Abstract
The phenomenon of drug resistance in micro-organisms necessitates the search for new compounds capable of combating them. Fungi emerge as a promising source of such compounds as they produce a wide range of secondary metabolites with bacteriostatic or fungistatic activity. These compounds can serve as alternatives for commonly used antibiotics. Furthermore, fungi also accumulate compounds with antiviral activity. This review focuses on filamentous fungi and macrofungi as sources of antimicrobial compounds. The article describes both individual isolated compounds and extracts that exhibit antibacterial, antifungal, and antiviral activity. These compounds are produced by the fruiting bodies and mycelium, as well as the biomass of mycelial cultures. Additionally, this review characterizes the chemical compounds extracted from mushrooms used in the realm of cosmetology; specifically, their antimicrobial activity.
1. Introduction
In the kingdom of fungi, extensive biosynthetic capabilities are leading to the production of compounds with complex chemical structures that exhibit high biological activity. Operating as saprophytic organisms, fungi are strongly biochemically related to the composition of the substrate on which they reside. This fact largely determines their biosynthetic capabilities [1]. The main groups of biogenetic compounds originating from fungi can be divided into primary and secondary metabolites [2]. Primary metabolites are compounds derived from the primary metabolism of sugars (monosaccharides, disaccharides, polysaccharides, sugar alcohols, and quaternary amine bases). Secondary metabolites are compounds that can be further subdivided into compounds derived from the metabolism of active acetate (polyketides, isoprenoids, and sterols); compounds derived from the metabolism of fatty acids (polyacetylenes); compounds derived biogenetically from shikimic acid (phenols and phenolic acids); compounds formed from the transformation of amino acids (amines, toxic amines, alkaloids, and peptides); and compounds formed from the transformation of aromatic amino acids (ergot alkaloids) [2]. Another division of fungal metabolites includes nitrogenous compounds—including urea, amino acids, peptides, proteins, lectins, amines, alkaloids, indole derivatives, vitamins, purine compounds, isoxazole derivatives, and phenoxazine derivatives—as well as non-nitrogenous compounds such as carbohydrates, lipids, polyacetylenes, polyketides, isoprenoids, sterols, organic acids, and phenolic compounds [2,3].
The phenomenon of drug resistance among micro-organisms necessitates the search for new compounds capable of combating them. Fungi are a promising source of such compounds as they produce a wide range of secondary metabolites exhibiting bacteriostatic or fungistatic activity. These compounds can serve as alternatives to commonly used antibiotics. Furthermore, fungi also accumulate compounds with antiviral activity [4].
The development of new technologies, such as metabolomics, offers great potential for the identification and characterization of new natural antimicrobial compounds in the future. This knowledge may be crucial for the development of future therapeutic strategies. While the majority of natural compounds, including those stemming from fungal origin, do not have sufficient therapeutic effectiveness in monotherapy, their use in combination therapy with traditional antibiotics may contribute to the synergy effect of eliminating side effects and improving pharmacokinetic and pharmacodynamic properties [5].
Progress in fermentation technology, separation methodologies, and techniques for structural determination have rendered other micro-organisms appealing as potential reservoirs of novel bioactive secondary metabolites [6,7].
Among the many compounds that are fungal metabolites, many are used in cosmetology, but may also have important clinical significance if only in terms of increasing drug resistance. This problem also applies to aesthetic medicine. There are many biomaterials used in cosmetology and aesthetic medicine, as well as procedures that pose a potential risk of infection with multidrug-resistant strains. The compounds described here are examples of known natural substances whose application in the context of drug resistance may be important. Currently, researchers are looking for compounds of natural origin whose application could be considered in aspects of global problems. One of these is microbial drug resistance [8,9].
Categorizing antimicrobial compounds extracted from macro- and microfungi can depend on their chemical characteristics, modes of operation, or potential uses. Under the framework of classifying compounds by their chemical structure, we can highlight various antimicrobial agents, such as β-glucans present in macrofungi. Their primary mechanism centers around immune modulation. Additionally, we have the terpenoids, primarily originating from macrofungi, whose effects include the inhibition of microbial enzymes. An essential category comprises peptides, chiefly sourced from microfungi, which exert influence by impacting cell membrane permeability and disrupting cellular processes [10,11,12].
The objective of this review article is to present an updated and comprehensive overview of antimicrobial compounds derived from fungi, highlighting recent advancements in the field. The focus of this review encompassed both filamentous fungi and macromycetes. The paper describes the realm of antibacterial, antifungal, and antiviral compounds produced by both the fruiting bodies and mycelium (in the case of filamentous species) and those accumulated by the biomass of mycelial cultures. In addition, the paper delineates the characteristics of chemical compounds extracted from fungi with applications in cosmetology within the context of their antimicrobial activity. The main question posed in this review paper is as follows: how can fungi serve as a source of antimicrobial compounds to combat drug resistance of micro-organisms and what is the scope of their potential applications, including use in cosmetics?
2. Compounds of Fungal Origin with Antimicrobial Activity
2.1. Antibacterial Activity of Substances of Fungal Origin
2.1.1. Compounds of Fungal Origin with Antibacterial Activity
Numerous scientific studies indicate the antimicrobial activity of individual compounds and specific extracts obtained from fungal fruiting bodies.
It is believed that the presence of fungal fruiting bodies with such properties is due to defense mechanisms formed by fungi to survive in the environment. As the challenge of bacterial resistance to existing antibiotics grows, a variety of naturally occurring compounds exhibiting antimicrobial activity against pathogenic organisms is garnering increasing attention. Notably, one of the first compounds with antibacterial activity was the antibiotic substance sparassol, which was isolated from Sparassis crispa in 1920 [13] (Table 1). Over the following decades, the antibiotic activity of more than 2000 macromycetes species was subsequently validated.

Table 1.
Chemical structures of examples of compounds with antibacterial activity of fungal origin.
Key Antibacterial Compounds of Fungal Origin
Fungi are known for producing a wide variety of compounds endowed with antibacterial activity [6]. These compounds exhibit a dual nature, yielding a broad spectrum of activity while also manifesting selective efficacy against specific bacterial strains. Antibiotics are substances that hinder the growth and division of bacteria. The term “antibiotic” was coined by microbiologist Selman Waksman, who discovered two antibiotics: streptomycin and neomycin [26]. Nowadays, antibiotics encompass a spectrum spanning natural substances, semisynthetic derivatives, and synthetic analogs. These agents selectively target various bacterial structures, leading to either a bactericidal or bacteriostatic effect. Antibiotics are categorized into distinct groups based on factors such as their mode of action, chemical structure, or spectrum of activity. For instance, certain antibiotics inhibit the synthesis of bacterial cell walls (e.g., β-lactams), while others impede protein production (e.g., chloramphenicol, tetracycline), or interfere with bacterial RNA and DNA nucleic acids (e.g., quinolones) [27]. In the early 20th century, small doses of penicillin proved highly effective in controlling a significant proportion of bacterial infections. However, as the use of penicillin increased, its effectiveness waned against infections. This phenomenon, identified as antibiotic resistance, stems from the defense mechanisms micro-organisms develop to counteract antibiotics. It is important to note that antibiotic resistance is not a recent occurrence but rather an outcome of bacteria evolving various mechanisms to protect themselves from harmful substances within their environment. This process allows them to quickly adapt to adverse changes [28]. Filamentous fungi, particularly those belonging to the genera Penicillium, Cephalosporium, Aspergillus, and Fusidium, are vital organisms in pharmaceutical biotechnology, particularly in the pharmaceutical industry. These fungi are well-known prolific producers of antibiotics, and alongside actinomycetes, they are recognized as the primary sources of antibiotics [29]. Among the most significant antibiotic classes produced by fungi are penicillins, cephalosporins, fusidans, fusafungin, and fumigacin (helvolic acid).
- Penicillin
Penicillins belong to the β-lactam group of antibiotics (Figure 1). In terms of their chemical structure, they contain a thiazolidine ring conjugated with a β-lactam ring. The operational mechanism of penicillins, as well as other β-lactam antibiotics, entails binding to penicillin-binding proteins (PBPs), subsequently obstructing their function. Antibiotics within this grouping exhibit extremely low levels of general and organ toxicity against human cells because they only affect cells where peptidoglycan synthesis occurs. The commercial production of penicillins involves selected species such as Penicillium chrysogenum, Penicillium baculatum, Penicillium turbatum, Aspergillus persicinum, Aspergillus flavus, Aspergillus giganteus, Aspergillus nidulans, Aspergillus oryzae, and Aspergillus parasiticus [30,31]. Penicillins are primarily utilized in medicine to treat bacterial infections. They can be employed for addressing conditions such as impetigo, erysipelas, and acne, thereby contributing to the acceleration of wound healing [31].
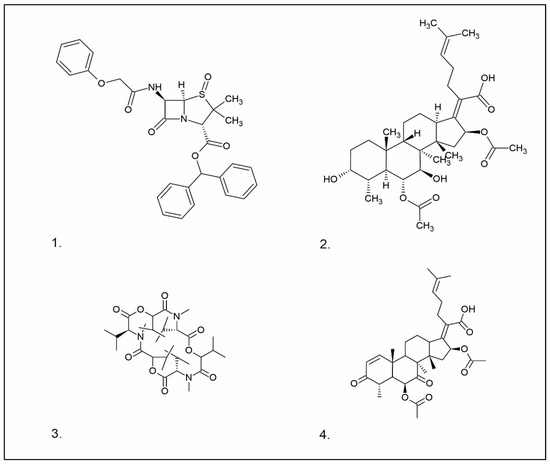
Figure 1.
Chemical structures of 1. Penicillin; 2. Cephalosporin; 3. Fusafungine; and 4. helvolic acid.
- Cephalosporin
The precursors of cephalosporins were initially isolated from cultures of Cephalosporium acremonium in 1948 by Giuseppe Brotzu [32,33] (Figure 1). The mechanism of action of cephalosporins is analogous to that of β-lactam antibiotics. These compounds find commercial production through strains of C. acremonium and Paecilomyces persicinius. Cephalosporins can be subdivided into several subgroups based on their chemical structure (P1–P5) [33]. Similar to all β-lactam antibiotics, cephalosporins inhibit the formation of bonds that connect peptidoglycan (murein) subunits, thus preventing the formation of a complete cell wall. They form covalent attachments to the active centers of bacterial enzymes, namely, carboxypeptidase and transpeptidase, leading to the inhibition of their actions. Consequently, they hinder the synthesis of bacterial cell walls. Cephalosporins treat bacterial infections of various origins, including both Gram-positive and Gram-negative bacteria. These antibacterial agents are employed in the treatment of infections caused by pathogens such as Staphylococcus aureus and Escherichia coli, among others [34]. Cephalosporins can be used to treat skin diseases caused by micro-organisms. In dermatology, they have been applied to address skin conditions such as folliculitis or postoperative infections [35].
- Fusidans
One of the most well-recognized fusidans is fusidic acid. Fusidic acid has an inhibitory effect on the protein synthesis of Gram-positive bacteria. Initially isolated in 1962 from Fusidium coccineum, fusidic acid was subsequently extracted from Mucor ramannianus and Isaria kogana [36,37]. Currently, biotechnological methods are obtained to derive fusafungin from species such as Calcarisporium arbuscula, Fusidium coccophilum, and Mortierella ramanniana [36]. Sporting a steroidal configuration, fusidic acid functions as an antibiotic with bacteriostatic properties. Its operational mechanism inhibits the synthesis of bacterial proteins, thereby preventing the growth and multiplication of bacterial cells. Notably, fusidic acid exhibits a narrow spectrum of activity, with a primary focus on Gram-positive bacteria, particularly those that are resistant to penicillin, such as Staphylococcus strains. The administration of fusidic acid during treatment might lead to the emergence of resistant strains of Staphylococcus. This antibiotic can be used in the form of creams and ointments for the topical treatment of infections such as impetigo, boils, inflammation of sweat glands and hair follicles, atrophy, acne vulgaris, and infections caused by the genus Staphylococcus spp. [37,38]. Of significance, fusidic acid can permeate intact skin, with the extent of penetration influenced by factors such as antibiotic exposure duration and skin condition. The biological half-life of fusidic acid is approximately 4–5 h. After being absorbed into the bloodstream, fusidic acid undergoes significant metabolism in the liver. While it is primarily excreted through the bile, a minor portion is eliminated unchanged in the urine [39]. While these compounds are not typically directly utilized in cosmetics due to their medical nature, they do exhibit effectiveness against pathogens responsible for skin diseases, such as S. aureus and Staphylococcus epidermidis [37].
Fusafungine emerges as a peptide antibiotic that exerts a bacteriostatic effect on numerous pathogenic micro-organisms (Figure 1). In addition to its antibacterial attributes, it also independently demonstrates anti-inflammatory activity. The probable mechanism of action involves enhancing the activity of NK cells, stimulating lymphocytes to produce IL-2, and inhibiting proinflammatory cytokines [40]. Fusafungine has proven efficacy in treating pharyngitis, offering an alternative to systemic antibiotics, steroids, or anti-inflammatory drugs. Sourced from the entomopathogenic fungus Fusarium lateritium (Ascomycota), this compound boasts an expansive activity spectrum without inducing bacterial resistance. As an ionophore antibiotic, it amalgamates enniatins and exhibits a unique ability to selectively form complexes with potassium cations, thereby transporting them across the lipid membranes of liposomes [41]. The topical application of fusafungine has been utilized, while its aerosol form has shown promise in treating inflammation of the upper and lower respiratory tract. Clinical trials have confirmed the effectiveness of the aerosolized form of this medication [41].
