24, 4648–4659. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Shao, Q.; Tang, Y.; Li, X.; Qi, X.; Jiang, R.; Liang, Y.; Kang, F. HIF-1α Regulates Osteoclast Activation and Mediates Osteogenesis during Mandibular Bone Repair via CT-1. Oral. Dis. 2022, 28, 428–441. [Google Scholar] [CrossRef] [PubMed]
- Frey, J.L.; Stonko, D.P.; Faugere, M.-C.; Riddle, R.C. Hypoxia-Inducible Factor-1α Restricts the Anabolic Actions of Parathyroid Hormone. Bone Res. 2014, 2, 14005. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, M.; Feng, H.; Peng, Y.; Sun, J.; Qu, X.; Li, C. Lactate Induces Osteoblast Differentiation by Stabilization of HIF1α. Mol. Cell Endocrinol. 2017, 452, 84–92. [Google Scholar] [CrossRef]
- Miyauchi, Y.; Sato, Y.; Kobayashi, T.; Yoshida, S.; Mori, T.; Kanagawa, H.; Katsuyama, E.; Fujie, A.; Hao, W.; Miyamoto, K.; et al. HIF1α Is Required for Osteoclast Activation by Estrogen Deficiency in Postmenopausal Osteoporosis. Proc. Natl. Acad. Sci. USA 2013, 110, 16568–16573. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wan, C.; Deng, L.; Liu, X.; Cao, X.; Gilbert, S.R.; Bouxsein, M.L.; Faugere, M.-C.; Guldberg, R.E.; Gerstenfeld, L.C.; et al. The Hypoxia-Inducible Factor Alpha Pathway Couples Angiogenesis to Osteogenesis during Skeletal Development. J. Clin. Investig. 2007, 117, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Kho, D.H.; Yanagawa, T.; Harazono, Y.; Gao, X.; Hogan, V.; Raz, A. Galectin-3 Inhibits Osteoblast Differentiation through Notch Signaling. Neoplasia 2014, 16, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Malaval, L.; Gupta, A.K.; Aubin, J.E. Simultaneous Detection of Multiple Bone-Related mRNAs and Protein Expression during Osteoblast Differentiation: Polymerase Chain Reaction and Immunocytochemical Studies at the Single Cell Level. Dev. Biol. 1994, 166, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-G.; Sun, J.; Shen, W.-W.; Yang, S.-Z.; Zhang, Y.; Hu, X.; Qiu, H.; Xu, S.-C.; Chu, T.-W. Sema4D Expression and Secretion Are Increased by HIF-1α and Inhibit Osteogenesis in Bone Metastases of Lung Cancer. Clin. Exp. Metastasis 2019, 36, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.B.; Cargill, S.L.; Anderson, G.B.; Carey, J.R. Ovarian Status Influenced the Rate of Body-Weight Change but Not the Total Amount of Body-Weight Gained or Lost in Female CBA/J Mice. Exp. Gerontol. 2010, 45, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jiang, H.; Wang, B.; Gu, M.; Zhang, N.; Liang, W.; Wang, Y. Effect of Leptin on Marrow Adiposity in Ovariectomized Rabbits Assessed by Proton Magnetic Resonance Spectroscopy. J. Comput. Assist. Tomogr. 2018, 42, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Beekman, K.M.; Veldhuis-Vlug, A.G.; den Heijer, M.; Maas, M.; Oleksik, A.M.; Tanck, M.W.; Ott, S.M.; van ’t Hof, R.J.; Lips, P.; Bisschop, P.H.; et al. The Effect of Raloxifene on Bone Marrow Adipose Tissue and Bone Turnover in Postmenopausal Women with Osteoporosis. Bone 2019, 118, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Wojdasiewicz, P.; Turczyn, P.; Lach-Gruba, A.; Poniatowski, Ł.A.; Purrahman, D.; Mahmoudian-Sani, M.-R.; Szukiewicz, D. The Role of Rosavin in the Pathophysiology of Bone Metabolism. Int. J. Mol. Sci. 2024, 25, 2117. [Google Scholar] [CrossRef]
