


Research Progress of Flavonoids Regulating Endothelial Function

Abstract
:1. Introduction
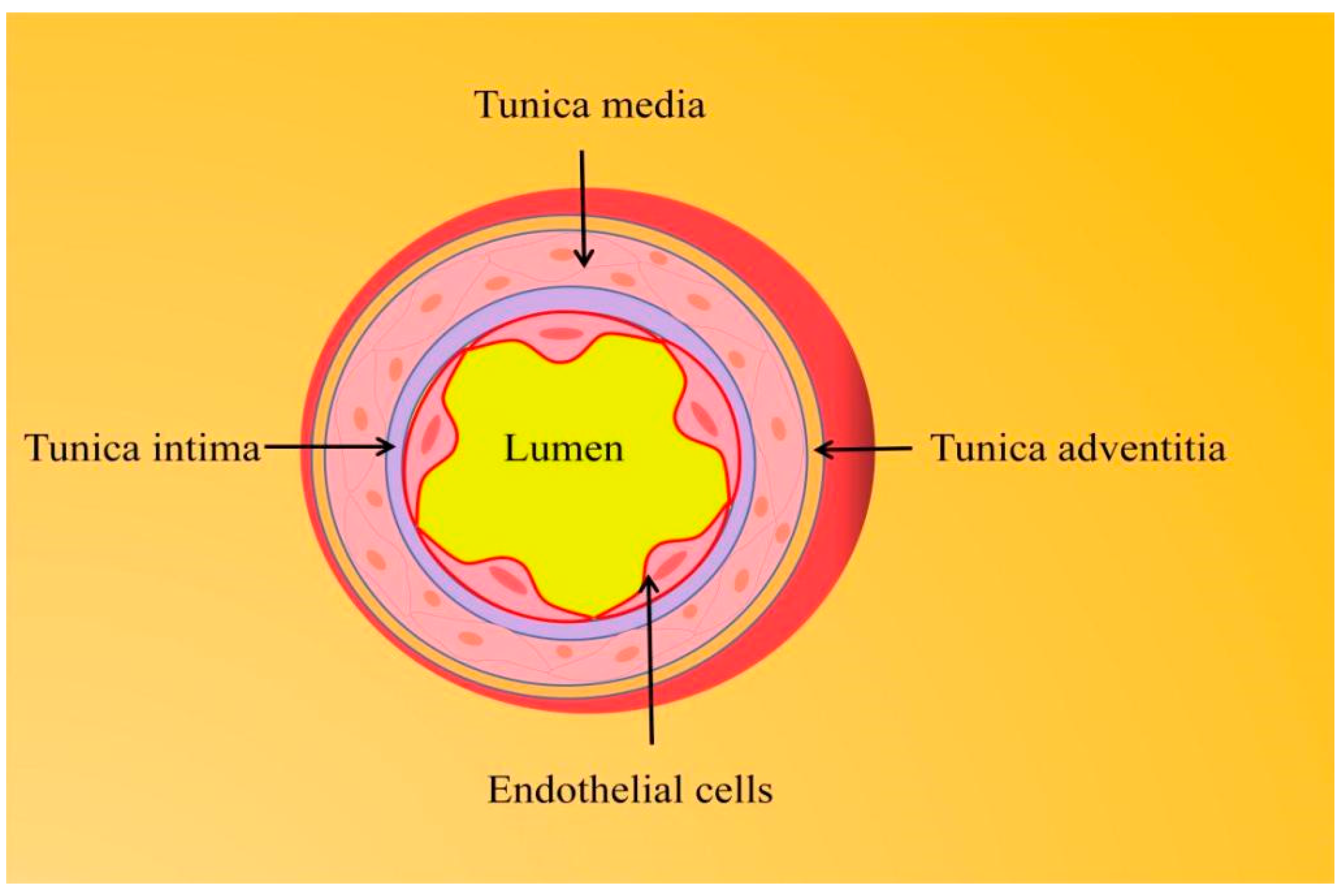
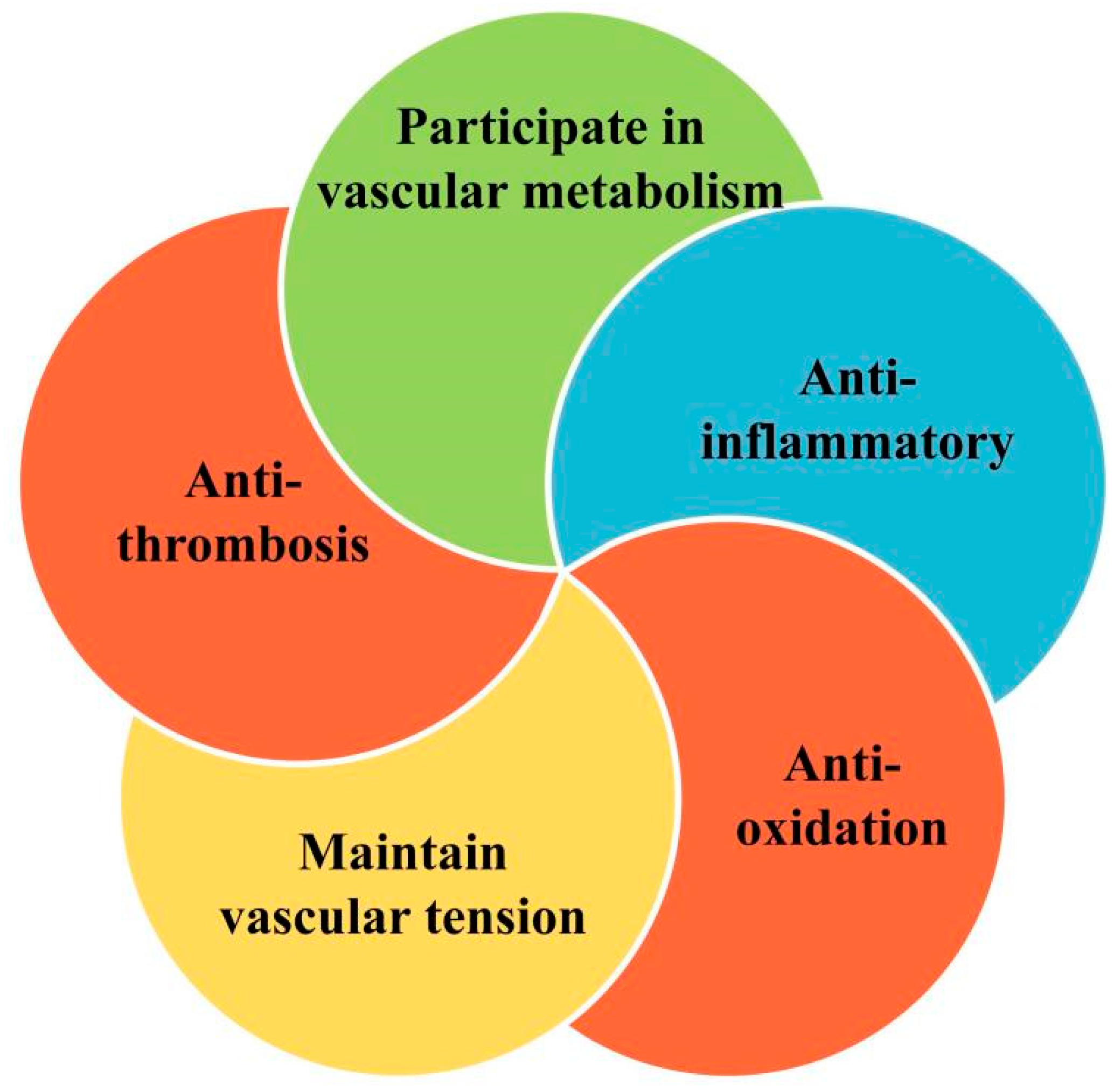
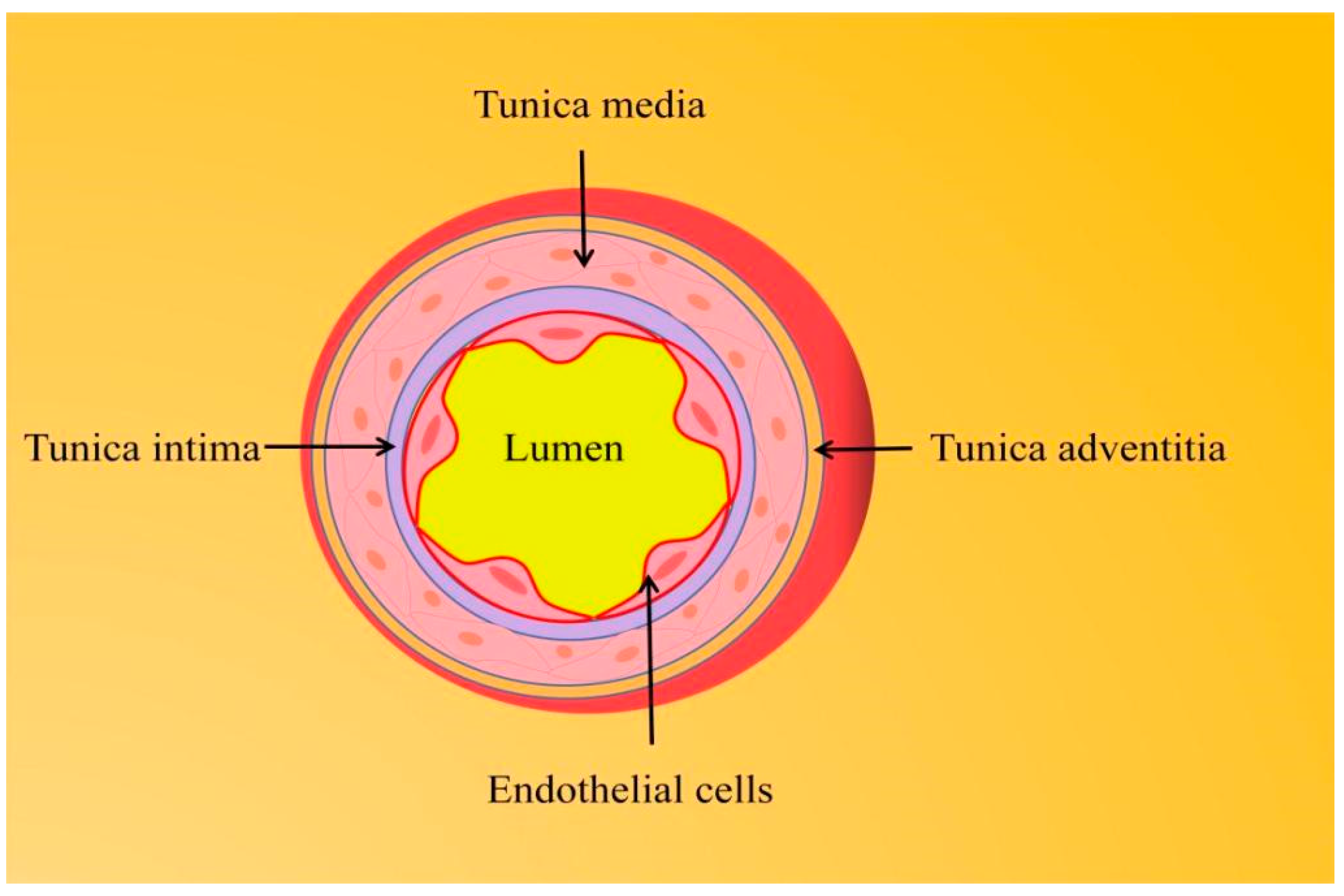
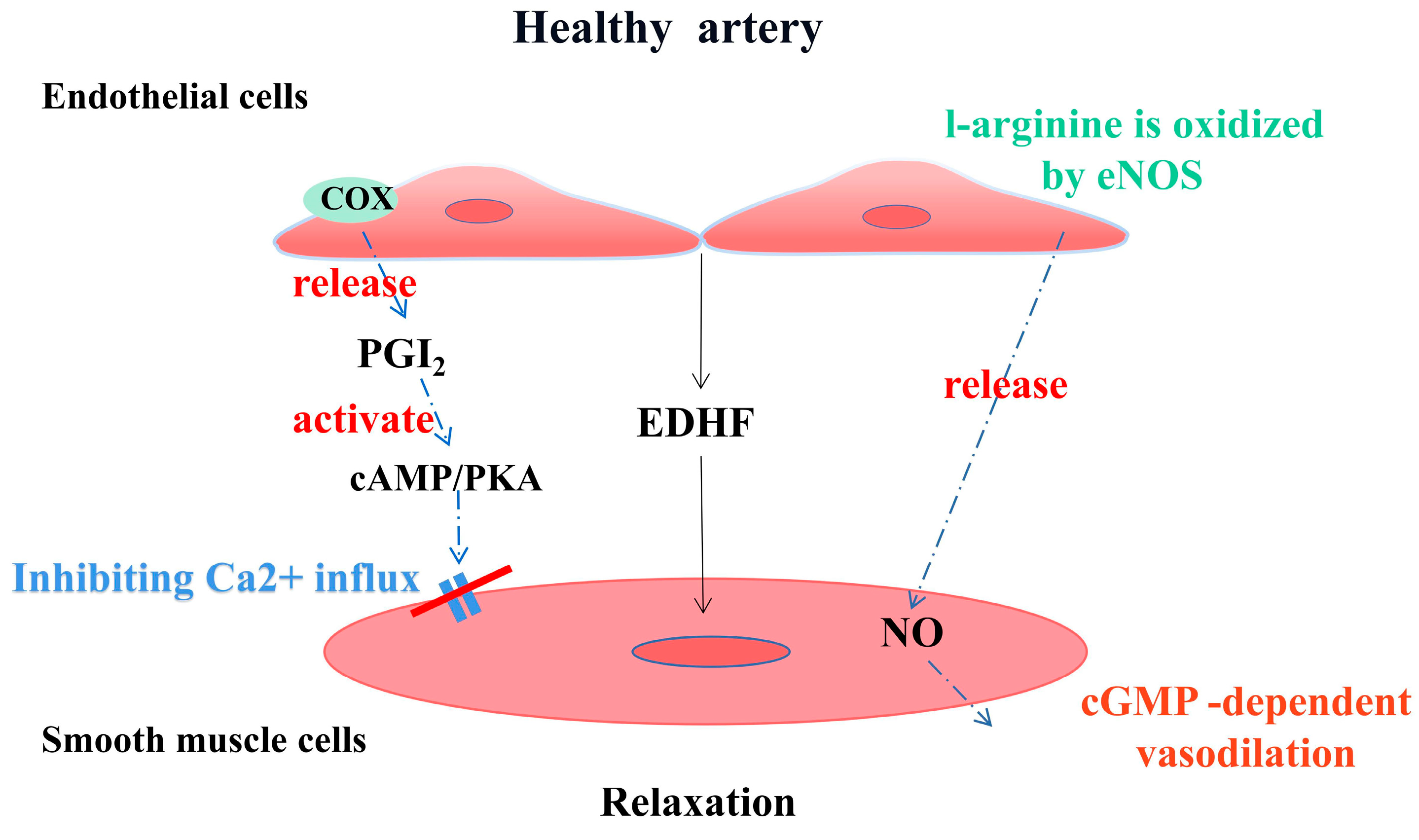
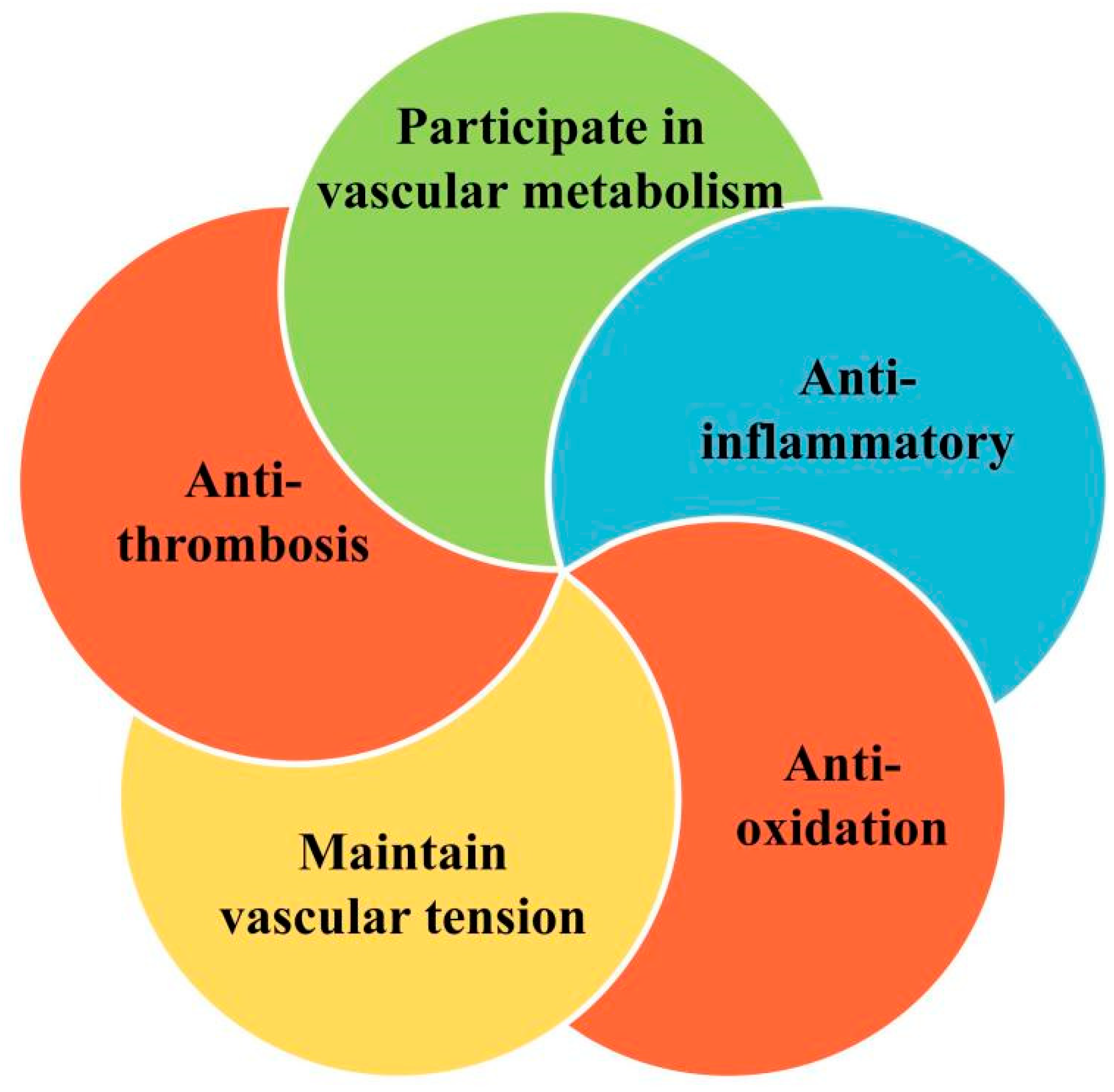
1.1. Endothelial Function and Vascular Homeostasis
1.2. Endothelial Dysfunction
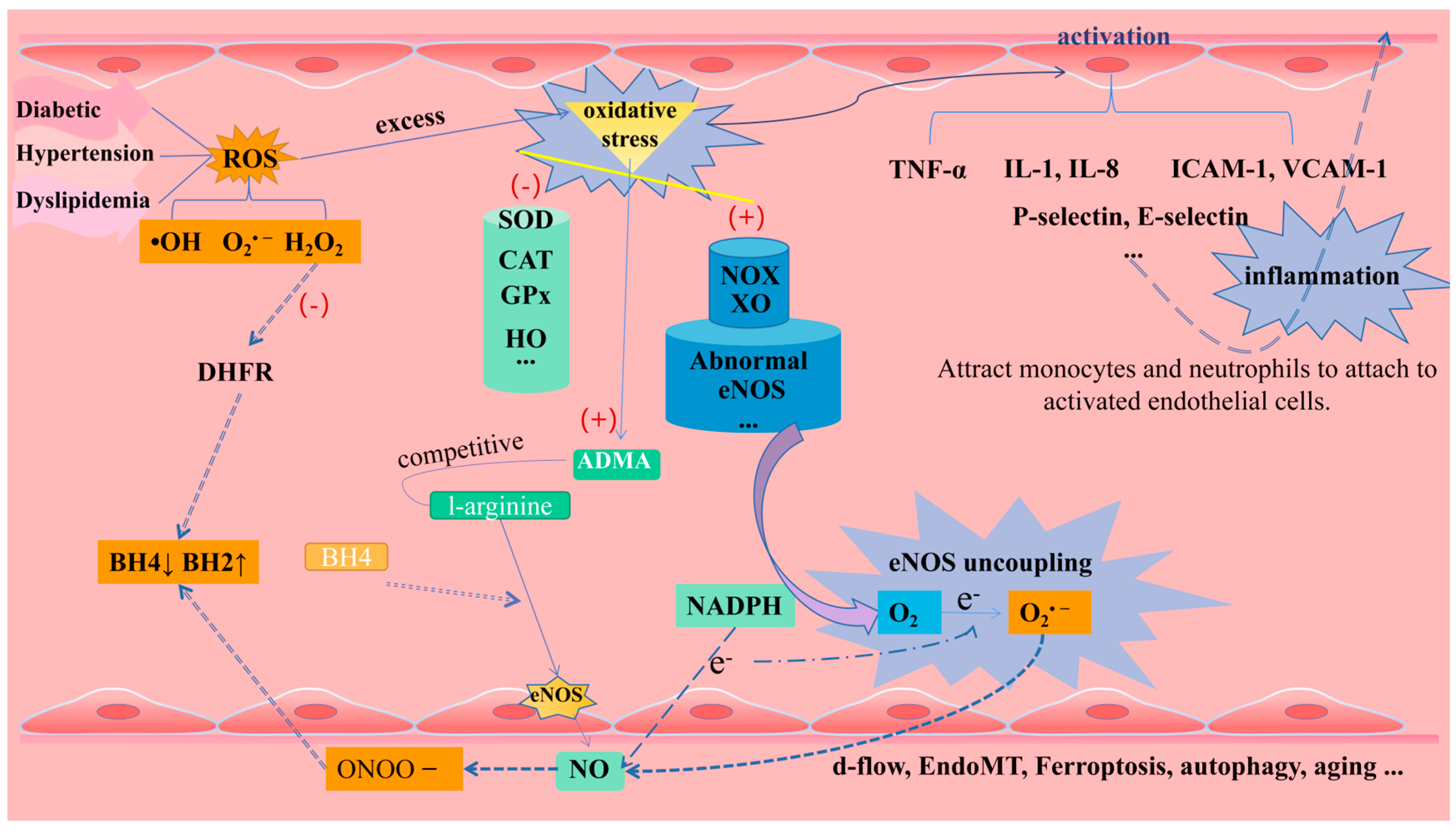
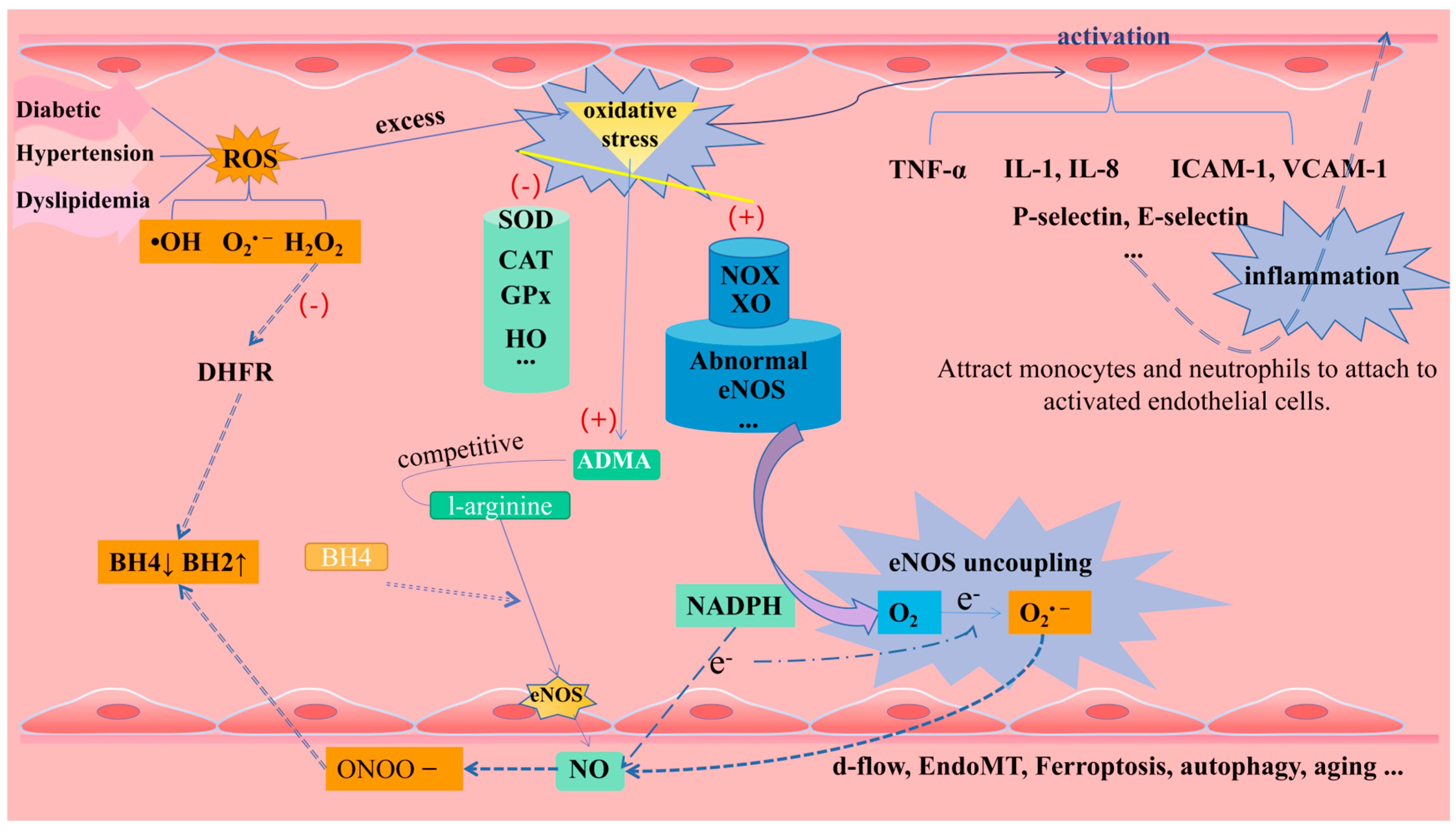
1.2.1. Oxidative Stress and eNOS Uncoupling
1.2.2. Inflammation
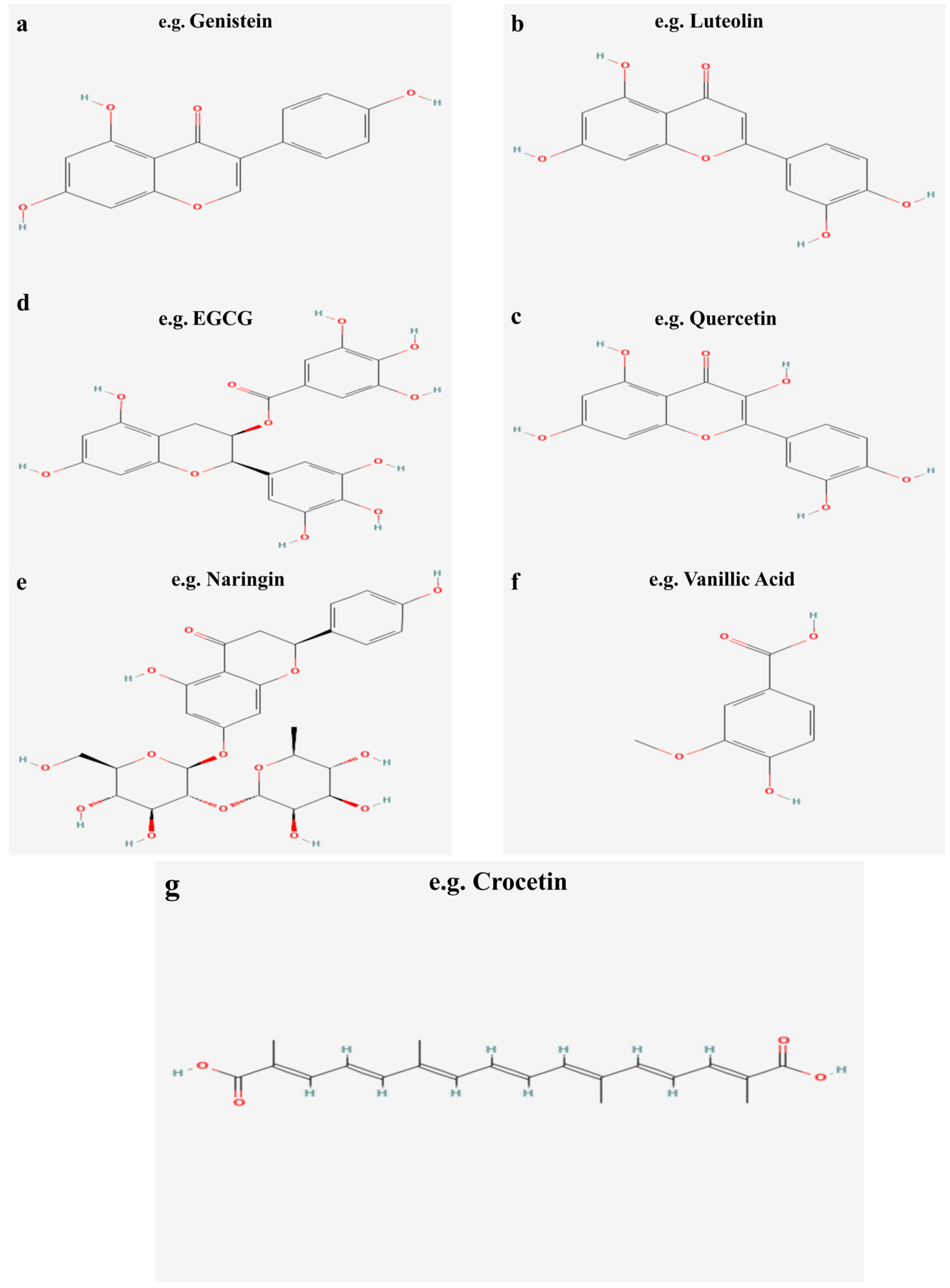
2. Flavonoids
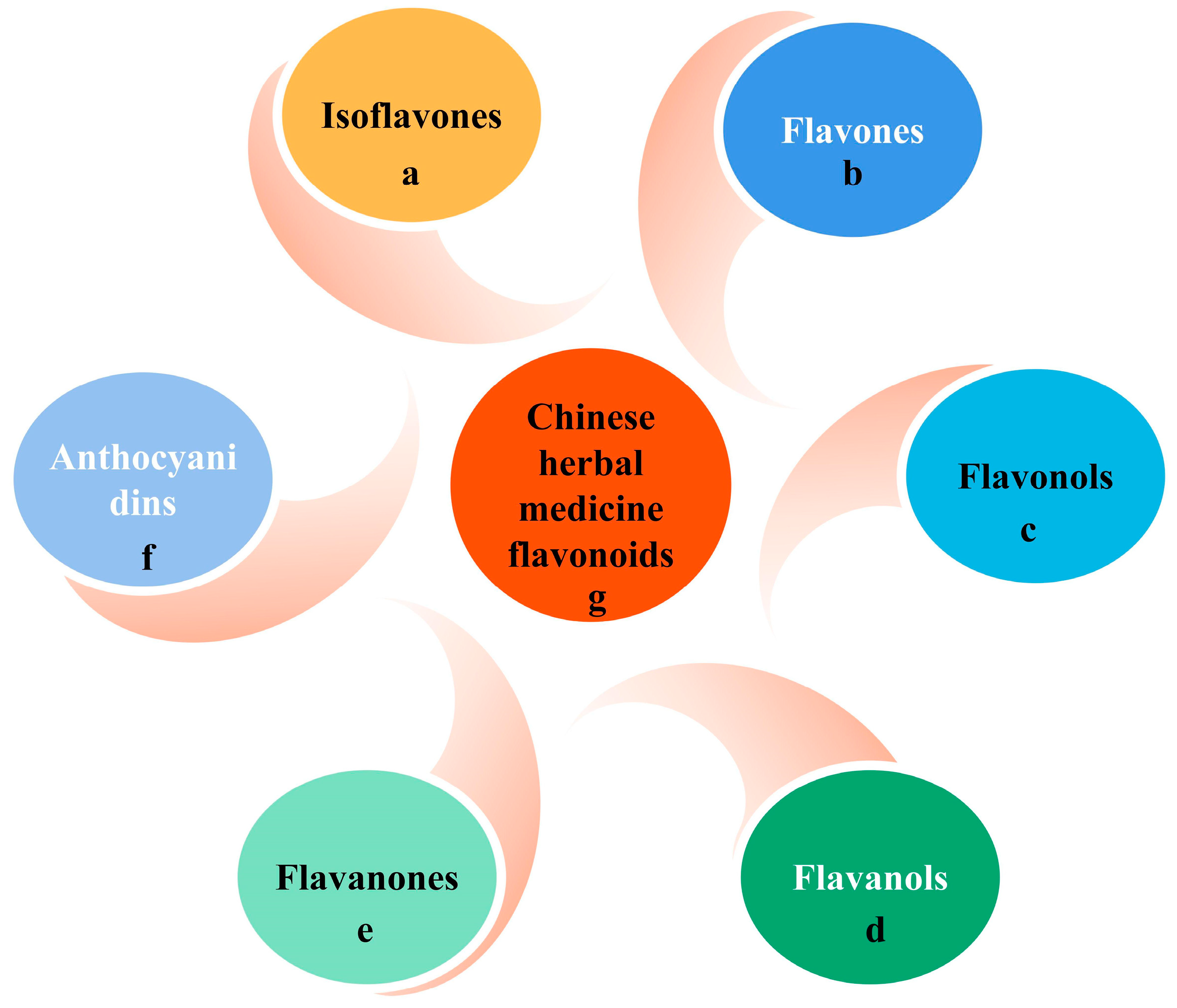
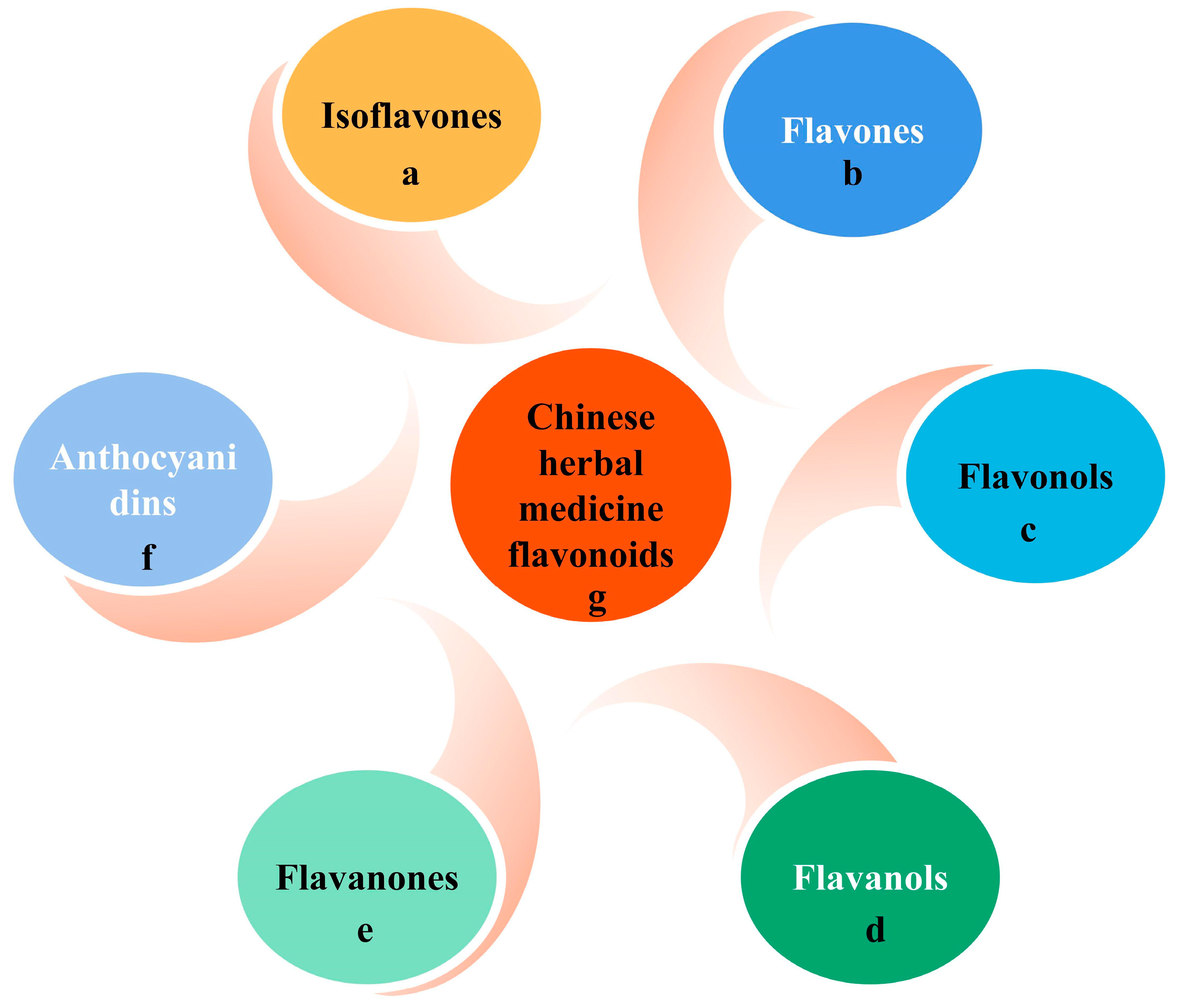
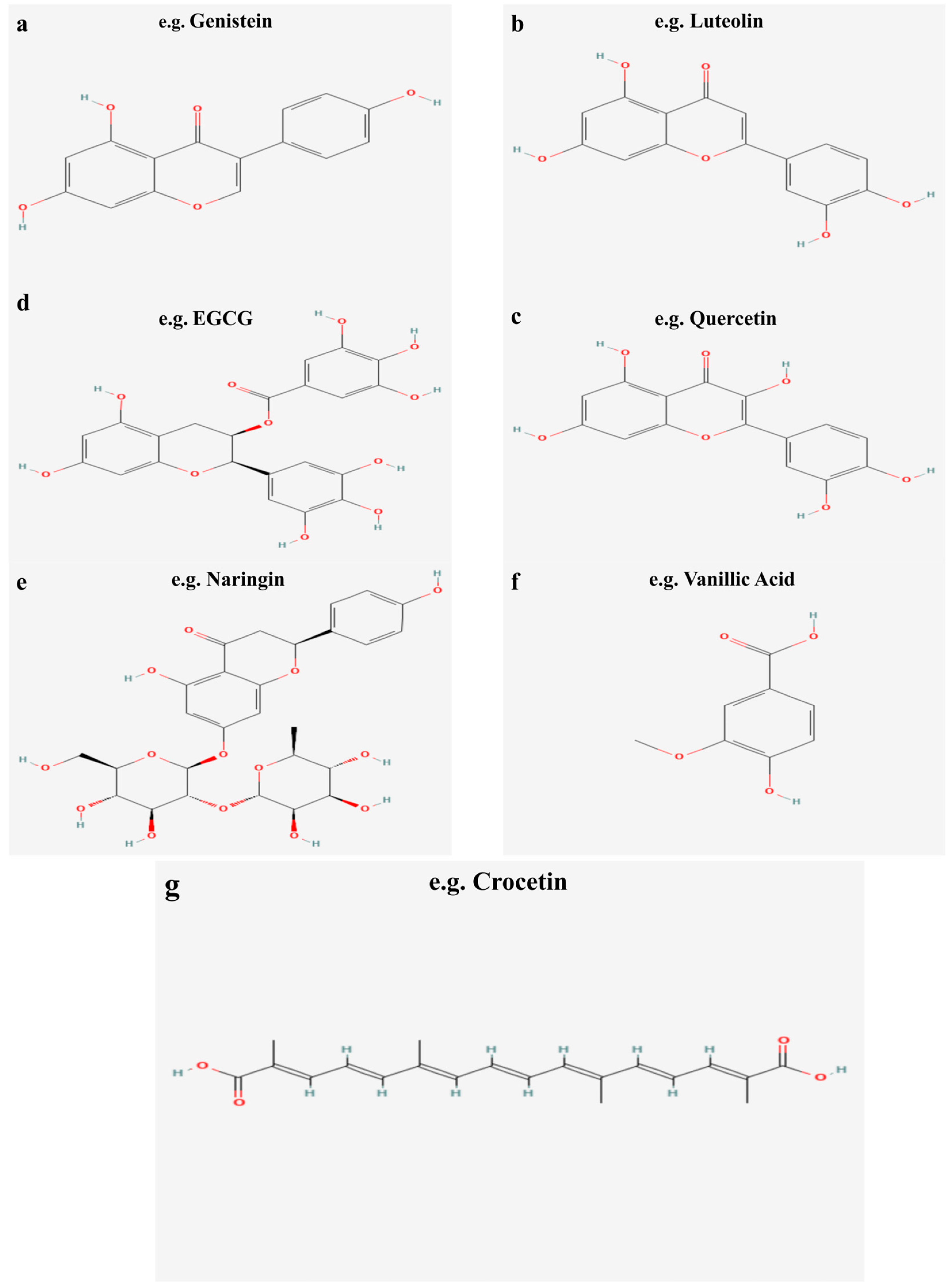
2.1. Sources, Classification, and Chemical Properties of Flavonoids
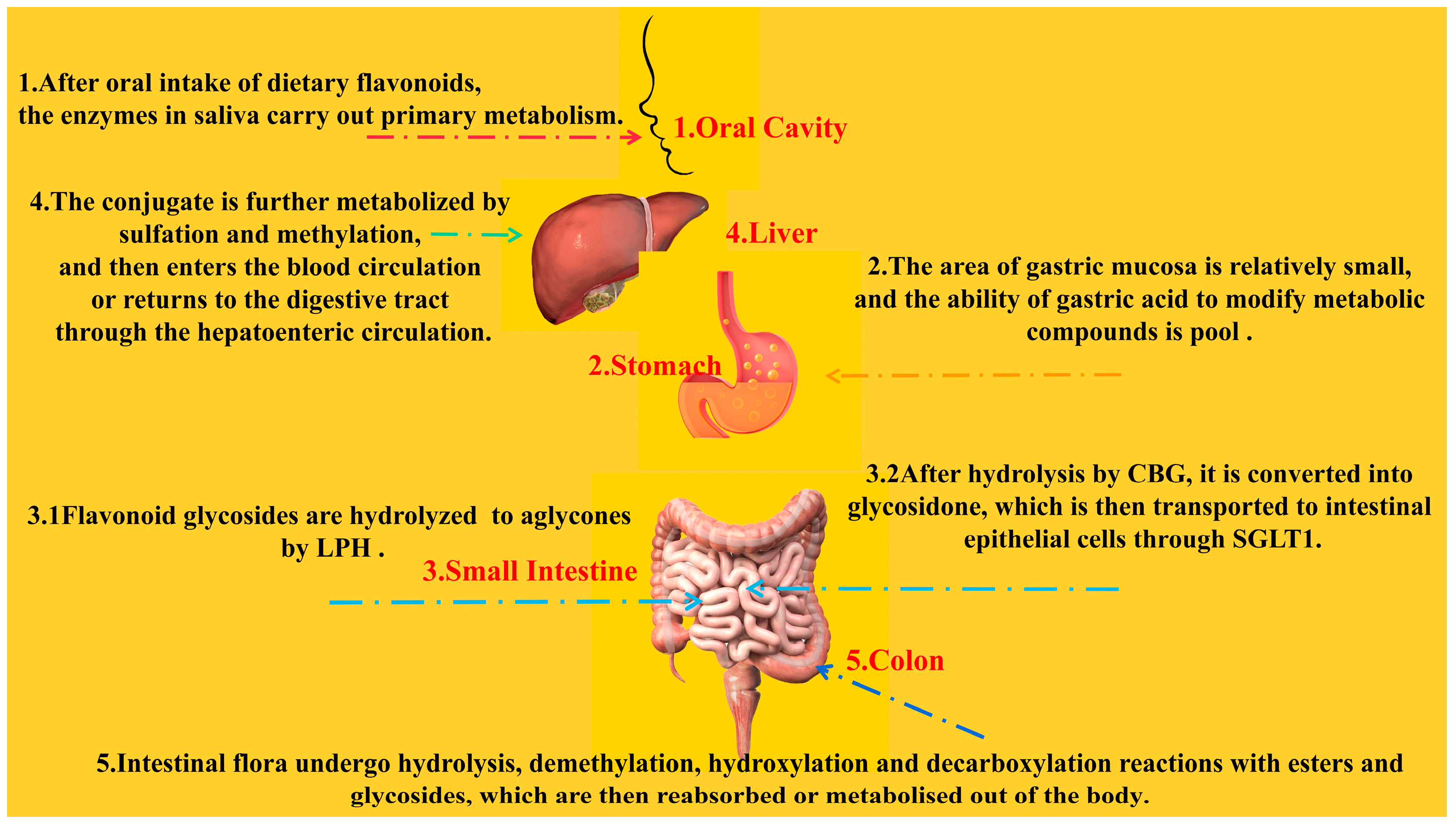
2.2. Bioavailability of Flavonoids
3. Protective Effects of Flavonoids on Endothelial Cells
3.1. Flavones (Luteolin and Baicalin)
3.2. Flavonols (Quercetin)
3.3. Flavanols (EGCG)
3.4. Flavanones (Hesperidin and Naringin)
3.5. Isoflavones (Genistein and Daidzein)
3.6. Anthocyanins
3.7. Chinese Herbal Medicine Flavonoids (Crocetin and Crocin)
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sacks, F.M.; Lichtenstein, A.H.; Wu, J.H.Y.; Appel, L.J.; Creager, M.A.; Kris-Etherton, P.M.; Miller, M.; Rimm, E.B.; Rudel, L.L.; Robinson, J.G.; et al. Dietary Fats and Cardiovascular Disease: A Presidential Advisory from the American Heart Association. Circulation 2017, 136, e1–e23. [Google Scholar] [CrossRef]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative Stress and Reactive Oxygen Species in Endothelial Dysfunction Associated with Cardiovascular and Metabolic Diseases. Vasc. Pharmacol. 2018, 100, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Endemann, D.H.; Schiffrin, E.L. Endothelial Dysfunction. J. Am. Soc. Nephrol. 2004, 15, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Konukoglu, D.; Uzun, H. Endothelial Dysfunction and Hypertension. Adv. Exp. Med. Biol. 2017, 956, 511–540. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, P.M.; Shimokawa, H.; Feletou, M.; Tang, E.H.C. Endothelial Dysfunction and Vascular Disease—A 30th Anniversary Update. Acta Physiol. 2017, 219, 22–96. [Google Scholar] [CrossRef] [PubMed]
- Salas-Salvadó, J.; Becerra-Tomás, N.; García-Gavilán, J.F.; Bulló, M.; Barrubés, L. Mediterranean Diet and Cardiovascular Disease Prevention: What Do We Know? Prog. Cardiovasc. Dis. 2018, 61, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A Concise Overview on the Chemistry, Occurrence, and Human Health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The Effects of Plant Flavonoids on Mammalian Cells: Implications for Inflammation, Heart Disease, and Cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Bohn, T. Dietary Factors Affecting Polyphenol Bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef]
- Gao, J.; Hou, T. Cardiovascular Disease Treatment Using Traditional Chinese Medicine: Mitochondria as the Achilles’ Heel. Biomed. Pharmacother. 2023, 164, 114999. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhao, Q.; Li, L.; Huang, S.; Yi, S.; Hu, Z. Effect of Traditional Chinese Medicine on the Cardiovascular Diseases. Front. Pharmacol. 2022, 13, 806300. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Bai, W. Bioactive Phytochemicals. Crit. Rev. Food Sci. Nutr. 2019, 59, 827–829. [Google Scholar] [CrossRef]
- Gouverneur, M.; Van Den Berg, B.; Nieuwdorp, M.; Stroes, E.; Vink, H. Vasculoprotective Properties of the Endothelial Glycocalyx: Effects of Fluid Shear Stress. J. Intern. Med. 2006, 259, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W.S. Vascular Nitric Oxide: Beyond ENOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef]
- Versteeg, H.H.; Heemskerk, J.W.M.; Levi, M.; Reitsma, P.H.; Middleton, E.A.; Weyrich, A.S.; Zimmerman, G.A.; Bevers, E.M.; Williamson, P.L.; Wu, X.; et al. New Fundamentals in Hemostasis. Physiol. Rev. 2013, 93, 327–358. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S. Endothelial Mechanotransduction, Redox Signaling and the Regulation of Vascular Inflammatory Pathways. Front. Physiol. 2018, 9, 524. [Google Scholar] [CrossRef]
- Huang, H.; Vandekeere, S.; Kalucka, J.; Bierhansl, L.; Zecchin, A.; Brüning, U.; Visnagri, A.; Yuldasheva, N.; Goveia, J.; Cruys, B.; et al. Role of Glutamine and Interlinked Asparagine Metabolism in Vessel Formation. EMBO J. 2017, 36, 2334–2352. [Google Scholar] [CrossRef]
- Yetik-Anacak, G.; Catravas, J.D. Nitric Oxide and the Endothelium: History and Impact on Cardiovascular Disease. Vasc. Pharmacol. 2006, 45, 268–276. [Google Scholar] [CrossRef]
- Puddu, G.M.; Cravero, E.; Arnone, G.; Muscari, A.; Puddu, P. Molecular Aspects of Atherogenesis: New Insights and Unsolved Questions. J. Biomed. Sci. 2005, 12, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Venditti, P. Evolution of the Knowledge of Free Radicals and Other Oxidants. Oxid. Med. Cell. Longev. 2020, 2020, 9829176. [Google Scholar] [CrossRef] [PubMed]
- Shaito, A.; Aramouni, K.; Assaf, R.; Parenti, A.; Orekhov, A.; Yazbi, A.E.; Pintus, G.; Eid, A.H. Oxidative Stress-Induced Endothelial Dysfunction in Cardiovascular Diseases. Front. Biosci. 2022, 27, 105. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Harrison, D.G. Endothelial Dysfunction in Cardiovascular Diseases: The Role of Oxidant Stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef]
