
HIV-1 Integrase Inhibition Activity by Spiroketals Derived from Plagius flosculosus, an Endemic Plant of Sardinia (Italy) and Corsica (France)

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
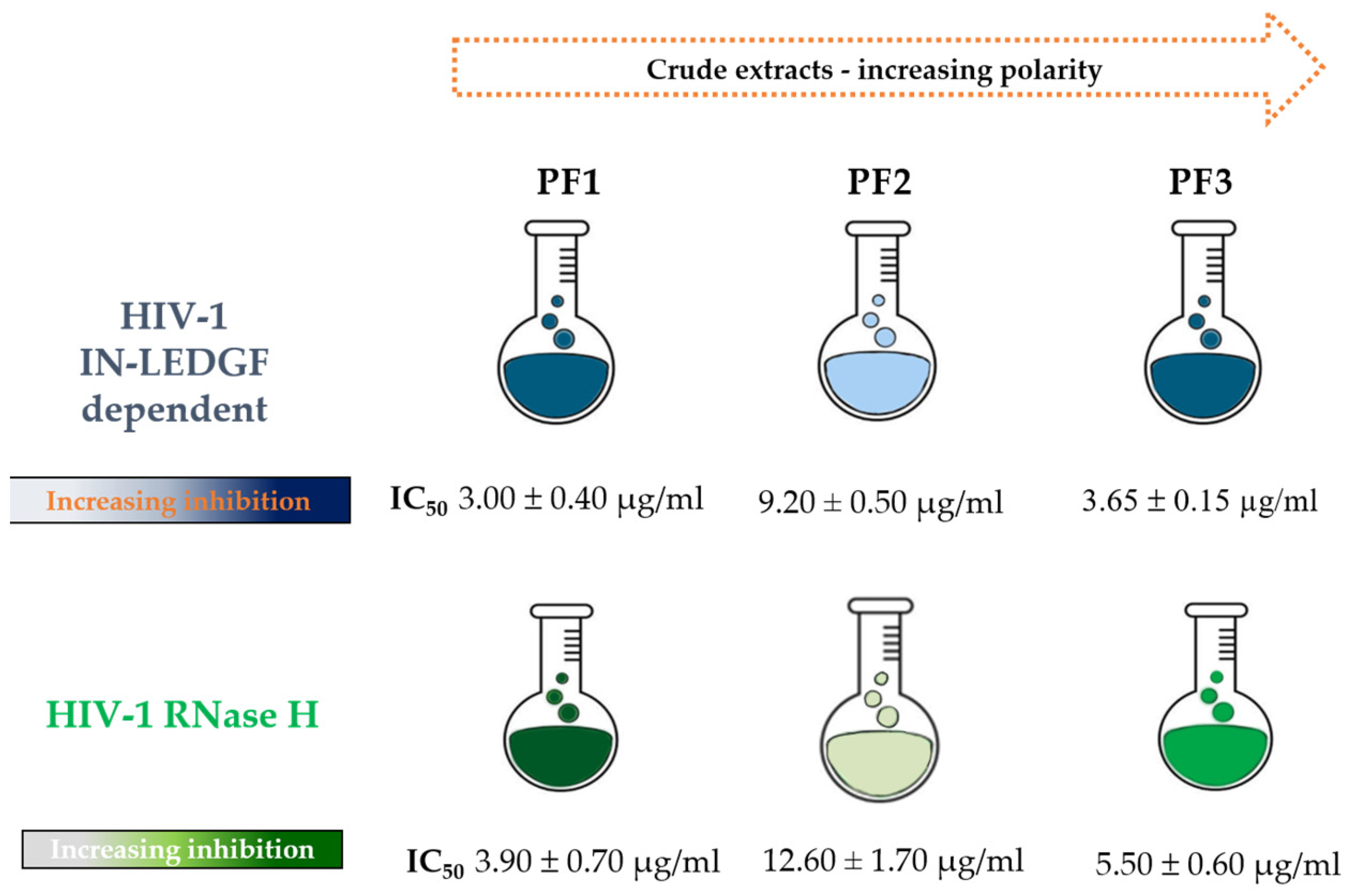
2.1. Effect of P. flosculosus Extracts on HIV-1 IN Activity in the Presence of LEDGF/p75 Cellular Cofactor
2.2. Effect of P. flosculosus Extracts on HIV-1 RT-Associated RNase H Activity
2.3. Chemical Characterization of Bioactive Crude Extract
2.4. Effects of Pure Compounds on HIV-1 IN Activity in the Presence of LEDGF/p75 Cellular Cofactor and on HIV-1 RNase H Activity
2.5. Pure Compounds’ Spectra in the Presence of Magnesium
2.6. Evaluation of Inhibition of Pure Compounds against HIV-1 IN Activities
2.7. Inhibition of HIV-1 Replication in a Cell-Based Assay by SPK3
3. Materials and Methods
3.1. Plant Material and Sample Preparation
3.2. Preparation of Crude Extracts for Bioassay
3.3. Bioguided-Fractionation of the Active Extract and Compounds Purification
3.3.1. General Chromatographic Procedures
3.3.2. NMR Experiments
3.3.3. Compound Purification
3.4. Anti-HIV Biochemical Assays
3.4.1. Recombinant Proteins Preparation
3.4.2. HTRF LEDGF-Dependent and -Independent Assays
3.4.3. IN–LEDGF Binding Assay
3.4.4. IN–IN Binding Assay
3.4.5. RT-Associated RNase H Assay
3.4.6. Compound Spectra in the Presence of MgCl2
3.5. Cytotoxicity Assay
3.6. Determination of Anti-HIV-1 Activity in TZM-bl Cell Line
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fact Sheet—Latest Global and Regional Statistics on the Status of the AIDS Epidemic. UNAIDS. Available online: https://www.unaids.org/en/resources/documents/2022/UNAIDS_FactSheet (accessed on 3 April 2023).
- Hu, W.; Hughes, S.H. HIV-1 Reverse Transcription. Cold Spring Harb. Perspect. Med. 2012, 2, a006882. [Google Scholar] [CrossRef]
- Yang, W.E.I.; Steitz, T.A. Recombining the Structures of HIV Integrase, RuvC and RNase H The Recently Reported Crystal Structures of Two Recombination Enzymes, Endonuclease, Are Surprisingly Similar to That of Ribonuclease H Suggesting the Possibility That They Have a Common Enz. Structure 1995, 3, 131–134. [Google Scholar] [CrossRef]
- Delelis, O.; Carayon, K.; Saïb, A.; Deprez, E.; Mouscadet, J.F. Integrase and Integration: Biochemical Activities of HIV-1 Integrase. Retrovirology 2008, 5, 114. [Google Scholar] [CrossRef] [PubMed]
- Engelman, A.; Kessl, J.J.; Kvaratskhelia, M. Allosteric Inhibition of HIV-1 Integrase Activity. Curr. Opin. Chem. Biol. 2013, 17, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Rozman, M.; Zidovec-Lepej, S.; Jambrosic, K.; Babić, M.; Drmić Hofman, I. Role of TLRs in HIV-1 Infection and Potential of TLR Agonists in HIV-1 Vaccine Development and Treatment Strategies. Pathogens 2023, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Finzi, D.; Blankson, J.; Siliciano, J.D.; Margolick, J.B.; Chadwick, K.; Pierson, T.; Smith, K.; Lisziewicz, J.; Lori, F.; Flexner, C.; et al. Latent Infection of CD4+ T Cells Provides a Mechanism for Lifelong Persistence of HIV-1, Even in Patients on Effective Combination Therapy. Nat. Med. 1999, 5, 512–517. [Google Scholar] [CrossRef]