Fumigacin and helvolic acid (Figure 1) encompass antibiotics and phytotoxic substances produced by fungi belonging to the Ascomycota category, including Aspergillus fumigatus, Cephalosporium caeruleus, and Sarocladium oryzae (known as plant pathogens), alongside the species Emericellopsis terricola. Possessing distinctive properties and a varied range of action, fumigacin draws parallels to cephalosporins, especially those within the P1 group [42].
Selected Compounds of Fungal Origin from the Group of Isoprenoids, Peptides, and Acetylene Derivatives
Other substances with antibiotic properties, isolated from macrofungi, are compounds classified as isoprenoids, peptides, nucleosides, and acetylene derivatives.
- Isoprenoids
Isoprenoid compounds constitute a diverse group of secondary metabolites found in Basidiomycota. These compounds are intrinsically linked to the biogenetic pathway that originates from active acetate and proceeds through mevalonic acid, ultimately leading to the formation of “active isoprene.” Subsequent transformations of the latter undergo a series of transformations, resulting in the production of monoterpenes, sesquiterpenes, diterpenes, triterpenes, tetraterpenes, and steroids [43,44].
Merulidial, which contains an “unsaturated dialdehyde” functional group, emerges from liquid cultures of Merulius tremellosus (Table 1). This compound exhibits formidable activity against an array of Gram-positive bacteria, including Micrococcus roseus, Corynebacterium insidiosum, Bacillus brevis, Bacillus subtilis, Streptomyces viridochrontogenes, Sarcina lutea, and Arthrobacter citreus, as well as Gram-negative bacteria such as Proteus vulgaris [14,45]. Pilatin, a derivative of marasman (Table 1), is isolated from Flagelloscypha pilati. It proves effective in inhibiting the growth of Gram-negative bacteria, including Salmonella typhimurium, within a concentration range of 5–50 µg/mL [15]. From mycelial cultures of Pleurotellus hypnophilus, three metabolites with antibiotic activity have been identified. These include hypnophilin, pleurotellol, and pleurotellic acid (Table 1), all of which are sesquiterpenes derived from hirsutane. The common structural feature shared by all three metabolites is the α-methylenketone moiety [16]. Hypnophilin has been the subject of investigation due to its antimicrobial and antioxidant properties. Its potential utilization in skin care and cosmetics could be attributed to its antioxidant activity, which has the potential to safeguard the skin against oxidative stress and enhance overall skin health. Pleurotellol has been studied for its antibacterial and antifungal properties. In the context of skin care and cosmetics, pleurotellol’s antimicrobial activity could be relevant for formulations targeting skin conditions caused by microbial overgrowth [46]. Lentinellic acid, an iludane-type sesquiterpene (Table 1), exhibits robust antibacterial properties and has been isolated from two species of the genus Lentinellus: Lentinellus omphalodes and Lentinellus ursinus. It exhibits activity against Gram-positive bacteria, including B. brevis, Aerobacter aerogenes, and C. insidiosum, at concentrations ranging from 1 to 5 µL/mL [17]. Sesquiterpenoids with antimicrobial properties could potentially contribute to the development of novel skin care and cosmetic formulations aimed at addressing skin-related concerns caused by micro-organisms. Additionally, their potential antioxidant and anti-inflammatory activities may further enhance their suitability for cosmetic applications, promoting skin health and overall product quality. Lentinellic acid methyl ester also possesses antifungal activity [17]. In the context of skin care and cosmetics, this compound may have potential applications as a preservative in cosmetic formulations to help solve skin problems caused by fungal infections such as athlete’s foot or fungal acne [47].
Sulphurenic acid (Table 1) and eburicoic acid are triterpenes isolated from Laetiporus sulphureus [22]. Pleuromutilin, a diterpene compound, was isolated by Kavanagh in 1951 from a saprophytic fungus Clitopilus passeckerianus (formerly Pleurotus passeckerianus) (Table 1). Pleuromutilin and its derivatives inhibit bacterial protein synthesis by binding to the peptidyltransferase component of the 50S subunit of ribosomes [20]. Striatins A, B, and C are kyatan diterpenes isolated from Cyathus striatus (Table 1). These compounds exhibit antibiotic and cytotoxic effects at concentrations of 2 µg/mL. These compounds have been found in both the fruiting bodies and in vitro mycelium of the species. They demonstrate activity against various bacteria, including A. citreus, B. brevis, B. subtilis, E. coli, Leuconostoc mesenteroides, Mycobacterium phlei, Nocardia brasiliensis, P. vulgaris, Pseudomonas fluorescens, S. lutea, S. aureus, and Streptomyces viridochromogenes, along with the fungus Saccharomyces cerevisiae and the yeast Rhodotoula rubra. In the context of cosmetics and skin care, secondary metabolites such as striatins could have potential applications such as antimicrobial and antioxidant effects [21]. Armillaric acid, isolated from mycelial cultures of Armillaria mellea, is a sesquiterpene compound (Table 1) [18]. An aryl ester of this compound, known as melleolide, has exhibited antibacterial activity [48]. Flammulina velutipes mycelium has yielded four sesquiterpenes with antibacterial activity: enokipodins A, B, C, and D (Table 1). These compounds exhibit activity against B. subtilis, with enokipodins A and C also demonstrating activity against S. aureus Enokipodins, with a diverse range of biological properties that could have applications in cosmetics: antioxidant effects, skin brightening, and anti-inflammatory properties [19,49]. From the fruiting bodies of the saprophytic species Jahnoporus hirtus (Basidiomycota), a steroid named (24Z)-3,11-dioxolanosta-8,24-dien-26-oic acid has been isolated. This compound displays activity against Bacillus cereus and Enterococcus faecalis [50]. Ganomycin A and B (Table 1), isolated from Ganoderma pfeifferi, exhibit activity against B. subtilis, Micrococcus flavus, and S. aureus [24].
- Peptides
One of the peptides produced by fungi is plectasin, isolated from the fruiting bodies of Pseudoplectania nigrella (Ascomycota). Plectasin belongs to the defensin group of peptides and carries a cationic character. Comprising 40 amino acids, it exhibits activity against Gram-positive bacteria such as S. aureus and Streptococcus pneumoniae, primarily affecting the stability of their cell membranes [51]. In vitro, plectasin’s impact on S. pneumoniae mirrors that of penicillin and vancomycin. In addition, this peptide targets Gram-positive bacteria of genera such as Streptococcus (S. pneumoniae, S. pyogenes), Staphylococcus (S. aureus, S. epidermidis), Enterococcus (E. faecalis, E. faecium), Corynebacterium (C. diphtheriae, C. jeikeium), and Bacillus (B. cereus, B. thuringiensis) [52]. Zervamicins, a group of peptides with antibacterial activity, is produced by Emericellopsis salmosynnnemata (Ascomycota) (Figure 2). These peptides, classified as peptaibols, are linear and characterized by a high content of α,α-dialkyl amino acids, such as α-aminoisobutyric acid [53,54]. Another set of peptaibols includes peptaibol boletusin, peptaibol chrysospermin-3, and peptaibol chrysospermin-5, all extracted from Boletus spp. These compounds demonstrate efficacy against B. subtilis, Corynebacterium lilium, and S. aureus. Peptaibol chrysospermin-3 also shows activity against various Streptococcus strains [55]. Derived from the fungal fermentation of Tolypocladium niveum or Aspergillus terreus strains (Figure 2), cyclosporin is a cyclic peptide. The cyclosporin family comprises cyclic peptides with specific amino acids and demonstrates a mild antibiotic effect. However, their primary utility lies in their role as immunosuppressive agents. Cyclosporin A, for instance, is used in medicine to prevent organ rejection in transplant patients and treat autoimmune diseases [56]. Enniatins, cyclic hexapolipeptides, are synthesized by various strains of the Fusarium genus within the family Nectriaceae (Ascomycota) (Figure 2). In addition to their antibiotic properties, they exhibit insecticidal and anticancer activities [57].
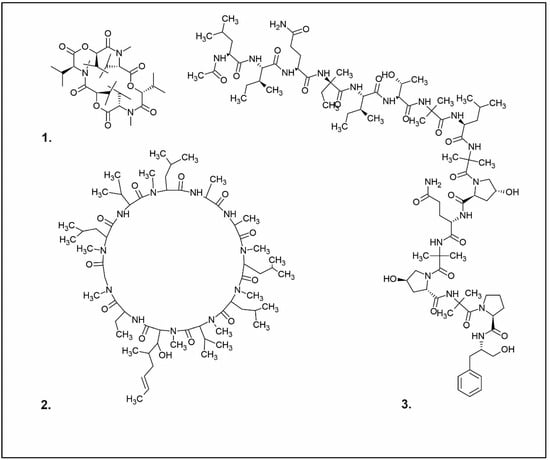
Figure 2.
Examples of antibacterial compounds with peptides structure: 1. enniatin; 2. Cyclosporin; and 3. Zervamicin.
An example of a nucleoside exhibiting antibacterial activity is nebularine, which has been isolated from Clitocybe nebularis, a saprotrophic toxic species [58]. Notably, an enzyme with multifaceted attributes, ribonuclease, is sourced from the edible species Pleurotus sajor-caju. This enzyme demonstrates antimicrobial, antimitogenic, and antiproliferative effects and exhibits activity against Pseudomonas aeruginosa and S. aureus, functioning by targeting RNA [59].
- Acetylene derivatives
Polyacetylenes stem from gradual desaturation processes involving saturated fatty acids, and they are a common occurrence within the fungal kingdom. Many acetylene derivatives identified in Basidiomycota exhibit antibacterial, cytotoxic, and antifungal activities, such as scorodonin, obtained from Marasmius scorodonius (Table 1), and 1-hydroxy-2-nonyn-3-one, extracted from Ischnoderma benzoinum [25]. Scorodonin derived from fungi holds potential for diverse cosmetic uses due to its antioxidant, anti-inflammatory, and skin-lightening properties [60]. Aqueveque has isolated two polyacetylenic compounds, namely, hepta-4,6-diyn-3-ol and 7-chloro-hepta-4,6-diyn-3-ol, from Gymnophilus spectabilis. These compounds are thought to arise from the desaturation pathway of saturated fatty acids and serve as precursors for the synthesis of polyacetylene compounds in fungi. Their antibacterial activity against Gram-positive and Gram-negative bacteria, as well as their antifungal activity, can be attributed to the presence of unsaturated triple bonds [61].
Other Compounds of Fungal Origin with Antibacterial Activity
Pleurotin, a derivative of quinone, along with leucopleurotin and dihydropleurotinic acid, has been isolated from Pleurotus griseus (now classified as Hohenbuehelia grisea). These compounds exhibit activity against Gram-positive bacteria and specific pathogenic fungi. Pleurotin has shown antimicrobial activity against certain bacteria and fungi. In the context of cosmetics, its antimicrobial properties could be explored for potential use as a natural preservative to prevent microbial growth in cosmetic products [62]. Oxalic acid, isolated from the mycelium of Lentinus edodes, shows activity against B. cereus, S. aureus, and E. faecalis [63]. Oxalic acid derived from fungi presents an array of cosmetic uses due to its exfoliating, brightening, antibacterial, and antioxidant properties [64]. Another compound, cloratin A, a benzoic acid derivative, has been isolated from the saprotrophic inedible fungus Xylaria intracolarata. This compound displays activity against E. coli, Klebsiella pneumoniae, P. aeruginosa, and Salmonella enterica, with particularly potent inhibitory activity observed against K. pneumoniae, surpassing the control group [65]. Antibacterial activity is also evident in anthraquinone derivatives such as 6–methylxanthopurpurin-3-O-methyl ether, (1 S, 3 S), austrocortilutein, (1 S, 3 R), austrocortilutein, (1 S, 3 S), austrocortirubin, and torosachryson, isolated from Cortinarius basirubencens. Compounds of erythroglaucine and emodin, isolated from other Cortinarius species, also demonstrated efficacy against S. aureus [66]. A fraction labeled B from Pycnoporus sanguineus, mainly composed of loccosee-3-one, exhibited activity against S. aureus and various strains of Streptococcus (A, B, C, and G). Compounds isolated from G. pfeifferi showed moderate activity against E. coli, Proteus mirabilis, and Serratia marcescens. Quinoline, isolated from the fungus Leucopaxillus albissimus, showed activity against Achromobacter xyloxidans, Acinetobacter baumannii, Burkholderia cenocepacia, Burkholderia loccose, Burkholderia multivorans, Cytophaga johnsonae, and P. aeruginosa, with the highest activity observed against C. johnsonae [67].
Polysaccharides, such as β-glucans, chitin, and its derivative chitosan, are vital components of the fungal cell wall. Chitosan, featuring amino sugars in its composition, exhibits antibacterial activity. Notably, chitosan is found not only in fungi but also present in the shells of arthropods such as crabs, shrimp, squid, and crayfish [68]. Exhibiting a wide antibacterial activity, chitosan proves effective against certain Gram-negative bacteria, Gram-positive bacteria, and fungi. Specifically, it has shown a higher effect on Gram-positive bacteria, including Listeria monocytogenes, Bacillus megaterium, B. cereus, S. aureus, Lactobacillus plantarum, L. brevis, and L. bulgaris. While it does display activity against Gram-negative bacteria, such as E. coli, P. fluorescens, S. typhimurium, and Vibrio parahaemolyticus, its potency is comparatively weaker [69,70].
Recent studies indicate that chitosan can be obtained biotechnologically from the cell wall of the filamentous fungus Rhizopus oryzae. Its antibacterial properties have been tested against E. coli, K. pneumoniae, and S. aureus [71,72]. A summary of the antibacterial activity of compounds derived from fungi is provided in Table 2.
Table 2.
Summary of antibacterial activity of compounds of fungal origin.
2.1.2. Extracts of Fungal Origin with Antibacterial Activity
A considerable number of studies have focused on evaluating the antibacterial activity of natural raw materials, often by investigating the analysis of complete extracts. Notably, several types of extracts have been extensively examined, including aqueous, ethanol, methanol, chloroform, dichloromethane, ether, and acetone extracts.