- Daiber, A.; Xia, N.; Steven, S.; Oelze, M.; Hanf, A.; Kröller-Schön, S.; Münzel, T.; Li, H. New Therapeutic Implications of Endothelial Nitric Oxide Synthase (ENOS) Function/Dysfunction in Cardiovascular Disease. Int. J. Mol. Sci. 2019, 20, 187. [Google Scholar] [CrossRef] [PubMed]
- Milstien, S.; Katusic, Z. Oxidation of Tetrahydrobiopterin by Peroxynitrite: Implications for Vascular Endothelial Function. Biochem. Biophys. Res. Commun. 1999, 263, 681–684. [Google Scholar] [CrossRef]
- Chalupsky, K.; Cai, H. Endothelial Dihydrofolate Reductase: Critical for Nitric Oxide Bioavailability and Role in Angiotensin II Uncoupling of Endothelial Nitric Oxide Synthase. Proc. Natl. Acad. Sci. USA 2005, 102, 9056–9061. [Google Scholar] [CrossRef]
- Böger, R.H.; Sydow, K.; Borlak, J.; Thum, T.; Lenzen, H.; Schubert, B.; Tsikas, D.; Bode-Böger, S.M. LDL Cholesterol Upregulates Synthesis of Asymmetrical Dimethylarginine in Human Endothelial Cells: Involvement of S-Adenosylmethionine-Dependent Methyltransferases. Circ. Res. 2000, 87, 99–105. [Google Scholar] [CrossRef]
- Chen, C.A.; Wang, T.Y.; Varadharaj, S.; Reyes, L.A.; Hemann, C.; Talukder, M.A.H.; Chen, Y.R.; Druhan, L.J.; Zweier, J.L. S-Glutathionylation Uncouples ENOS and Regulates Its Cellular and Vascular Function. Nature 2010, 468, 1115–1120. [Google Scholar] [CrossRef]
- Ali, L.; Schnitzler, J.G.; Kroon, J. Metabolism: The Road to Inflammation and Atherosclerosis. Curr. Opin. Lipidol. 2018, 29, 474–480. [Google Scholar] [CrossRef]
- Tabas, I.; García-Cardeña, G.; Owens, G.K. Recent Insights into the Cellular Biology of Atherosclerosis. J. Cell Biol. 2015, 209, 13–22. [Google Scholar] [CrossRef]
- Zhu, Y.; Xian, X.; Wang, Z.; Bi, Y.; Chen, Q.; Han, X.; Tang, D.; Chen, R. Research Progress on the Relationship between Atherosclerosis and Inflammation. Biomolecules 2018, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Libby, P. Interleukin-1 Beta as a Target for Atherosclerosis Therapy: Biological Basis of CANTOS and Beyond. J. Am. Coll. Cardiol. 2017, 70, 2278–2289. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Roumeliotis, N.; Sawamura, T.; Renier, G. C-Reactive Protein Enhances LOX-1 Expression in Human Aortic Endothelial Cells: Relevance of LOX-1 to C-Reactive Protein-Induced Endothelial Dysfunction. Circ. Res. 2004, 95, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Binesh, A.; Devaraj, S.N.; Halagowder, D. Molecular Interaction of NFκB and NICD in Monocyte-Macrophage Differentiation Is a Target for Intervention in Atherosclerosis. J. Cell. Physiol. 2019, 234, 7040–7050. [Google Scholar] [CrossRef]
- Kleinbongard, P.; Heusch, G.; Schulz, R. TNFalpha in Atherosclerosis, Myocardial Ischemia/Reperfusion and Heart Failure. Pharmacol. Ther. 2010, 127, 295–314. [Google Scholar] [CrossRef]
- Trojanowicz, B.; Ulrich, C.; Seibert, E.; Fiedler, R.; Girndt, M. Uremic Conditions Drive Human Monocytes to Pro-Atherogenic Differentiation via an Angiotensin-Dependent Mechanism. PLoS ONE 2014, 9, e102137. [Google Scholar] [CrossRef]
- Das, S.; Zhang, E.; Senapati, P.; Amaram, V.; Reddy, M.A.; Stapleton, K.; Leung, A.; Lanting, L.; Wang, M.; Chen, Z.; et al. A Novel Angiotensin II-Induced Long Noncoding RNA Giver Regulates Oxidative Stress, Inflammation, and Proliferation in Vascular Smooth Muscle Cells. Circ. Res. 2018, 123, 1298–1312. [Google Scholar] [CrossRef]
- Park, M.Y.; Ha, S.E.; Kim, H.H.; Bhosale, P.B.; Abusaliya, A.; Jeong, S.H.; Park, J.S.; Heo, J.D.; Kim, G.S. Scutellarein Inhibits LPS-Induced Inflammation through NF-ΚB/MAPKs Signaling Pathway in RAW264.7 Cells. Molecules 2022, 27, 3782. [Google Scholar] [CrossRef]
- Dhingra, S.; Sharma, A.K.; Singla, D.K.; Singal, P.K. P38 and ERK1/2 MAPKs Mediate the Interplay of TNF-Alpha and IL-10 in Regulating Oxidative Stress and Cardiac Myocyte Apoptosis. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H3524–H3531. [Google Scholar] [CrossRef]
- Souilhol, C.; Harmsen, M.C.; Evans, P.C.; Krenning, G. Endothelial-Mesenchymal Transition in Atherosclerosis. Cardiovasc. Res. 2018, 114, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liu, J.; Piao, H.; Zhu, Z.; Wei, R.; Liu, K. ROS-Triggered Endothelial Cell Death Mechanisms: Focus on Pyroptosis, Parthanatos, and Ferroptosis. Front. Immunol. 2022, 13, 1039241. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.; You, J.; Zhi, C.; Li, P.; Lin, X.; Tan, X.; Ma, W.; Li, L.; Xie, W. Ferroptosis: The Potential Value Target in Atherosclerosis. Cell Death Dis. 2021, 12, 782. [Google Scholar] [CrossRef]
- Bravo-San Pedro, J.M.; Kroemer, G.; Galluzzi, L. Autophagy and Mitophagy in Cardiovascular Disease. Circ. Res. 2017, 120, 1812–1824. [Google Scholar] [CrossRef] [PubMed]
- Mameli, E.; Martello, A.; Caporali, A. Autophagy at the Interface of Endothelial Cell Homeostasis and Vascular Disease. FEBS J. 2022, 289, 2976–2991. [Google Scholar] [CrossRef]
- Khan, H.; Belwal, T.; Efferth, T.; Farooqi, A.A.; Sanches-Silva, A.; Vacca, R.A.; Nabavi, S.F.; Khan, F.; Prasad Devkota, H.; Barreca, D.; et al. Targeting Epigenetics in Cancer: Therapeutic Potential of Flavonoids. Crit. Rev. Food Sci. Nutr. 2021, 61, 1616–1639. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Huff, M.W. Antiatherogenic Properties of Flavonoids: Implications for Cardiovascular Health. Can. J. Cardiol. 2010, 26 (Suppl. A), 17A–21A. [Google Scholar] [CrossRef]