- Ndashimye, E.; Reyes, P.S.; Arts, E.J. New Antiretroviral Inhibitors and HIV-1 Drug Resistance: More Focus on 90% HIV-1 Isolates? FEMS Microbiol. Rev. 2023, 47, fuac040. [Google Scholar] [CrossRef]
- Esposito, F.; Ambrosio, F.A.; Maleddu, R.; Costa, G.; Rocca, R.; Maccioni, E.; Catalano, R.; Romeo, I.; Eleftheriou, P.; Karia, D.C.; et al. Chromenone Derivatives as a Versatile Scaffold with Dual Mode of Inhibition of HIV-1 Reverse Transcriptase-Associated Ribonuclease H Function and Integrase Activity. Eur. J. Med. Chem. 2019, 182, 111617. [Google Scholar] [CrossRef]
- Mahboubi-Rabbani, M.; Abbasi, M.; Hajimahdi, Z.; Zarghi, A. HIV-1 Reverse Transcriptase/Integrase Dual Inhibitors: A Review of Recent Advances and Structure-Activity Relationship Studies. Iran. J. Pharm. Res. 2021, 20, 333–369. [Google Scholar] [CrossRef]
- Métifiot, M.; Leon, O.; Tarrago-Litvak, L.; Litvak, S.; Andréola, M.L. Targeting HIV-1 Integrase with Aptamers Selected against the Purified RNase H Domain of HIV-1 RT. Biochimie 2005, 87, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Kessl, J.J.; Jena, N.; Koh, Y.; Taskent-Sezgin, H.; Slaughter, A.; Feng, L.; De Silva, S.; Wu, L.; Le Grice, S.F.J.; Engelman, A.; et al. Multimode, Cooperative Mechanism of Action of Allosteric HIV-1 Integrase Inhibitors. J. Biol. Chem. 2012, 287, 16801–16811. [Google Scholar] [CrossRef]
- Le Rouzic, E.; Bonnard, D.; Chasset, S.; Bruneau, J.M.; Chevreuil, F.; Le Strat, F.; Nguyen, J.; Beauvoir, R.; Amadori, C.; Brias, J.; et al. Dual Inhibition of HIV-1 Replication by Integrase-LEDGF Allosteric Inhibitors Is Predominant at the Post-Integration Stage. Retrovirology 2013, 10, 144. [Google Scholar] [CrossRef]
- Dinh, L.P.; Sun, J.; Glenn, C.D.; Patel, K.; Pigza, J.A.; Donahue, M.G.; Yet, L.; Kessl, J.J. Multi-Substituted Quinolines as HIV-1 Integrase Allosteric Inhibitors. Viruses 2022, 14, 1466. [Google Scholar] [CrossRef] [PubMed]
- Christ, F.; Voet, A.; Marchand, A.; Nicolet, S.; Desimmie, B.A.; Marchand, D.; Bardiot, D.; Van Der Veken, N.J.; Van Remoortel, B.; Strelkov, S.V.; et al. Rational Design of Small-Molecule Inhibitors of the LEDGF/P75-Integrase Interaction and HIV Replication. Nat. Chem. Biol. 2010, 6, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Slaughter, A.; Jena, N.; Feng, L.; Kessl, J.J.; Fadel, H.J.; Malani, N.; Male, F.; Wu, L.; Poeschla, E.; et al. A New Class of Multimerization Selective Inhibitors of HIV-1 Integrase. PLoS Pathog. 2014, 10, e1004171. [Google Scholar] [CrossRef]
