
Lactoferrins in Their Interactions with Molecular Targets: A Structure-Based Overview

Abstract
1. Introduction
Further Insight on the C-Lobe of Lactoferrin
2. Structural Characterization of Lactoferrin Interactions with Bacterial Proteins
2.1. Evolutionary Overview on Lactoferrin Receptors in Bacteria
2.2. Characterization of Lactoferrin Interactions with Bacterial Membranes
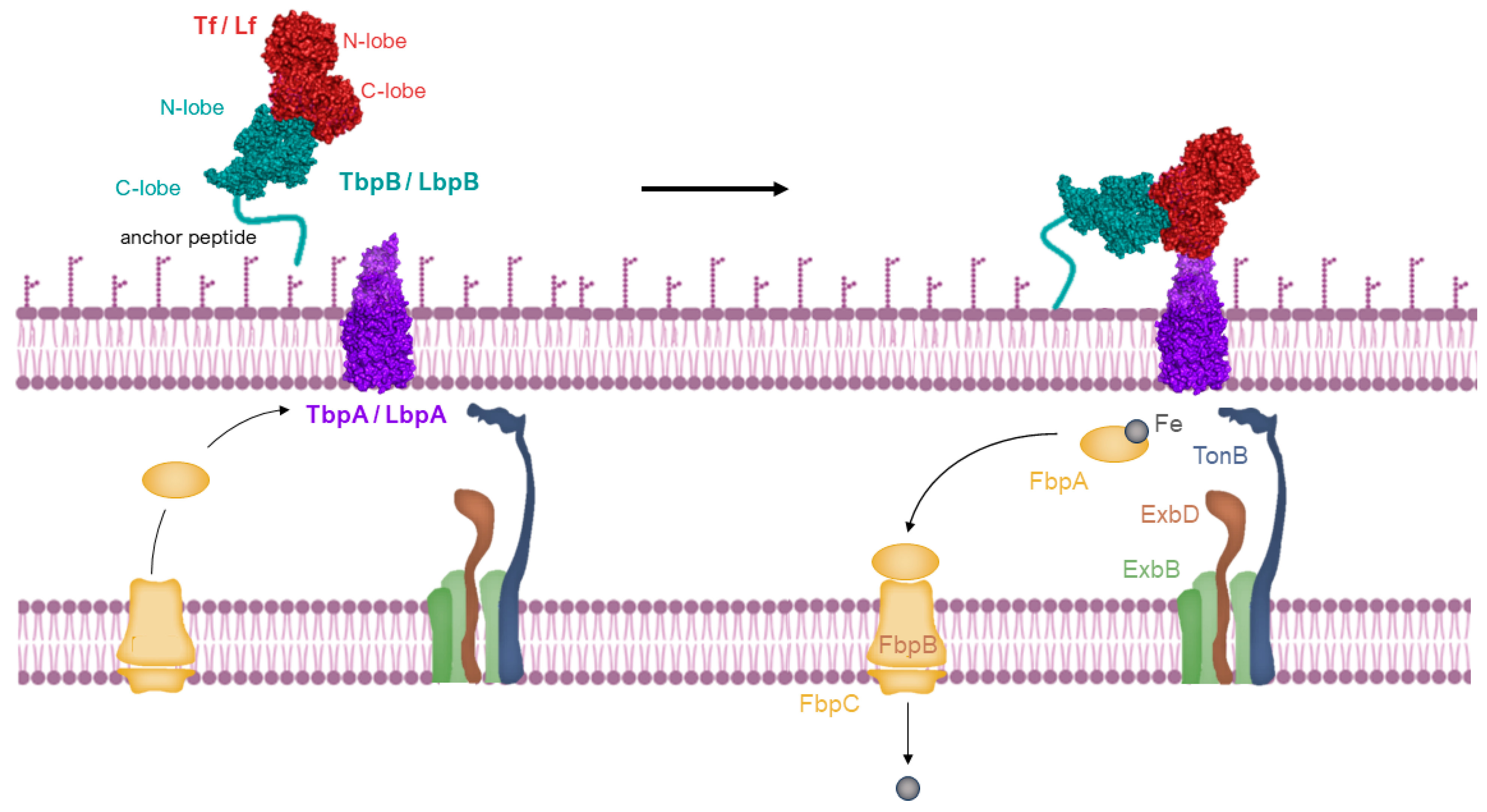
2.3. The Process of Iron Uptake Mediated by Membrane Receptors
2.4. Crystalloghaphic and Cryo-EM Data of Lactoferrin-Binding Protein
Evidence of Interactions with Porins
3. Lactoferrin Interactions with Viral Proteins
4. Computational Approaches for the Investigation of Lactoferrin Activity against SARS-CoV-2 Infection
5. Lactoferrin-Derived Peptides and Chimera
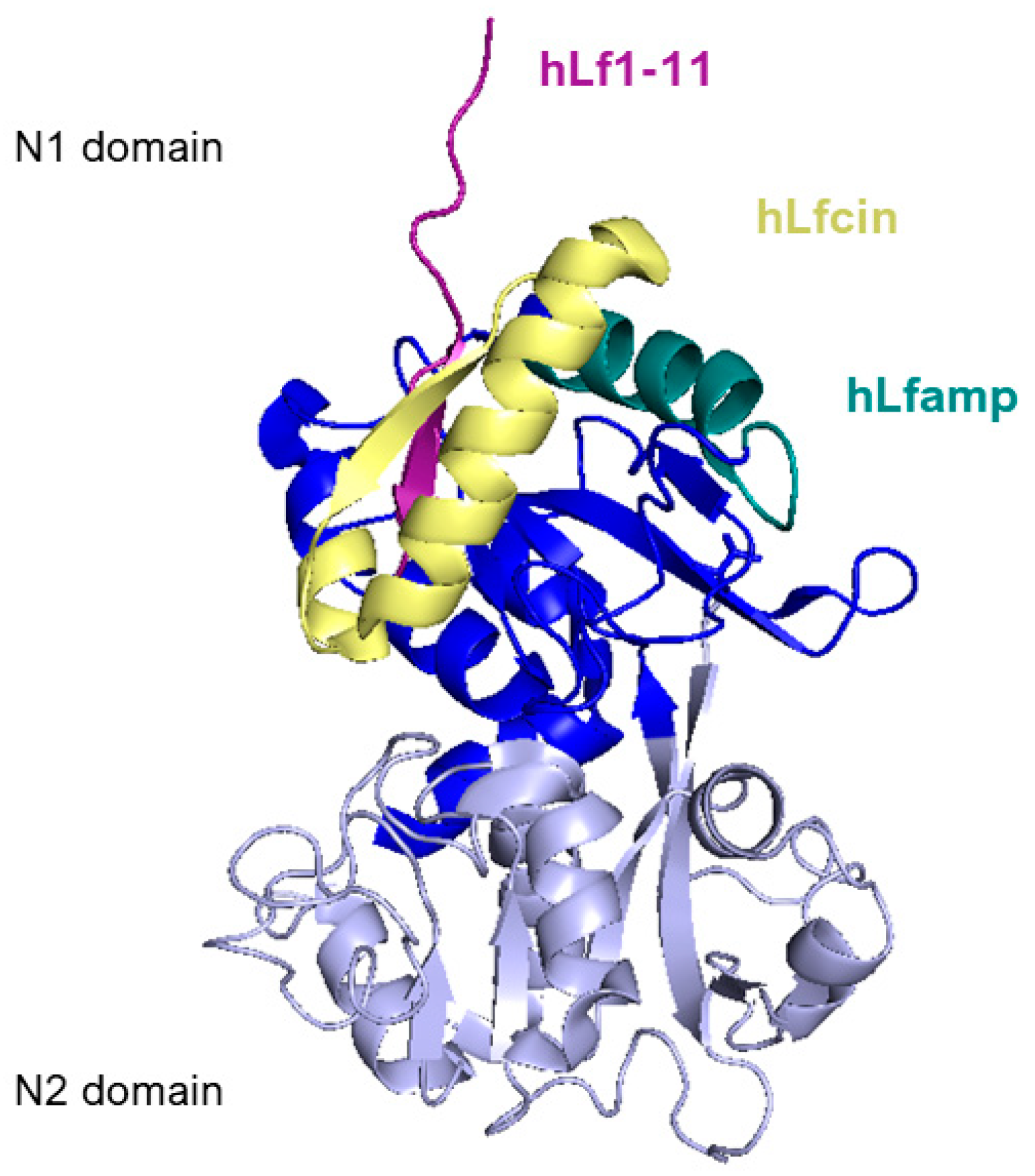
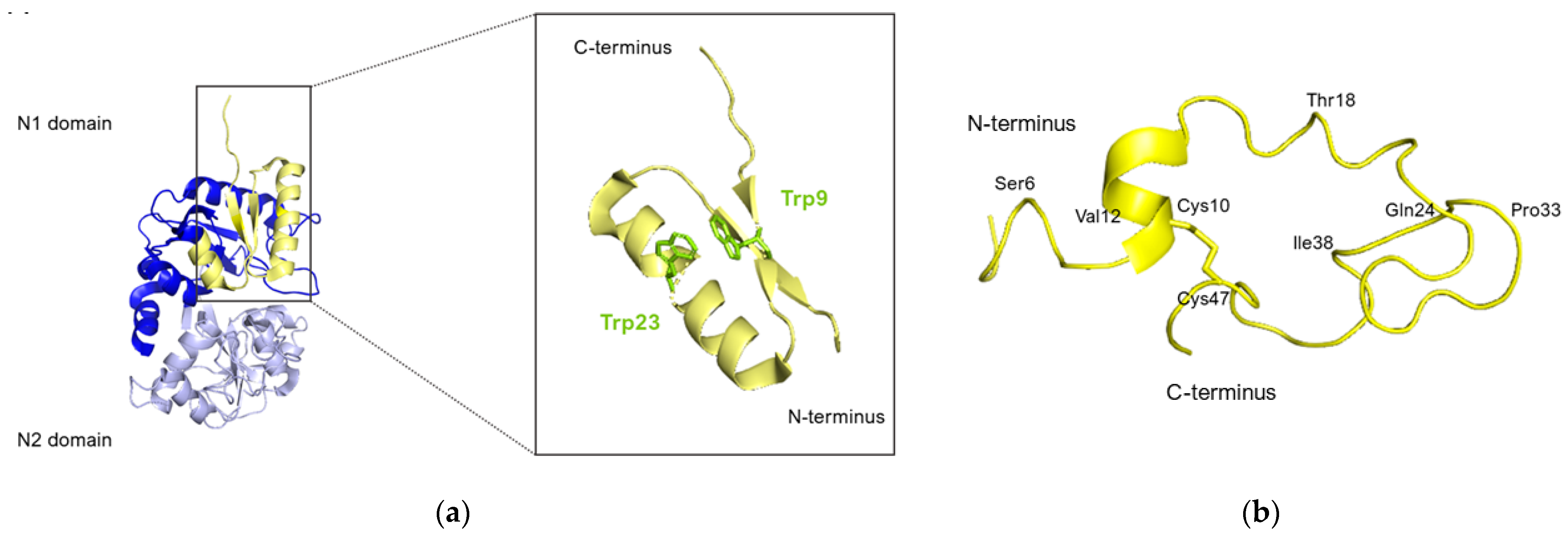
5.1. Lactoferricin
5.2. Lactoferrampin
5.3. Lf1-11
5.4. Other Lactoferrin-Derived Peptides with Antimicrobial Activities
5.5. Lactoferrin-Chimera
6. Lactoferrin Interaction with Plasma Proteins and Human Cell Receptors
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lambert, L.A. Molecular Evolution of the Transferrin Family and Associated Receptors. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.N.; Anderson, B.F.; Baker, H.M.; Day, C.L.; Haridas, M.; Norris, G.E.; Rumball, S.V.; Smith, C.A.; Thomas, D.H. Three-Dimensional Structure of Lactoferrin in Various Functional States. Adv. Exp. Med. Biol. 1994, 357, 1–12. [Google Scholar] [PubMed]
- Lambert, L.A.; Perri, H.; Halbrooks, P.J.; Mason, A.B. Evolution of the Transferrin Family: Conservation of Residues Associated with Iron and Anion Binding. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 142, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Schaeffer, E.; Sidoli, A.; Baralle, F.E.; Cohen, G.N.; Zakin, M.M. Organization of the Human Transferrin Gene: Direct Evidence That It Originated by Gene Duplication. Proc. Natl. Acad. Sci. USA 1985, 82, 3149–3153. [Google Scholar] [CrossRef] [PubMed]
- Masson, P.L.; Heremans, J.F.; Dive, C.H. An Iron-Binding Protein Common to Many External Secretions. Clin. Chim. Acta 1966, 14, 735–739. [Google Scholar] [CrossRef]
- Berlov, M.N.; Korableva, E.S.; Andreeva, Y.V.; Ovchinnikova, T.V.; Kokryakov, V.N. Lactoferrin from Canine Neutrophils: Isolation and Physicochemical and Antimicrobial Properties. Biochemistry 2007, 72, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.F.; Baker, H.M.; Dodson, E.J.; Norris, G.E.; Rumball, S.V.; Waters, J.M.; Baker, E.N. Structure of Human Lactoferrin at 3.2 Å Resolution. Proc. Natl. Acad. Sci. USA 1987, 84, 1769–1773. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.F.; Baker, H.M.; Norris, G.E.; Rice, D.W.; Baker, E.N. Structure of Human Lactoferrin: Crystallographic Structure Analysis and Refinement at 2.8 Å Resolution. J. Mol. Biol. 1989, 209, 711–734. [Google Scholar] [CrossRef]
- Haridas, M.; Anderson, B.F.; Baker, E.N. Structure of Human Diferric Lactoferrin Refined at 2.2 Å Resolution. Acta Crystallogr. D Biol. Crystallogr. 1995, 51, 629–646. [Google Scholar] [CrossRef]
- Moore, S.A.; Anderson, B.F.; Groom, C.R.; Haridas, M.; Baker, E.N. Three-Dimensional Structure of Diferric Bovine Lactoferrin at 2.8 Å Resolution. J. Mol. Biol. 1997, 274, 222–236. [Google Scholar] [CrossRef]
- Khan, J.A.; Kumar, P.; Paramasivam, M.; Yadav, R.S.; Sahani, M.S.; Sharma, S.; Srinivasan, A.; Singh, T.P. Camel Lactoferrin, a Transferrin-Cum-Lactoferrin: Crystal Structure of Camel Apolactoferrin at 2.6 Å Resolution and Structural Basis of Its Dual Role. J. Mol. Biol. 2001, 309, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Yadav, S.; Paramasivam, M.; Srinivasan, A.; Singh, T.P. Structure of Buffalo Lactoferrin at 3.3 Å Resolution at 277 K. Acta Crystallogr. D Biol. Crystallogr. 2000, 56, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Paramasivam, M.; Yadav, S.; Srinivasan, A.; Singh, T.P. Structure of Buffalo Lactoferrin at 2.5 Å Resolution Using Crystals Grown at 303 K Shows Different Orientations of the N and C Lobes. Acta Crystallogr. D Biol. Crystallogr. 1999, 55, 1805–1813. [Google Scholar] [CrossRef]
- Sharma, A.K.; Paramasivam, M.; Srinivasan, A.; Yadav, M.P.; Singh, T.P. Three-Dimensional Structure of Mare Diferric Lactoferrin at 2.6 Å Resolution. J. Mol. Biol. 1999, 289, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.M.; Baker, E.N. Lactoferrin and Iron: Structural and Dynamic Aspects of Binding and Release. BioMetals 2004, 17, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.N.; Baker, H.M. Lactoferrin. Cell. Mol. Life Sci. 2005, 62, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Mason, A.B.; Halbrooks, P.J.; James, N.G.; Connolly, S.A.; Larouche, J.R.; Smith, V.C.; MacGillivray, R.T.A.; Chasteen, N.D. Mutational Analysis of C-Lobe Ligands of Human Serum Transferrin: Insights into the Mechanism of Iron Release. Biochemistry 2005, 44, 8013–8021. [Google Scholar] [CrossRef] [PubMed]
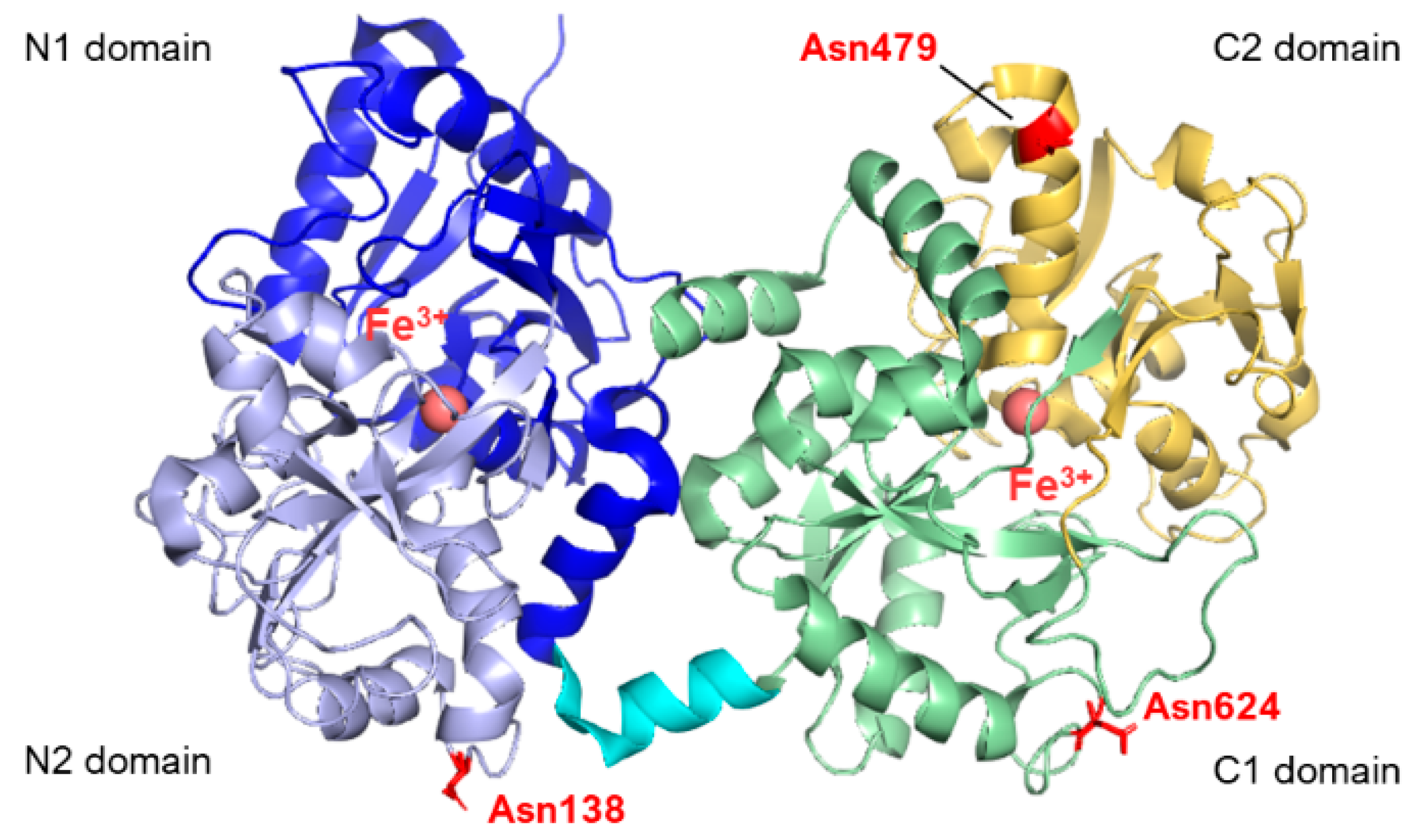
- Zlatina, K.; Galuska, S.P. The N-Glycans of Lactoferrin: More than Just a Sweet Decoration. Biochem. Cell Biol. 2021, 99, 117–127. [Google Scholar] [CrossRef]
- Karav, S.; German, J.; Rouquié, C.; Le Parc, A.; Barile, D. Studying Lactoferrin N-Glycosylation. Int. J. Mol. Sci. 2017, 18, 870. [Google Scholar] [CrossRef]
- Van Veen, H.A.; Geerts, M.E.J.; Van Berkel, P.H.C.; Nuijens, J.H. The Role of N-Linked Glycosylation in the Protection of Human and Bovine Lactoferrin against Tryptic Proteolysis. Eur. J. Biochem. 2004, 271, 678–684. [Google Scholar] [CrossRef]
- van Berkel, P.H.C.; van Veen, H.A.; Geerts, M.E.J.; de Boer, H.A.; Nuijens, J.H. Heterogeneity in Utilization of N-Glycosylation Sites Asn624 and Asn138 in Human Lactoferrin: A Study with Glycosylation-Site Mutants. Biochem. J. 1996, 319, 117–122. [Google Scholar] [CrossRef]
- Wei, Z.; Nishimura, T.; Yoshida, S. Presence of a Glycan at a Potential N-Glycosylation Site, Asn-281, of Bovine Lactoferrin. J. Dairy Sci. 2000, 83, 683–689. [Google Scholar] [CrossRef]
- Ye, X.-Y.; Nishimura, T.; Yoshida, S. Characterization of the Protein and Glycan Moieties in Different Forms of Bovine Lactoferrin. Biosci. Biotechnol. Biochem. 1997, 61, 782–786. [Google Scholar] [CrossRef]