- Xiao, J.; Capanoglu, E.; Jassbi, A.R.; Miron, A. Advance on the Flavonoid C-Glycosides and Health Benefits. Crit. Rev. Food Sci. Nutr. 2016, 56 (Suppl. 1), S29–S45. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef]
- Walle, T.; Browning, A.M.; Steed, L.L.; Reed, S.G.; Walle, U.K. Flavonoid Glucosides Are Hydrolyzed and Thus Activated in the Oral Cavity in Humans. J. Nutr. 2005, 135, 48–52. [Google Scholar] [CrossRef]
- Chen, L.; Cao, H.; Huang, Q.; Xiao, J.; Teng, H. Absorption, Metabolism and Bioavailability of Flavonoids: A Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 7730–7742. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.Y.; Lin, Y.K.; Lin, C.F.; Wang, P.W.; Chen, E.L.; Fang, J.Y. Elucidating the Skin Delivery of Aglycone and Glycoside Flavonoids: How the Structures Affect Cutaneous Absorption. Nutrients 2017, 9, 1304. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Fang, X.; Yang, J.; Yao, Y.; Nandakumar, K.S.; Salem, M.L.; Cheng, K. Recent Research on Flavonoids and Their Biomedical Applications. Curr. Med. Chem. 2021, 28, 1042–1066. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Su, S.; Xin, M.; Zhang, Z.; Nan, X.; Li, Z.; Lu, D. Luteolin Ameliorates Hypoxia-Induced Pulmonary Hypertension via Regulating HIF-2α-Arg-NO Axis and PI3K-AKT-ENOS-NO Signaling Pathway. Phytomedicine 2022, 104, 154329. [Google Scholar] [CrossRef]
- Ou, H.C.; Pandey, S.; Hung, M.Y.; Huang, S.H.; Hsu, P.T.; Day, C.H.; Pai, P.; Viswanadha, V.P.; Kuo, W.W.; Huang, C.Y. Luteolin: A Natural Flavonoid Enhances the Survival of HUVECs against Oxidative Stress by Modulating AMPK/PKC Pathway. Am. J. Chin. Med. 2019, 47, 541–557. [Google Scholar] [CrossRef]
- Chen, H.I.; Hu, W.S.; Hung, M.Y.; Ou, H.C.; Huang, S.H.; Hsu, P.T.; Day, C.H.; Lin, K.H.; Viswanadha, V.P.; Kuo, W.W.; et al. Protective Effects of Luteolin against Oxidative Stress and Mitochondrial Dysfunction in Endothelial Cells. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 1032–1043. [Google Scholar] [CrossRef]
- Jia, Z.; Nallasamy, P.; Liu, D.; Shah, H.; Li, J.Z.; Chitrakar, R.; Si, H.; McCormick, J.; Zhu, H.; Zhen, W.; et al. Luteolin Protects against Vascular Inflammation in Mice and TNF-Alpha-Induced Monocyte Adhesion to Endothelial Cells via Suppressing IΚBα/NF-ΚB Signaling Pathway. J. Nutr. Biochem. 2015, 26, 293–302. [Google Scholar] [CrossRef]
- De Stefano, A.; Caporali, S.; Di Daniele, N.; Rovella, V.; Cardillo, C.; Schinzari, F.; Minieri, M.; Pieri, M.; Candi, E.; Bernardini, S.; et al. Anti-Inflammatory and Proliferative Properties of Luteolin-7-O-Glucoside. Int. J. Mol. Sci. 2021, 22, 1321. [Google Scholar] [CrossRef]
- Wei, X.; Zhu, X.; Hu, N.; Zhang, X.; Sun, T.; Xu, J.; Bian, X. Baicalin Attenuates Angiotensin II-Induced Endothelial Dysfunction. Biochem. Biophys. Res. Commun. 2015, 465, 101–107. [Google Scholar] [CrossRef]
- Hsu, W.L.; Lin, Y.C.; Jeng, J.R.; Chang, H.Y.; Chou, T.C. Baicalein Ameliorates Pulmonary Arterial Hypertension Caused by Monocrotaline through Downregulation of ET-1 and ETAR in Pneumonectomized Rats. Am. J. Chin. Med. 2018, 46, 769–783. [Google Scholar] [CrossRef]
- Wang, P.; Wu, J.; Wang, Q.; Zhuang, S.; Zhao, J.; Yu, Y.; Zhang, W.; Zheng, Y.; Liu, X. Baicalin Inhibited Both the Furin/TGFβ1/Smad3/TSP-1 Pathway in Endothelial Cells and the AKT/Ca2+/ROS Pathway in Platelets to Ameliorate Inflammatory Coagulopathy. Eur. J. Pharmacol. 2023, 949, 175674. [Google Scholar] [CrossRef] [PubMed]
- Vissenaekens, H.; Grootaert, C.; Raes, K.; De Munck, J.; Smagghe, G.; Boon, N.; Van Camp, J. Quercetin Mitigates Endothelial Activation in a Novel Intestinal-Endothelial-Monocyte/Macrophage Coculture Setup. Inflammation 2022, 45, 1600–1611. [Google Scholar] [CrossRef] [PubMed]
- Ozyel, B.; Le Gall, G.; Needs, P.W.; Kroon, P.A. Anti-Inflammatory Effects of Quercetin on High-Glucose and Pro-Inflammatory Cytokine Challenged Vascular Endothelial Cell Metabolism. Mol. Nutr. Food Res. 2021, 65, e2000777. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, X.; Zhu, G.; Liu, H.; Chen, J.; Wang, Y.; He, X. Quercetin Inhibits TNF-α Induced HUVECs Apoptosis and Inflammation via Downregulating NF-KB and AP-1 Signaling Pathway in Vitro. Medicine 2020, 99, E22241. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, F.; Qian, X.; Luo, S.; Wang, Z.; Gao, X.; Kong, X.; Zhang, J.; Chen, S. Quercetin Protects Endothelial Function from Inflammation Induced by Localized Disturbed Flow by Inhibiting NRP2 -VEGFC Complex. Int. Immunopharmacol. 2023, 116, 109842. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zheng, L.; Zhang, B.; Deng, Z.; Li, H. Synergistic Protection of Quercetin and Lycopene against Oxidative Stress via SIRT1-Nox4-ROS Axis in HUVEC Cells. Curr. Res. food Sci. 2022, 5, 1985–1993. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Tian, R.; Lu, N. Quercetin Inhibited Endothelial Dysfunction and Atherosclerosis in Apolipoprotein E-Deficient Mice: Critical Roles for NADPH Oxidase and Heme Oxygenase-1. J. Agric. Food Chem. 2020, 68, 10875–10883. [Google Scholar] [CrossRef]
- Li, M.T.; Ke, J.; Guo, S.F.; Wu, Y.; Bian, Y.F.; Shan, L.L.; Liu, Q.Y.; Huo, Y.J.; Guo, C.; Liu, M.Y.; et al. The Protective Effect of Quercetin on Endothelial Cells Injured by Hypoxia and Reoxygenation. Front. Pharmacol. 2021, 12, 732874. [Google Scholar] [CrossRef]