- Ohata, Y.; Tomonaga, M.; Watanabe, Y.; Tomura, K.; Kimura, K.; Akaki, T.; Adachi, K.; Kodama, E.N.; Matsuzaki, Y.; Hayashi, H. Antiviral Activity and Resistance Profile of the Novel HIV-1 Non-Catalytic Site Integrase Inhibitor JTP-0157602. J. Virol. 2022, 96, e01843-21. [Google Scholar] [CrossRef] [PubMed]
- Parcella, K.; Wang, T.; Eastman, K.; Zhang, Z.; Yin, Z.; Patel, M.; Tu, Y.; Zheng, B.Z.; Walker, M.A.; Saulnier, M.G.; et al. Discovery and Preclinical Profiling of GSK3839919, a Potent HIV-1 Allosteric Integrase Inhibitor. ACS Med. Chem. Lett. 2022, 13, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Engelman, A.N.; Kvaratskhelia, M. Multimodal Functionalities of HIV-1 Integrase. Viruses 2022, 14, 926. [Google Scholar] [CrossRef] [PubMed]
- Cos, P.; Maes, L.; Vlietinck, A.; Pieters, L. Plant-Derived Leading Compounds for Chemotherapy of Human Immunodeficiency Virus (HIV) Infection—An Update (1998–2007). Planta Med. 2008, 74, 1323–1337. [Google Scholar] [CrossRef]
- Sanna, C.; Maxia, A.; Fenu, G.; Loi, M.C. So Uncommon and so Singular, but Underexplored: An Updated Overview on Ethnobotanical Uses, Biological Properties and Phytoconstituents of Sardinian Endemic Plants. Plants 2020, 9, 958. [Google Scholar] [CrossRef]
- Guzzo, F.; Russo, R.; Sanna, C.; Celaj, O.; Caredda, A.; Corona, A.; Tramontano, E.; Fiorentino, A.; Esposito, F.; D’abrosca, B. Chemical Characterization and Anti-HIV-1 Activity Assessment of Iridoids and Flavonols from Scrophularia trifoliata. Molecules 2021, 26, 4777. [Google Scholar] [CrossRef] [PubMed]
- Sanna, C.; Scognamiglio, M.; Fiorentino, A.; Corona, A.; Graziani, V.; Caredda, A.; Cortis, P.; Montisci, M.; Ceresola, E.R.; Canducci, F.; et al. Prenylated Phloroglucinols from Hypericum scruglii, an Endemic Species of Sardinia (Italy), as New Dual HIV-1 Inhibitors Effective on HIV-1 Replication. PLoS ONE 2018, 12, e0195168. [Google Scholar] [CrossRef] [PubMed]
- Fois, M.; Farris, E.; Calvia, G.; Campus, G.; Fenu, G.; Porceddu, M.; Bacchetta, G. The Endemic Vascular Flora of Sardinia: A Dynamic Checklist with an Overview of Biogeography and Conservation Status. Plants 2022, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, B.; Piras, A.; Porcedda, S.; Tuveri, E.; Laconi, S.; Deidda, D.; Maxia, A. Chemical and Biological Comparisons on Supercritical Extracts of Tanacetum cinerariifolium (Trevir) Sch. Bip. with Three Related Species of Chrysanthemums of Sardinia (Italy). Nat. Prod. Res. 2009, 23, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Casu, L.; Bonsignore, L.; Pinna, M.; Casu, M.; Floris, C.; Gertsch, J.; Cottiglia, F. Cytotoxic Diacetylenic Spiroketal Enol Ethers from Plagius flosculosus. J. Nat. Prod. 2006, 69, 295–298. [Google Scholar] [CrossRef]
