

Novel Deep Sea Isoindole Alkaloid FGFC1 Exhibits Its Fibrinolytic Effects by Inhibiting Thrombin-Activatable Fibrinolysis Inhibitor
 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Regulatory Effects on TAFI by FGFC1
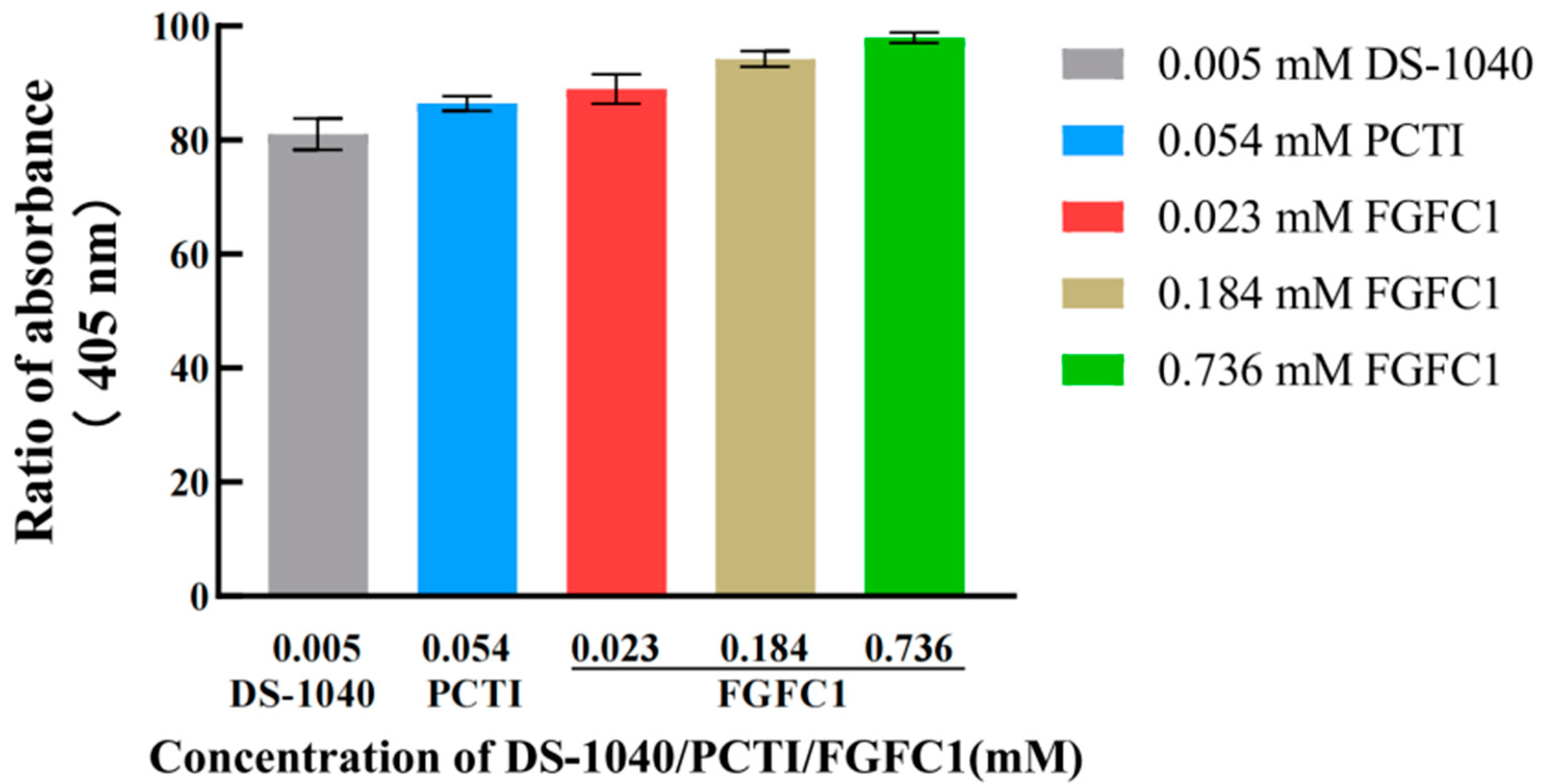
2.1.1. FGFC1 Inhibited TAFI Activation
2.1.2. FGFC1 Inhibited TAFIa–Fibrin Combination and Initiated Its Proteolysis
2.2. Inhibitory Effect of FGFC1 on TAFI-Mediating Coagulation Ex Vivo and In Vitro
2.2.1. FGFC1 Inhibited TAFI-Mediating Coagulation in the hPPP
2.2.2. FGFC1 Exhibited Little Influence on TM in the hPPP
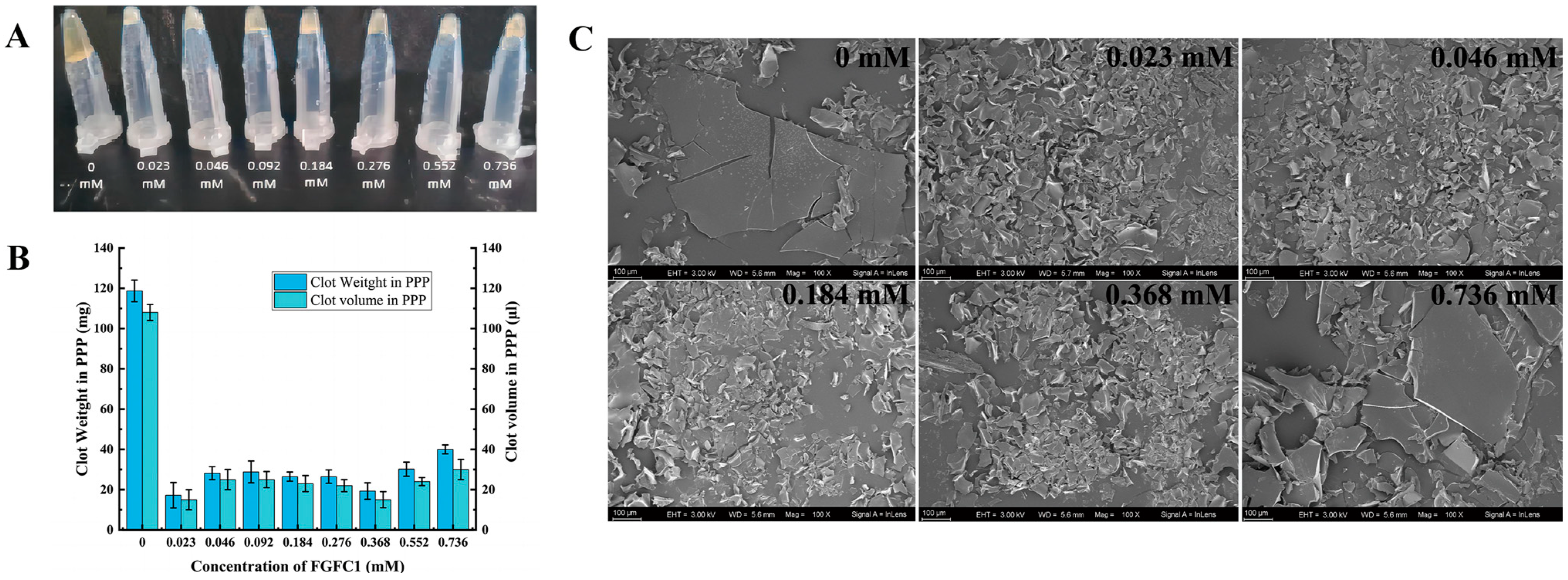
2.3. FGFC1 Reduced Coagulum Level and Obstacled Platelet Anchoring in hPPP
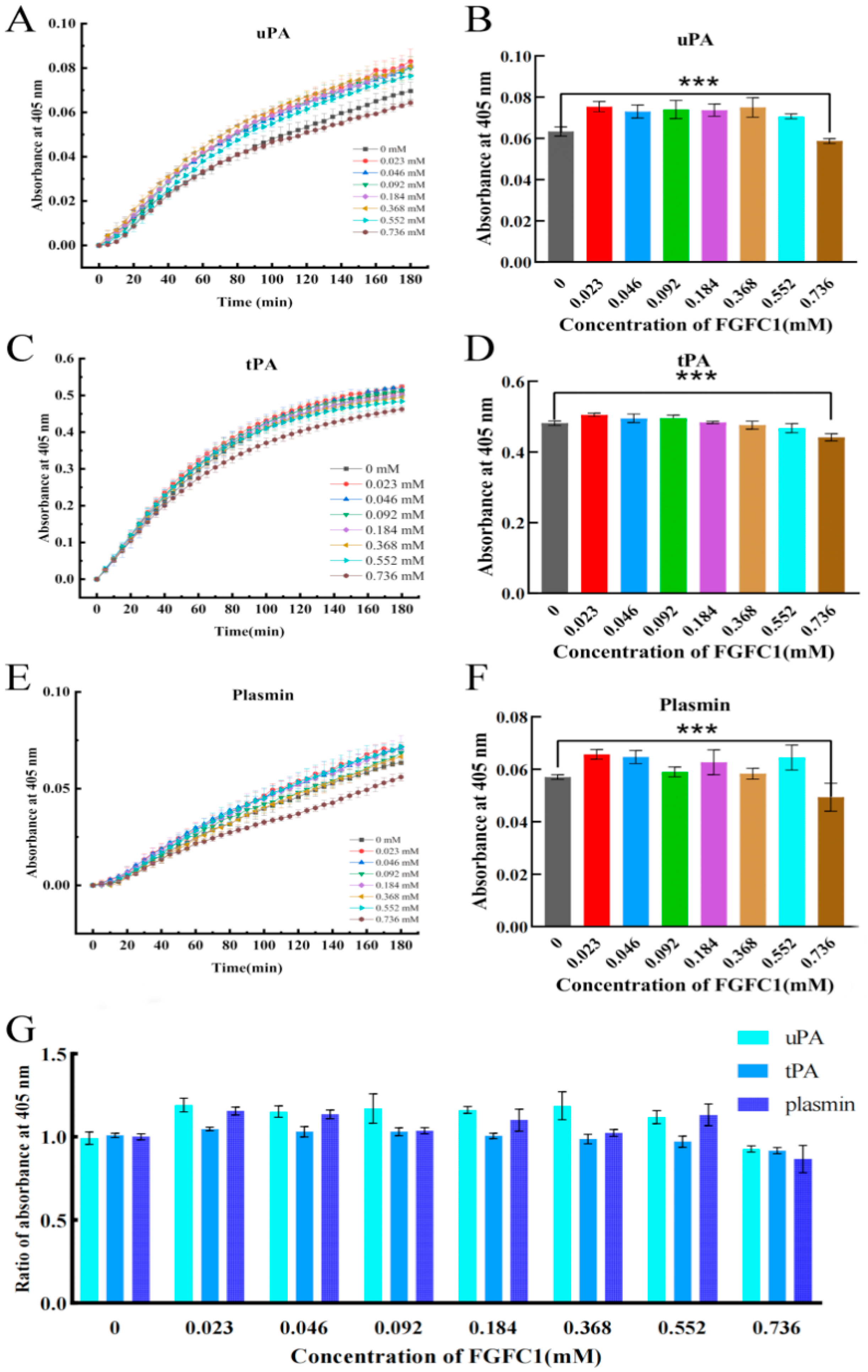
2.4. FGFC1 Facilitated TAFI-Mediating Fibrinolysis Ex Vivo
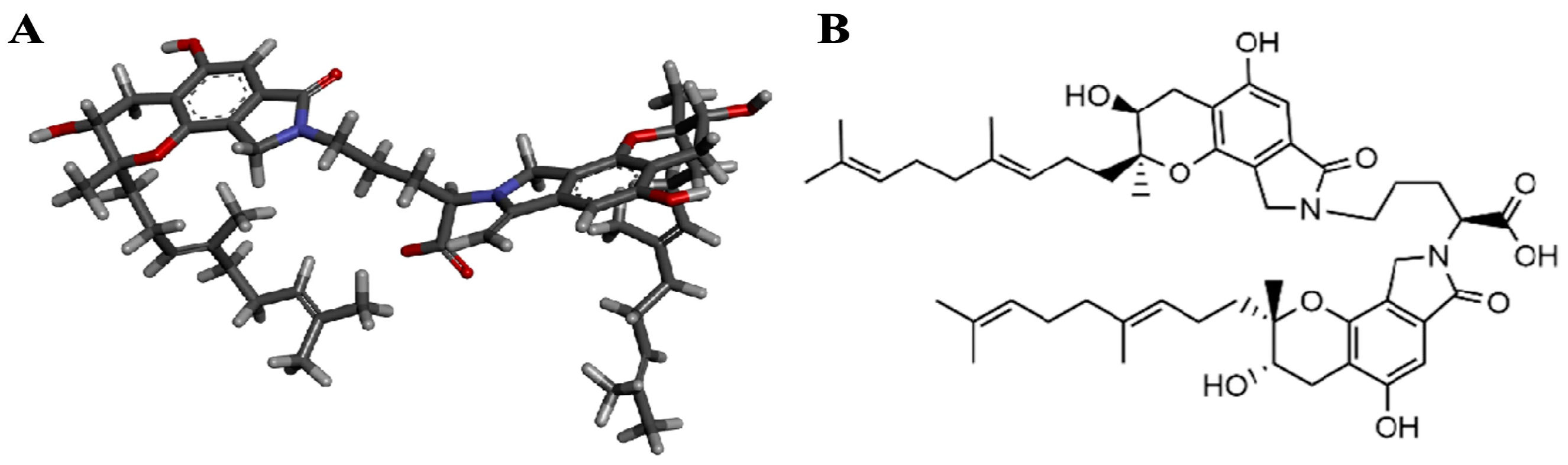
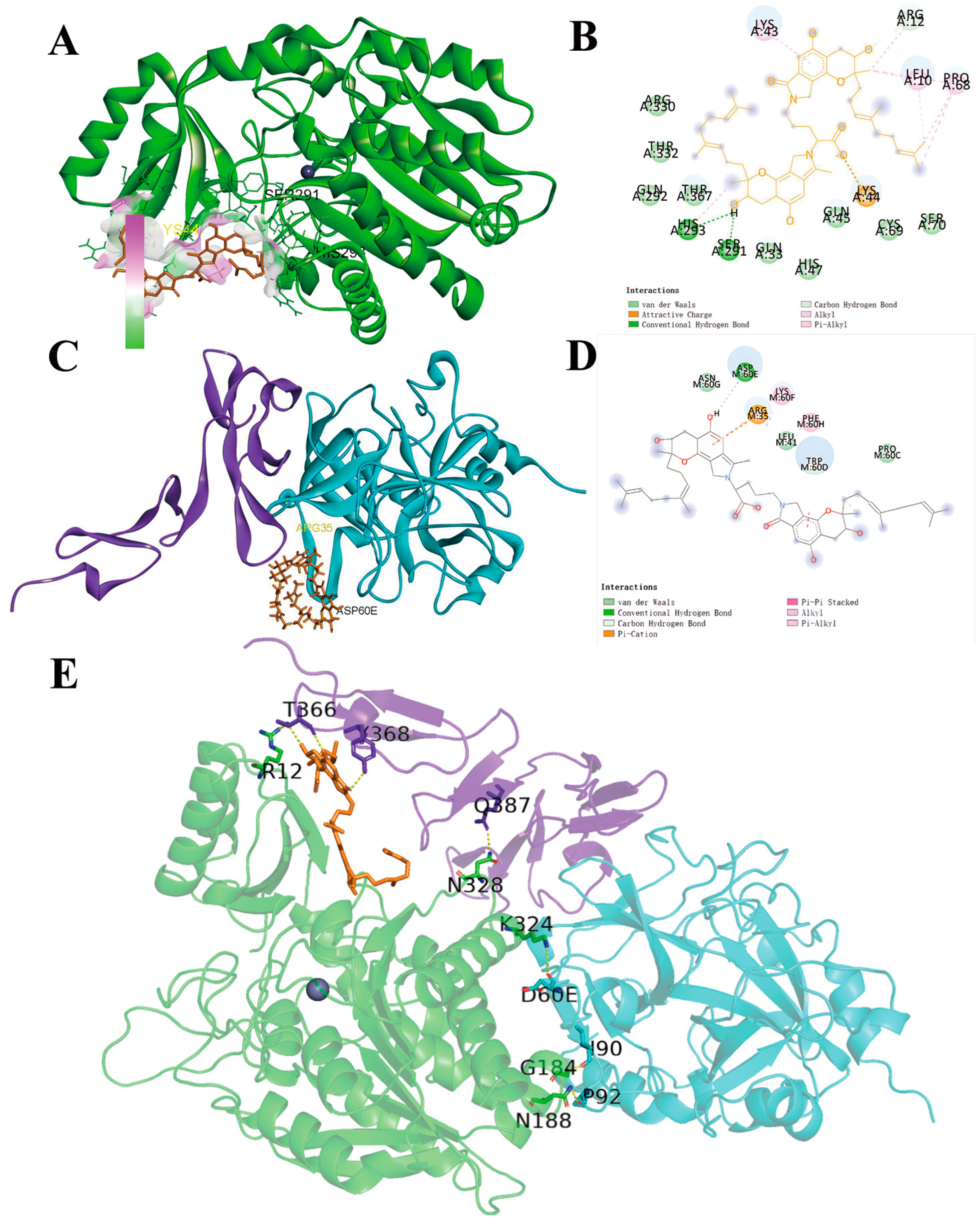
2.5. Molecular Docking of FGFC1 to TAFI
3. Discussion
4. Materials and Methods
4.1. The Construction of the hPPP-Based Ex Vivo System
4.2. Investigation of FGFC1’s Regulatory Effects on TAFI
4.3. ELISA Assays for TM Detection
4.4. Investigation on the Anti-Coagulation Effect of FGFC1
4.5. Investigation on the Fibrinolytic Effect of FGFC1
4.6. Molecular Docking
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TAFI | Thrombin-activated fibrinolysis inhibitor |
| TM | Thrombomodulin |
| FGFC1 | Fungi fibrinolytic compound 1 |
| hPPP | Human platelet-poor plasma |
| TAFIa | Activated TAFI |
| PC | Protein C |
| hCMEC/D3 | Human cerebral microvessel endothelial cells, Clone D |
| uPA | Uridylyl phosphate adenosine |
| tPA | Tissue plasminogen activator |
References