- Mohd Sabri, N.A.; Lee, S.K.; Murugan, D.D.; Ling, W.C. Epigallocatechin Gallate (EGCG) Alleviates Vascular Dysfunction in Angiotensin II-Infused Hypertensive Mice by Modulating Oxidative Stress and ENOS. Sci. Rep. 2022, 12, 17633. [Google Scholar] [CrossRef]
- Pai, P.Y.; Chou, W.C.; Chan, S.H.; Wu, S.Y.; Chen, H.I.; Li, C.W.; Hsieh, P.L.; Chu, P.M.; Chen, Y.A.; Ou, H.C.; et al. Epigallocatechin Gallate Reduces Homocysteine-Caused Oxidative Damages through Modulation SIRT1/AMPK Pathway in Endothelial Cells. Am. J. Chin. Med. 2021, 49, 113–129. [Google Scholar] [CrossRef]
- Meng, J.; Chen, Y.; Wang, J.; Qiu, J.; Chang, C.; Bi, F.; Wu, X.; Liu, W. EGCG Protects Vascular Endothelial Cells from Oxidative Stress-Induced Damage by Targeting the Autophagy-Dependent PI3K-AKT-MTOR Pathway. Ann. Transl. Med. 2020, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.T.; Lakshmi, S.P.; Maruthi Prasad, E.; Varadacharyulu, N.C.; Kodidhela, L.D. Epigallocatechin Gallate Suppresses Inflammation in Human Coronary Artery Endothelial Cells by Inhibiting NF-ΚB. Life Sci. 2020, 258, 118136. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Liu, M.; Liu, H.; Suo, R.; Lu, C. Naringin Protects Endothelial Cells from Apoptosis and Inflammation by Regulating the Hippo-YAP Pathway. Biosci. Rep. 2020, 40, BSR20193431. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, L.; Cao, Z.; Li, W.; Liu, R.; Chen, Y.; Li, C.; Song, Y.; Liu, G.; Hu, J.; et al. Naringenin Ameliorates Homocysteine Induced Endothelial Damage via the AMPKα/Sirt1 Pathway. J. Adv. Res. 2021, 34, 137–147. [Google Scholar] [CrossRef]
- Feng, J.; Luo, J.; Deng, L.; Zhong, Y.; Wen, X.; Cai, Y.; Li, J. Naringenin-Induced HO-1 Ameliorates High Glucose or Free Fatty Acids-Associated Apoptosis via PI3K and JNK/Nrf2 Pathways in Human Umbilical Vein Endothelial Cells. Int. Immunopharmacol. 2019, 75, 105769. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.S.; Tew, W.Y.; Jingying, C.; Yam, M.F. Vasorelaxant Effect of 5,7,4’- Trihydroxyflavanone (Naringenin) via Endothelium Dependent, Potassium and Calcium Channels in Sprague Dawley Rats: Aortic Ring Model. Chem. Biol. Interact. 2021, 348, 109620. [Google Scholar] [CrossRef]
- Gao, G.; Nakamura, S.; Asaba, S.; Miyata, Y.; Nakayama, H.; Matsui, T. Hesperidin Preferentially Stimulates Transient Receptor Potential Vanilloid 1, Leading to NO Production and Mas Receptor Expression in Human Umbilical Vein Endothelial Cells. J. Agric. Food Chem. 2022, 70, 11290–11300. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, X.; Pang, X.; Zhao, Z.; Yu, H.; Zhou, H. Genistein Protects against Ox-LDL-Induced Senescence through Enhancing SIRT1/LKB1/AMPK-Mediated Autophagy Flux in HUVECs. Mol. Cell. Biochem. 2019, 455, 127–134. [Google Scholar] [CrossRef]
- Wu, G.; Li, S.; Qu, G.; Hua, J.; Zong, J.; Li, X.; Xu, F. Genistein Alleviates H2O2-Induced Senescence of Human Umbilical Vein Endothelial Cells via Regulating the TXNIP/NLRP3 Axis. Pharm. Biol. 2021, 59, 1388–1401. [Google Scholar] [CrossRef]
- XIE, X.; CONG, L.; LIU, S.; XIANG, L.; FU, X. Genistein Alleviates Chronic Vascular Inflammatory Response via the MiR-21/NF-κB P65 Axis in Lipopolysaccharide-treated Mice. Mol. Med. Rep. 2021, 23, 11831. [Google Scholar] [CrossRef]
- Huang, W.; Hutabarat, R.P.; Chai, Z.; Zheng, T.; Zhang, W.; Li, D. Antioxidant Blueberry Anthocyanins Induce Vasodilation via PI3K/Akt Signaling Pathway in High-Glucose-Induced Human Umbilical Vein Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 1575. [Google Scholar] [CrossRef]
- Lee, G.H.; Hoang, T.H.; Jung, E.S.; Jung, S.J.; Han, S.K.; Chung, M.J.; Chae, S.W.; Chae, H.J. Anthocyanins Attenuate Endothelial Dysfunction through Regulation of Uncoupling of Nitric Oxide Synthase in Aged Rats. Aging Cell 2020, 19, e13279. [Google Scholar] [CrossRef]
- Li, W.; Zhang, G.; Tan, S.; Gong, C.; Yang, Y.; Gu, M.; Mi, Z.; Yang, H.Y. Polyacylated Anthocyanins Derived from Red Radishes Protect Vascular Endothelial Cells Against Palmitic Acid-Induced Apoptosis via the P38 MAPK Pathway. Plant Foods Hum. Nutr. 2022, 77, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Kam, H.T.; Chen, Y.; Gong, G.; Hoi, M.P.M.; Skalicka-Woźniak, K.; Dias, A.C.P.; Lee, S.M.Y. Crocetin and Its Glycoside Crocin, Two Bioactive Constituents from Crocus sativus L. (Saffron), Differentially Inhibit Angiogenesis by Inhibiting Endothelial Cytoskeleton Organization and Cell Migration Through VEGFR2/SRC/FAK and VEGFR2/MEK/ERK Signaling Pathways. Front. Pharmacol. 2021, 12, 675359. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.T.; Zhang, P.L.; Li, Y.C.; Sun, M.R.; Wang, Y.T.; Wang, S.P.; Yang, H.; Liu, B.L.; Wang, M.; et al. Hydroxysafflor Yellow A Blocks HIF-1 α Induction of NOX2 and Protects ZO-1 Protein in Cerebral Microvascular Endothelium. Antioxidants 2022, 11, 728. [Google Scholar] [CrossRef]
- Yang, J.; Wang, R.; Cheng, X.; Qu, H.C.; Qi, J.; Li, D.; Xing, Y.; Bai, Y.; Zheng, X. The Vascular Dilatation Induced by Hydroxysafflor Yellow A (HSYA) on Rat Mesenteric Artery through TRPV4-Dependent Calcium Influx in Endothelial Cells. J. Ethnopharmacol. 2020, 256, 112790. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A.; Ming, Y.; Pengyi, H.; Jie, K.Y.; Yali, L.; Haiyan, Z.; Shuai, X.; Wenjing, L.; Ling, W.; Xia, Z.M.; et al. The Fate of Flavonoids after Oral Administration: A Comprehensive Overview of Its Bioavailability. Crit. Rev. Food Sci. Nutr. 2022, 62, 6169–6186. [Google Scholar] [CrossRef] [PubMed]