- Xiao, C.Y.; Lan, J.E.; Liu, X.; Sun, Z.L.; Li, X.J.; Yin, Y.H.; Gibbons, S.; Mu, Q. Acetylenic spiroketal enol ethers from Artemisia rupestris and their synergistic antibacterial effects on methicillin-resistant Staphylococcus aureus. Nat. Prod. Res. 2023, 1–5. [Google Scholar] [CrossRef]
- Nakamura, Y.; Ohto, Y.; Murakami, A.; Jiwajinda, S.; Ohigashi, H. Isolation and Identification of Acetylenic Spiroketal Enol Ethers from Artemisia lactiflora as Inhibitors of Superoxide Generation Induced by a Tumor Promoter in Differentiated HL-60 Cells. J. Agric. Food Chem. 1998, 46, 5031–5036. [Google Scholar] [CrossRef]
- Calzado, M.A.; Lüdi, K.S.; Fiebich, B.; Ben-Neriah, Y.; Bacher, S.; Munoz, E.; Ballero, M.; Prosperini, S.; Appendino, G.; Schmitz, M.L. Inhibition of NF-KappaB Activation and Expression of Inflammatory Mediators by Polyacetylene Spiroketals from Plagius flosculosus. Biochim. Biophys. Acta 2005, 1729, 88–93. [Google Scholar] [CrossRef]
- Álvarez, Á.L.; Habtemariam, S.; Abdel Moneim, A.E.; Melón, S.; Dalton, K.P.; Parra, F. A spiroketal-enol ether derivative from Tanacetum vulgare selectively inhibits HSV-1 and HSV-2 glycoprotein accumulation in Vero cells. Antivir. Res. 2015, 119, 8–18. [Google Scholar] [CrossRef]
- Cos, P.; Vlietinck, A.J.; Berghe, D.V.; Maes, L. Anti-Infective Potential of Natural Products: How to Develop a Stronger in Vitro “Proof-of-Concept”. J. Ethnopharmacol. 2006, 106, 290–302. [Google Scholar] [CrossRef]
- Sanna, C.; Rigano, D.; Corona, A.; Piano, D.; Formisano, C.; Farci, D.; Franzini, G.; Ballero, M.; Chianese, G.; Tramontano, E.; et al. Dual HIV-1 Reverse Transcriptase and Integrase Inhibitors from Limonium morisianum Arrigoni, an endemic species of Sardinia (Italy). Nat. Prod. Res. 2019, 33, 1798–1803. [Google Scholar] [CrossRef] [PubMed]
- Hnatyszyn, O.; Broussalis, A.; Herrera, G.; Muschietti, L.; Coussio, J.; Martino, V.; Ferraro, G.; Font, M.; Monge, A.; Martínez-Irujo, J.J.; et al. Argentine Plant Extracts Active against Polymerase and Ribonuclease H Activities of HIV-1 Reverse Transcriptase. Phytother. Res. 1999, 13, 206–209. [Google Scholar] [CrossRef]
- Buono-Core, G.E.; Nunez, M.V.; Lucero, A.; Vargas, M.R.; Jullian, C. Structural elucidation of bioactive principles in floral extracts of German chamomille (Matricaria recutita L.). J. Chil. Chem. Soc. 2011, 56, 549–553. [Google Scholar] [CrossRef]
- Maqua, M.P.; Vines, A.C.G.; Caballero, E.; Grande, M.C.; Medarde, M.; Bellido, I.S. Components from Santolina rosmarinifolia, subspecies rosmarinifolia and canescens. Phytochemistry 1988, 27, 3664–3667. [Google Scholar] [CrossRef]
- Konovalov, D.A. Polyacetylene Compounds of Plants of the Asteraceae Family (Review). Pharm. Chem. J. 2014, 48, 613–631. [Google Scholar] [CrossRef]