- Li, D.; Yuan, T.; Zhang, X.; Xiao, Y.; Wang, R.; Fan, Y.; Zhang, X. Icariin: A Potential Promoting Compound for Cartilage Tissue Engineering. Osteoarthr. Cartil. 2012, 20, 1647–1656. [Google Scholar] [CrossRef]
- Gao, S.; Cheng, L.; Li, K.; Liu, W.-H.; Xu, M.; Jiang, W.; Wei, L.-C.; Zhang, F.; Xiao, W.; Xiong, Y.; et al. Effect of Epimedium Pubescen Flavonoid on Bone Mineral Status and Bone Turnover in Male Rats Chronically Exposed to Cigarette Smoke. BMC Musculoskelet. Disord. 2012, 13, 105. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, B.; Che, J.; Shang, P. Hypoxia Pathway in Osteoporosis: Laboratory Data for Clinical Prospects. Int. J. Environ. Res. Public Health 2023, 20, 3129. [Google Scholar] [CrossRef]
- Fujimoto, H.; Fujimoto, K.; Ueda, A.; Ohata, M. Hypoxemia Is a Risk Factor for Bone Mass Loss. J. Bone Miner. Metab. 1999, 17, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fan, X.; Tao, J.; Xu, T.; Zhang, Y.; Zhang, W.; Li, L.; Li, X.; Ding, H.; Sun, M.; et al. Impact of Prenatal Hypoxia on Fetal Bone Growth and Osteoporosis in Ovariectomized Offspring Rats. Reprod. Toxicol. 2018, 78, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fayed, H.A.; Barakat, B.M.; Elshaer, S.S.; Abdel-Naim, A.B.; Menze, E.T. Antiosteoporotic Activities of Isoquercitrin in Ovariectomized Rats: Role of Inhibiting Hypoxia Inducible Factor-1 Alpha. Eur. J. Pharmacol. 2019, 865, 172785. [Google Scholar] [CrossRef]
- Knowles, H.J. Distinct Roles for the Hypoxia-Inducible Transcription Factors HIF-1α and HIF-2α in Human Osteoclast Formation and Function. Sci. Rep. 2020, 10, 21072. [Google Scholar] [CrossRef]
- Wang, X.; Wei, L.; Li, Q.; Lai, Y. HIF-1α Protects Osteoblasts from ROS-Induced Apoptosis. Free Radic. Res. 2022, 56, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Sato, Y.; Iwasaki, R.; Kobayashi, T.; Watanabe, R.; Oike, T.; Miyamoto, K.; Toyama, Y.; Matsumoto, M.; Nakamura, M.; et al. Selective Estrogen Receptor Modulators Suppress Hif1α Protein Accumulation in Mouse Osteoclasts. PLoS ONE 2016, 11, e0165922. [Google Scholar] [CrossRef]
- Chen, D.; Li, Y.; Zhou, Z.; Xing, Y.; Zhong, Y.; Zou, X.; Tian, W.; Zhang, C. Synergistic Inhibition of Wnt Pathway by HIF-1α and Osteoblast-Specific Transcription Factor Osterix (Osx) in Osteoblasts. PLoS ONE 2012, 7, e52948. [Google Scholar] [CrossRef]
- Li, L.; Li, A.; Zhu, L.; Gan, L.; Zuo, L. Roxadustat Promotes Osteoblast Differentiation and Prevents Estrogen Deficiency-Induced Bone Loss by Stabilizing HIF-1α and Activating the Wnt/β-Catenin Signaling Pathway. J. Orthop. Surg. Res. 2022, 17, 286. [Google Scholar] [CrossRef]