- Legrand, D.; Mazurier, J.; Colavizza, D.; Montreuil, J.; Spik, G. Properties of the Iron-Binding Site of the N-Terminal Lobe of Human and Bovine Lactotransferrins. Importance of the Glycan Moiety and of the Non-Covalent Interactions between the N- and C-Terminal Lobes in the Stability of the Iron-Binding Site. Biochem. J. 1990, 266, 575–581. [Google Scholar]
- Li, Z.; Furmanski, P. Role of Sialic Acid Residues in Iron Binding by Human Lactoferrin-α. Chin. J. Cancer Res. 1995, 7, 79–85. [Google Scholar] [CrossRef]
- Barboza, M.; Pinzon, J.; Wickramasinghe, S.; Froehlich, J.W.; Moeller, I.; Smilowitz, J.T.; Ruhaak, L.R.; Huang, J.; Lönnerdal, B.; German, J.B.; et al. Glycosylation of Human Milk Lactoferrin Exhibits Dynamic Changes During Early Lactation Enhancing Its Role in Pathogenic Bacteria-Host Interactions. Mol. Cell. Proteom. 2012, 11, M111.015248. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.; Giansanti, F.; Boffi, A.; Ajello, M.; Valenti, P.; Chiancone, E.; Antonini, G. Ca2+ Binding to Bovine Lactoferrin Enhances Protein Stability and Influences the Release of Bacterial Lipopolysaccharide. Biochem. Cell Biol. 2002, 80, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Karabi, Z.; Moradian, F.; Kheirabadi, M. The Effect of Lactoferrin on ULK1 and ATG13 Genes Expression in Breast Cancer Cell Line MCF7 and Bioinformatics Studies of Protein Interaction between Lactoferrin and the Autophagy Initiation Complex. Cell Biochem. Biophys. 2022, 80, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Cutone, A.; Rosa, L.; Ianiro, G.; Lepanto, M.S.; Bonaccorsi di Patti, M.C.; Valenti, P.; Musci, G. Lactoferrin’s Anti-Cancer Properties: Safety, Selectivity, and Wide Range of Action. Biomolecules 2020, 10, 456. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sinha, M.; Kaushik, S.; Kaur, P.; Singh, T.P. C-Lobe of Lactoferrin: The Whole Story of the Half-Molecule. Biochem. Res. Int. 2013, 2013, 271641. [Google Scholar] [CrossRef] [PubMed]
- Di Biase, A.M.; Pietrantoni, A.; Tinari, A.; Siciliano, R.; Valenti, P.; Antonini, G.; Seganti, L.; Superti, F. Heparin-interacting Sites of Bovine Lactoferrin Are Involved in Anti-adenovirus Activity. J. Med. Virol. 2003, 69, 495–502. [Google Scholar] [CrossRef]
- Superti, F.; Agamennone, M.; Pietrantoni, A.; Ammendolia, M.G. Bovine Lactoferrin Prevents Influenza A Virus Infection by Interfering with the Fusogenic Function of Viral Hemagglutinin. Viruses 2019, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-S.; Shimazaki, K.; Tamura, T. Expression of Bovine Lactoferrin C-Lobe in Rhodococcus erythropolis and Its Purification and Characterization. Biosci. Biotechnol. Biochem. 2006, 70, 2641–2645. [Google Scholar] [CrossRef] [PubMed]
- Cao, J. Functional Divergence of the N-Lobe and C-Lobe of Transferrin Gene in Pungitius sinensis (Amur Stickleback). Animals 2022, 12, 3458. [Google Scholar] [CrossRef]
- Kim, W.-S.; Ohashi, M.; Shimazaki, K. Inhibitory Effects of Synthetic Peptides Containing Bovine Lactoferrin C-Lobe Sequence on Bacterial Growth. Korean J. Food Sci. Anim. Resour. 2016, 36, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Li, L.; Zhang, W.; Zhang, R.; Xu, Y. Heterologous Expression of Bovine Lactoferrin C-Lobe in Bacillus subtilis and Comparison of Its Antibacterial Activity with N-Lobe. Syst. Microbiol. Biomanuf. 2022, 2, 345–354. [Google Scholar] [CrossRef]
- D’Onofrio, A.; Crawford, J.M.; Stewart, E.J.; Witt, K.; Gavrish, E.; Epstein, S.; Clardy, J.; Lewis, K. Siderophores from Neighboring Organisms Promote the Growth of Uncultured Bacteria. Chem. Biol. 2010, 17, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Morgenthau, A.; Livingstone, M.; Adamiak, P.; Schryvers, A.B. The Role of Lactoferrin Binding Protein B in Mediating Protection against Human Lactoferricin 1. Biochem. Cell Biol. 2012, 90, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Ogunnariwo, J.A.; Schryvers, A.B. Characterization of a Novel Transferrin Receptor in Bovine Strains of Pasteurella multocida. J. Bacteriol. 2001, 183, 890–896. [Google Scholar] [CrossRef]
- Ekins, A.; Bahrami, F.; Sijercic, A.; Maret, D.; Niven, D.F. Haemophilus somnus Possesses Two Systems for Acquisition of Transferrin-Bound Iron. J. Bacteriol. 2004, 186, 4407–4411. [Google Scholar] [CrossRef]
- Barber, M.F.; Elde, N.C. Escape from Bacterial Iron Piracy through Rapid Evolution of Transferrin. Science 2014, 346, 1362–1366. [Google Scholar] [CrossRef]
- Ogunnariwo, J.A.; Schryvers, A.B. Correlation between the Ability of Haemophilus paragallinarum to Acquire Ovotransferrin-Bound Iron and the Expression of Ovotransferrin-Specific Receptors. Avian Dis. 1992, 36, 655. [Google Scholar] [CrossRef]
- Schryvers, A.B.; Morris, L.J. Identification and Characterization of the Human Lactoferrin-Binding Protein from Neisseria meningitidis. Infect. Immun. 1988, 56, 1144–1149. [Google Scholar] [CrossRef]
- Schryvers, A.B.; Lee, B.C. Comparative Analysis of the Transferrin and Lactoferrin Binding Proteins in the Family Neisseriaceae. Can. J. Microbiol. 1989, 35, 409–415. [Google Scholar] [CrossRef]
- Bonnah, R.A.; Yu, R.; Schryvers, A.B. Biochemical Analysis of Lactoferrin Receptors in the Neisseriaceae: Identification of a Second Bacterial Lactoferrin Receptor Protein. Microb. Pathog. 1995, 19, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Bonnah, R.A.; Schryvers, A.B. Preparation and Characterization of Neisseria meningitidis Mutants Deficient in Production of the Human Lactoferrin-Binding Proteins LbpA and LbpB. J. Bacteriol. 1998, 180, 3080–3090. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.-H.; Schryvers, A.B. Bacterial Lactoferrin Receptors: Insights from Characterizing the Moraxella bovis Receptors. Biochem. Cell Biol. 2002, 80, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Du, R.-P.; Wang, Q.; Yang, Y.-P.; Schryvers, A.B.; Chong, P.; Klein, M.H.; Loosmore, S.M. Cloning and Expression of the Moraxella catarrhalis Lactoferrin Receptor Genes. Infect. Immun. 1998, 66, 3656–3665. [Google Scholar] [CrossRef]
- Bonnah, R.A.; Wong, H.; Loosmore, S.M.; Schryvers, A.B. Characterization of Moraxella (Branhamella) catarrhalis LbpB, LbpA, and Lactoferrin Receptor Orf3 Isogenic Mutants. Infect. Immun. 1999, 67, 1517–1520. [Google Scholar] [CrossRef]