- Takahama, U.; Yamauchi, R.; Hirota, S. Antioxidative Flavonoids in Adzuki-Meshi (Rice Boiled with Adzuki Bean) React with Nitrite under Simulated Stomach Conditions. J. Funct. Foods 2016, 26, 657–666. [Google Scholar] [CrossRef]
- Babadi, D.; Dadashzadeh, S.; Osouli, M.; Daryabari, M.S.; Haeri, A. Nanoformulation Strategies for Improving Intestinal Permeability of Drugs: A More Precise Look at Permeability Assessment Methods and Pharmacokinetic Properties Changes. J. Control. Release 2020, 321, 669–709. [Google Scholar] [CrossRef]
- Wang, S.; Mateos, R.; Goya, L.; Amigo-Benavent, M.; Sarriá, B.; Bravo, L. A Phenolic Extract from Grape By-Products and Its Main Hydroxybenzoic Acids Protect Caco-2 Cells against pro-Oxidant Induced Toxicity. Food Chem. Toxicol. 2016, 88, 65–74. [Google Scholar] [CrossRef]
- Day, A.J.; Gee, J.M.; DuPont, M.S.; Johnson, I.T.; Williamson, G. Absorption of Quercetin-3-Glucoside and Quercetin-4′-Glucoside in the Rat Small Intestine: The Role of Lactase Phlorizin Hydrolase and the Sodium-Dependent Glucose Transporter. Biochem. Pharmacol. 2003, 65, 1199–1206. [Google Scholar] [CrossRef] [PubMed]
- Németh, K.; Plumb, G.W.; Berrin, J.G.; Juge, N.; Jacob, R.; Naim, H.Y.; Williamson, G.; Swallow, D.M.; Kroon, P.A. Deglycosylation by Small Intestinal Epithelial Cell Beta-Glucosidases Is a Critical Step in the Absorption and Metabolism of Dietary Flavonoid Glycosides in Humans. Eur. J. Nutr. 2003, 42, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Kottra, G.; Daniel, H. Flavonoid Glycosides Are Not Transported by the Human Na+/Glucose Transporter When Expressed in Xenopus Laevis Oocytes, but Effectively Inhibit Electrogenic Glucose Uptake. J. Pharmacol. Exp. Ther. 2007, 322, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Van Der Woude, H.; Boersma, M.G.; Vervoort, J.; Rietjens, I.M.C.M. Identification of 14 Quercetin Phase II Mono- and Mixed Conjugates and Their Formation by Rat and Human Phase II in Vitro Model Systems. Chem. Res. Toxicol. 2004, 17, 1520–1530. [Google Scholar] [CrossRef]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, Excretion and Metabolite Profiling of Methyl-, Glucuronyl-, Glucosyl- and Sulpho-Conjugates of Quercetin in Human Plasma and Urine after Ingestion of Onions. Br. J. Nutr. 2006, 96, 107. [Google Scholar] [CrossRef]
- Hollman, P.C.H. Absorption, Bioavailability, and Metabolism of Flavonoids. Pharm. Biol. 2004, 42, 74–83. [Google Scholar] [CrossRef]
- Steed, A.L.; Christophi, G.P.; Kaiko, G.E.; Sun, L.; Goodwin, V.M.; Jain, U.; Esaulova, E.; Artyomov, M.N.; Morales, D.J.; Holtzman, M.J.; et al. The Microbial Metabolite Desaminotyrosine Protects from Influenza through Type I Interferon. Science 2017, 357, 498–502. [Google Scholar] [CrossRef]
- Williamson, G.; Clifford, M.N. Role of the Small Intestine, Colon and Microbiota in Determining the Metabolic Fate of Polyphenols. Biochem. Pharmacol. 2017, 139, 24–39. [Google Scholar] [CrossRef]
- Clifford, M.N. Diet-Derived Phenols in Plasma and Tissues and Their Implications for Health. Planta Med. 2004, 70, 1103–1114. [Google Scholar] [CrossRef]
- Crozier, A.; Del Rio, D.; Clifford, M.N. Bioavailability of Dietary Flavonoids and Phenolic Compounds. Mol. Asp. Med. 2010, 31, 446–467. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [PubMed]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Miean, K.H.; Mohamed, S. Flavonoid (Myricetin, Quercetin, Kaempferol, Luteolin, and Apigenin) Content of Edible Tropical Plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kong, X.; Wang, M.; Li, J.; Chen, W.; Jiang, D. Luteolin Partially Inhibits LFA-1 Expression in Neutrophils Through the ERK Pathway. Inflammation 2019, 42, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, M.; Lei, X.; Huang, M.; Ye, W.; Zhang, R.; Zhang, D. Luteolin Inhibits Angiogenesis by Blocking Gas6/Axl Signaling Pathway. Int. J. Oncol. 2017, 51, 677–685. [Google Scholar] [CrossRef]
- Kampa, R.P.; Flori, L.; Sęk, A.; Spezzini, J.; Brogi, S.; Szewczyk, A.; Calderone, V.; Bednarczyk, P.; Testai, L. Luteolin-Induced Activation of Mitochondrial BKCa Channels: Undisclosed Mechanism of Cytoprotection. Antioxidants 2022, 11, 1892. [Google Scholar] [CrossRef]
- Kang, M.J.; Ko, G.S.; Oh, D.G.; Kim, J.S.; Noh, K.; Kang, W.; Yoon, W.K.; Kim, H.C.; Jeong, H.G.; Jeong, T.C. Role of Metabolism by Intestinal Microbiota in Pharmacokinetics of Oral Baicalin. Arch. Pharmacal Res. 2014, 37, 371–378. [Google Scholar] [CrossRef]
- Ding, L.; Jia, C.; Zhang, Y.; Wang, W.; Zhu, W.; Chen, Y.; Zhang, T. Baicalin Relaxes Vascular Smooth Muscle and Lowers Blood Pressure in Spontaneously Hypertensive Rats. Biomed. Pharmacother. 2019, 111, 325–330. [Google Scholar] [CrossRef]
- Bai, J.; Wang, Q.; Qi, J.; Yu, H.; Wang, C.; Wang, X.; Ren, Y.; Yang, F. Promoting Effect of Baicalin on Nitric Oxide Production in CMECs via Activating the PI3K-AKT-ENOS Pathway Attenuates Myocardial Ischemia-Reperfusion Injury. Phytomedicine 2019, 63, 153035. [Google Scholar] [CrossRef]