Ganoderma lucidum stands as a prominent fungal raw material in East Asian traditional medicine, including TCM [73]. Notably, diverse extracts including aqueous, ethanol, methanol, and acetone have demonstrated comparable efficacy against gentamicin sulfate, an aminoglycoside antibiotic. This effectiveness extends to various bacterial species: E. coli, S. aureus, K. pneumoniae, B. subtilis, S. typhimurium, and P. aeruginosa [74]. Other studies have confirmed that acetone extract of G. lucidum exhibits antibacterial activity, mainly against Gram-negative K. pneumoniae bacteria. Additionally, a synergistic interaction was observed when combining G. lucidum extracts with antimicrobial agents such as ampicillin, cefazolin, oxytetracycline, and chloramphenicol. This synergy was particularly pronounced with cefazolin against B. subtilis and Klebsiella oxytoca [75]. Conversely, a chloroform extract from the edible mycorrhizal fungus Hygrophorus agathosmus exhibited inhibition against various pathogenic bacteria, including E. coli, Enterobacter aerogenes, S. typhimurium, P. aeruginosa, S. aureus, S. epidermidis, and B. subtilis. Furthermore, this extract demonstrated inhibitory effects on Candida albicans and S. cerevisiae [76]. In a similar vein, a dichloromethane extract from Suillus collitinus observed activity against Gram-positive bacteria, including S. epidermidis and B. subtilis. This extract exhibited substantial antibacterial activity, with MIC values of 7.81 µg/mL, surpassing the reference antibiotic streptomycin (MIC = 15.62 µg/mL). For S. aureus, MIC values remained equal to those of streptomycin, at 15.62 µg/mL [76]. Finally, the methanolic extract of Hypholoma fasciculare, a saprotrophic poisonous fungus, exhibited notable antibacterial activity against Gram-positive bacteria such as B. cereus, B. subtilis, and S. aureus [77]. Turkoglu conducted a study investigating the antibacterial activity of ethanol extracts from L. sulphureus. The extract displayed inhibitory activity against the growth of Gram-positive bacteria, including B. subtilis, B. cereus, Micrococcus luteus, and M. flavus [78]. Another study analyzed the antibacterial activity of different extracts (chloroform, ethyl acetate, and water) from Lentinula edodes fruiting bodies. These extracts showed antibacterial activity against Streptococcus spp., Actinomyces spp., Lactobacillus spp., Prevotella spp., and Porphyromonas spp., which are known to cause various oral infections. Specifically, chloroform extracts exhibited bactericidal activity against both growing and resting bacterial cells of Streptococcus mutans and Prevotella intermedia, while the other two extracts exhibited bacteriostatic activity against both growing and resting bacterial cells of S. mutans and resting bacterial cells of P. intermedia [79]. Furthermore, a low molecular weight fraction study was conducted on an extract of L. edodes formulated as a mouthwash and administered to a group of volunteers [80]. Methanolic extract from the mycelium of Leucopaxillus giganteus, an inedible saprophytic species, showed antibacterial properties against Gram-positive bacteria in the order of potency: S. aureus > B. cereus > B. subtilis. This study also revealed that diammonium hydrogen phosphate was the preferred nitrogen source for enhancing the production of bioactive compounds inhibiting the growth of Gram-positive bacteria [81]. Studies on methanolic extracts of Phellinus rimosus and Navesporus loccose demonstrated moderate antibacterial activity against Gram-positive bacteria B. subtilis and S. aureus [82]. Ethanolic extracts from Pleurotus ostreatus and Meripilus giganteus exhibited broad-spectrum antibacterial activity, particularly against S. lutea [83]. Evaluating extracts from fruiting bodies and mycelial cultures of Trametes versicolor, researchers found varying antibacterial activity based on the type of solvent used for the extraction (water, organic solvents, or mixtures). The study revealed significant antibacterial activity against Gram-positive bacteria, with lower activity against Gram-negative bacteria. This effect was attributed to coriolin, a sesquiterpene compound found in Trametes (formerly Coriolus) spp. (Table 1). Extracts from Clavariadelphus loccose and T. versicolor have exhibited activity against a range of bacteria, including E. coli, E. aerogenes, S. typhimurium, S. aureus, and B. subtilis [76]. Aqueous extracts of Cordyceps sinensis and Cordyceps militaris, which are species that parasitize invertebrates, have demonstrated antibacterial activity against S. aureus, probably as a result of an increase in phagocytic macrophage activity and cytokine expression [83]. Ethanol extracts containing polysaccharides from Grifola loccose fruiting bodies have been tested against Gram-positive bacteria such as S. aureus, E. faecalis, B. cereus, L. monocytogenes, and Gram-negative bacteria such as E. coli, Salmonella enteritidis, Shigella sonnei, and Yersinia enterocolitica. The most notable antibacterial activity was observed against B. cereus [84]. Acetyl acetate extracts from various species growing in Brazil, including Phellinus sp., Gloeoporus thelephoroides, Hexagonia hydnoides, and Nothopanus hygrophanus, demonstrated inhibition of growth against bacteria such as B. cereus, L. monocytogenes, and S. aureus [85]. Aqueous, ethanol, methanol, and xylene extracts of Agaricus bisporus and P. sajor-caju, both saprophytic edible fungi, have shown antibacterial activity against E. coli, E. aerogenes, P. aeruginosa, and K. pneumoniae. Consumption of these fungi may provide natural protection against common pathogenic organisms [86]. Methanolic extracts of Hydnum repandum, an edible saprophytic species, have demonstrated activity against the Gram-negative bacteria P. aeruginosa [87]. The methanolic extract of the fruiting bodies of Lepista nuda, another edible fungus, exhibited antibacterial activity against E. coli and P. aeruginosa [88]. Dichloromethane extract from S. collitinus displayed activity against a range of bacteria, including E. coli, E. aerogenes, S. typhimurium, S. aureus, and S. epidermidis, B. subtilis, as well as C. albicans and S. cerevisiae [76,89]. Regarding L. sulphureus, both ethanolic and aqueous extracts from its fruiting bodies have shown antibacterial effects against various strains, including B. subtilis, B. cereus, M. luteus, M. flavus, and K. pneumoniae. Among these strains, M. flavus exhibited the highest susceptibility, while K. pneumoniae showed resistance. Although the efficacy of the active extracts was lower compared to commercial drugs, they still demonstrated potential as antibacterial agents. Furthermore, the aqueous extract of L. sulphureus fruiting bodies has shown antibacterial effects against M. flavus and L. monocytogenes [78,90]. Notably, the extract displayed significant efficacy against L. monocytogenes, a strain resistant to streptomycin [90]. A summary of the antibacterial activity of fungal extracts can be found in Table 3.
Table 3.
Summary of antibacterial activity of fungal origin extracts.
2.2. Antifungal Activity of Substances of Fungal Origin
2.2.1. Compounds of Fungal Origin with Antifungal Activity
Due to concerns over the toxicity of polyene antibiotics and synthetic azole derivatives, researchers have turned to exploring natural compounds with antifungal properties. This pursuit is driven by the increasing resistance of Candida species to traditional antifungal drugs, prompting the search for alternative resources. The antifungal activity of substances produced by fungi is attributed to both high-molecular-weight compounds such as proteins and peptides, as well as low-molecular-weight compounds, including terpenes (sesquiterpenes), steroids, and organic acids [9]. Numerous studies conducted in this field involve screening extracts derived from fungal materials [92]. Biforminic acid and biformin, a polyacetylene compound, are examples of substances with antifungal properties produced by the saprophytic fungus Trichaptum biforme. These compounds were among the earliest bioactive substances of fungal origin to be identified [93]. Griseofulvin is an antibiotic used for treating fungal infections in both humans and animals. Its mechanism of action involves inhibiting the cell divisions of dermatophytes belonging to the genera Microsporum, Epidermophyton, and Trichophyton. Griseofulvin is produced by various species of the Penicillium genus, particularly Penicillium griseofulvum, Penicillium aethiopicum, Penicillium janezewski, and Penicillium lanosus. Commercial production of griseofulvin involves biotechnological methods using Penicillium patulum [94,95]. During the fermentation processes leading to griseofulvin synthesis, other metabolites are formed, primarily intermediates in the antibiotic’s production, such as dehydrogriseofulvin, griseophenone A, griseoxanthone C, griseophenone Y, and dechlorogriseofulvinic acid. These metabolites might have significant implications in the search for new therapeutic agents. Griseofulvin’s mechanism of action involves inhibiting RNA biosynthesis and chitin synthesis, leading to damage to the fungal cell wall. It is commonly used for treating fungal skin, nail, and hair infections. When administered orally, it is well absorbed from the gastrointestinal tract and reaches peak concentration after about 4 h. The half-life of griseofulvin is 16–20 h, and the recommended dosage for treating superficial infections is typically 250 mg every 6 h. However, it is worth noting that griseofulvin has been discontinued from use [94,95]. Agrocybin, a peptide displaying activity against plant pathogens such as Mycosphaerella arachidicola and Fusarium oxysporum, was isolated from the fruiting bodies of Agrocybe cylindracea, a saprophytic Basidiomycota species [96]. Cordymin, a peptide, functions as an inhibitor of the growth of plant pathogens including Bipolaris maydis, M. arachidicola, Rhizoctonia solani, and C. albicans (with IC50 values of 50 μM, 10 μM, 80 μM, and 0.75 mM, respectively, for the fungal species). C. albicans, a yeast species (Saccharomycetes), is known to cause opportunistic infections in immunocompromised patients, while also constituting the natural flora of the digestive tract in 40–80% of the population. Cordymin was isolated from C. militaris, a parasitic species classified under Ascomycota [97]. Cordymin is well-known for its ability to improve the health and look of the skin. Its moisturizing qualities help in retaining skin hydration. This substance efficiently fights against oxidative stress triggered by environmental elements such as UV radiation and pollution. By counteracting damaging free radicals, cordymin helps in thwarting premature skin aging. Alongside its antioxidative attributes, cordymin also demonstrates anti-inflammatory effects and properties that promote skin regeneration [97].
In turn, screening studies aimed at identifying naturally occurring fungicides from fungi have demonstrated the potent antifungal impact of the ethanolic extract derived from the fruiting bodies of Albatrellus dispansus. Grifolin, the active compound isolated from these bodies, has been identified as the crucial component. Against Erysiphe graminis, Sclerotinia sclerotiorum, and Fusarium graminearum, it exhibited antifungal activity levels of 86.4% and 80.9%, respectively, at a concentration of 304.9 μM [98,99]. Cloratin A, sourced from X. intracolarata, displayed activity against Aspergillus niger (with an inhibition zone diameter of 15 mm) and C. albicans (with an inhibition zone diameter of 17 mm), comparable to the control substance nystatin, which also had an inhibition zone diameter of 17 mm. The diameter of the inhibition zone (IZD) serves as a reliable indicator of the antifungal activity present in the sample [100]. Lactarius rufus, an inedible mycorrhizal fungus, accumulates the sesquiterpene rufuslactone, an isomer of the previously described 3,8-oxa-13-hydroxylactar-6-en-5-lactaric acid γ-lactone from Lactarius necator. Rufuslactone exhibits antifungal properties against plant pathogens such as Alternaria alternata, Alternaria brassicae, Botrytis cinerea, and Fusarium graminearum [101]. Sesquiterpenoids Enokipodins F, G, and I were isolated from the mycelium of F. velutipes, a saprotrophic tree species. These compounds exhibit moderate activity against A. fumigatus, a pathogenic species affecting mammals, birds, and insects [49]. The protein isolated from the fruit bodies of Tricholoma giganteum exhibits antifungal activity against plant pathogens including Fusarium oxysporum, M. arachidicola, and Physalospora piricola. Furthermore, in the same study, trichogin was confirmed to inhibit HIV-1 reverse transcriptase with an IC50 value of 83 nM [102]. Oospolactone, sourced from Gloeophyllum sepiarium, possesses antifungal activity against Alternaria spp., Fusarium spp., Giberella spp., Penicillium spp., and Aspergillus spp. [103]. Lentin, isolated from L. edodes, also exhibits antifungal activity against M. arachidicola [104]. Compounds characterized by the acylcyclopentenodione structure, identified as chrysotriones A and B from Hygrophorus chrysodon, an edible saprophytic species, exhibit activity against Fusarium verticillioides [105]. Three sterol-structured compounds and five terpene-structured compounds were isolated from the fruiting bodies of Ganoderma annulare: 5-ergosta-7-en-3-ol; 5-ergosta-7,22-dien-3-ol; 5,8-epidioxy-5-ergost-6,22-dien-3-ol; and aplplanoxidic acids A, C, F, G, and H. All these compounds were subjected to antifungal activity testing against the dermatophytes Microsporum canis and Trichophyton mentagrophytes. Applanoxidic acid A displayed the highest level of activity [106]. A summary of the antifungal activity of fungal compounds is presented in Table 4.
Table 4.
Summary of antifungal activity of compounds of fungal origin.
2.2.2. Extracts of Fungal Origin with Antifungal Activity
The antifungal activity of both aqueous and ethanol extracts from A. bisporus was determined against A. flavus [111]. Ethanolic extracts and the protein fraction obtained from the mycelium of Ophiocordyceps sobolifera displayed potent antifungal effects against both pathogenic and saprophytic fungi, including C. albicans [112]. Methanolic extracts of A. bisporus, Agaricus bitorquis, and Agaricus sylvicola demonstrated antifungal activity against C. albicans and Candida tropicalis [113]. The chloroform extract of H. agathosmus exhibited antifungal activity against S. cerevisiae [76]. Additionally, the dichloromethane extract from S. collitinus showed activity against both S. cerevisiae and C. albicans [76]. Numerous fungal fruiting body extracts have been tested for their antifungal activity against C. albicans strains. Notably, the ethanolic extract derived from L. sulphureus fruiting bodies exhibited significant activity, with an inhibition zone diameter (IZD) measuring 21 ± 1 mm. This result surpassed the positive control, nystatin, which had an IZD of 19 mm [78].