- Ostan, N.K.H.; Moraes, T.F.; Schryvers, A.B. Lactoferrin Receptors in Gram-Negative Bacteria: An Evolutionary Perspective. Biochem. Cell Biol. 2021, 99, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Morgenthau, A.; Beddek, A.; Schryvers, A.B. The Negatively Charged Regions of Lactoferrin Binding Protein B, an Adaptation against Anti-Microbial Peptides. PLoS ONE 2014, 9, e86243. [Google Scholar] [CrossRef]
- Brooks, C.L.; Arutyunova, E.; Lemieux, M.J. The Structure of Lactoferrin-Binding Protein B from Neisseria meningitidis Suggests Roles in Iron Acquisition and Neutralization of Host Defences. Acta Crystallogr. F Struct. Biol. Commun. 2014, 70, 1312–1317. [Google Scholar] [CrossRef]
- Arutyunova, E.; Brooks, C.L.; Beddek, A.; Mak, M.W.; Schryvers, A.B.; Lemieux, M.J. Crystal Structure of the N-Lobe of Lactoferrin Binding Protein B from Moraxella bovis. Biochem. Cell Biol. 2012, 90, 351–361. [Google Scholar] [CrossRef]
- Arnold, R.R.; Cole, M.F.; McGhee, J.R. A Bactericidal Effect for Human Lactoferrin. Science 1977, 197, 263–265. [Google Scholar] [CrossRef]
- Yamauchi, K.; Tomita, M.; Giehl, T.J.; Ellison, R.T. Antibacterial Activity of Lactoferrin and a Pepsin-Derived Lactoferrin Peptide Fragment. Infect. Immun. 1993, 61, 719–728. [Google Scholar] [CrossRef]
- Ellison, R.T.; Giehl, T.J. Killing of Gram-Negative Bacteria by Lactoferrin and Lysozyme. J. Clin. Investig. 1991, 88, 1080–1091. [Google Scholar] [CrossRef]
- Ellison, R.T.; Giehl, T.J.; LaForce, F.M. Damage of the Outer Membrane of Enteric Gram-Negative Bacteria by Lactoferrin and Transferrin. Infect. Immun. 1988, 56, 2774–2781. [Google Scholar] [CrossRef] [PubMed]
- Ellison, R.T.; LaForce, F.M.; Giehl, T.J.; Boose, D.S.; Dunn, B.E. Lactoferrin and Transferrin Damage of the Gram-Negative Outer Membrane Is Modulated by Ca2+ and Mg2+. J. Gen. Microbiol. 1990, 136, 1437–1446. [Google Scholar] [CrossRef] [PubMed]
- Elass-Rochard, E.; Roseanu, A.; Legrand, D.; Trif, M.; Salmon, V.; Motas, C.; Montreuil, J.; Spik, G. Lactoferrin-Lipopolysaccharide Interaction: Involvement of the 28-34 Loop Region of Human Lactoferrin in the High-Affinity Binding to Escherichia coli 055B5 Lipopolysaccharide. Biochem. J. 1995, 312, 839–845. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular Basis of Bacterial Outer Membrane Permeability Revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Appelmelk, B.J.; An, Y.Q.; Geerts, M.; Thijs, B.G.; De Boer, H.A.; MacLaren, D.M.; De Graaff, J.; Nuijens, J.H. Lactoferrin Is a Lipid A-Binding Protein. Infect. Immun. 1994, 62, 2628–2632. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pabst, K.M.; Aida, Y.; Pabst, M.J. Lipopolysaccharide-Inactivating Activity of Neutrophils Is Due to Lactoferrin. J. Leukoc. Biol. 1995, 57, 865–874. [Google Scholar] [CrossRef]
- Machnicki, M.; Zimecki, M.; Zagulski, T. Lactoferrin Regulates the Release of Tumor Necrosis Factor Alpha and Interleukin 6 in Vivo. Int. J. Exp. Pathol. 1993, 74, 433–439. [Google Scholar] [PubMed]
- van Berkel, P.H.C.; Geerts, E.J.M.; van Veen, A.H.; Mericskay, M.; De Boer, A.H.; Nuijens, H.J. N-Terminal Stretch Arg2, Arg3, Arg4 and Arg5 of Human Lactoferrin Is Essential for Binding to Heparin, Bacterial Lipopolysaccharide, Human Lysozyme and DNA. Biochem. J. 1997, 328, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Schryvers, A.B.; Morris, L.J. Identification and Characterization of the Transferrin Receptor from Neisseria meningitidis. Mol. Microbiol. 1988, 2, 281–288. [Google Scholar] [CrossRef]
- Mickelsen, P.A.; Blackman, E.; Sparling, P.F. Ability of Neisseria gonorrhoeae, Neisseria geningitidis, and Commensal Neisseria Species to Obtain Iron from Lactoferrin. Infect. Immun. 1982, 35, 915–920. [Google Scholar] [CrossRef]
- Gray-Owen, S.D.; Schyvers, A.B. Bacterial Transferrin and Lactoferrin Receptors. Trends Microbiol. 1996, 4, 185–191. [Google Scholar] [CrossRef]
- Morgenthau, A.; Pogoutse, A.; Adamiak, P.; Moraes, T.F.; Schryvers, A.B. Bacterial Receptors for Host Transferrin and Lactoferrin: Molecular Mechanisms and Role in Host–Microbe Interactions. Future Microbiol. 2013, 8, 1575–1585. [Google Scholar] [CrossRef]
- Moraes, T.F.; Yu, R.; Strynadka, N.C.J.; Schryvers, A.B. Insights into the Bacterial Transferrin Receptor: The Structure of Transferrin-Binding Protein B from Actinobacillus pleuropneumoniae. Mol. Cell 2009, 35, 523–533. [Google Scholar] [CrossRef]
- Yang, X.; Yu, R.; Calmettes, C.; Moraes, T.F.; Schryvers, A.B. Anchor Peptide of Transferrin-Binding Protein B Is Required for Interaction with Transferrin-Binding Protein A. J. Biol. Chem. 2011, 286, 45165–45173. [Google Scholar] [CrossRef] [PubMed]
- Noinaj, N.; Easley, N.C.; Oke, M.; Mizuno, N.; Gumbart, J.; Boura, E.; Steere, A.N.; Zak, O.; Aisen, P.; Tajkhorshid, E.; et al. Structural Basis for Iron Piracy by Pathogenic Neisseria. Nature 2012, 483, 53–58. [Google Scholar] [CrossRef]
- Noto, J.M.; Cornelissen, C.N. Identification of TbpA Residues Required for Transferrin-Iron Utilization by Neisseria gonorrhoeae. Infect. Immun. 2008, 76, 1960–1969. [Google Scholar] [CrossRef] [PubMed]
- Renauld-Mongénie, G.; Poncet, D.; Mignon, M.; Fraysse, S.; Chabanel, C.; Danve, B.; Krell, T.; Quentin-Millet, M.-J. Role of Transferrin Receptor from a Neisseria meningitidis TbpB Isotype II Strain in Human Transferrin Binding and Virulence. Infect. Immun. 2004, 72, 3461–3470. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.; Schryvers, A.B. Bacterial Lactoferrin-Binding Protein A Binds to Both Domains of the Human Lactoferrin C-Lobe. Microbiology 2003, 149, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- Noinaj, N.; Cornelissen, C.N.; Buchanan, S.K. Structural Insight into the Lactoferrin Receptors from Pathogenic Neisseria. J. Struct. Biol. 2013, 184, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Biswas, G.D.; Anderson, J.E.; Chen, C.-J.; Cornelissen, C.N.; Sparling, P.F. Identification and Functional Characterization of the Neisseria gonorrhoeae LbpB Gene Product. Infect. Immun. 1999, 67, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Biswas, G.D.; Sparling, P.F. Characterization of LbpA, the Structural Gene for a Lactoferrin Receptor in Neisseria gonorrhoeae. Infect. Immun. 1995, 63, 2958–2967. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, A.; Maas, A.; Tommassen, J. Identification of the IroA Gene Product of Neisseria meningitidis as a Lactoferrin Receptor. J. Bacteriol. 1994, 176, 1764–1766. [Google Scholar] [CrossRef] [PubMed]