- Tsai, C.L.; Tsai, C.W.; Chang, W.S.; Lin, J.C.; Hsia, T.C.; Bau, D.T. Protective Effects of Baicalin on Arsenic Trioxide-Induced Oxidative Damage and Apoptosis in Human Umbilical Vein Endothelial Cells. In Vivo 2021, 35, 155–162. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Shi, R.; Wei, Z.; Zhu, D.; Fu, N.; Wang, C.; Yin, S.; Liang, Y.; Xing, J.; Wang, X.; Wang, Y. Baicalein Attenuates Monocrotaline-Induced Pulmonary Arterial Hypertension by Inhibiting Vascular Remodeling in Rats. Pulm. Pharmacol. Ther. 2018, 48, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Zhang, Z.; Zhao, Y.; Wang, D. Baicalin Reduces Immune Cell Infiltration by Inhibiting Inflammation and Protecting Tight Junctions in Ischemic Stroke Injury. Am. J. Chin. Med. 2023, 51, 355–372. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Ku, S.K.; Bae, J.S. Antiplatelet, Anticoagulant, and Profibrinolytic Activities of Baicalin. Arch. Pharmacal Res. 2015, 38, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yu, G.; Yu, Q.; Wang, L.; Wu, L.; Tao, Z.; Ding, J.; Lin, D. Baicalin Promotes Random-Pattern Skin Flap Survival by Inducing Autophagy via AMPK-Regulated TFEB Nuclear Transcription. Phytother. Res. 2023. [Google Scholar] [CrossRef]
- Zhao, L.; Xiong, M.; Liu, Y. Baicalin Enhances the Proliferation and Invasion of Trophoblasts and Suppresses Vascular Endothelial Damage by Modulating Long Non-Coding RNA NEAT1/MiRNA-205-5p in Hypertensive Disorder Complicating Pregnancy. J. Obstet. Gynaecol. Res. 2021, 47, 3060–3070. [Google Scholar] [CrossRef]
- Liu, L.; Lan, X.; Chen, X.; Dai, S.; Wang, Z.; Zhao, A.; Lu, L.; Huang, N.; Chen, J.; Yang, P.; et al. Multi-Functional Plant Flavonoids Regulate Pathological Microenvironments for Vascular Stent Surface Engineering. Acta Biomater. 2023, 157, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Perez-Vizcaino, F.; Duarte, J. Flavonols and Cardiovascular Disease. Mol. Asp. Med. 2010, 31, 478–494. [Google Scholar] [CrossRef]
- Terao, J. Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health. Antioxidants 2023, 12, 258. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Ge, C.; Li, W.; Li, R. 3-(3-Hydroxyphenyl)Propionic Acid, a Microbial Metabolite of Quercetin, Inhibits Monocyte Binding to Endothelial Cells via Modulating E-Selectin Expression. Fitoterapia 2022, 156, 105071. [Google Scholar] [CrossRef] [PubMed]
- Suganya, N.; Mani, K.P.; Sireesh, D.; Rajaguru, P.; Vairamani, M.; Suresh, T.; Suzuki, T.; Chatterjee, S.; Ramkumar, K.M. Establishment of Pancreatic Microenvironment Model of ER Stress: Quercetin Attenuates β-Cell Apoptosis by Invoking Nitric Oxide-CGMP Signaling in Endothelial Cells. J. Nutr. Biochem. 2018, 55, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Suganya, N.; Dornadula, S.; Chatterjee, S.; Mohanram, R.K. Quercetin Improves Endothelial Function in Diabetic Rats through Inhibition of Endoplasmic Reticulum Stress-Mediated Oxidative Stress. Eur. J. Pharmacol. 2018, 819, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Tumova, S.; Kerimi, A.; Williamson, G. Long Term Treatment with Quercetin in Contrast to the Sulfate and Glucuronide Conjugates Affects HIF1α Stability and Nrf2 Signaling in Endothelial Cells and Leads to Changes in Glucose Metabolism. Free Radic. Biol. Med. 2019, 137, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Rezabakhsh, A.; Rahbarghazi, R.; Malekinejad, H.; Fathi, F.; Montaseri, A.; Garjani, A. Quercetin Alleviates High Glucose-Induced Damage on Human Umbilical Vein Endothelial Cells by Promoting Autophagy. Phytomedicine 2019, 56, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.; Du, Y.; Zhang, M.; Zhu, A.R.; Zhang, J. Senolytics Cocktail Dasatinib and Quercetin Alleviate Human Umbilical Vein Endothelial Cell Senescence via the TRAF6-MAPK-NF-ΚB Axis in a YTHDF2-Dependent Manner. Gerontology 2022, 68, 920–934. [Google Scholar] [CrossRef] [PubMed]
- Dagher, O.; Mury, P.; Thorin-Trescases, N.; Noly, P.E.; Thorin, E.; Carrier, M. Therapeutic Potential of Quercetin to Alleviate Endothelial Dysfunction in Age-Related Cardiovascular Diseases. Front. Cardiovasc. Med. 2021, 8, 658400. [Google Scholar] [CrossRef]
- Márquez Campos, E.; Jakobs, L.; Simon, M.C. Antidiabetic Effects of Flavan-3-Ols and Their Microbial Metabolites. Nutrients 2020, 12, 1592. [Google Scholar] [CrossRef]
- Eng, Q.Y.; Thanikachalam, P.V.; Ramamurthy, S. Molecular Understanding of Epigallocatechin Gallate (EGCG) in Cardiovascular and Metabolic Diseases. J. Ethnopharmacol. 2018, 210, 296–310. [Google Scholar] [CrossRef]
- Kim, S.; Lee, H.; Moon, H.; Kim, R.; Kim, M.; Jeong, S.; Kim, H.; Kim, S.H.; Hwang, S.S.; Lee, M.Y.; et al. Epigallocatechin-3-Gallate Attenuates Myocardial Dysfunction via Inhibition of Endothelial-to-Mesenchymal Transition. Antioxidants 2023, 12, 1059. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, H.; Qiu, J.; Wang, X.; Zhong, Y.; Yao, C.; Yao, L.; Zheng, Q.; Xiong, C.H. Stability of Glycosylated Complexes Loaded with Epigallocatechin 3-Gallate (EGCG). Food Chem. 2023, 410, 135364. [Google Scholar] [CrossRef]
- Zhang, W.; Shen, H.; Li, Y.; Yang, K.; Lei, P.; Gu, Y.; Sun, L.; Xu, H.; Wang, R. Preparation of Type-A Gelatin/Poly-γ-Glutamic Acid Nanoparticles for Enhancing the Stability and Bioavailability of (-)-Epigallocatechin Gallate. Foods 2023, 12, 1748. [Google Scholar] [CrossRef] [PubMed]
- Imperatrice, M.; Cuijpers, I.; Troost, F.J.; Sthijns, M.M.J.P.E. Hesperidin Functions as an Ergogenic Aid by Increasing Endothelial Function and Decreasing Exercise-Induced Oxidative Stress and Inflammation, Thereby Contributing to Improved Exercise Performance. Nutrients 2022, 14, 2955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, H.; Hu, X.; He, Y.; Li, L.; Yang, X.; Wang, C.; Hu, M.; Tao, S. Heat Shock Protein 70 Mediates the Protective Effect of Naringenin on High-Glucose-Induced Alterations of Endothelial Function. Int. J. Endocrinol. 2022, 2022, 7275765. [Google Scholar] [CrossRef] [PubMed]