- Yin, B.L.; Fan, J.F.; Gao, Y.; Wu, Y.L. Progress in Molecular Diversity of Tonghaosu and Its Analogs. ARKIVOC 2003, ii, 70–83. [Google Scholar] [CrossRef]
- Amoros, M.; Simões, C.M.; Girre, L.; Sauvager, F.; Cormier, M. Synergistic effect of flavones and flavonols against herpes simplex virus type 1 in cell culture. Comparison with the antiviral activity of propolis. J. Nat. Prod. 1992, 55, 1732–1740. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, H.; Lyu, L.; Huang, Z.; Fan, S.; Wu, W.; Li, W. Synergistic effect of natural antifungal agents for postharvest diseases of blackberry fruits. J. Sci. Food Agric. 2019, 9, 3343–3349. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef]
- Corona, A.; Di Leva, F.S.; Thierry, S.; Pescatori, L.; Cuzzucoli Crucitti, G.; Subra, F.; Delelis, O.; Esposito, F.; Rigogliuso, G.; Costi, R.; et al. Identification of highly conserved residues involved in inhibition of HIV-1 RNase H function by Diketo acid derivatives. Antimicrob. Agents Chemother. 2014, 58, 6101–6110. [Google Scholar] [CrossRef]
- Summa, V.; Petrocchi, A.; Bonelli, F.; Crescenzi, B.; Donghi, M.; Ferrara, M.; Fiore, F.; Gardelli, C.; Gonzalez Paz, O.; Hazuda, D.J.; et al. Discovery of raltegravir, a potent, selective orally bioavailable HIV-integrase inhibitor for the treatment of HIV-AIDS infection. J. Med. Chem. 2008, 51, 5843–5855. [Google Scholar] [CrossRef] [PubMed]
- Tramontano, E.; Esposito, F.; Badas, R.; Di Santo, R.; Costi, R.; La Colla, P. 6-[1-(4-Fluorophenyl)Methyl-1H-Pyrrol-2-Yl)]-2,4-Dioxo-5-Hexenoic Acid Ethyl Ester a Novel Diketo Acid Derivative Which Selectively Inhibits the HIV-1 Viral Replication in Cell Culture and the Ribonuclease H Activity in Vitro. Antivir. Res. 2005, 65, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; McKee, C.J.; Kessl, J.J.; Santos, W.L.; Daigle, J.E.; Engelman, A.; Verdine, G.; Kvaratskhelia, M. Subunit-specific protein footprinting reveals significant structural rearrangements and a role for N-terminal Lys-14 of HIV-1 Integrase during viral DNA binding. J. Biol. Chem. 2008, 283, 5632–5641. [Google Scholar] [CrossRef] [PubMed]
- Kessl, J.J.; Li, M.; Ignatov, M.; Shkriabai, N.; Eidahl, J.O.; Feng, L.; Musier-Forsyth, K.; Craigie, R.; Kvaratskhelia, M. FRET analysis reveals distinct conformations of IN tetramers in the presence of viral DNA or LEDGF/p75. Nucleic Acids Res. 2011, 39, 9009–9022. [Google Scholar] [CrossRef]
- Saladini, F.; Giannini, A.; Boccuto, A.; Vicenti, I.; Zazzi, M. Agreement between an In-House Replication Competent and a Reference Replication Defective Recombinant Virus Assay for Measuring Phenottypic Resistance to HIV-1 Protease, Reverse Transcriptase, and Integrase Inhibitors. J. Clin. Lab. Anal. 2018, 32, e22206. [Google Scholar] [CrossRef]