- Prinz, T.; Meyer, M.; Pettersson, A.; Tommassen, J. Structural Characterization of the Lactoferrin Receptor from Neisseria meningitidis. J. Bacteriol. 1999, 181, 4417–4419. [Google Scholar] [CrossRef]
- Yadav, R.; Govindan, S.; Daczkowski, C.; Mesecar, A.; Chakravarthy, S.; Noinaj, N. Structural Insight into the Dual Function of LbpB in Mediating Neisserial Pathogenesis. Elife 2021, 10, e71683. [Google Scholar] [CrossRef]
- Sill, C.; Biehl, R.; Hoffmann, B.; Radulescu, A.; Appavou, M.-S.; Farago, B.; Merkel, R.; Richter, D. Structure and Domain Dynamics of Human Lactoferrin in Solution and the Influence of Fe(III)-Ion Ligand Binding. BMC Biophys. 2016, 9, 7. [Google Scholar] [CrossRef]
- Naidu, S.S.; Svensson, U.; Kishore, A.R.; Naidu, A.S. Relationship between Anti-bacterial Activity and Porin Binding of Lactoferrin in Escherichia coli and Salmonella typhimurium. Antimicrob. Agents Chemother. 1993, 37, 240–245. [Google Scholar] [CrossRef]
- Erdei, J.; Forsgren, A.; Naidu, A.S. Lactoferrin Binds to Porins OmpF and OmpC in Escherichia coli. Infect. Immun. 1994, 62, 1236–1240. [Google Scholar] [CrossRef]
- Sallmann, F.R.; Baveye-Descamps, S.; Pattus, F.; Salmon, V.; Branza, N.; Spik, G.; Legrand, D. Porins OmpC and PhoE of Escherichia coli as Specific Cell-Surface Targets of Human Lactoferrin. J. Biol. Chem. 1999, 274, 16107–16114. [Google Scholar] [CrossRef]
- Ziere, G.J.; Bijsterbosch, M.K.; van Berkel, T.J. Removal of 14 N-Terminal Amino Acids of Lactoferrin Enhances Its Affinity for Parenchymal Liver Cells and Potentiates the Inhibition of Beta- Very Low Density Lipoprotein Binding. J. Biol. Chem. 1993, 268, 27069–27075. [Google Scholar] [CrossRef] [PubMed]
- Ziere, G.J.; Kruijt, J.K.; Bijsterbosch, M.K.; van Berkel, T.J.C. Recognition of Lactoferrin and Aminopeptidase M-Modified Lactoferrin by the Liver: Involvement of Proteoglycans and the Remnant Receptor. Biochem. J. 1996, 313, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; Mazurier, J.; Elass, A.; Rochard, E.; Vergoten, G.; Maes, P.; Montreuil, J.; Spik, G. Molecular Interactions between Human Lactotransferrin and the Phytohemagglutinin-Activated Human Lymphocyte Lactotransferrin Receptor Lie in Two Loop-Containing Regions of the N-Terminal Domain I of Human Lactotransferrin. Biochemistry 1992, 31, 9243–9251. [Google Scholar] [CrossRef] [PubMed]
- Huettinger, M.; Retzek, H.; Hermann, M.; Goldenberg, H. Lactoferrin Specifically Inhibits Endocytosis of Chylomicron Remnants but Not Alpha-Macroglobulin. J. Biol. Chem. 1992, 267, 18551–18557. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; Van Berkel, P.H.C.; Salmon, V.; Van Veen, A.H.; Slomianny, M.-C.; Nuijens, H.J.; Spik, G. The N-Terminal Arg2, Arg3 and Arg4 of Human Lactoferrin Interact with Sulphated Molecules but Not with the Receptor Present on Jurkat Human Lymphoblastic T-Cells. Biochem. J. 1997, 327, 841–846. [Google Scholar] [CrossRef]
- Mann, D.M.; Romm, E.; Migliorini, M. Delineation of the Glycosaminoglycan-Binding Site in the Human Inflammatory Response Protein Lactoferrin. J. Biol. Chem. 1994, 269, 23661–23667. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, K.; Tazume, T.; Uji, K.; Tanaka, M.; Kumura, H.; Mikawa, K.; Shimo-oka, T. Properties of a Heparin-Binding Peptide Derived from Bovine Lactoferrin. J. Dairy Sci. 1998, 81, 2841–2849. [Google Scholar] [CrossRef]
- Wu, H.F.; Monroe, D.M.; Church, F.C. Characterization of the Glycosaminoglycan-Binding Region of Lactoferrin. Arch. Biochem. Biophys. 1995, 317, 85–92. [Google Scholar] [CrossRef]
- Lévy-Toledano, S.; Grelac, F.; Caen, J.P.; Maclouf, J. KRDS, a Peptide Derived from Human Lactotransferrin, Inhibits Thrombin-Induced Thromboxane Synthesis by a Cyclooxygenase-Independent Mechanism. Thromb. Haemost. 1995, 73, 857–861. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the Bactericidal Domain of Lactoferrin. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- Elass-Rochard, E.; Legrand, D.; Salmon, V.; Roseanu, A.; Trif, M.; Tobias, P.S.; Mazurier, J.; Spik, G. Lactoferrin Inhibits the Endotoxin Interaction with CD14 by Competition with the Lipopolysaccharide-Binding Protein. Infect. Immun. 1998, 66, 486–491. [Google Scholar] [CrossRef]
- Harmsen, M.C.; Swart, P.J.; Bethune, M.-P.d.; Pauwels, R.; Clercq, E.D.; The, T.B.; Meijer, D.K.F. Antiviral Effects of Plasma and Milk Proteins: Lactoferrin Shows Potent Activity against Both Human Immunodeficiency Virus and Human Cytomegalovirus Replication In Vitro. J. Infect. Dis. 1995, 172, 380–388. [Google Scholar] [CrossRef]
- Berkhout, B.; van Wamel, J.L.B.; Beljaars, L.; Meijer, D.K.F.; Visser, S.; Floris, R. Characterization of the Anti-HIV Effects of Native Lactoferrin and Other Milk Proteins and Protein-Derived Peptides. Antivir. Res. 2002, 55, 341–355. [Google Scholar] [CrossRef]
- Groot, F.; Geijtenbeek, T.B.H.; Sanders, R.W.; Baldwin, C.E.; Sanchez-Hernandez, M.; Floris, R.; van Kooyk, Y.; de Jong, E.C.; Berkhout, B. Lactoferrin Prevents Dendritic Cell-Mediated Human Immunodeficiency Virus Type 1 Transmission by Blocking the DC-SIGN--Gp120 Interaction. J. Virol. 2005, 79, 3009–3015. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Longhi, C.; Conte, M.P.; Pisani, S.; Valenti, P.; Seganti, L. Lactoferrin Inhibits Herpes Simplex Virus Type 1 Adsorption to Vero Cells. Antivir. Res. 1996, 29, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Pisani, S.; Antonini, G.; Valenti, P.; Seganti, L.; Orsi, N. Metal Complexes of Bovine Lactoferrin Inhibit in Vitro Replication of Herpes Simplex Virus Type 1 and 2. Biometals 1998, 11, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Trybala, E.; Superti, F.; Johansson, M.; Bergström, T. Inhibition of Herpes Simplex Virus Infection by Lactoferrin Is Dependent on Interference with the Virus Binding to Glycosaminoglycans. Virology 2004, 318, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Ammendolia, M.G.; Marchetti, M.; Superti, F. Bovine Lactoferrin Prevents the Entry and Intercellular Spread of Herpes Simplex Virus Type 1 in Green Monkey Kidney Cells. Antivir. Res. 2007, 76, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Ammendolia, M.G.; Superti, F. Glycosaminoglycans Are Not Indispensable for the Anti-Herpes Simplex Virus Type 2 Activity of Lactoferrin. Biochimie 2009, 91, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, W.; Ye, Q.; Zhou, Y.; Xiong, W.; He, W.; Deng, M.; Zhou, M.; Guo, X.; Chen, P.; et al. Inhibition of Epstein-Barr Virus Infection by Lactoferrin. J. Innate Immun. 2012, 4, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Superti, F.; Ammendolia, M.G.; Rossi, P.; Valenti, P.; Seganti, L. Inhibition of Poliovirus Type 1 Infection by Iron-, Manganese- and Zinc-Saturated Lactoferrin. Med. Microbiol. Immunol. 1999, 187, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Kaneko, S.; Yu, D.; Murakami, S. Hepatitis C Virus Envelope Proteins Bind Lactoferrin. J. Virol. 1997, 71, 5997–6002. [Google Scholar] [CrossRef]