- Tang, Y.-X.; Liu, M.; Liu, L.; Zhen, B.-R.; Wang, T.-T.; Li, N.; Lv, N.; Zhu, Z.; Sun, G.; Wang, X.; et al. Lipophilic Constituents in Salvia Miltiorrhiza Inhibit Activation of the Hepatic Stellate Cells by Suppressing the JAK1/STAT3 Signaling Pathway: A Network Pharmacology Study and Experimental Validation. Front. Pharmacol. 2022, 13, 770344. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, X.; Zhu, D.; Liu, T.; Liang, X.; Liu, F.; Zhang, Y.; Dong, X.; Sun, B. HIF-1α Promoted Vasculogenic Mimicry Formation in Hepatocellular Carcinoma through LOXL2 up-Regulation in Hypoxic Tumor Microenvironment. J. Exp. Clin. Cancer Res. 2017, 36, 60. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candidate Compound | CAS (Chemical Abstracts Service Number) | Tc Similarity |
|---|---|---|
| 2″-O-Rhamnosylicariside II | 135293-13-9 | 0.65983897 |
| Epimedoside A | 39012-04-9 | 0.650410123 |
| Afzelin | 482-39-3 | 0.630783002 |
| Epimedin C | 110642-44-9 | 0.624386306 |
| Epimedin A1 | 140147-77-9 | 0.596296546 |
| Korepimedoside A | 106441-31-0 | 0.502801148 |
| Compound | Concentration for MC3T3-E1 |
|---|---|
| Epimedium aqueous extract | 6.25 μg/mL |
| 2″-O-RhamnosylIcariside II | 6.25 μM |
| Epimedoside A | 62.5 μM |
| Afzelin | 62.5 μM |
| Epimedin C | 31.25 μM |
| Epimedin A1 | 31.25 μM |
| Korepimedoside A | 12.5 μM |
| Control | Hypoxia | Treatment | |
|---|---|---|---|
| Induction medium | ✓ | ✓ | ✓ |
| Treatment | ✓ | ||
| CoCl2 (100 μM) | ✓ | ✓ |
| Gene | Primer Sequence (5′→3′) |
|---|---|
| Mouse Hif1a | Upstream: GTGCACAGAGCCTCCTCG |
| Downstream: AGCTGGAAGGTTTGTGGTGT | |
| Mouse Atcb | Upstream: AGCCATGTACGTAGCCATCC |
| Downstream: GACTCCATCACAATGCCAGT |
| Blank | Control | Hypoxia | Treatment | |
|---|---|---|---|---|
| Normal medium | ✓ | |||
| Induction medium | ✓ | ✓ | ✓ | |
| Treatment | ✓ | |||
| CoCl2 (100 μM) | ✓ | ✓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, H.; Tang, F.; Zhao, Z.; Huang, W.; Wan, X.; Hong, Z.; Liu, Y.; Dong, X.; Chen, S. The Bioactive Compounds of Epimedium and Their Potential Mechanism of Action in Treating Osteoporosis: A Network Pharmacology and Experimental Validation Study. Pharmaceuticals 2024, 17, 706. https://doi.org/10.3390/ph17060706
Dong H, Tang F, Zhao Z, Huang W, Wan X, Hong Z, Liu Y, Dong X, Chen S. The Bioactive Compounds of Epimedium and Their Potential Mechanism of Action in Treating Osteoporosis: A Network Pharmacology and Experimental Validation Study. Pharmaceuticals. 2024; 17(6):706. https://doi.org/10.3390/ph17060706
Chicago/Turabian StyleDong, Huizhong, Fen Tang, Zilu Zhao, Wenxuan Huang, Xiangyang Wan, Zhanying Hong, Ying Liu, Xin Dong, and Si Chen. 2024. "The Bioactive Compounds of Epimedium and Their Potential Mechanism of Action in Treating Osteoporosis: A Network Pharmacology and Experimental Validation Study" Pharmaceuticals 17, no. 6: 706. https://doi.org/10.3390/ph17060706
APA StyleDong, H., Tang, F., Zhao, Z., Huang, W., Wan, X., Hong, Z., Liu, Y., Dong, X., & Chen, S. (2024). The Bioactive Compounds of Epimedium and Their Potential Mechanism of Action in Treating Osteoporosis: A Network Pharmacology and Experimental Validation Study. Pharmaceuticals, 17(6), 706. https://doi.org/10.3390/ph17060706