Furthermore, the methanolic extract from Lactarius camphoratus, chloroform extract from L. edodes, and methanolic extract from Lepista nuda also exhibited activity against C. albicans [87]. Studies have revealed that extracts from G. lucidum inhibit the growth of micro-organisms responsible for surface mycoses caused by Pityrosporum ovale, S. epidermidis, and Propionibacterium acnes [106,110].
An aqueous extract sourced from L. sulphureus demonstrated potent antifungal activity, with ketoconazole serving as the benchmark antifungal drug. Although the L. sulphureus extract displayed a slightly milder effect in comparison to ketoconazole, it still held promising potential as an antifungal agent. Furthermore, the aqueous–ethanol extract extracted from L. sulphureus exhibited antifungal activity targeting a range of fungi, including A. niger, B. cinerea, F. oxysporum, Penicillium gladioli, and Sclerotinia sclerotiorum [114,115].
T. versicolor extracts exhibit not only antifungal activity but also antifungal activity. In a cross-sectional study that evaluated the antifungal activity of various Basidiomycota representatives, the methanolic extract derived from T. versicolor showed activity against A. fumigatus. However, no discernible activity against C. albicans was observed [91]. A summary of the antifungal activity of fungal extracts is provided in Table 5.
Table 5.
Summary of antifungal activity of extracts of fungal origin.
2.3. Extracts and Chemical Compounds of Fungal Origin with Antiviral Activity
The antiviral mechanisms of fungal-derived substances often involve blocking viral enzymes, disrupting nucleic acid synthesis, or indirectly boosting the immunostimulatory effects. While numerous chemical compounds with proven antiviral activity are registered drugs, ongoing intensive research aims to search for substances of natural origin, including those of fungal origin. The scientific literature broadly describes the antiviral effect of both fruiting body extracts and single, isolated compounds [117]. For instance, triterpenes such as ganoderiol, ganodermanontriol (Table 1), and ganodermic acid derived from G. lucidum exhibit activity against HIV-1 [110]. Similarly, ganodermadiol, lucidadiol, and lucidumol B obtained from Ganoderma pfeiferi demonstrate effectiveness against the influenza A virus [118]. Ganodermadiol also combats the herpes virus HSV-1 [110]. Phenolic compounds sourced from Inonotus hispidus exhibit activity against influenza viruses of types A and B [119]. Among the macromolecular compounds with antiviral activity isolated from fungi, the most noteworthy is the PSK complex (Krestin). This polysaccharide peptide, derived from the mycelium of T. versicolor, boasts anticancer and immunostimulatory properties. Scientific studies have confirmed the antiviral activity of PSK against cytomegalovirus and its ability to inhibit HIV replication [120].
In the study of natural substances, special attention has been given to the analysis of aqueous extracts. This is attributed to the logistical challenges and potential hazards associated with the utilization of organic solvents as extraction agents for raw materials. Compounds found within the fruiting bodies of species such as G. pfeifferi, Rozites caperata, and Agaricus brasiliensis have exhibited activity against herpes viruses. Notably, sulfated polysaccharides from A. brasiliensis, RC28 proteoglycan from R. caperata, and triterpenoids from G. pfeifferi (present in aqueous extracts) exhibit noteworthy antiviral potential. These compounds hold the ability to effectively counteract various stages of herpes virus replication [118,121,122].
Aqueous extracts containing polysaccharides and ethanol extracts sourced from Pleurotus pulmonarius fruiting bodies have demonstrated antiviral activity against the influenza A (H1N1pdm) virus [123]. Similarly, the acidic polysaccharide fraction obtained from C. militaris fruiting bodies has shown identical antiviral activity against the influenza A (H1N1) virus [124].
Aqueous–methanol extracts derived from the fruiting bodies of L. sulphureus have demonstrated inhibitory effects on HIV reverse transcriptase. This enzyme plays a crucial role in the transcription process, and its inhibition leads to the suppression of virus replication. The observed antiviral activity within the tested extracts is believed to be influenced by the presence of immunomodulatory polysaccharides [125]. Polysaccharides from A. brasiliensis show antiviral activity against poliovirus type (PV-1) [126]. In the case of polysaccharides EP-AV1 and EP-AV2 sourced from an aqueous extract of the fruiting body of Porodaedalea pini (also known as Phellinus pini), their presence inhibits plaque formation in Vero cells induced by herpes simplex virus 1 (HSV-1) and by Coxsackie virus B3 (CVB3) in HeLa cells. These polysaccharides have been demonstrated to affect the initial stage of virus replication [127].
Polyphenols were isolated from the ethanol extract of the fruiting bodies of Phellinus baumii. Through spectroscopic techniques, compounds including hispidin, hypholomine B, inoscavin A, davallialactone, and pelligridin D were identified. These compounds demonstrated inhibitory effects on the neuraminidase activity, an enzyme specific to the H1N1, H5N1, and H3N2 strains of the influenza virus. Additionally, they exhibited a reduction in the virus-induced cytopathic effect (CPE). Neuraminidase serves as an enzyme that allows viruses to exit cells by breaking down the cell membrane of an infected cell. It also plays a role in facilitating virus attachment to cell membranes, aiding their entry into the cell due to its high affinity for the sialic acid of membrane receptors [128]. Laccase isolated from Pleurotus ostreatus and tyrosinase from A. bisporus show activity against HCV. Laccase from P. ostreatus has been shown to block viral entry and replication into PBMC and HepG2 cells, while tyrosinase from A. bisporus inhibits viral replication into replicon-containing Huh-5-2 cells [129]. Another noteworthy species that show significant antiviral activity is Grifola frondosa. The main active compound is β-glucan (GF-D). It has been shown that a combination of GF-D with IFN human interferon α–2b could potentially offer effective therapy against chronic HBV infections [130,131]. In 2018, structural identification of lentinan from L. edodes mycelium LNT-1 was conducted, followed by an investigation of its antiviral activity against hematopoietic necrosis virus (IHNV) [132]. Notably, its immunostimulatory activity was also demonstrated. As proven, the innate immune response is a critical factor in the course of COVID-19 disease. COVID-19 patients show high titers of inflammatory cytokines, so the effect of LNT-1 on SARS-CoV-2 should be considered [133].
A potential candidate in the battle against SARS-CoV-2 is Inonotus obliquus, commonly known as the chaga fungus, which possesses a robust enzyme system and defense mechanism due to its parasitic lifestyle [133]. SARS-CoV-2, the virus responsible for COVID-19, primarily targets the human respiratory system and other vital organs. Currently, no specific treatment for SARS-CoV-2 infection exists, although certain drugs have displayed potential efficacy in inhibiting the virus. Natural substances, including fungi, have exhibited potent antiviral and anti-inflammatory effects positioning them as promising candidates for effective COVID-19 treatments [133]. I. obliquus, commonly found in Asia, Europe, and North America, serves as a widely utilized natural resource for various ailments. A specific polysaccharide fraction derived from I. obliquus, named IOP, has shown the ability to inhibit the production of NO and similar cytokines associated with COVID-19 [133]. COVID-19 patients often experience inflammatory responses, resulting in elevated plasma levels of cytokines and leukocytes. Since IOPs have shown promising results in treating various viral diseases, their potential effect on COVID-19 infection holds considerable promise. Furthermore, an aqueous extract of I. obliquus has demonstrated virucidal activity against the hepatitis C virus, remarkably reducing its infectivity by 100-fold within a span of 10 min [134,135]. A summary of the antiviral activity of compounds and fungal extracts is provided in Table 6.
Table 6.
Summary of antiviral activity of compounds and extracts of fungal origin.
3. Cosmetic Applications of Antimicrobial Fungal-Derived Compounds
Antimicrobial fungal-derived compounds have gained significant attention in various fields, including cosmetics, due to their potential benefits in preventing and treating microbial infections. These compounds offer several applications within cosmetic formulations, particularly in skin care and hair care products (Figure 3) [10,11,12,138,139].
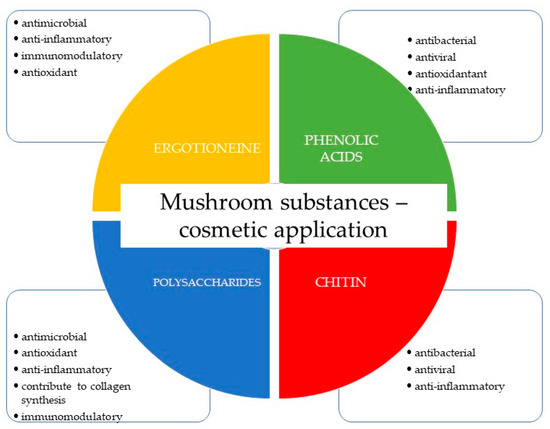
Figure 3.
Cosmetic applications of ingredients of mushroom origin are the most used.
The antimicrobial properties of mushrooms have been extensively explored in recent studies, revealing their potential as valuable sources of natural compounds with antibacterial and antioxidant activities. In the study by Eiamthaworn et al., focused on Cordyceps militaris extracts and determining the biological activity of extracts against skin pathogenic bacteria, results demonstrate that Cordyceps militaris extracts exhibit significant antibacterial activity against various skin pathogens. This finding underscores the potential of mushrooms as a natural source of antimicrobial agents, holding promise for the development of therapeutic solutions targeting skin infections [140]. In turn, Taofiq et al. describe the development of mushroom-based cosmeceutical formulations. These formulations are designed to possess not only antimicrobial properties but also anti-inflammatory, antityrosinase, and antioxidant activities. The study reveals the multifunctional nature of mushroom compounds, highlighting their potential to address multiple skin-related concerns simultaneously. This research supports the notion that mushrooms can serve as versatile ingredients in skin care products, offering holistic benefits beyond conventional antibacterial effects. The studies suggest that mushroom-derived compounds can contribute to the creation of effective solutions for skin care, particularly in combatting skin infections, reducing inflammation, and providing antioxidant support [139].
3.1. Potential Practical Applications
One of the foremost practical applications within the realm of mushroom-derived bioactive compounds lies in their preservative efficacy. Fungal metabolites bearing antimicrobial attributes offer a compelling natural substitute for conventional synthetic preservatives, thereby augmenting the longevity of cosmetic formulations [141].
Furthermore, the utility of mushroom metabolites extends to formulations tailored for acne-prone or sensitive cutaneous conditions. Their antimicrobial prowess can be harnessed to effectively modulate the proliferation of bacteria associated with acne pathology.
Moreover, the strategic integration of fungal metabolites with antimicrobial characteristics into sun protection formulations represents an additional avenue of application. Such products offer a comprehensive approach to skin well-being, amalgamating solar safeguarding, cutaneous hydration preservation, and antimicrobial properties [142].
3.2. Microbiological Factors Causing Skin Diseases
Numerous cutaneous disorders, such as acne and fungal infections, stem from pathogenic micro-organisms. Mushroom-derived metabolites possessing potent antibacterial and antifungal attributes hold promising potential in counteracting these infections. Through their incorporation into topical formulations, the capacity arises to selectively target and eradicate detrimental micro-organisms, concurrently fostering skin health. In the realm of bacterial-induced skin maladies, exemplars such as S. aureus prevail. This species is a prevalent instigator of conditions ranging from boils and impetigo to cellulitis, and even more severe manifestations such as methicillin-resistant S. aureus (MRSA) infections. Research endeavors are oriented towards comprehending the virulence determinants and mechanisms that facilitate S. aureus’ cutaneous colonization and invasiveness, with the aim of developing efficacious antibiotic interventions [143]. Additionally, Streptococcus pyogenes, recognized as Group A Streptococcus (GAS), emerges as a causative agent for afflictions including impetigo, erysipelas, and cellulitis, warranting meticulous scrutiny to unravel its pathogenic intricacies [144]. Similarly, P. acnes assumes prominence as the causal agent of acne vulgaris, a prevalent dermatological concern. Scientific inquiry has delved into deciphering the role of P. acnes in acne genesis, its interplay with the immune system, and potential therapeutic modalities targeting this bacterium [145]. Among dermatophytes that assail the integument, noteworthy is the prevalence of Trichophyton species, attributed to a spectrum of fungal skin infections spanning tinea corporis, tinea pedis (athlete’s foot), and tinea cruris (jock itch). Another cluster of fungi, encompassing Microsporum and Epidermophyton, contributes to parallel conditions such as athlete’s foot. Comprehensively exploring mushroom-derived metabolites endowed with potent antibacterial and antifungal characteristics, with the goal of addressing this gamut of micro-organisms, underscores an auspicious avenue in advancing therapeutic approaches for diverse cutaneous infections [146].
3.3. Selected Compounds of Fungal Origin with Cosmetic Properties
Ergothioneine, an exogenous amino acid derived from thiourea-histidine, emerges as a notable example. This compound boasts high solubility in water and stability at high temperatures. The primary source of ergothioneine lies within macrofungi. Its ability to chelate metal ions aids in preventing the buildup of reactive oxygen species in the body. Furthermore, ergothioneine exhibits antimicrobial and anti-inflammatory properties while contributing to the regulation of immune system function [12,138]. Ergothioneine, derived from fungi, has found extensive applications in cosmetic products. It offers protective properties against mitochondrial DNA damage in fibroblasts and keratinocytes, thereby safeguarding the skin against the harmful effects of free radicals. Its principal application resides in antiaging preparations. In the realm of cosmetology, ergothioneine extracted from A. bisporus, G. frondosa, and L. edodes stands as a widely employed resource [10].