- Shah, N.B.; Sharedalal, P.; Shafi, I.; Tang, A.; Zhao, H.; Lakhter, V.; Kolluri, R.; Rao, A.K.; Bashir, R. Prevalence and outcomes of heparin-induced thrombocytopenia in hospitalized patients with venous thromboembolic disease: Insight from national inpatient sample. J. Vasc. Surg. Venous Lymphat. Disord. 2023, 11, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Michels, A.; Lillicrap, D.; Yacob, M. Role of von Willebrand factor in venous thromboembolic disease. JVS-Vasc. Sci. 2022, 3, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Nazir, S.A.; Ganeshan, A.; Nazir, S.; Uberoi, R. Endovascular Treatment Options in the Management of Lower Limb Deep Venous Thrombosis. Cardiovasc. Interv. Radiol. 2009, 32, 861–876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Lu, B.; Gao, Y.; Zhang, Y.; Li, Y.; Niu, H.; Fan, L.; Pang, Z.; Qiao, Y. Functional and binding studies of gallic acid showing platelet aggregation inhibitory effect as a thrombin inhibitor. Chin. Herb. Med. 2022, 14, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, S.M.; Velde, A.C.V.d.; Luque, D.E.; Cardozo, C.M.; Kraemer, S.; Pereira, M.d.C.G.; Gay, C.C. Platelet aggregation inhibitors from Bothrops alternatus snake venom. Toxicon 2023, 223, 107014. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Mikhalina, G.; Satoskar, A.A.; Biederman, L.; Nadasdy, T.; Rovin, B.; Parikh, S.; Brodsky, S.V. Clinical Outcomes in Patients with Biopsy Proven Anticoagulant-Related Nephropathy. Kidney Int. Rep. 2023, 8, 1445–1448. [Google Scholar] [CrossRef]
- Ren, Y.; Wu, H.; Lai, F.; Yang, M.; Li, X.; Tang, Y. Isolation and identification of a novel anticoagulant peptide from enzymatic hydrolysates of scorpion (Buthus martensii Karsch) protein. Food Res. Int. 2014, 64, 931–938. [Google Scholar] [CrossRef]
- Brouwers, K.; Kruit, A.S.; Hummelink, S.; Ulrich, D.J.O. Management of free flap salvage using thrombolytic drugs: A systematic review. J. Plast. Reconstr. Aesthetic Surg. 2020, 73, 1806–1814. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Rai, S.K.; Thakur, R.; Chattopadhyay, P.; Kar, S.K. Bafibrinase: A non-toxic, non-hemorrhagic, direct-acting fibrinolytic serine protease from Bacillus sp. strain AS-S20-I exhibits in vivo anticoagulant activity and thrombolytic potency. Biochimie 2012, 94, 1300–1308. [Google Scholar] [CrossRef]