- Guzzo, F.; Buommino, E.; Landrum, L.; Russo, R.; Lembo, F.; Fiorentino, A.; D’Abrosca, B. Phytochemical Investigation of Myrcianthes cisplatensis: Structural Characterization of New p-Coumaroyl Alkylphloroglucinols and Antimicrobial Evaluation against Staphylococcus aureus. Plants 2023, 12, 1046. [Google Scholar] [CrossRef]
- Esposito, F.; Tintori, C.; Martini, R.; Christ, F.; Debyser, Z.; Ferrarese, R.; Cabiddu, G.; Corona, A.; Ceresola, E.R.; Calcaterra, A.; et al. Kuwanon-L as a New Allosteric HIV-1 Integrase Inhibitor: Molecular Modeling and Biological Evaluation. ChemBioChem 2015, 16, 2507–2512. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IN-LEDGF-Dependent 1 IC50 (µM) | RNase H 1 IC50 (µM) |
|---|---|---|
| SPK1 | 1.69 ± 0.38 | 10.02 ± 5.40 |
| SPK2 | 14.70 ± 0.30 | 20.0 ± 6.10 |
| SPK3 | 1.46 ± 0.16 | 3.60 ± 0.60 |
| Raltegravir | 0.05 ± 0.02 | - |
| RDS1759 | - | 0.0067 ± 0.90 |
| Code | IN–LEDGF Binding 1 IC50(µM) | IN–IN Subunit Exchange 2 IC50(µM) | IN Multimerization 3 MI50(µM) | LEDGF-Independent IN Activity 4 IC50(µM) |
|---|---|---|---|---|
| SPK3 | >100 | >100 | 1.00 ± 0.01 | 27.3 ± 2.3 |
| Kuwanon-L | 22.0 ± 0.5 | >100 | 38.0 ± 0.02 | 34.0 ± 0.5 |
| Compound | 1 EC50 (µM) | 2 CC50 (µM) |
|---|---|---|
| SPK3 | NA | 50.1 ± 4.3 |
| Raltegravir | 0.0087 ± 0.059 | >100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanna, C.; D’Abrosca, B.; Fiorentino, A.; Giammarino, F.; Vicenti, I.; Corona, A.; Caredda, A.; Tramontano, E.; Esposito, F. HIV-1 Integrase Inhibition Activity by Spiroketals Derived from Plagius flosculosus, an Endemic Plant of Sardinia (Italy) and Corsica (France). Pharmaceuticals 2023, 16, 1118. https://doi.org/10.3390/ph16081118
Sanna C, D’Abrosca B, Fiorentino A, Giammarino F, Vicenti I, Corona A, Caredda A, Tramontano E, Esposito F. HIV-1 Integrase Inhibition Activity by Spiroketals Derived from Plagius flosculosus, an Endemic Plant of Sardinia (Italy) and Corsica (France). Pharmaceuticals. 2023; 16(8):1118. https://doi.org/10.3390/ph16081118
Chicago/Turabian StyleSanna, Cinzia, Brigida D’Abrosca, Antonio Fiorentino, Federica Giammarino, Ilaria Vicenti, Angela Corona, Alessia Caredda, Enzo Tramontano, and Francesca Esposito. 2023. "HIV-1 Integrase Inhibition Activity by Spiroketals Derived from Plagius flosculosus, an Endemic Plant of Sardinia (Italy) and Corsica (France)" Pharmaceuticals 16, no. 8: 1118. https://doi.org/10.3390/ph16081118
APA StyleSanna, C., D’Abrosca, B., Fiorentino, A., Giammarino, F., Vicenti, I., Corona, A., Caredda, A., Tramontano, E., & Esposito, F. (2023). HIV-1 Integrase Inhibition Activity by Spiroketals Derived from Plagius flosculosus, an Endemic Plant of Sardinia (Italy) and Corsica (France). Pharmaceuticals, 16(8), 1118. https://doi.org/10.3390/ph16081118