Chitin forms the cell wall of mushrooms, comprising a complex structure of poly-N-acetyl-1,4-D-glucosamine subunits linked by β-(1→4) glycosidic bonds. This compound exhibits insolubility in water while displaying hygroscopic and sorptive characteristics. Remarkably, chitin showcases properties including anti-inflammatory, antiviral, and antibacterial properties [147]. Chitin finds application in cosmetic products, mainly prized for its moisturizing effects. It offers antiaging, firming, and regenerative benefits, reinstating skin elasticity, delivering antioxidant effects, and stimulating collagen synthesis. Its presence is particularly prominent in formulations tailored towards mature and dry skin, effectively maintaining appropriate hydration levels [148].
Chitosan, a deacetylated derivative of chitin, exhibits an affinity for keratin and can create bonds with amino acids. Even at low concentrations, it can create an elastic film that is resistant to moisture. These traits render chitosan suitable for various products, including hair care products [149]. Within both filamentous and macromycetes fungi, polysaccharides are prevalent, and their biological activity is determined by their chemical structures. Predominantly represented by glucans, fungal polysaccharides can also possess galactan or mannans structures. The type of glycosidic bond (α or β) and the arrangement of the polysaccharide molecule significantly impact their biological activity. Of particular note are β-glucans, which exhibit primary biological activity. These polysaccharides often possess substantial molecular weights ranging from 100 to 1000 kDa. The most active polysaccharides tend to sport a linear structure, devoid of long side chains, facilitating enhanced solubility and assimilation. The primary chain typically contains β-(1→3) bonds, with branching occurring through β-(1→6) bonds [147]. Some polysaccharides might even incorporate an additional peptide fragment. Higher fungi-derived polysaccharides have primarily been employed as immunostimulants in the noninvasive treatment of cancer, with lentinan serving as a prime example. These polysaccharides also exhibit diverse effects encompassing antiviral, antibacterial, antiparasitic, anti-inflammatory, hypotensive, hypoglycemic, and vasoprotective effects [69]. Mushroom-derived polysaccharides in cosmetic preparations are utilized for their antioxidant properties. These compounds contribute to collagen synthesis, protect skin cells from UV radiation, demonstrate anti-inflammatory and antimicrobial effects, and are employed in cosmetics for their moisturizing effects [72].
Phenolic acids, notably gallic acid, not only exhibit antioxidant properties but also wield anti-inflammatory, antibacterial, and antiviral properties. Gallic acid finds application in combating pigmentary disorders, which stand among the most common dermatological conditions [150].
Vitamin E, a term often used to depict a collection of closely related compounds named tocopherols and tocotrienols, possesses a shared structure featuring a chromanol and an isoprene side chain. This vitamin is recognized for its capability to scavenge loose radicals, which, in turn, is thought to safeguard against degenerative ailments such as cancer and cardiovascular disorders. Moreover, vitamin E showcases prowess in combatting diverse micro-organisms and impeding their growth. This valuable attribute of vitamin E contributes to its potential role in bolstering immune system well-being and fortifying the body’s resistance against infections. In the realm of cosmetology, vitamin E holds significant sway due to its favorable impacts on the skin, encompassing antioxidant, moisturizing, regenerative, UV radiation shielding, anti-inflammatory, and anti-aging properties [151]. As a potent antioxidant, vitamin E diligently counters free radicals that pose harm to skin cells and expedite the aging process. It erects a safeguarding barrier to stave off moisture loss, thereby preserving skin suppleness and thwarting desiccation. The vitamin aids in the regeneration of compromised skin cells and exhibits particular efficacy in reducing the visibility of scars. While not a replacement for sunscreen, vitamin E furnishes a measure of defense against UV-induced harm. Scientifically established as a formidable antioxidant with anti-inflammatory attributes, vitamin E further amplifies its merits. Furthermore, the vitamin’s adeptness in fending off oxidative stress and facilitating skin restoration enhances its prowess in the battle against aging. Mushrooms stand as a natural wellspring of vitamin E. Among the species that have been scrutinized for their tocopherol and tocotrienol content, Lentinula edodes, Agaricus bisporus, Pleurotus ostreatus, and Grifola frondosa [152].
Inhibiting tyrosinase, the enzyme responsible for melanin formation, is a pivotal strategy for skin brightening. Many substances used for skin brightening modify tyrosinase activity [139]. Among compounds derived from fungi boasting skin-whitening qualities are kojic acid, azelaic acid, and 3,4-dihydroxybenzaldehyde [153]. Kojic acid is a crucial inhibitor of melanin biosynthesis, mirroring the action of hydroquinone. It functions by obstructing tyrosinase activity while additionally offering antimicrobial and antioxidant properties. This compound is primarily produced by fungal species belonging to the Aspergillus and Penicillium genera [154]. Azelaic acid stands as another example of a compound influencing tyrosinase activity. It is produced, among other sources, by the species P. ovale (Basidiomycota). Azelaic acid’s effectiveness against hyperpigmentation has been substantiated, even surpassing that of hydroquinone. In addition, it is characterized by keratolytic, anti-inflammatory, and antibacterial effects [139].
4. Conclusions
The escalating prevalence of infectious diseases has escalated into a global concern, primarily attributed to the increasing resistance that micro-organisms and viruses have developed against conventional antimicrobial drugs utilized for both therapeutic and preventive purposes. In response to this critical challenge, there has been a notable surge in the exploration of natural sources endowed with potent antimicrobial properties. Within the scientific literature, fungi emerge as compelling candidates, showcasing robust antimicrobial potential that spans across a diverse spectrum, encompassing Gram-positive and Gram-negative bacterial strains, fungal pathogens, and even viruses. This burgeoning interest in fungi’s antimicrobial capabilities is particularly intriguing in the context of the cosmetics industry, which is actively harnessing these therapeutic attributes to elevate the quality and efficacy of its products. A comprehensive assessment of conducted studies underscores the remarkable presence of substances with substantial antimicrobial activity within mushrooms, emblematic representatives of the fungal kingdom. These bioactive compounds hold immense promise for diverse applications, particularly in the formulation of skin care products tailored to address a range of persistent skin conditions. Furthermore, the therapeutic potential of these fungal substances extends beyond cosmetics, demonstrating effectiveness in the treatment of various dermatological diseases. This signifies a significant stride towards merging the realms of natural antimicrobial agents derived from fungi with the intricacies of dermatological care.
Author Contributions
Conceptualization, K.S.-Z.; investigation, K.S.-Z. and M.T.; writing—original draft preparation, K.S.-Z., M.T., A.O.-J. and P.N.; writing—review and editing, K.S.-Z., M.Z., K.K. and B.M.; visualization, K.S.-Z. and M.Z.; supervision K.S.-Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by projects supported by the Polish Ministry of Science and Higher Education: Grants PL: N42/DBS/000271.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Deacon, J. Introduction: The Fungi and Fungal Activities. In Fungal Biology; Blackwell Pub: New York, NY, USA, 2006; pp. 1–15. ISBN 978-1-405-13066-0. [Google Scholar]
- Kohlmünzer, S. Farmakognozja. Podręcznik Dla Studentów Farmacji; Wydawnictwa Lekarskie PZWL: Warszawa, Poland, 1998. [Google Scholar]
- Chen, H.P.; Liu, J.K. Secondary Metabolites from Higher Fungi. Prog. Chem. Org. Nat. 2017, 106, 1–201. [Google Scholar]
- Suay, I.; Arenal, F.; Asensio, F.J.; Basilio, A.; Cabello, M.A.; Díez, M.T.; García, J.B.; González del Val, A.; Gorrochategui, J.; Hernández, P. Screening of basidiomycetes for antimicrobial activities. Anton. Leeuw. Int. J. G 2000, 78, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, D.; Dussart, F. Selected fungal natural products with antimicrobial properties. Molecules 2020, 25, 911. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, K.; Verma, P. Fungal Metabolites: A Recent Trend and Its Potential Biotechnological Applications. In New and Future Developments in Microbial Biotechnology and Bioengineering: Recent Advances in Application of Fungi and Fungal Metabolites: Current Aspects; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–14. [Google Scholar]
- Di Hsu, K.; Cheng, K.C. From nutraceutical to clinical trial: Frontiers in Ganoderma development. Appl. Microbiol. Biotechnol. 2018, 102, 9037–9051. [Google Scholar] [CrossRef]
- Shah, R.A.; Hsu, J.I.; Patel, R.R.; Mui, U.N.; Tyring, S.K. Antibiotic resistance in dermatology: The scope of the problem and strategies to address It. J. Am. Acad. Dermatol. 2022, 86, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.; Kumar, V.; Naik, B.; Verma, A.; Saris, P.E.J.; Kumar, V.; Gupta, S. Antifungal metabolites, their novel sources, and targets to combat drug resistance. Front. Microbiol. 2022, 13, 1061603. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Bahkali, A.H.; Moslem, M.A. Fungi—An unusual source for cosmetics. Fungal. Divers. 2010, 43, 1–9. [Google Scholar] [CrossRef]
- Wu, Y.; Choi, M.H.; Li, J.; Yang, H.; Shin, H.J. Mushroom cosmetics: The present and future. Cosmetics 2016, 3, 22. [Google Scholar] [CrossRef]
- Bowe, W.P.; Pugliese, S. Cosmetic benefits of natural ingredients. J. Drugs. Dermatol. 2014, 13, 1021–1025. [Google Scholar]
- Sharma, N.; Tapwal, A.; Verma, R.; Kumar, D.; Nepovimova, E.; Kuca, K. Medicinal, nutritional, and nutraceutical potential of Sparassis Crispa s. lat.: A review. IMA Fungus 2022, 13, 8. [Google Scholar] [CrossRef]