- Austin, C.A.; Fisher, L.M. Isolation and characterization of a human cDNA clone encoding a novel DNA topoisomerase II homologue from HeLa cells. FEBS Lett. 1990, 266, 115–117. [Google Scholar] [CrossRef]
- Hendriks, D.; Wang, W.; van Sande, M.; Scharpe, S. Human serum carboxypeptidase U: A new kininase? Agents Actions Suppl. 1992, 38 Pt 1, 407–413. [Google Scholar] [PubMed]
- Hendriks, D.; Scharpé, S.; Sande, M.; Lommaert, M.P. Characterisation of a Carboxypeptidase in Human Serum Distinct from Carboxypeptidase N. Clin. Chem. Lab. Med. 2009, 27, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Bouma, B.N.; Meijers, J.C.M. Thrombin-activatable fibrinolysis inhibitor (TAFI, plasma procarboxypeptidase B, procarboxypeptidase R, procarboxypeptidase U). J. Thromb. Haemost. JTH 2003, 1, 1566–1574. [Google Scholar] [CrossRef] [PubMed]
- Esmon, C. The protein C pathway. Crit. Care Med. 2000, 28, S44–S48. [Google Scholar] [CrossRef] [PubMed]
- van Moorsel, M.V.A.; Poolen, G.C.; Koekman, C.A.; Verhoef, S.; de Maat, S.; Barendrecht, A.; van Kleef, N.D.; Meijers, J.C.M.; Schiffelers, R.M.; Maas, C.; et al. VhH anti-thrombomodulin clone 1 inhibits TAFI activation and enhances fibrinolysis in human whole blood under flow. J. Thromb. Haemost. 2022, 20, 1213–1222. [Google Scholar] [CrossRef]
- Sillen, M.; Declerck, P.J. Thrombin Activatable Fibrinolysis Inhibitor (TAFI): An Updated Narrative Review. Int. J. Mol. Sci. 2021, 22, 3670. [Google Scholar] [CrossRef]
- Gurewich, V.; Pannell, R. Structure-function relationships in thrombin-activatable fibrinolysis inhibitor: Comment. J. Thromb. Haemost. 2016, 14, 1899–1900. [Google Scholar] [CrossRef]