- Ikeda, M.; Sugiyama, K.; Tanaka, T.; Tanaka, K.; Sekihara, H.; Shimotohno, K.; Kato, N. Lactoferrin Markedly Inhibits Hepatitis C Virus Infection in Cultured Human Hepatocytes. Biochem. Biophys. Res. Commun. 1998, 245, 549–553. [Google Scholar] [CrossRef]
- Ikeda, M.; Nozaki, A.; Sugiyama, K.; Tanaka, T.; Naganuma, A.; Tanaka, K.; Sekihara, H.; Shimotohno, K.; Saito, M.; Kato, N. Characterization of Antiviral Activity of Lactoferrin against Hepatitis C Virus Infection in Human Cultured Cells. Virus Res. 2000, 66, 51–63. [Google Scholar] [CrossRef]
- Liao, Y.; El-Fakkarany, E.; Lönnerdal, B.; Redwan, E.M. Inhibitory Effects of Native and Recombinant Full-Length Camel Lactoferrin and Its N and C Lobes on Hepatitis C Virus Infection of Huh7.5 Cells. J. Med. Microbiol. 2012, 61, 375–383. [Google Scholar] [CrossRef]
- Picard-Jean, F.; Bouchard, S.; Larivée, G.; Bisaillon, M. The Intracellular Inhibition of HCV Replication Represents a Novel Mechanism of Action by the Innate Immune Lactoferrin Protein. Antivir. Res. 2014, 111, 13–22. [Google Scholar] [CrossRef]
- Pietrantoni, A.; Di Biase, A.M.; Tinari, A.; Marchetti, M.; Valenti, P.; Seganti, L.; Superti, F. Bovine Lactoferrin Inhibits Adenovirus Infection by Interacting with Viral Structural Polypeptides. Antimicrob. Agents Chemother. 2003, 47, 2688–2691. [Google Scholar] [CrossRef]
- Persson, B.D.; Lenman, A.; Frängsmyr, L.; Schmid, M.; Ahlm, C.; Plückthun, A.; Jenssen, H.; Arnberg, N. Lactoferrin-Hexon Interactions Mediate CAR-Independent Adenovirus Infection of Human Respiratory Cells. J. Virol. 2020, 94, e00542-20. [Google Scholar] [CrossRef]
- Tanaka, T.; Nakatani, S.; Xuan, X.; Kumura, H.; Igarashi, I.; Shimazaki, K. Antiviral Activity of Lactoferrin against Canine Herpesvirus. Antivir. Res. 2003, 60, 193–199. [Google Scholar] [CrossRef]
- Ammendolia, M.G.; Pietrantoni, A.; Tinari, A.; Valenti, P.; Superti, F. Bovine Lactoferrin Inhibits Echovirus Endocytic Pathway by Interacting with Viral Structural Polypeptides. Antivir. Res. 2007, 73, 151–160. [Google Scholar] [CrossRef]
- Weng, T.; Chen, L.; Shyu, H.; Chen, S.; Wang, J.; Yu, C.; Lei, H.; Yeh, T. Lactoferrin Inhibits Enterovirus 71 Infection by Binding to VP1 Protein and Host Cells. Antivir. Res. 2005, 67, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Miotto, M.; Di Rienzo, L.; Bò, L.; Boffi, A.; Ruocco, G.; Milanetti, E. Molecular Mechanisms Behind Anti SARS-CoV-2 Action of Lactoferrin. Front. Mol. Biosci. 2021, 8, 607443. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, R.; Centi, L.; Miotto, M.; Milanetti, E.; Di Rienzo, L.; Pitea, M.; Piazza, P.; Ruocco, G.; Boffi, A.; Parisi, G. Lactoferrin Inhibition of the Complex Formation between ACE2 Receptor and SARS-CoV-2 Recognition Binding Domain. Int. J. Mol. Sci. 2022, 23, 5436. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Xu, S.; Xu, Z.; Cheng, S.; Wu, D.; Liu, H.; Du, M. Identification of Dual-Function Bovine Lactoferrin Peptides Released Using Simulated Gastrointestinal Digestion. Food Biosci. 2021, 39, 100806. [Google Scholar] [CrossRef]
- Berlutti, F.; Pantanella, F.; Natalizi, T.; Frioni, A.; Paesano, R.; Polimeni, A.; Valenti, P. Antiviral Properties of Lactoferrin—A Natural Immunity Molecule. Molecules 2011, 16, 6992–7018. [Google Scholar] [CrossRef]
- Chang, R.; Ng, T.B.; Sun, W.-Z. Lactoferrin as Potential Preventative and Adjunct Treatment for COVID-19. Int. J. Antimicrob. Agents 2020, 56, 106118. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Li, X.; Yu, Z.; Wu, S.; Ding, L.; Liu, J. Identification of Lactoferrin-Derived Peptides as Potential Inhibitors against the Main Protease of SARS-CoV-2. LWT 2022, 154, 112684. [Google Scholar] [CrossRef] [PubMed]
- Cutone, A.; Rosa, L.; Bonaccorsi di Patti, M.C.; Iacovelli, F.; Conte, M.P.; Ianiro, G.; Romeo, A.; Campione, E.; Bianchi, L.; Valenti, P.; et al. Lactoferrin Binding to SARS-CoV-2 Spike Glycoprotein Blocks Pseudoviral Entry and Relieves Iron Protein Dysregulation in Several In Vitro Models. Pharmaceutics 2022, 14, 2111. [Google Scholar] [CrossRef]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro Web Server for Protein–Protein Docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin Against SARS-CoV-2: In Vitro and In Silico Evidences. Front. Pharmacol. 2021, 12, 666600. [Google Scholar] [CrossRef]
- Milanetti, E.; Miotto, M.; Di Rienzo, L.; Monti, M.; Gosti, G.; Ruocco, G. 2D Zernike Polynomial Expansion: Finding the Protein-Protein Binding Regions. Comput. Struct. Biotechnol. J. 2021, 19, 29–36. [Google Scholar] [CrossRef]
- Milanetti, E.; Miotto, M.; Di Rienzo, L.; Nagaraj, M.; Monti, M.; Golbek, T.W.; Go-sti, G.; Roeters, S.J.; Weidner, T.; Otzen, D.E.; et al. In-Silico Evidence for a Two Receptor Based Strategy of SARS-CoV-2. Front. Mol. Biosci. 2021, 8, 690655. [Google Scholar] [CrossRef]
- Bò, L.; Miotto, M.; Di Rienzo, L.; Milanetti, E.; Ruocco, G. Exploring the Association Between Sialic Acid and SARS-CoV-2 Spike Protein through a Molecular Dynamics-Based Approach. Front. Med. Technol. 2021, 2, 614652. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.; Milanetti, E.; Frans, M.T.; Miotto, M.; Di Rienzo, L.; Baranov, M.V.; Gosti, G.; Somavarapu, A.K.; Nagaraj, M.; Golbek, T.W.; et al. Two Receptor Binding Strategy of SARS-CoV-2 Is Mediated by Both the N-Terminal and Receptor-Binding Spike Domain. J. Phys. Chem. B 2024, 128, 451–464. [Google Scholar] [CrossRef]
- Nguyen, L.; McCord, K.A.; Bui, D.T.; Bouwman, K.M.; Kitova, E.N.; Elaish, M.; Kumawat, D.; Daskhan, G.C.; Tomris, I.; Han, L.; et al. Sialic Acid-Containing Glycolipids Mediate Binding and Viral Entry of SARS-CoV-2. Nat. Chem. Biol. 2022, 18, 81–90. [Google Scholar] [CrossRef]
- Unione, L.; Moure, M.J.; Lenza, M.P.; Oyenarte, I.; Ereño-Orbea, J.; Ardá, A.; Jiménez-Barbero, J. The SARS-CoV-2 Spike Glycoprotein Directly Binds Exogeneous Sialic Acids: A NMR View. Angew. Chem. Int. Ed. 2022, 61, e202201432. [Google Scholar] [CrossRef]