- Quack, W.; Anke, T.; Oberwinkler, F.; Giannetti, B.M.; Steglich, W. Antibiotics from Basidiomycetes. V. Merulidial, a new antibiotic from the basidiomycete Merulius tremellosus Fr. J. Antibiot. 1978, 31, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Heim, J.; Anke, T.; Mocek, U.; Steffan, B.; Steglich, W. Antibiotics from Basidiomycetes. XXIX: Pilatin, a new antibiotically active marasmane derivative from cultures of Flagelloscypha pilatii Agerer. J. Antibiot. 1988, 41, 1752–1757. [Google Scholar] [CrossRef] [PubMed]
- Kupka, J.; Anke, T.; Giannetti, B.M.; Steglich, W. Antibiotics from Basidiomycetes. XIV. Isolation and biological characterization of hypnophilin, pleurotellol, and pleurotellic acid from Pleurotellus hypnophilus (Berk.) Sacc. Arch. Microbiol. 1981, 130, 223–227. [Google Scholar] [CrossRef]
- Stärk, A.; Anke, T.; Mocek, U.; Steglich, W.; Kirfel, A.; Will, G. Lentinellic acid, a biologically active protoilludane derivative from Lentinellus species (Basidiomycetes). Z Naturforsch. C J. Biosci. 1988, 43, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Obuchi, T.; Kondoh, H.; Watanabe, N.; Tamai, M.; Omura, S.; Jun-Shan, Y.; Xiao-Tian, L. Armillaric acid, a new antibiotic produced by Armillaria mellea. Planta. Med. 1990, 56, 198–201. [Google Scholar] [CrossRef]
- Tabuchi, A.; Fukushima-Sakuno, E.; Osaki-Oka, K.; Futamura, Y.; Motoyama, T.; Osada, H.; Ishikawa, N.K.; Nagasawa, E.; Tokimoto, K. Productivity and bioactivity of enokipodins A–D of Flammulina rossica and Flammulina velutipes. Biosci. Biotechnol. Biochem. 2020, 84, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Novak, R.; Shlaes, D.M. The pleuromutilin antibiotics: A new class for human use. Curr Opin Investig Drugs 2010, 11, 182–191. [Google Scholar]
- Hofle, G.; Oberwinkler, F. The striatins-new antibiotics from the Basidiomycete Cyathus striatus (Huds. Ex Pers.) Willd. J. Antibiot. 1977, 30, 221–225. [Google Scholar]
- Šiljegović, J.D.; Stojković, D.S.; Nikolić, M.M.; Glamočlija, J.M.; Soković, M.D.; Ćirić, A.M. Antimicrobial activity of aqueous extract of Laetiporus sulphureus (Bull.: Fr.) Murill. Zb Matice. Srp. Prir. Nauk. 2011, 299–305. [Google Scholar] [CrossRef]
- Ma, B.; Ren, W.; Zhou, Y.; Ma, J.; Ruan, Y.; Wen, C.N. Triterpenoids from the spores of Ganoderma lucidum. N. Am. J. Med. Sci. 2011, 3, 495–498. [Google Scholar] [CrossRef]
- Mothana, R.A.A.; Jansen, R.; Jülich, W.D.; Lindequist, U. Ganomycins A and B, new antimicrobial farnesyl hydroquinones from the Basidiomycete Ganoderma pfeifferi. J. Nat. Prod. 2000, 63, 416–418. [Google Scholar] [CrossRef] [PubMed]
- Anke, T.; Kupka, J.; Schramm, G.; Steglich, W. Antibiotics from Basidiomycetes. X. Scorodonin, a new antibacterial and antifungal metabolite from Marasmius scorodonius (Fr.) Fr. J. Antibiot. 1980, 33, 463–467. [Google Scholar] [CrossRef]
- Clardy, J.; Fischbach, M.A.; Currie, C.R. The natural history of antibiotics. Curr. Biol. 2009, 19, R437–R441. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.; Truman, A.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Dutta, B.; Lahiri, D.; Nag, M.; Ghosh, S.; Dey, A.; Ray, R.R. Fungi in pharmaceuticals and production of antibiotics. In Applied Mycology: Entrepreneurship with Fungi; Springer International Publishing: Cham, Germany, 2022; pp. 233–257. [Google Scholar]
- Conrado, R.; Gomes, T.C.; Roque, G.S.C.; De Souza, A.O. Overview of bioactive fungal secondary metabolites: Cytotoxic and antimicrobial compounds. Antibiotics 2022, 11, 1604. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. β-lactams and β-lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.L. The penicillins: A review and update. J. Midwifery Womens Health 2002, 47, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Bo, G. Giuseppe Brotzu and the discovery of cephalosporins. Clin. Microbiol. Infect. 2000, 6, 6–8. [Google Scholar] [CrossRef]
- Fernandez, J.; Jimenez-Rodriguez, T.W.; Blanca-Lopez, N. Classifying Cephalosporins: From generation to cross-reactivity. Curr. Opin. Allergy Clin. Immunol. 2021, 21, 346–354. [Google Scholar] [CrossRef]
- Khan, D.A.; Banerji, A.; Bernstein, J.A.; Bilgicer, B.; Blumenthal, K.; Castells, M.; Ein, D.; Lang, D.M.; Phillips, E. Cephalosporin allergy: Current understanding and future challenges. J. Allergy Clin. Immunol. Pract. 2019, 7, 2105–2114. [Google Scholar] [CrossRef]
- Harrison, C.J.; Bratcher, D. Cephalosporins: A review. Pediatr. Rev. 2008, 29, 264–273. [Google Scholar] [CrossRef]
- Cordes, M.G. Fusidic Acid. In xPharm: The Comprehensive Pharmacology Reference; Elsevier: Amsterdam, The Netherlands, 2007; pp. 1–4. [Google Scholar]
- Schöfer, H.; Simonsen, L. Fusidic acid in dermatology: An updated review. Eur. J. Dermatol. 2010, 20, 006–015. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Ji, W.; Zhang, D.; Zhu, Y.; Bi, Y. Bioactivities and structure-activity relationships of fusidic acid derivatives: A review. Front. Pharmacol. 2021, 12, 759220. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P. Fusidic Acid: A bacterial elongation factor inhibitor for the oral treatment of acute and chronic staphylococcal infections. Cold Spring Harb. Perspect. Med. 2016, 6, a025437. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.; Bluzat, A.; Seigneuret, M.; Rigaud, J.L. Alkali cation transport through liposomes by the antimicrobial fusafungine and its constitutive enniatins. Biochem. Pharmacol. 1995, 50, 2105–2107. [Google Scholar] [CrossRef] [PubMed]
- Lund, V.J.; Grouin, J.M.; Eccles, R.; Bouter, C.; Chabolle, F. Efficacy of fusafungine in acute rhinopharyngitis: A pooled analysis. Rhinology 2004, 42, 207–212. [Google Scholar] [PubMed]
- Tschen, J.S.M.; Chen, L.L.; Hsieh, S.T.; Wu, T.S. Isolation and phytotoxic effects of helvolic acid from plant pathogenic fungus Sarocladium Oryzae. Bot. Bull. Acad. Sin. 1997, 38, 251–256. [Google Scholar]
- Savidov, N.; Gloriozova, T.A.; Poroikov, V.V.; Dembitsky, V.M. Highly oxygenated isoprenoid lipids derived from fungi and fungal endophytes: Origin and biological activities. Steroids 2018, 140, 114–124. [Google Scholar] [CrossRef]
- Ninkuu, V.; Zhang, L.; Yan, J.; Fu, Z.; Yang, T.; Zeng, H. Biochemistry of terpenes and recent advances in plant protection. Int. J. Mol. Sci. 2021, 22, 5710. [Google Scholar] [CrossRef]
- Anke, H.; Sterner, O.; Steglich, W. Structure-activity relationships for unsaturated dialdehydes. Mutagenic, antimicrobial, cytotoxic, and phytotoxic activities of merulidial derivatives. J. Antibiot. 1989, 42, 738–744. [Google Scholar] [CrossRef]
- Mehta, G.; Murthy, A.S.K. The first total synthesis of the novel triquinane natural products pleurotellol and pleurotellic acid. Tetrahedron. Lett. 2003, 44, 5243–5246. [Google Scholar] [CrossRef]
- Cadelis, M.M.; Copp, B.R.; Wiles, S. A Review of fungal protoilludane sesquiterpenoid natural products. Antibiotics 2020, 9, 928. [Google Scholar] [CrossRef] [PubMed]
- Midland, S.L.; Izac, R.R.; Wing, R.M.; Zaki, A.I.; Munnecke, D.E.; Sims, J.J. Melleolide, a new antibiotic from Armillaria mellea. Tetrahedron. Lett. 1982, 23, 2515–2518. [Google Scholar] [CrossRef]
- Ishikawa, N.K.; Fukushi, Y.; Yamaji, K.; Tahara, S.; Takahashi, K. Antimicrobial cuparene-type sesquiterpenes, enokipodins c and d, from a mycelial culture of Flammulina velutipes. J. Nat. Prod. 2001, 64, 932–934. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.T.; Winkler, A.L.; Schwan, W.R.; Volk, T.J.; Rott, M.A.; Monte, A. Antibacterial Compounds from Mushrooms I: A Lanostane-type triterpene and prenylphenol derivatives from Jahnoporus hirtus and Albatrellus flettii and their activities against Bacillus cereus and Enterococcus faecalis. Planta. Med. 2010, 76, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Mygind, P.H.; Fischer, R.L.; Schnorr, K.M.; Hansen, M.T.; Sönksen, C.P.; Ludvigsen, S.; Raventós, D.; Buskov, S.; Christensen, B.; De Maria, L. Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus. Nature 2005, 437, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Brinch, K.S.; Sandberg, A.; Baudoux, P.; Van Bambeke, F.; Tulkens, P.M.; Frimodt-Møller, N.; Høiby, N.; Kristensen, H.H. Plectasin shows intracellular activity against Staphylococcus aureus in Human THP-1 monocytes and in a mouse peritonitis model. Antimicrob. Agents Chemother. 2009, 53, 4801–4808. [Google Scholar] [CrossRef] [PubMed]
- Raap, J.; Erkelens, K.; Ogrel, A.; Skladnev, D.A.; Brückner, H. Fungal biosynthesis of non-ribosomal peptide antibiotics and α, α-dialkylated amino acid constituents. J. Pept. Sci. 2005, 11, 331–338. [Google Scholar] [CrossRef]
- Rossi, C.; Eid, M.; Rippa, S.; Castano, S.; Desbat, B.; Chopineau, J.; Béven, L. Exploring the membrane mechanism of the bioactive peptaibol ampullosporin a using lipid monolayers and supported biomimetic membranes. J. Biophys. 2010, 2010, 179641. [Google Scholar]
- Lee, S.J.; Yeo, W.H.; Yun, B.S.; Yoo, I.D. Isolation and sequence analysis of new peptaibol, boletusin, from Boletus spp. J. Pept. Sci. 1999, 5, 374–378. [Google Scholar] [CrossRef]
- Isaac, C.E.; Jones, A.; Pickard, M.A. Production of cyclosporins by Tylypocladium niveum strains. Antimicrob. Agents Chemother. 1990, 34, 121–127. [Google Scholar] [CrossRef]
- Jonsson, M. The Toxicity of Fusarium Mycotoxins Enniatin and Moniliformin; University of Helsinki: Helsinki, Finland, 2017; ISBN 978-952-225-164-0. [Google Scholar]
- McMorris, T.C.; Turner, W.B. Fungal metabolites. Mycologia 1972, 64, 464. [Google Scholar] [CrossRef]
- Ngai, P.H.K.; Ng, T.B. A ribonuclease with antimicrobial, antimitogenic and antiproliferative activities from the edible mushroom Pleurotus sajor-caju. Peptides 2004, 25, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.; De Oliveira, A.; Batista, R. Occurrence, biological activities and synthesis of kaurane diterpenes and their glycosides. Molecules 2007, 12, 455–483. [Google Scholar] [CrossRef] [PubMed]
- Aqueveque, P.; Becerra, J.; Palfner, G.; Silva, M.; Alarcon, J.; Anke, T.; Sterner, O. Antimicrobial activity of metabolites from mycelial cultures of Chilean Basidiomycetes. J. Chil. Chem. Soc. 2006, 51, 1057–1060. [Google Scholar] [CrossRef]
- Sandargo, B.; Thongbai, B.; Praditya, D.; Steinmann, E.; Stadler, M.; Surup, F. Antiviral 4-hydroxypleurogrisein and antimicrobial pleurotin derivatives from cultures of the nematophagous Basidiomycete Hohenbuehelia grisea. Molecules 2018, 23, 2697. [Google Scholar] [CrossRef] [PubMed]