- Conway, E.M. Thrombomodulin and its role in inflammation. Semin. Immunopathol. 2012, 34, 107–125. [Google Scholar] [CrossRef]
- Sanglas, L.; Arolas, J.L.; Valnickova, Z.; Aviles, F.X.; Enghild, J.J.; Gomis-Rüth, F.X. Insights into the molecular inactivation mechanism of human activated thrombin-activatable fibrinolysis inhibitor. J. Thromb. Haemost. 2010, 8, 1056–1065. [Google Scholar] [CrossRef]
- Marx, P.F.; Brondijk, T.H.C.; Plug, T.; Romijn, R.A.; Hemrika, W.; Meijers, J.C.M.; Huizinga, E.G. Crystal structures of TAFI elucidate the inactivation mechanism of activated TAFI: A novel mechanism for enzyme autoregulation. Blood 2008, 112, 2803–2809. [Google Scholar] [CrossRef]
- Marx, P.F.; Dawson, P.E.; Bouma, B.N.; Meijers, J.C.M. Plasmin-mediated activation and inactivation of thrombin-activatable fibrinolysis inhibitor. Biochemistry 2002, 41, 6688–6696. [Google Scholar] [CrossRef] [PubMed]
- Sansilvestri-Morel, P.; Rupin, A.; Schaffner, A.-P.; Bertin, F.; Mennecier, P.; Lapret, I.; Declerck, P.J.; Baumy, P.; Vallez, M.-O.; Petit-Dop, F.; et al. S62798, a potent TAFIa inhibitor, accelerates endogenous fibrinolysis in a murine model of pulmonary thromboembolism. Thromb. Res. 2021, 204, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wu, W.; Zhu, Q.; Fu, S.; Wang, X.; Hong, S.; Guo, R.; Bao, B. Identification and Fibrinolytic Evaluation of an Isoindolone Derivative Isolated from a Rare Marine Fungus Stachybotrys longispora FG216. Chin. J. Chem. 2015, 33, 1089–1095. [Google Scholar] [CrossRef]
- Guo, R.; Duan, D.; Hong, S.; Zhou, Y.; Wang, F.; Wang, S.; Wu, W.; Bao, B. A marine fibrinolytic compound FGFC1 stimulating enzymatic kinetic parameters of a reciprocal activation system based on a single chain urokinase-type plasminogen activator and plasminogen. Process Biochem. 2018, 68, 190–196. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, H.; Sheng, R.; Fu, Z.; Fan, J.; Wu, W.; Tu, Q.; Guo, R. Synthesis and Bioactivities of Marine Pyran-Isoindolone Derivatives as Potential Antithrombotic Agents. Mar. Drugs 2021, 19, 218. [Google Scholar] [CrossRef]