- Fuior, E.V.; Mocanu, C.A.; Deleanu, M.; Voicu, G.; Anghelache, M.; Rebleanu, D.; Simionescu, M.; Calin, M. Evaluation of VCAM-1 Targeted Naringenin/Indocyanine Green-Loaded Lipid Nanoemulsions as Theranostic Nanoplatforms in Inflammation. Pharmaceutics 2020, 12, 1066. [Google Scholar] [CrossRef]
- Zaheer, K.; Humayoun Akhtar, M. An Updated Review of Dietary Isoflavones: Nutrition, Processing, Bioavailability and Impacts on Human Health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1280–1293. [Google Scholar] [CrossRef] [PubMed]
- Toro-Funes, N.; Morales-Gutiérrez, F.J.; Veciana-Nogués, M.T.; Vidal-Carou, M.C.; Spencer, J.P.E.; Rodriguez-Mateos, A. The Intracellular Metabolism of Isoflavones in Endothelial Cells. Food Funct. 2015, 6, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Poasakate, A.; Maneesai, P.; Potue, P.; Bunbupha, S.; Tong-Un, T.; Settheetham-Ishida, W.; Khamseekaew, J.; Pakdeechote, P. Genistein Alleviates Renin-Angiotensin System Mediated Vascular and Kidney Alterations in Renovascular Hypertensive Rats. Biomed. Pharmacother. 2022, 146, 112601. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Q.; Shi, L.; Qin, L.; Zhang, Q.; Mi, M. Equol Attenuates Atherosclerosis in Apolipoprotein E-Deficient Mice by Inhibiting Endoplasmic Reticulum Stress via Activation of Nrf2 in Endothelial Cells. PLoS ONE 2016, 11, e0167020. [Google Scholar] [CrossRef]
- Park, M.H.; Ju, J.W.; Kim, M.; Han, J.S. The Protective Effect of Daidzein on High Glucose-Induced Oxidative Stress in Human Umbilical Vein Endothelial Cells. Z. Naturforsch. C. 2016, 71, 21–28. [Google Scholar] [CrossRef]
- Dong, Y.; Wu, X.; Han, L.; Bian, J.; He, C.; El-Omar, E.; Gong, L.; Wang, M. The Potential Roles of Dietary Anthocyanins in Inhibiting Vascular Endothelial Cell Senescence and Preventing Cardiovascular Diseases. Nutrients 2022, 14, 2836. [Google Scholar] [CrossRef]
- De Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.A.; Cassidy, A.; Kay, C.D. The Pharmacokinetics of Anthocyanins and Their Metabolites in Humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef] [PubMed]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human Metabolism and Elimination of the Anthocyanin, Cyanidin-3-Glucoside: A (13)C-Tracer Study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.; Czank, C.; Woodward, G.M.; Cassidy, A.; Kay, C.D. Phenolic Metabolites of Anthocyanins Modulate Mechanisms of Endothelial Function. J. Agric. Food Chem. 2015, 63, 2423–2431. [Google Scholar] [CrossRef] [PubMed]
- Krga, I.; Monfoulet, L.E.; Konic-Ristic, A.; Mercier, S.; Glibetic, M.; Morand, C.; Milenkovic, D. Anthocyanins and Their Gut Metabolites Reduce the Adhesion of Monocyte to TNFα-Activated Endothelial Cells at Physiologically Relevant Concentrations. Arch. Biochem. Biophys. 2016, 599, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Krga, I.; Tamaian, R.; Mercier, S.; Boby, C.; Monfoulet, L.E.; Glibetic, M.; Morand, C.; Milenkovic, D. Anthocyanins and Their Gut Metabolites Attenuate Monocyte Adhesion and Transendothelial Migration through Nutrigenomic Mechanisms Regulating Endothelial Cell Permeability. Free Radic. Biol. Med. 2018, 124, 364–379. [Google Scholar] [CrossRef]
- Herrera-Bravo, J.; Beltrán, J.F.; Huard, N.; Saavedra, K.; Saavedra, N.; Alvear, M.; Lanas, F.; Salazar, L.A. Anthocyanins Found in Pinot Noir Waste Induce Target Genes Related to the Nrf2 Signalling in Endothelial Cells. Antioxidants 2022, 11, 1239. [Google Scholar] [CrossRef]
- Chen, B.H.; Inbaraj, B.S. Nanoemulsion and Nanoliposome Based Strategies for Improving Anthocyanin Stability and Bioavailability. Nutrients 2019, 11, 1052. [Google Scholar] [CrossRef]
- Xi, L.; Qian, Z.; Du, P.; Fu, J. Pharmacokinetic Properties of Crocin (Crocetin Digentiobiose Ester) Following Oral Administration in Rats. Phytomedicine 2007, 14, 633–636. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-Brain Barrier: From Physiology to Disease and Back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef]
- Taguchi, K.; Okudaira, K.; Matsumoto, T.; Kobayashi, T. Ginkgolide B Caused the Activation of the Akt/ENOS Pathway through the Antioxidant Effect of SOD1 in the Diabetic Aorta. Pflugers Arch. 2023, 475, 453–463. [Google Scholar] [CrossRef]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, Biology and Role in Disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.; Mahmoud, M.; Liu, R.; Andueza, A.; Kumar, S.; Kang, D.W.; Zhang, J.; Tamargo, I.; Villa-Roel, N.; Baek, K.I.; et al. Stable Flow-Induced Expression of KLK10 Inhibits Endothelial Inflammation and Atherosclerosis. Elife 2022, 11, e72579. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Wang, S.; Shao, J.; Jin, Y.J.; Xu, L.; Yan, Y.; Günther, S.; Wang, L.; Offermanns, S. Tenascin-X Mediates Flow-Induced Suppression of EndMT and Atherosclerosis. Circ. Res. 2022, 130, 1647–1659. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Components | Dose | Function | Signal Passage | Ref. |
|---|---|---|---|---|---|
| Hypoxia-induced pulmonary hypertension in rats | Luteolin | 10–100 μmol/L, 28 days | Aortic ring relaxation; Mean pulmonary arterial hypertension ↓; | HIF-2α-Arg-NO axis ↓ and PI3K-AKT-eNOS-NO ↑ | [54] |