- Bender, S.; Dumitrache-Anghel, C.N.; Backhaus, J.; Christie, G.; Cross, R.F.; Lonergan, G.T.; Baker, W.L. A case for caution in assessing the antibiotic activity of extracts of culinary-medicinal shiitake mushroom [Lentinus edodes (Berk.) Singer] (Agaricomycetideae). Int. J. Med. Mushrooms. 2003, 5, 6. [Google Scholar] [CrossRef]
- Palmieri, F.; Estoppey, A.; House, G.L.; Lohberger, A.; Bindschedler, S.; Chain, P.S.G.; Junier, P. Oxalic Acid, a Molecule at the Crossroads of Bacterial-Fungal Interactions. Adv. Appl. Microbiol. 2019, 106, 49–77. [Google Scholar] [PubMed]
- Quang, D.N.; Bach, D.D.; Hashimoto, T.; Asakawa, Y. Chemical constituents of the vietnamese inedible mushroom Xylaria intracolorata. Nat. Prod. Res. 2006, 20, 317–321. [Google Scholar] [CrossRef]
- Beattie, K.D.; Rouf, R.; Gander, L.; May, T.W.; Ratkowsky, D.; Donner, C.D.; Gill, M.; Grice, I.D.; Tiralongo, E. Antibacterial metabolites from australian macrofungi from the genus Cortinarius. Phytochemistry 2010, 71, 948–955. [Google Scholar] [CrossRef]
- Schwan, W.R.; Dunek, C.; Gebhardt, M.; Engelbrecht, K.; Klett, T.; Monte, A.; Toce, J.; Rott, M.; Volk, T.J.; LiPuma, J.J. Screening a mushroom extract library for activity against Acinetobacter baumannii and Burkholderia cepacia and the identification of a compound with anti-burkholderia activity. Ann. Clin. Microbiol. Antimicrob. 2010, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- No, H.K.; Park, N.Y.; Lee, S.H.; Meyers, S.P. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Coma, V.; Deschamps, A.; Martial-Gros, A. Bioactive packaging materials from edible chitosan polymer—Antimicrobial activity assessment on dairy-related contaminants. J. Food Sci. 2003, 68, 2788–2792. [Google Scholar] [CrossRef]
- Kumar, M. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Jeihanipour, A.; Keikhosro, K.; Taherzadeh, M. Antimicrobial Properties of Fungal Chitosan. Res. J. Biol. Sci. 2007, 2, 239–243. [Google Scholar]
- Hosseinnejad, M.; Jafari, S.M. Evaluation of different factors affecting antimicrobial properties of chitosan. Int. J. Biol. Macromol. 2016, 85, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R.M. Ganoderma—A therapeutic fungal biofactory. Phytochemistry 2006, 67, 1985–2001. [Google Scholar] [CrossRef]
- Quereshi, S.; Pandey, A.K.; Sandhu, S.S. Evaluation of antibacterial activity of different Ganoderma lucidum extracts. PJSR 2010, 3, 9–13. [Google Scholar]
- Yoon, S.Y.; Eo, S.K.; Kim, Y.S.; Lee, C.K.; Han, S.S. Antimicrobial activity of Ganoderma lucidum extract alone and in combination with some antibiotics. Arch. Pharm. Res. 1994, 17, 438–442. [Google Scholar] [CrossRef]
- Yamaç, M.; Bilgili, F. Antimicrobial activities of fruit bodies and/or mycelial cultures of some mushroom isolates. Pharm. Biol. 2006, 44, 660–667. [Google Scholar] [CrossRef]
- Barros, L.; Venturini, B.A.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C.F.R. Chemical composition and biological properties of Portuguese wild mushrooms: A comprehensive study. J. Agric. Food Chem. 2008, 56, 3856–3862. [Google Scholar] [CrossRef] [PubMed]
- Turkoglu, A.; Duru, M.E.; Mercan, N.; Kivrak, I.; Gezer, K. Antioxidant and antimicrobial activities of Laetiporus sulphureus (Bull.) Murrill. Food Chem. 2006, 101, 267–273. [Google Scholar] [CrossRef]
- Hirasawa, M.; Shouji, N.; Neta, T.; Fukushima, K.; Takada, K. Three kinds of antibacterial substances from Lentinus edodes (Berk.) Sing. (Shiitake, an Edible Mushroom). Int. J. Antimicrob. Agents 1999, 11, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Signoretto, C.; Burlacchini, G.; Marchi, A.; Grillenzoni, M.; Cavalleri, G.; Ciric, L.; Lingström, P.; Pezzati, E.; Daglia, M.; Zaura, E. Testing a low molecular mass fraction of a mushroom (Lentinus edodes) extract formulated as an oral rinse in a cohort of volunteers. J. Biomed. Biotechnol. 2011, 2011, 857987. [Google Scholar] [CrossRef]
- Barros, L.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C.F.R. Bioactive properties of the medicinal mushroom Leucopaxillus giganteus mycelium obtained in the presence of different nitrogen sources. Food Chem. 2007, 105, 179–186. [Google Scholar] [CrossRef]
- Sheena, N.; Ajith, T.A.; Mathew, A.; Janardhanan, K.K. Antibacterial activity of three macrofungi, Ganoderma lucidum, Navesporus floccosa and Phellinus rimosus occurring in South India. Pharm. Biol. 2003, 41, 564–567. [Google Scholar] [CrossRef]
- Kalyoncu, F.; Oskay, M.; Sağlam, H.; Erdoğan, T.F.; Tamer, A.U. Antimicrobial and antioxidant activities of mycelia of 10 wild mushroom species. J. Med. Food 2010, 13, 415–419. [Google Scholar] [CrossRef]
- Klaus, A.; Kozarski, M.; Vunduk, J.; Todorovic, N.; Jakovljevic, D.; Zizak, Z.; Pavlovic, V.; Levic, S.; Niksic, M.; Van Griensven, L.J.L.D. Biological potential of extracts of the wild edible basidiomycete mushroom Grifola frondosa. Food Res. Int. 2015, 67, 272–283. [Google Scholar] [CrossRef]
- Rosa, L.H.; Gomes Machado, K.M.; Jacob, C.; Capelari, M.; Rosa, C.A.; Zani, C.L. Screening of Brazilian Basidiomycetes for antimicrobial activity. Mem. Inst. Oswaldo. Cruz. 2003, 98, 967–974. [Google Scholar] [CrossRef]
- Tambekar, D.H.; Sonar, T.P.; Khodke, M.V.; Khante, B.S. The novel antibacterials from two edible mushrooms: Agaricus bisporus and Pleurotus sajor -caju. Int. J. Pharmacol. 2006, 2, 584–587. [Google Scholar]
- Ozen, T.; Darcan, C.; Aktop, O.; Turkekul, I. Screening of antioxidant, antimicrobial activities and chemical contents of edible mushrooms wildly grown in the Black Sea region of Turkey. Comb. Chem. High Throughput. Screen. 2011, 14, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Dulger, B.; Ergul, C.C.; Gucin, F. Antimicrobial activity of the macrofungus Lepista nuda. Fitoterapia 2002, 73, 695–697. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.; Ferreira, I.F.R.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M. A review on antimicrobial activity of mushroom (Basidiomycetes) extracts and isolated compounds. Planta. Med. 2012, 78, 1707–1718. [Google Scholar] [CrossRef] [PubMed]
- Petrović, J.; Stojković, D.; Reis, F.S.; Barros, L.; Glamočlija, J.; Ćirić, A.; Ferreira, I.C.F.R.; Soković, M. Study on chemical, bioactive and food preserving properties of Laetiporus sulphureus (Bull.: Fr.) Murr. Food Funct. 2014, 5, 1441–1451. [Google Scholar] [CrossRef] [PubMed]
- Hleba, L. Antimicrobial activity of crude methanolic extracts from Ganoderma lucidum and Trametes versicolor. Anim. Sci. Biotechnol. 2014, 47, 89–93. [Google Scholar]
- Kontoyiannis, D.P.; Lewis, R.E. Antifungal drug resistance of pathogenic fungi. Lancet 2002, 359, 1135–1144. [Google Scholar] [CrossRef] [PubMed]
- Kuklev, D.V.; Domb, A.J.; Dembitsky, V.W. Bioactive acetylenic metabolites. Phytomedicine 2013, 20, 1145–1159. [Google Scholar] [CrossRef]
- Develoux, M. Griseofulvin. Ann. Dermatol. Venereol. 2001, 128, 1317–1325. [Google Scholar]
- Muralidhar, R.; Gummadi, S.N.; Dasu, V.V.; Panda, T. Statistical analysis on some critical parameters affecting the formation of protoplasts from the mycelium of Penicillium griseofulvum. Biochem. Eng. J. 2003, 3, 229–235. [Google Scholar] [CrossRef]
- Ngai, P.H.K.; Zhao, Z.; Ng, T.B. Agrocybin, an antifungal peptide from the edible mushroom Agrocybe cylindracea. Peptides 2005, 26, 191–196. [Google Scholar] [CrossRef]
- Wong, J.H.; Ng, T.B.; Wang, H.; Sze, S.C.W.; Zhang, K.Y.; Li, Q.; Lu, X. Cordymin, an antifungal peptide from the medicinal fungus Cordyceps militaris. Phytomedicine 2011, 18, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.Q.; Shao, H.J.; Zhu, H.J.; Liu, J.K. Activity in Vitro and in Vivo against Plant Pathogenic Fungi of Grifolin Isolated from the Basidiomycete Albatrellus Dispansus. Z. Naturforsch. C. J. Biosci. 2005, 60, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.J.; Li, L.L.; Deng, Q.P.; Yu, X.F.; Yang, L.F.; Luo, F.J.; Xiao, L.B.; Chen, X.Y.; Ye, M.; Liu, J.K. Grifolin, a potent antitumour natural product upregulates death-associated protein kinase 1 DAPK1 via P53 in nasopharyngeal carcinoma cells. Eur. J. Cancer 2011, 47, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.; Ferreira, I.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M. A Review on antifungal activity of mushroom (Basidiomycetes) extracts and isolated compounds. Curr. Top Med. Chem. 2013, 13, 2648–2659. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.Q.; Wang, F.; Bian, X.Y.; Liu, J.K. Rufuslactone, a new antifungal sesquiterpene from the fruiting bodies of the Basidiomycete Lactarius rufus. J. Antibiot. 2005, 58, 456–459. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, H.; Ng, T.B. Isolation of trichogin, an antifungal protein from fresh fruiting bodies of the edible mushroom Tricholoma giganteum. Peptides 2005, 26, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Kawai, K.; Yamada, S.; Sawai, Y. Isolation of oospolactone as antifungal principle of Gloeophyllum sepiarium. Agric. Biol. Chem. 2014, 40, 811–812. [Google Scholar]
- Ngai, P.H.K.; Ng, T.B. Lentin, a novel and potent antifungal protein from shitake mushroom with inhibitory effects on activity of human immunodeficiency virus-1 reverse transcriptase and proliferation of leukemia cells. Life Sci. 2003, 73, 3363–3374. [Google Scholar] [CrossRef]
- Gilardoni, G.; Clericuzio, M.; Tosi, S.; Zanoni, G.; Vidari, G. Antifungal acylcyclopentenediones from fruiting bodies of Hygrophorus chrysodon. J. Nat. Prod. 2007, 70, 137–139. [Google Scholar] [CrossRef]
- Smania, E.F.; Delle Monache, F.; Smania, A.; Yunes, R.A.; Cuneo, R.S. Antifungal activity of sterols and triterpenes isolated from Ganoderma annulare. Fitoterapia 2003, 74, 375–377. [Google Scholar] [CrossRef]
- Wang, H.; Ng, T.B. Eryngin, a novel antifungal peptide from fruiting bodies of the edible mushroom Pleurotus eryngii. Peptides 2004, 25, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; Kim, S.; Oh, Y.L.; Kong, W.S.; Park, H.; Cho, H.; Jang, K.Y.; Kim, J.G.; Choi, I.G. Genomic discovery of the hypsin gene and biosynthetic pathways for terpenoids in Hypsizygus marmoreus. BMC Genom. 2018, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Steglich, W.; Schramm, G. The strobilurins—New antifungal antibiotics from the Basidiomycete Strobilurus tenacellus. J. Antibiot. 1977, 30, 806–810. [Google Scholar]