- Hang, S.; Chen, H.; Wu, W.; Wang, S.; Fang, Y.; Sheng, R.; Tu, Q.; Guo, R. Progress in Isoindolone Alkaloid Derivatives from Marine Microorganism: Pharmacology, Preparation, and Mechanism. Mar. Drugs 2022, 20, 405. [Google Scholar] [CrossRef]
- Gao, C.; Shen, Q.; Tang, P.; Cao, Y.; Lin, H.; Li, B.; Sun, P.; Bao, B.; Wu, W. In Vitro Study of the Fibrinolytic Activity via Single Chain Urokinase-Type Plasminogen Activator and Molecular Docking of FGFC1. Molecules 2021, 26, 1816. [Google Scholar] [CrossRef]
- Gao, C.; Tang, S.; Zhang, H.; Zhang, H.; Zhang, T.; Bao, B.; Zhu, Y.; Wu, W. A Novel Marine Pyran-Isoindolone Compound Enhances Fibrin Lysis Mediated by Single-Chain Urokinase-Type Plasminogen Activator. Mar. Drugs 2022, 20, 495. [Google Scholar] [CrossRef]
- Su, T.; Wu, W.; Yan, T.; Zhang, C.; Zhu, Q.; Bao, B. Pharmacokinetics and tissue distribution of a novel marine fibrinolytic compound in Wistar rat following intravenous administrations. J. Chromatogr. B-Anal. Technol. Biomed. Life Sci. 2013, 942, 77–82. [Google Scholar] [CrossRef]
- Juhan-Vague, I.; Renucci, J.F.; Grimaux, M.; Morange, P.E.; Gouvernet, J.; Gourmelin, Y.; Alessi, M.C. Thrombin-activatable fibrinolysis inhibitor antigen levels and cardiovascular risk factors. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2156–2161. [Google Scholar] [CrossRef]
- Eaton, D.L.; Malloy, B.E.; Tsai, S.P.; Henzel, W.; Drayna, D. Isolation, molecular cloning, and partial characterization of a novel carboxypeptidase B from human plasma. J. Biol. Chem. 1991, 266, 21833–21838. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.K.; Eaton, D.L. Activation and Characterization of Procarboxypeptidase B from Human Plasma. Biochemistry 1995, 34, 5811–5816. [Google Scholar] [CrossRef]