- Iacovelli, F.; Costanza, G.; Romeo, A.; Cosio, T.; Lanna, C.; Bagnulo, A.; Di Maio, U.; Sbardella, A.; Gaziano, R.; Grelli, S.; et al. Interaction of Pelargonium Sidoides Compounds with Lactoferrin and SARS-CoV-2: Insights from Molecular Simulations. Int. J. Environ. Res. Public Health 2022, 19, 5254. [Google Scholar] [CrossRef] [PubMed]
- Furlund, C.B.; Ulleberg, E.K.; Devold, T.G.; Flengsrud, R.; Jacobsen, M.; Sekse, C.; Holm, H.; Vegarud, G.E. Identification of Lactoferrin Peptides Generated by Digestion with Human Gastrointestinal Enzymes. J. Dairy Sci. 2013, 96, 75–88. [Google Scholar] [CrossRef]
- Kuwata, H.; Yip, T.-T.; Tomita, M.; Hutchens, T.W. Direct Evidence of the Generation in Human Stomach of an Antimicrobial Peptide Domain (Lactoferricin) from Ingested Lactoferrin. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1998, 1429, 129–141. [Google Scholar] [CrossRef]
- Kuwata, H.; Yamauchi, K.; Teraguchi, S.; Hayasawa, H.; Ushida, Y.; Shimokawa, Y.; Toida, T. Functional Fragments of Ingested Lactoferrin Are Resistant to Proteolytic Degradation in the Gastrointestinal Tract of Adult Rats. J. Nutr. 2001, 131, 2121–2127. [Google Scholar] [CrossRef]
- Tomita, M.; Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K. Potent Antibacterial Peptides Generated by Pepsin Digestion of Bovine Lactoferrin. J. Dairy Sci. 1991, 74, 4137–4142. [Google Scholar] [CrossRef]
- Umeyama, M.; Kira, A.; Nishimura, K.; Naito, A. Interactions of Bovine Lactoferricin with Acidic Phospholipid Bilayers and Its Antimicrobial Activity as Studied by Solid-State NMR. Biochim. Biophys. Acta (BBA)-Biomembr. 2006, 1758, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Diarra, M.S. Ultrastructural and Cytochemical Study of Cell Wall Modification by Lactoferrin, Lactoferricin and Penicillin G against Staphylococcus aureus. J. Electron. Microsc. 2003, 52, 207–215. [Google Scholar] [CrossRef]
- Farnaud, S.; Spiller, C.; Moriarty, L.C.; Patel, A.; Gant, V.; Odell, E.W.; Evans, R.W. Interactions of Lactoferricin-Derived Peptides with LPS and Anti-microbial Activity. FEMS Microbiol. Lett. 2004, 233, 193–199. [Google Scholar] [CrossRef]
- Hunter, H.N.; Demcoe, A.R.; Jenssen, H.; Gutteberg, T.J.; Vogel, H.J. Human Lactoferricin Is Partially Folded in Aqueous Solution and Is Better Stabilized in a Membrane Mimetic Solvent. Antimicrob. Agents Chemother. 2005, 49, 3387–3395. [Google Scholar] [CrossRef]
- Gruden, Š.; Poklar Ulrih, N. Diverse Mechanisms of Antimicrobial Activities of Lactoferrins, Lactoferricins, and Other Lactoferrin-Derived Peptides. Int. J. Mol. Sci. 2021, 22, 11264. [Google Scholar] [CrossRef]
- Chapple, D.S.; Hussain, R.; Joannou, C.L.; Hancock, R.E.W.; Odell, E.; Evans, R.W.; Siligardi, G. Structure and Association of Human Lactoferrin Peptides with Escherichia coli Lipopolysaccharide. Antimicrob. Agents Chemother. 2004, 48, 2190–2198. [Google Scholar] [CrossRef] [PubMed]
- Farnaud, S.; Patel, A.; Odell, E.W.; Evans, R.W. Variation in Antimicrobial Activity of Lactoferricin-Derived Peptides Explained by Structure Modelling. FEMS Micro-Biol. Lett. 2004, 238, 221–226. [Google Scholar] [CrossRef]
- Pei, J.; Xiong, L.; Bao, P.; Chu, M.; Yan, P.; Guo, X. Secondary Structural Transformation of Bovine Lactoferricin Affects Its Antibacterial Activity. Probiotics Antimicrob. Proteins 2021, 13, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Hilde Ulvatne, L.H.V. Bactericidal Kinetics of 3 Lactoferricins Against Staphylococcus aureus and Escherichia coli. Scand. J. Infect. Dis. 2001, 33, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Han, F.; Xie, Y.; Wang, Y. Comparative Antimicrobial Activity and Mechanism of Action of Bovine Lactoferricin-Derived Synthetic Peptides. BioMetals 2011, 24, 1069–1078. [Google Scholar] [CrossRef]
- Haug, B.E.; Svendsen, J.S. The Role of Tryptophan in the Antibacterial Activity of a 15-residue Bovine Lactoferricin Peptide. J. Pept. Sci. 2001, 7, 190–196. [Google Scholar] [CrossRef]
- Vorland, L.H.; Ulvatne, H.; Andersen, J.; Haukland, H.H.; Rekdal, O.; Svendsen, J.S.; Gutteberg, T.J. Antibacterial Effects of Lactoferricin B. Scand. J. Infect. Dis. 1999, 31, 179–184. [Google Scholar] [CrossRef]
- van der Kraan, M.I.A.; Groenink, J.; Nazmi, K.; Veerman, E.C.I.; Bolscher, J.G.M.; Nieuw Amerongen, A. V Lactoferrampin: A Novel Antimicrobial Peptide in the N1-Domain of Bovine Lactoferrin. Peptides 2004, 25, 177–183. [Google Scholar] [CrossRef]
- Bolscher, J.G.M.; van der Kraan, M.I.A.; Nazmi, K.; Kalay, H.; Grün, C.H.; van’t Hof, W.; Veerman, E.C.I.; Nieuw Amerongen, A.V. A One-Enzyme Strategy to Release an Antimicrobial Peptide from the LFampin-Domain of Bovine Lactoferrin. Peptides 2006, 27, 1–9. [Google Scholar] [CrossRef]
- Haney, E.; Nazmi, K.; Lau, F.; Bolscher, J.G.M.; Vogel, H. Novel Lactoferrampin Antimicrobial Peptides Derived from Human Lactoferrin. Biochimie 2009, 91, 141–154. [Google Scholar] [CrossRef]
- Dijkshoorn, L.; Brouwer, C.P.J.M.; Bogaards, S.J.P.; Nemec, A.; van den Broek, P.J.; Nibbering, P.H. The Synthetic N-Terminal Peptide of Human Lactoferrin, HLF(1-11), Is Highly Effective against Experimental Infection Caused by Multi-drug-Resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2004, 48, 4919–4921. [Google Scholar] [CrossRef]
- Faber, C.; Stallmann, H.P.; Lyaruu, D.M.; Joosten, U.; von Eiff, C.; van Nieuw Amerongen, A.; Wuisman, P.I.J.M. Comparable Efficacies of the Antimicrobial Peptide Human Lactoferrin 1-11 and Gentamicin in a Chronic Methicillin-Resistant Staphylococcus aureus Osteomyelitis Model. Antimicrob. Agents Chemother. 2005, 49, 2438–2444. [Google Scholar] [CrossRef]
- Denardi, L.B.; de Arruda Trindade, P.; Weiblen, C.; Ianiski, L.B.; Stibbe, P.C.; Pin-to, S.C.; Santurio, J.M. In Vitro Activity of the Antimicrobial Peptides H-Lf1-11, MSI-78, LL-37, Fengycin 2B, and Magainin-2 against Clinically Important Bacteria. Braz. J. Microbiol. 2022, 53, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Lupetti, A.; Brouwer, C.P.J.M.; Bogaards, S.J.P.; Welling, M.M.; de Heer, E.; Campa, M.; van Dissel, J.T.; Friesen, R.H.E.; Nibbering, P.H. Human Lactoferrin-Derived Peptide’s Antifungal Activities against Disseminated Candida albicans Infection. J. Infect. Dis. 2007, 196, 1416–1424. [Google Scholar] [CrossRef] [PubMed]