- Galappaththi, M.C.A.; Patabendige, N.M.; Premarathne, B.M.; Hapuarachchi, K.K.; Tibpromma, S.; Dai, D.Q.; Suwannarach, N.; Rapior, S.; Karunarathna, S.C. A Review of Ganoderma triterpenoids and their bioactivities. Biomolecules 2022, 13, 24. [Google Scholar] [CrossRef]
- Sultan, S. The antifungal activity of aqueous and alcoholic extract of mushroom (Agaricus Bisporus) against Aspergillus flavus. In Proceedings of the 1st International Multi-Disciplinary Conference Theme: Sustainable Development and Smart Planning, IMDC-SDSP, Online, 28–30 June 2020. [Google Scholar]
- Sangdee, A.; Sangdee, K.; Buranrat, B.; Thammawat, S. Effects of mycelial extract and crude protein of the medicinal mushroom, Ophiocordyceps sobolifera, on the pathogenic fungus, Candida albicans. Trop. Pharm. Res. 2018, 17, 2449–2454. [Google Scholar] [CrossRef]
- Öztürk, M.; Duru, M.E.; Kivrak, Ş.; Mercan-Doĝan, N.; Türkoglu, A.; Özler, M.A. In vitro antioxidant, anticholinesterase and antimicrobial activity studies on three Agaricus species with fatty acid compositions and iron contents: A comparative study on the three most edible mushrooms. Food Chem.Toxicol. 2011, 49, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Pârvu, M.; Andrei, A.Ş.; Roşca-Casian, O. Antifungal activity of Laetiporus sulphureus mushroom extract. Contrib. Bot. 2010, 45, 2015. [Google Scholar]
- Sułkowska-Ziaja, K.; Muszyńska, B.; Gawalska, A.; Sałaciak, K. Laetiporus sulphureus—Chemical composition and medicinal value. Acta. Sci. Pol. Hortorum. Cultus. 2018, 17, 87–96. [Google Scholar] [CrossRef]
- Barros, L.; Baptista, P.; Estevinho, L.; Ferreira, I. Effect of fruiting body maturity stage on chemical composition and antimicrobial activity of Lactarius sp. mushrooms. J. Agric. Food Chem. 2007, 55, 8766–8771. [Google Scholar] [CrossRef]
- Linnakoski, R.; Reshamwala, D.; Veteli, P.; Cortina-Escribano, M.; Vanhanen, H.; Marjomäki, V. Antiviral agents from fungi: Diversity, mechanisms and potential applications. Front. Microbiol. 2018, 9, 2325. [Google Scholar] [CrossRef]
- Mothana, R.A.A.; Awadh Ali, N.A.; Jansen, R.; Wegner, U.; Mentel, R.; Lindequist, U. Antiviral lanostanoid triterpenes from the fungus Ganoderma pfeifferi. Fitoterapia 2003, 74, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.A.A.; Mothana, R.A.A.; Lesnau, A.; Pilgrim, H.; Lindequist, U. Antiviral activity of Inonotus hispidus. Fitoterapia 2003, 74, 483–485. [Google Scholar]
- Tochikura, T.S.; Nakashima, H.; Hirose, K.; Yamamoto, N. A biological response modifier, PSK, inhibits human immunodeficiency virus infection in vitro. Biochem. Biophys. Res. Commun. 1987, 148, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, F.T.; Camelini, C.M.; Mascarello, A.; Rossi, M.J.; Nunes, R.J.; Barardi, C.R. Antiherpetic activity of a sulfated polysaccharide from Agaricus brasiliensis mycelia. Antiviral. Res. 2011, 92, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; He, F.; Piraino, F.F.; Xiang, H.; Chen, J.; Wang, Y. Antiviral activity of a cloned peptide rc28 isolated from the higher basidiomycetes mushroom Rozites caperata in a mouse model of HSV-1 keratitis. Int. J. Med. Mushrooms 2015, 17, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Vlasenko, V.A.; Ilyicheva, T.N.; Teplyakova, T.V.; Svyatchenko, S.V.; Asbaganov, S.V.; Zmitrovich, I.V. Antiviral activity of total polysaccharide fraction of water and ethanol extracts of Pleurotus pulmonarius against the influenza A virus. Curr. Res. Environ. Appl. Mycol. 2020, 10, 224–235. [Google Scholar] [CrossRef]
- Ohta, Y.; Lee, J.B.; Hayashi, K.; Fujita, A.; Dong, K.P.; Hayashi, T. In vivo anti-influenza virus activity of an immunomodulatory acidic polysaccharide isolated from Cordyceps militaris grown on germinated soybeans. J. Agric. Food Chem. 2007, 55, 10194–10199. [Google Scholar] [CrossRef]
- Mlinaric, A.; Kac, J.; Pohleven, F. Screening of selected wood-damaging fungi for the HIV-1 reverse transcriptase inhibitors. Acta. Pharm. 2005, 55, 69–79. [Google Scholar]
- Faccin, L.C.; Benati, F.; Rincão, V.P.; Mantovani, M.S.; Soares, S.A.; Gonzaga, M.L. Antiviral activity of aqueous and ethanol extracts and of an isolated polysaccharide from Agaricus brasiliensis against poliovirus type 1. Lett. Appl. Microbiol. 2007, 45, 24–28. [Google Scholar] [CrossRef]
- Lee, S.M.; Kim, S.M.; Lee, Y.H.; Kim, W.J.; Park, J.K.; Park, Y.I. Macromolecules isolated from Phellinus pini fruiting body: Chemical characterization and antiviral activity. Macromol. Res. 2010, 18, 602–609. [Google Scholar] [CrossRef]
- Hwang, B.S.; Lee, I.K.; Choi, H.J.; Yun, B.S. Anti-influenza activities of polyphenols from the medicinal mushroom Phellinus baumii. Bioorg. Med. Chem. Lett. 2015, 25, 3256–3260. [Google Scholar] [CrossRef]
- Seo, D.J.; Choi, C. Antiviral bioactive compounds of mushrooms and their antiviral mechanisms: A review. Viruses 2021, 23, 350. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, X.; Hao, C.; Zeng, P.; Zhang, M.; Liu, Y. Grifola frondosa polysaccharide: A review of antitumor and other biological activity studies in China. Discov. Med. 2018, 25, 159–176. [Google Scholar]
- Gu, C.Q.; Li, J.; Chao, F.H. Inhibition of hepatitis B virus by D-fraction from Grifola frondosa: Synergistic effect of combination with interferon-alpha in HepG2 2.2.15. Antiviral. Res. 2006, 72, 162–165. [Google Scholar] [CrossRef]
- Ren, G.; Xu, L.; Lu, T.; Yin, J. Structural characterization and antiviral activity of lentinan from Lentinus edodes mycelia against infectious hematopoietic necrosis virus. Int. J. Biol. Macromol. 2018, 115, 1202–1210. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, F.; Anderson, D.; Najafzadeh, M. The antiviral, anti-inflammatory effects of natural medicinal herbs and mushrooms and SARS-CoV-2 infection. Nutrients 2020, 12, 2573. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Chen, H.; Dong, P.; Lu, X. Anti-inflammatory and anticancer activities of extracts and compounds from the mushroom Inonotus obliquus. Food Chem. 2013, 139, 503–508. [Google Scholar] [CrossRef]
- Shibnev, V.A.; Mishin, D.V.; Garaev, T.M.; Finogenova, N.P.; Botikov, A.G.; Deryabin, P.G. Antiviral activity of inonotus obliquus fungus extract towards infection caused by hepatitis C virus in cell cultures. Bull. Exp. Biol. Med. 2011, 151, 612–614. [Google Scholar] [CrossRef]
- Ershova, E.I.; Tikhonova, O.V.; Lur’e, L.M.; Efremenkova, O.V.; Kamzolkina, O.V.; Dudnik, I.V. Antimicrobial activity of Laetiporus sulphureus strains grown in submerged culture. Antibiot. Khimioter. 2003, 48, 18–22. [Google Scholar]
- Yang, S.S.; Cheng, M.J.; Chan, H.Y.; Hsieh, S.Y.; Wu, H.C.; Yuan, G.F. New secondary metabolites from an endophytic fungus in Porodaedalea pini. Rec. Nat. Prod. 2017, 11, 251–257. [Google Scholar]
- Taofiq, O.; González-Paramás, A.M.; Martins, A.; Barreiro, M.F.; Ferreira, I.C.F.R. Mushrooms extracts and compounds in cosmetics, cosmeceuticals and nutricosmetics—A review. Ind. Crops. Prod. 2016, 90, 38–48. [Google Scholar] [CrossRef]
- Taofiq, O.; Heleno, S.A.; Calhelha, R.C.; Alves, M.J.; Barros, L.; Barreiro, M.F.; González-Paramás, A.M.; Ferreira, I.C.F.R. Development of mushroom-based cosmeceutical formulations with anti-inflammatory, anti-tyrosinase, antioxidant, and antibacterial properties. Molecules 2016, 21, 1372. [Google Scholar] [CrossRef] [PubMed]
- Eiamthaworn, K.; Kaewkod, T.; Bovonsombut, S.; Tragoolpua, Y. Efficacy of Cordyceps militaris extracts against some skin pathogenic bacteria and antioxidant activity. J. Fungi. 2022, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Joshi, S.R. Industrial Perspectives of Fungi. In Industrial Microbiology and Biotechnology; Verma, P., Ed.; Springer: Singapore, 2022. [Google Scholar]
- Shankar, A.; Sharma, K.K. Fungal secondary metabolites in food and pharmaceuticals in the era of multi-omics. Appl. Microbiol. Biotechnol. 2022, 106, 3465–3488. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Avire, N.J.; Whiley, H.; Ross, K.A. Review of Streptococcus pyogenes: Public health risk factors, prevention and control. Pathogens 2021, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Platsidaki, E.; Dessinioti, C. Recent advances in understanding Propionibacterium acnes (Cutibacterium acnes) in acne. F1000Research 2018, 7, 1953. [Google Scholar] [CrossRef]
- Wang, R.; Huang, C.; Zhang, Y.; Li, R. Invasive dermatophyte infection: A systematic review. Mycoses 2021, 64, 340–348. [Google Scholar] [CrossRef]
- Ifuku, S. Chitin and Chitosan Nanofibers: Preparation and Chemical Modifications. Molecules 2014, 19, 18367–18380. [Google Scholar] [CrossRef]
- Park, B.K.; Kim, M.M. Applications of chitin and its derivatives in biological medicine. Int. J. Mol. Sci. 2010, 11, 5152–5164. [Google Scholar] [CrossRef]
- Vasylchenko, O.A.; Abramova, M.V. Comparative analysis of sources for chitosan obtaining. Problems Environ. Biotechnol. 2015, 1, 1–20. [Google Scholar] [CrossRef]
- Mattila, P.; Könkö, K.; Eurola, M.; Pihlava, J.M.; Astola, J.; Vahteristo, L. Contents of vitamins, mineral elements, and some phenolic compounds in cultivated mushrooms. J. Agric. Food Chem. 2001, 49, 2343–2348. [Google Scholar] [CrossRef] [PubMed]
- Fiume, M.M.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C. Safety assessment of tocopherols and tocotrienols as used in cosmetics. Int. J. Toxicol. 2018, 37, 61S–94S. [Google Scholar] [CrossRef]
- Sharif, S.; Rashid, S.; Atta, A.; Irshad, A.; Riaz, M.; Shahid, M. Phenolics, tocopherols and fatty acid profiling of wild and commercial mushrooms from Pakistan. J. Biol. Regul. Homeost. Agents. 2018, 32, 863–867. [Google Scholar] [PubMed]
- Chang, T.S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.M.; Kwak, S.Y.; Seo, H.S.; Seo, J.H.; Kim, B.G.; Lee, Y.S. Kojic acid-amino acid conjugates as tyrosinase inhibitors. Bioorg. Med. Chem. Lett. 2009, 19, 5586–5589. [Google Scholar] [CrossRef]
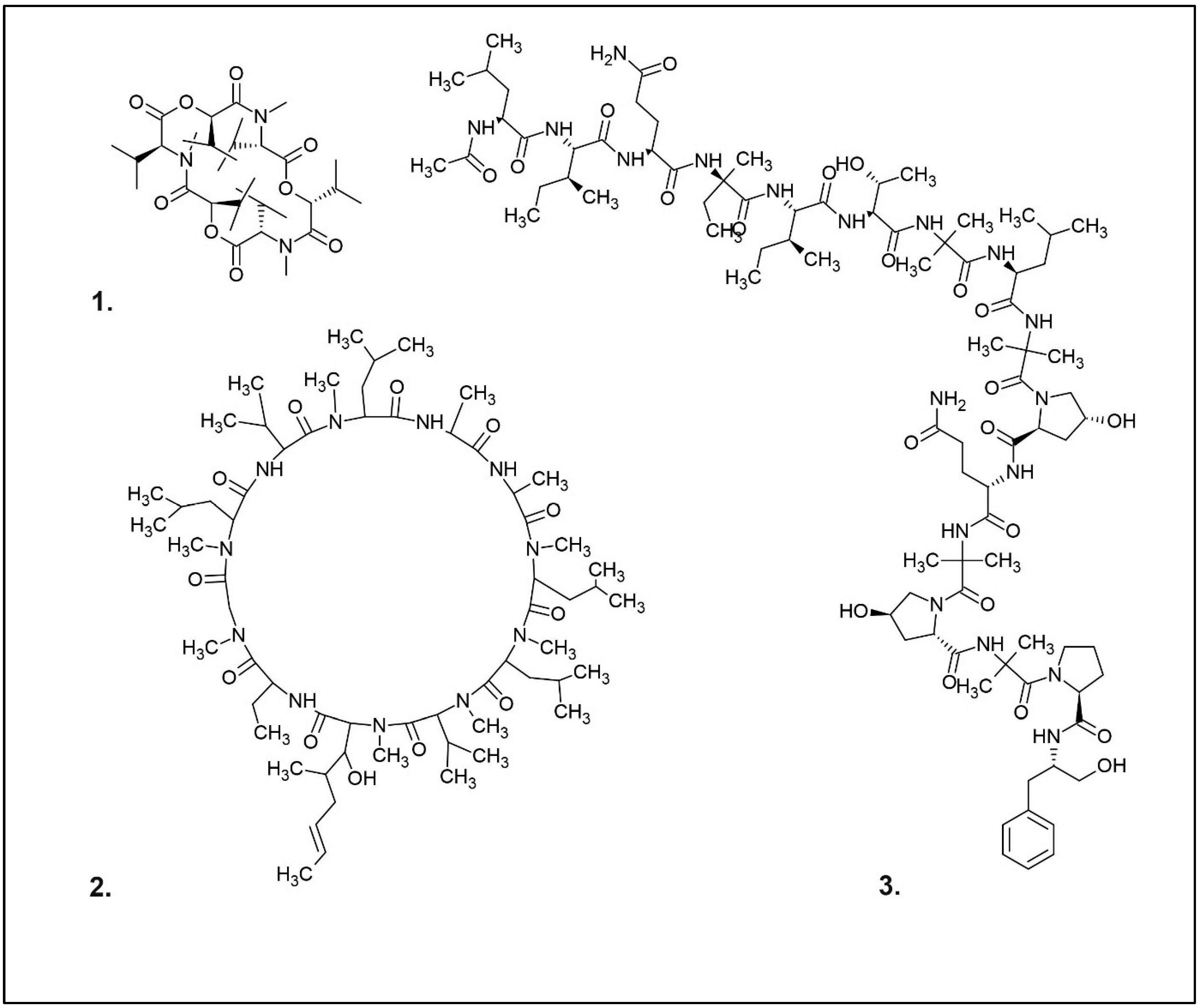
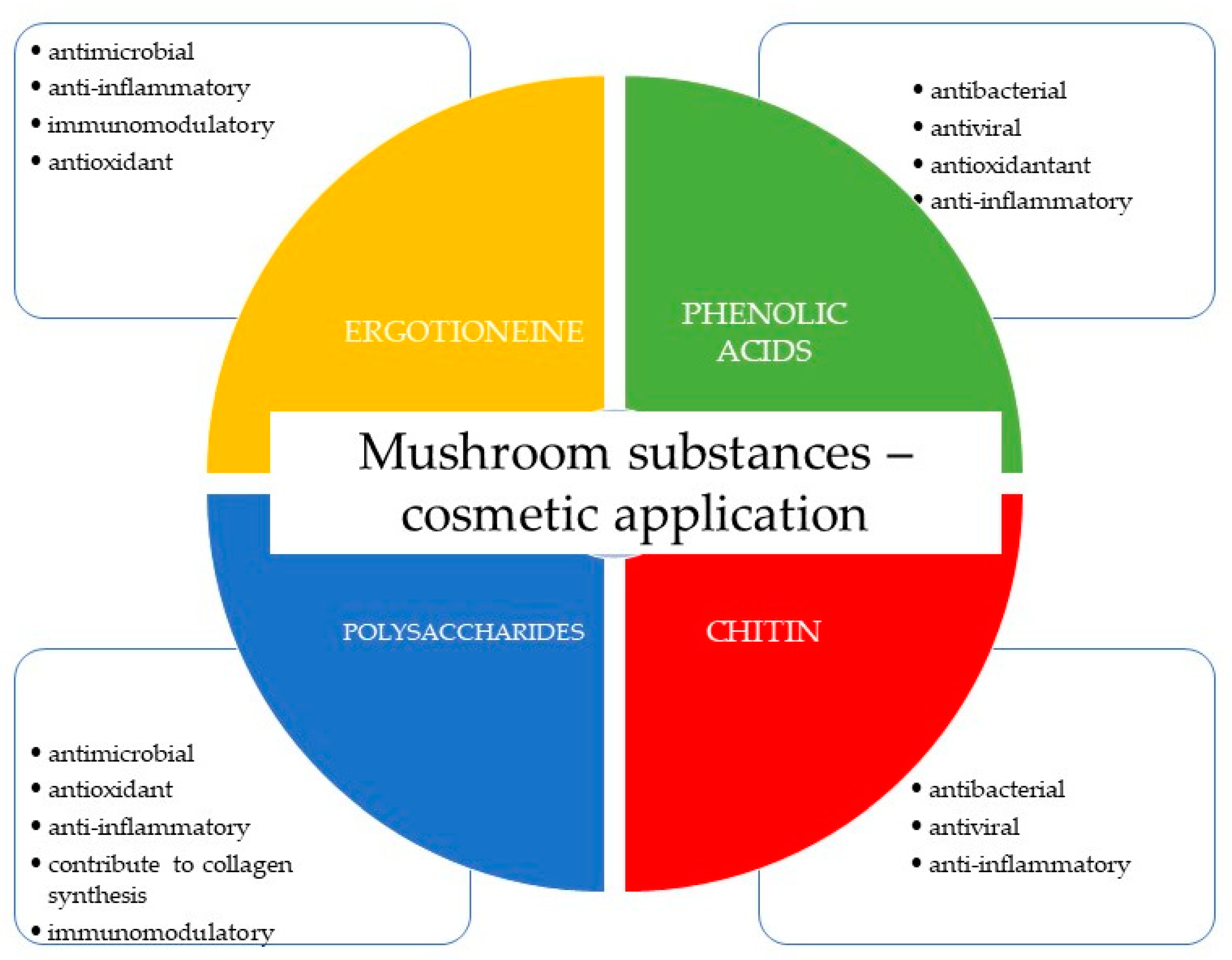
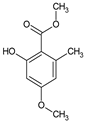
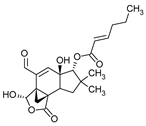
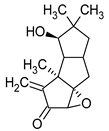
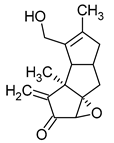
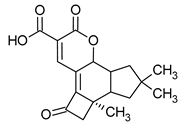
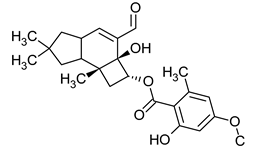
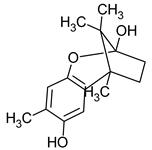
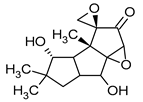
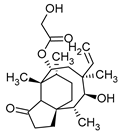
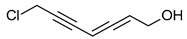
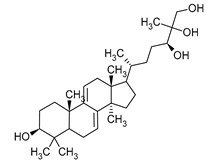
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).