- Mark, S.; Michael, B.; Ronald, S.; Mona, R.; Marlys, K.; Michael, N. Two naturally occurring variants of TAFI (Thr-325 and Ile-325) differ substantially with respect to thermal stability and antifibrinolytic activity of the enzyme. J. Biol. Chem. 2002, 277, 1021–1030. [Google Scholar]
- Predrag, M.; Evelien, H.; Johan, W.; Valentina, D.; Dragica, R.; Milica, C.; Ivo, E.; Dirk, H. Thrombin activatable fibrinolysis inhibitor (TAFI): A molecular link between coagulation and fibrinolysis. Srp. Arh. Celok. Lek. 2010, 138 (Suppl. 1), 74–78. [Google Scholar]
- Bajzar, L.; Morser, J.; Nesheim, M. TAFI, or plasma procarboxypeptidase B, couples the coagulation and fibrinolytic cascades through the thrombin-thrombomodulin complex. J. Biol. Chem. 1996, 271, 16603–16608. [Google Scholar] [CrossRef] [PubMed]
- Van de Wouwer, M.; Collen, D.; Conway, E.M. Thrombomodulin-protein C-EPCR system-Integrated to regulate coagulation and inflammation. Arter. Thromb. Vasc. Biol. 2004, 24, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Kim, P.Y.; Swystun, L.L.; Liaw, P.C.; Weitz, J.I. Activation of protein C and thrombin activable fibrinolysis inhibitor on cultured human endothelial cells. J. Thromb. Haemost. 2016, 14, 366–374. [Google Scholar] [CrossRef]
- Castaneda-Hurtado, D.F.; Perez-Samano, D.; Rios-Gomez, M.; Montes-Castaneda, J.E.; Montes-Ramirez, J.E. Acute Bilateral Stroke in a Moyamoya Patient with High Risk of Thrombosis Due to Multiple Myeloma With Chemotherapy. Cureus 2023, 15, e34172. [Google Scholar] [CrossRef]
- Castillo Tokumori, F.; Komrokji, R.; Kuykendall, A.T. Stepping out of antiquity: An update on emerging drugs for the treatment of polycythemia vera. Expert Opin. Emerg. Drugs 2021, 26, 209–218. [Google Scholar] [CrossRef]