- Lupetti, A.; Paulusma-Annema, A.; Welling, M.M.; Dogterom-Ballering, H.; Brouwer, C.P.J.M.; Senesi, S.; van Dissel, J.T.; Nibbering, P.H. Synergistic Activity of the N-Terminal Peptide of Human Lactoferrin and Fluconazole against Candida Species. Antimicrob. Agents Chemother. 2003, 47, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Lupetti, A.; Paulusma-Annema, A.; Welling, M.M.; Senesi, S.; van Dissel, J.T.; Nibbering, P.H. Candidacidal Activities of Human Lactoferrin Peptides Derived from the N Terminus. Antimicrob. Agents Chemother. 2000, 44, 3257–3263. [Google Scholar] [CrossRef] [PubMed]
- Nibbering, P.H.; Ravensbergen, E.; Welling, M.M.; van Berkel, L.A.; van Berkel, P.H.; Pauwels, E.K.; Nuijens, J.H. Human Lactoferrin and Peptides Derived from Its N Terminus Are Highly Effective against Infections with Antibiotic-Resistant Bacteria. Infect. Immun. 2001, 69, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Stallmann, H.P. Osteomyelitis Prevention in Rabbits Using Antimicrobial Peptide HLF1-11- or Gentamicin-Containing Calcium Phosphate Cement. J. Anti-Microb. Chemother. 2004, 54, 472–476. [Google Scholar] [CrossRef] [PubMed]
- van der Does, A.M.; Bogaards, S.J.P.; Ravensbergen, B.; Beekhuizen, H.; van Dissel, J.T.; Nibbering, P.H. Antimicrobial Peptide HLF1-11 Directs Granulocyte-Macrophage Colony-Stimulating Factor-Driven Monocyte Differentiation toward Macrophages with Enhanced Recognition and Clearance of Pathogens. Antimicrob. Agents Chemother. 2010, 54, 811–816. [Google Scholar] [CrossRef]
- van der Does, A.M.; Hensbergen, P.J.; Bogaards, S.J.; Cansoy, M.; Deelder, A.M.; van Leeuwen, H.C.; Drijfhout, J.W.; van Dissel, J.T.; Nibbering, P.H. The Human Lactoferrin-Derived Peptide HLF1-11 Exerts Immunomodulatory Effects by Specific Inhibition of Myeloperoxidase Activity. J. Immunol. 2012, 188, 5012–5019. [Google Scholar] [CrossRef]
- Sinha, M.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Antimicrobial Lactoferrin Peptides: The Hidden Players in the Protective Function of a Multifunctional Protein. Int. J. Pept. 2013, 2013, 1–12. [Google Scholar] [CrossRef]
- Dionysius, D.A.; Milne, J.M. Antibacterial Peptides of Bovine Lactoferrin: Purification and Characterization. J. Dairy Sci. 1997, 80, 667–674. [Google Scholar] [CrossRef]
- Recio, I.; Visser, S. Two Ion-Exchange Chromatographic Methods for the Isolation of Antibacterial Peptides from Lactoferrin. J. Chromatogr. A 1999, 831, 191–201. [Google Scholar] [CrossRef]
- Kim, W.-S.; Kim, P.-H.; Shimazaki, K.-I. Sensitivity of Pseudomonas syringae to Bovine Lactoferrin Hydrolysates and Identification of a Novel Inhibitory Peptide. Korean J. Food Sci. Anim. Resour. 2016, 36, 487–493. [Google Scholar] [CrossRef][Green Version]
- Lizzi, A.R.; Carnicelli, V.; Clarkson, M.M.; Nazzicone, C.; Segatore, B.; Celenza, G.; Aschi, M.; Dolo, V.; Strom, R.; Amicosante, G. Bovine Lactoferrin and Its Tryptic Peptides: Antibacterial Activity against Different Species. Appl. Biochem. Microbiol. 2016, 52, 435–440. [Google Scholar] [CrossRef]
- Rastogi, N.; Nagpal, N.; Alam, H.; Pandey, S.; Gautam, L.; Sinha, M.; Shin, K.; Manzoor, N.; Virdi, J.S.; Kaur, P.; et al. Preparation and Antimicrobial Action of Three Tryptic Digested Functional Molecules of Bovine Lactoferrin. PLoS ONE 2014, 9, e90011. [Google Scholar] [CrossRef]
- Hoek, K.S.; Milne, J.M.; Grieve, P.A.; Dionysius, D.A.; Smith, R. Antibacterial Activity in Bovine Lactoferrin-Derived Peptides. Antimicrob. Agents Chemother. 1997, 41, 54–59. [Google Scholar] [CrossRef] [PubMed]
- León-Sicairos, N.; Canizalez Roman, A.; De La Garza, M.; Reyes Lopez, M.; Zazueta-Beltran, J.; Nazmi, K.; Gomez-Gil, B.; Bolscher, J.G.M. Bactericidal Effect of Lactoferrin and Lactoferrin Chimera against Halophilic Vibrio parahaemolyticus. Biochimie 2009, 91, 133–140. [Google Scholar] [CrossRef] [PubMed]
- León-Sicairos, N.; Angulo-Zamudio, U.A.; Vidal, J.E.; López-Torres, C.A.; Bolscher, J.G.M.; Nazmi, K.; Reyes-Cortes, R.; Reyes-López, M.; de la Garza, M.; Canizalez-Román, A. Bactericidal Effect of Bovine Lactoferrin and Synthetic Peptide Lactoferrin Chimera in Streptococcus pneumoniae and the Decrease in LuxS Gene Expression by Lactoferrin. BioMetals 2014, 27, 969–980. [Google Scholar] [CrossRef]
- Sijbrandij, T.; Ligtenberg, A.J.; Nazmi, K.; van den Keijbus, P.A.M.; Veerman, E.C.I.; Bolscher, J.G.M.; Bikker, F.J. LFchimera protects HeLa cells from invasion by Yersinia spp. in vitro. BioMetals 2018, 31, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Puknun, A.; Bolscher, J.G.M.; Nazmi, K.; Veerman, E.C.I.; Tungpradabkul, S.; Wongratanacheewin, S.; Kanthawong, S.; Taweechaisupapong, S. A Heterodimer Comprised of Two Bovine Lactoferrin Antimicrobial Peptides Exhibits Powerful Bactericidal Activity against Burkholderia pseudomallei. World J. Microbiol. Biotechnol. 2013, 29, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Nazmi, K.; Bolscher, J.G.M.; Vogel, H.J. Structural and Biophysical Characterization of an Antimicrobial Peptide Chimera Comprised of Lactoferricin and Lactoferrampin. Biochim. Biophys. Acta (BBA)-Biomembr. 2012, 1818, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Bolscher, J.G.M.; Adao, R.; Nazmi, K.; Vandenkeybus, P.; Vanthof, W.; Nieuwamerongen, A.; Bastos, M.; Veerman, E. Bactericidal Activity of LFchimera Is Stronger and Less Sensitive to Ionic Strength than Its Constituent Lactoferricin and Lactoferrampin Peptides. Biochimie 2009, 91, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.V.; Zakahrova, E.T.; Kostevich, V.A.; Samygina, V.R.; Vasilyev, V.B. Lactoferrin, Myeloperoxidase, and Ceruloplasmin: Complementary Gearwheels Cranking Physiological and Pathological Processes. BioMetals 2014, 27, 815–828. [Google Scholar] [CrossRef]
- Sokolov, A.V.; Pulina, M.O.; Ageeva, K.V.; Ayrapetov, M.I.; Berlov, M.N.; Volgin, G.N.; Markov, A.G.; Yablonsky, P.K.; Kolodkin, N.I.; Zakharova, E.T.; et al. Interaction of Ceruloplasmin, Lactoferrin, and Myeloperoxidase. Biochemistry 2007, 72, 409–415. [Google Scholar] [CrossRef]
- Sokolov, A.V.; Ageeva, K.V.; Pulina, M.O.; Zakharova, E.T.; Vasilyev, V.B. Effect of Lactoferrin on Oxidative Features of Ceruloplasmin. BioMetals 2009, 22, 521–529. [Google Scholar] [CrossRef]
- Zakharova, E.T.; Shavlovski, M.M.; Bass, M.G.; Gridasova, A.A.; Pulina, M.O.; De Filippis, V.; Beltramini, M.; Di Muro, P.; Salvato, B.; Fontana, A.; et al. Interaction of Lactoferrin with Ceruloplasmin. Arch. Biochem. Biophys. 2000, 374, 222–228. [Google Scholar] [CrossRef]