| H2O2-induced injury of HUVECs | Luteolin | 2.5–20 μM, pretreatment 2 h | Anti-oxidative stress; improves mitochondrial function | AMPK/PKC ↑; P38 MAPK/NF-κB ↓ | [55,56] |
| TNF-α-induced adhesion of human EA.hy 926 ECs | Luteolin | 0.5–20 μM, pretreatment 1 h | (MCP-1, ICAM-1, VCAM-1) ↓ | IKBα/NF-κB ↓ | [57] |
| TNF-α-induced C57BL/6 mice | Luteolin | Modified diet containing 93.93% luteolin | Anti-inflammatory | IKBα/NF-κB ↓ | [57] |
| HUVECs | Luteolin- 7-O-Glucoside | 20 μL, treatment for 48 h | Anti-oxidative stress; Anti-inflammatory; Anti-proliferation | JAK/STAT3↓; Nox4/ROS-NF-κB↓; MAPK ↓ | [58] |
| AngⅡ-induced injury of HUVECs | Baicalin | 6.25–50 μM | Anti-oxidative stress; Anti-apoptosis | Activation of the ACE2/Ang- (1-7)/Mas axis; PI3K/AKT/eNOS ↑ | [59] |
| Norbascine-induced pulmonary hypertension in rats | Baicalein | 10 mg/kg/day, 28 days | Anti-oxidative stress; Mean pulmonary arterial hypertension ↓ | Akt/ERK1/2/GSK3β/ β-catenin ↓; ET-1/ETAR ↓; ROS ↓ | [60] |
| LPS-induced rats | Baicalin | 50, 100 mg/kg/day, 3 days | Inhibited platelet hyperactivation; Anti-inflammatory; TSP1 ↓ | Furin/TGFβ1/Smad3/TSP-1 ↓ | [61] |
| TNF-α-induced injury of HUVECs | Baicalin | / | Anti-platelet adhesion; TSP1, ICAM-1 ↓ | AKT/Ca2+/ROS ↓ | [61] |
| TNF-α-induced injury of HUVECs | Quercetin | 10 μM; 30 μg/mL | Anti-inflammatory; anti-apoptosis E-selectin, VCAM-1, ICAM-1, IL-6, IL-8 ↓ | Activator protein 1 (AP-1) ↓ NF-κB ↓ | [62,63,64] |
| DF-induced inflammation of HUVECs | Quercetin | 5 μM | Anti-inflammatory | NRP2 -VEGFC complex ↓ | [65] |
| H2O2-induced injury of HUVECs | Quercetin–lycopene combination (molar ratio 5:1) | 8 μM, 12 h | Anti-oxidative stress; Anti-inflammatory | SIRT1-Nox4-ROS ↓ | [66] |
| High-fat diet (HFD)-fed ApoE−/− mice | Quercetin | 4 mg/day, 8 weeks | Anti-oxidative stress | NOX ↓; HO-1↑ | [67] |
| H/R-induced injury of HBMECs | Quercetin | 0.1–1 μmol/L, 8 h | Anti-oxidative stress; Enhance cell viability; Anti-apoptosis; ICAM-1, VCAM-1 ↓ | Keap1/Nrf2 ↑ | [68] |
| AngⅡ-infused hypertensive mice | EGCG | 50 mg/kg/day | Anti-oxidative stress; Systolic blood pressure ↓ | NOX ↓; BH4-eNOS-NO ↑ | [69] |
| Homocysteine-induced injury of HUVECs | EGCG | Pretreatment 2 h | Anti-oxidative stress; Anti-apoptosis | SIRT1/AMPK ↑; Akt/eNOS ↑ | [70] |
| H2O2-induced injury of HUVECs | EGCG | 1–10 μmol/L, pretreatment 24 h | Anti-oxidative stress; Induced autophagy | PI3K-AKT-mTOR ↓ | [71] |
| TNF-α-induced injury of human coronary artery endothelial cells (HCAECs) | EGCG | / | Anti-inflammatory | NF-κB ↓ | [72] |
| ox-LDL-induced injury of HUVECs | Naringin | 50, 100 μM, pretreatment 2 h | Anti-inflammatory; Anti-apoptosis; Anti-EndMT | Hippo-YAP ↓ | [73] |
| Homocysteine-induced injury of HUVECs | Naringenin | 200 μM, treatment for 24 h | Anti-oxidative stress; Reduced eNOS uncoupling | AMPKα/Sirt1 ↑ | [74] |
| High glucose (HG)- or free fatty acids (FFA)-induced apoptosis in HUVECs | Naringenin | 0–100 μM | Anti-apoptosis | PI3K/Akt and JNK1 ↑; Nrf2 ↑; HO-1 ↑ | [75] |
| HUVECs | Hesperidin | 1 μM, 2 h | Promoted NO production and expression of MasR | TRPV1-CaMKII/p38 MAPK/MasR ↑; TRPV1-CaMKII/eNOS/NO ↑ | [76,77] |
| Ox-LDL-induced senescence of HUVECs | Genistein | 1 μM, pretreatment 30 min | Induced autophagy; Anti-aging | SIRT1/LKB1/AMPK ↑ | [78] |
| H2O2-induced senescence of HUVECs | Genistein | 40, 80 μg/mL, 24 h | Anti-apoptosis; Anti-aging | TXNIP/NLRP3 ↓ | [79] |
| LPS-induced chronic vascular inflammatory response in mice | Genistein | 10 mg/kg/day, 20 weeks | Anti-inflammatory | miR-21/NF-κB p65 ↓ | [80] |
| Vascular endothelial cells (VECs) | Genistein | 10 μM, pretreatment 2 h | Anti-inflammatory | miR-21/NF-κB p65 ↓ | [80] |
| High glucose (HG)-induced injury of HUVECs | Blueberry anthocyanins | 5 μg/mL, pretreatment 24 h | Anti-oxidative stress; Induced vasodilation | PI3K/Akt ↑; PKCζ ↓ | [81] |
| Aged SD rats | Mulberry extract | 300 mg/kg | Anti-oxidative stress; Anti-aging; Reduced eNOS uncoupling | SIRT1 ↑; | [82] |
| PA-treated SV 40 transfected aortic rat endothelial cells (SVAREC) | Anthocyanin from red radish | 50, 100, 200, 400 μ g/mL, 24 h | Anti-apoptosis | p38 MAPK ↓ | [83] |
| HUVECs | Crocetin | 10, 20, 40 μM | Inhibited cell migration and angiogenesis | VEGFR2/SRC/FAK ↓ | [84] |
| HUVECs | Crocin | 100, 200, 400 μM | Inhibited cell migration and angiogenesis | VEGFR2/MEK/ERK ↓ | [84] |
| LPS-stimulated brain microvascular endothelial cells | Hydroxysafflor Yellow A | / | Prevented ZO-1 degradation and protected the blood–brain barrier | HIF-1α/NOX2 ↓ | [85] |
| U-46,619- and PE-inhibited rat MAs | Hydroxysafflor Yellow A | 10−7, 10−6, 10−5, 10−4 M | Increased Ca2+ influx and expanded blood vessels | TRPV4-coupled Ca2+/PKA/eNOS ↑ | [86] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Zhang, Q. Research Progress of Flavonoids Regulating Endothelial Function. Pharmaceuticals 2023, 16, 1201. https://doi.org/10.3390/ph16091201
Li H, Zhang Q. Research Progress of Flavonoids Regulating Endothelial Function. Pharmaceuticals. 2023; 16(9):1201. https://doi.org/10.3390/ph16091201
Chicago/Turabian StyleLi, Hao, and Qi Zhang. 2023. "Research Progress of Flavonoids Regulating Endothelial Function" Pharmaceuticals 16, no. 9: 1201. https://doi.org/10.3390/ph16091201
APA StyleLi, H., & Zhang, Q. (2023). Research Progress of Flavonoids Regulating Endothelial Function. Pharmaceuticals, 16(9), 1201. https://doi.org/10.3390/ph16091201