- Diao, X.; Han, H.; Sun, H.; Zhang, H.; Wu, W. Protection of Tight Junctional Complexes between hCMEC/D3 Cells by Deep-Sea Fibrinolytic Compound FGFC1. Mar. Drugs 2024, 22, 341. [Google Scholar] [CrossRef]
- Schnaith, E.; Beyrau, R.; Buckner, B.; Klein, R.M.; Rick, W. Optimized determination of angiotensin I-converting enzyme activity with hippuryl-L-histidyl-L-leucine as substrate. Clin. Chim. Acta Int. J. Clin. Chem. 1994, 227, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Marx, P.F.; Plug, T.; Havik, S.R.; Mörgelin, M.; Meijers, J.C.M. The activation peptide of thrombin-activatable fibrinolysis inhibitor: A role in activity and stability of the enzyme? J. Thromb. Haemost. JTH 2009, 7, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.-C.; Chiou, Y.-N.; Hsu, H.-C.; Chen, Y.-H.; Chiou, J.-Y.; Lin, C.-P. Evaluation on the Analytical Methods for Tissue Plasminogen Activator (tPA) in Pharmaceutical Formulations. J. Food Drug Anal. 2009, 17, 7. [Google Scholar] [CrossRef]
- Valnickova, Z.; Christensen, T.; Skottrup, P.; Thogersen, I.B.; Hojrup, P.; Enghild, J.J. Post-translational modifications of human thrombin-activatable fibrinolysis inhibitor (TAFI): Evidence for a large shift in the isoelectric point and reduced solubility upon activation. Biochemistry 2006, 45, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Friberger, P.; Knos, M.; Gustavsson, S.; Aurell, L.; Claeson, G. Methods for determination of plasmin, antiplasmin and plasminogen by means of substrate S-2251. Haemostasis 1978, 7, 138–145. [Google Scholar]
- Oyebamiji, A.K.; Josiah, O.M.; Akintelu, S.A.; Adeoye, M.D.; Sabitu, B.O.; Latona, D.F.; Esan, A.O.; Soetan, E.A.; Semire, B. Dataset on insightful bio-evaluation of 2-(quinoline-4-yloxy)acetamide analogues as potential anti-Mycobacterium tuberculosis catalase-peroxidase agents via in silico mechanisms. Data Brief 2021, 38, 107441. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Binding Energy | Interaction | ||