- Pulina, M.O.; Zakharova, E.T.; Sokolov, A.V.; Shavlovski, M.M.; Bass, M.G.; Solovyov, K.V.; Kokryakov, V.N.; Vasilyev, V.B. Studies of the Ceruloplasmin-Lactoferrin Complex. Biochem. Cell Biol. 2002, 80, 35–39. [Google Scholar] [CrossRef]
- Barinov, N.A.; Vlasova, I.I.; Sokolov, A.V.; Kostevich, V.A.; Dubrovin, E.V.; Klinov, D.V. High-Resolution Atomic Force Microscopy Visualization of Metallo-proteins and Their Complexes. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 2862–2868. [Google Scholar] [CrossRef]
- Sabatucci, A.; Vachette, P.; Vasilyev, V.B.; Beltramini, M.; Sokolov, A.; Pulina, M.; Salvato, B.; Angelucci, C.B.; Maccarrone, M.; Cozzani, I.; et al. Structural Characterization of the Ceruloplasmin: Lactoferrin Complex in Solution. J. Mol. Biol. 2007, 371, 1038–1046. [Google Scholar] [CrossRef]
- Sokolov, A.V.; Pulina, M.O.; Zakharova, E.T.; Susorova, A.S.; Runova, O.L.; Ko-lodkin, N.I.; Vasilyev, V.B. Identification and Isolation from Breast Milk of Ceruloplasmin-Lactoferrin Complex. Biochemistry 2006, 71, 160–166. [Google Scholar] [CrossRef]
- Sokolov, A.V.; Voynova, I.V.; Kostevich, V.A.; Vlasenko, A.Y.; Zakharova, E.T.; Vasilyev, V.B. Comparison of Interaction between Ceruloplasmin and Lactoferrin/Transferrin: To Bind or Not to Bind. Biochemistry 2017, 82, 1073–1078. [Google Scholar] [CrossRef]
- Sakajiri, T.; Nakatsuji, M.; Teraoka, Y.; Furuta, K.; Ikuta, K.; Shibusa, K.; Sugano, E.; Tomita, H.; Inui, T.; Yamamura, T. Zinc Mediates the Interaction between Ceruloplasmin and Apo-Transferrin for the Efficient Transfer of Fe(III) Ions. Metallomics 2021, 13, mfab065. [Google Scholar] [CrossRef]
- Suzuki, Y.A.; Shin, K.; Lönnerdal, B. Molecular Cloning and Functional Expression of a Human Intestinal Lactoferrin Receptor. Biochemistry 2001, 40, 15771–15779. [Google Scholar] [CrossRef]
- Suzuki, Y.A.; Wong, H.; Ashida, K.-Y.; Schryvers, A.B.; Lönnerdal, B. The N1 Domain of Human Lactoferrin Is Required for Internalization by Caco-2 Cells and Targeting to the Nucleus. Biochemistry 2008, 47, 10915–10920. [Google Scholar] [CrossRef]
- Mancinelli, R.; Olivero, F.; Carpino, G.; Overi, D.; Rosa, L.; Lepanto, M.S.; Cutone, A.; Franchitto, A.; Alpini, G.; Onori, P.; et al. Role of Lactoferrin and Its Receptors on Biliary Epithelium. BioMetals 2018, 31, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Wesener, D.A.; Wangkanont, K.; McBride, R.; Song, X.; Kraft, M.B.; Hodges, H.L.; Zarling, L.C.; Splain, R.A.; Smith, D.F.; Cummings, R.D.; et al. Recognition of Microbial Glycans by Human Intelectin-1. Nat. Struct. Mol. Biol. 2015, 22, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Takayama, Y.; Aoki, R.; Uchida, R.; Tajima, A.; Aoki-Yoshida, A. Role of CXC Chemokine Receptor Type 4 as a Lactoferrin Receptor. Biochem. Cell Biol. 2017, 95, 57–63. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lactoferrin | Target Virus | Target of Exposure | Method of Exposure | Antiviral Activity | Reference |
|---|---|---|---|---|---|
| hLf/bLf | HIV-1 | MT4 cells |
| Interference with viral adsorption and proliferation | [96] |
| bLf/bLfcin | HIV-1 | SupT1 T/U87-CD4 cells |
| Blocking of HIV-1 variants that use CXCR4 and CCR5 coreceptors | [97] |
| bLf | HIV-1 | Dendritic cells |
| Inhibition of DC-mediated capture and transmission of HIV-1 and bLf-resistant HIV-1 through binding observed between Lf and DC-SIGN receptors | [98] |
| hLf/bLf | HCMV | Human fetal lung fibroblast cells |
| Receptor-mediated binding observed to the target cell membrane | [96] |
| hLf/bLf | HSV-1 | Vero cells |
| Inhibition of viral adsorption by blocking cellular receptors and glycans (LDL and HS) and/or viral attachment proteins (Lf in both apo- and holo-form) | [99] |
| bLf | HSV-1 HSV-2 | Vero cells |
|
| [100] |
| hLf/bLf | HSV-1 HSV-2 |
|
|
| [101] |
| bLf | HSV-1 | GMK cells |
|
| [102] |
| hLf/bLf | HSV-2 | GMK cells | Staining with crystal violet |
| [103] |
| hLf | EBV | Primary B cells |
| Direct binding of Lf to CD21 cell receptors | [104] |
| hLf/bLf | Poliovirus | Vero cells | Indirect immunofluorescence |
| [105] |
| hLf/bLf | HCV | HepG2 cells |
| Binding of Lf to viral envelope proteins E1/E2 | [106] |
| hLf/bLf | HCV | PH5CH8 cells |
| Prevention of infection of cells in presence of virus preincubated with Lf | [107] |
| bLf | HCV | MT-2C T cells | Nested RT-PCR |
| [108] |
| rcLf/ncLf | HCV | Huh7.5 cells |
|
| [109] |
| hLf | HCV | Huh-7 cells |
| Inhibition of viral ATPase/Helicase NS3 protein mediated by direct interaction between Lf and allosteric binding site on NS3 | [110] |
| bLf | HAdV |
|
| Binding of Lf to viral particle and external III and IIIa structural polypeptides | [111] |
| hLf/hLfcin | HAdV | A549 cells |
| Binding of Lfcin to hexon protein of viral capsid | [112] |
| hLf/bLf | CHV | MDCK cells |
|
| [113] |
| bLf | Echovirus | GMK cells |
|
| [114] |
| bLf | EV71 | RD and SK-N-SH cells | ELISA assay and indirect fluorescent stain |
| [115] |
| bLf | H1N1 Influenza A | MDCK cells |
| Lactoferrin binds to hemagglutinin and fusion peptide | [32] |
| hLf | SARS-CoV-2 | RBD region/ACE2 receptor | 2D Zernike polynomial expansion | Computational definition of regions of Lf with high affinity towards sialic acid and RBD | [116] |
| hLf | SARS-CoV-2 | RBD region/ACE2 receptor |
| Binding of Lf with ACE2 and observed inhibiting effects in viral interaction with receptor | [117] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piacentini, R.; Boffi, A.; Milanetti, E. Lactoferrins in Their Interactions with Molecular Targets: A Structure-Based Overview. Pharmaceuticals 2024, 17, 398. https://doi.org/10.3390/ph17030398
Piacentini R, Boffi A, Milanetti E. Lactoferrins in Their Interactions with Molecular Targets: A Structure-Based Overview. Pharmaceuticals. 2024; 17(3):398. https://doi.org/10.3390/ph17030398
Chicago/Turabian StylePiacentini, Roberta, Alberto Boffi, and Edoardo Milanetti. 2024. "Lactoferrins in Their Interactions with Molecular Targets: A Structure-Based Overview" Pharmaceuticals 17, no. 3: 398. https://doi.org/10.3390/ph17030398
APA StylePiacentini, R., Boffi, A., & Milanetti, E. (2024). Lactoferrins in Their Interactions with Molecular Targets: A Structure-Based Overview. Pharmaceuticals, 17(3), 398. https://doi.org/10.3390/ph17030398