|---|---|---|---|---|
| H Bond | Hydrophobic | Electrostatic | ||
| FGFC1-TAFI | −5.62 kcal/mol | SER291, HIS293 | LEU10, ARG12, LYS43, PRO68 | LYS44 |
| FGFC1-TM | −0.52 kcal/mol | ASP60E | LYS60F, PHE60H, TRP60D | ARG35M |
| TAFI-FGFC1-TM | R12-T366, G184-I90, G188-P92, K324-D60E, N328-Q387 | |||
| Tuning | Linear |
|---|---|
| Polarity | Positive |
| Mass range | 5–80 kDa |
| Laser rep. rate | 200 Hz |
| Laser power | 120 |
| Accumulation rate (shots/profile) | 50 |
| Profiles | 200 |
| Pulsed extract | 30,000.00 |
| Ion gate blanking | 5000.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Diao, X.; Jiang, T.; Wei, M.; Su, Y.; Shen, J.; Bao, C.; Wu, W. Novel Deep Sea Isoindole Alkaloid FGFC1 Exhibits Its Fibrinolytic Effects by Inhibiting Thrombin-Activatable Fibrinolysis Inhibitor. Pharmaceuticals 2024, 17, 1401. https://doi.org/10.3390/ph17101401
Zhang H, Diao X, Jiang T, Wei M, Su Y, Shen J, Bao C, Wu W. Novel Deep Sea Isoindole Alkaloid FGFC1 Exhibits Its Fibrinolytic Effects by Inhibiting Thrombin-Activatable Fibrinolysis Inhibitor. Pharmaceuticals. 2024; 17(10):1401. https://doi.org/10.3390/ph17101401
Chicago/Turabian StyleZhang, Haixing, Xiaozhen Diao, Tingting Jiang, Mingjun Wei, Yue Su, Jingjing Shen, Chunlin Bao, and Wenhui Wu. 2024. "Novel Deep Sea Isoindole Alkaloid FGFC1 Exhibits Its Fibrinolytic Effects by Inhibiting Thrombin-Activatable Fibrinolysis Inhibitor" Pharmaceuticals 17, no. 10: 1401. https://doi.org/10.3390/ph17101401
APA StyleZhang, H., Diao, X., Jiang, T., Wei, M., Su, Y., Shen, J., Bao, C., & Wu, W. (2024). Novel Deep Sea Isoindole Alkaloid FGFC1 Exhibits Its Fibrinolytic Effects by Inhibiting Thrombin-Activatable Fibrinolysis Inhibitor. Pharmaceuticals, 17(10), 1401. https://doi.org/10.3390/ph17101401