
Angelica keiskei Impacts the Lifespan and Healthspan of Drosophila melanogaster in a Sex and Strain-Dependent Manner



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
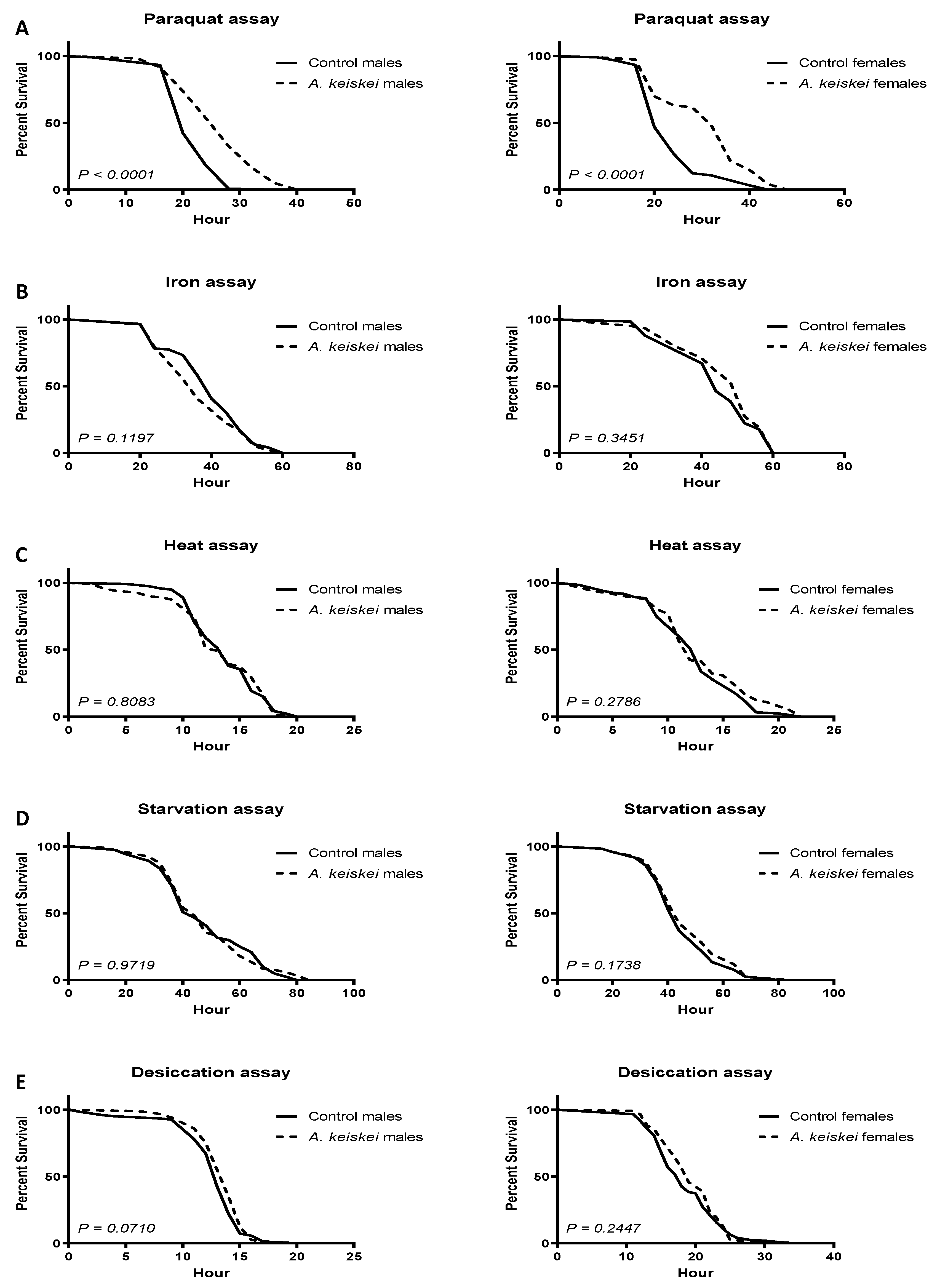
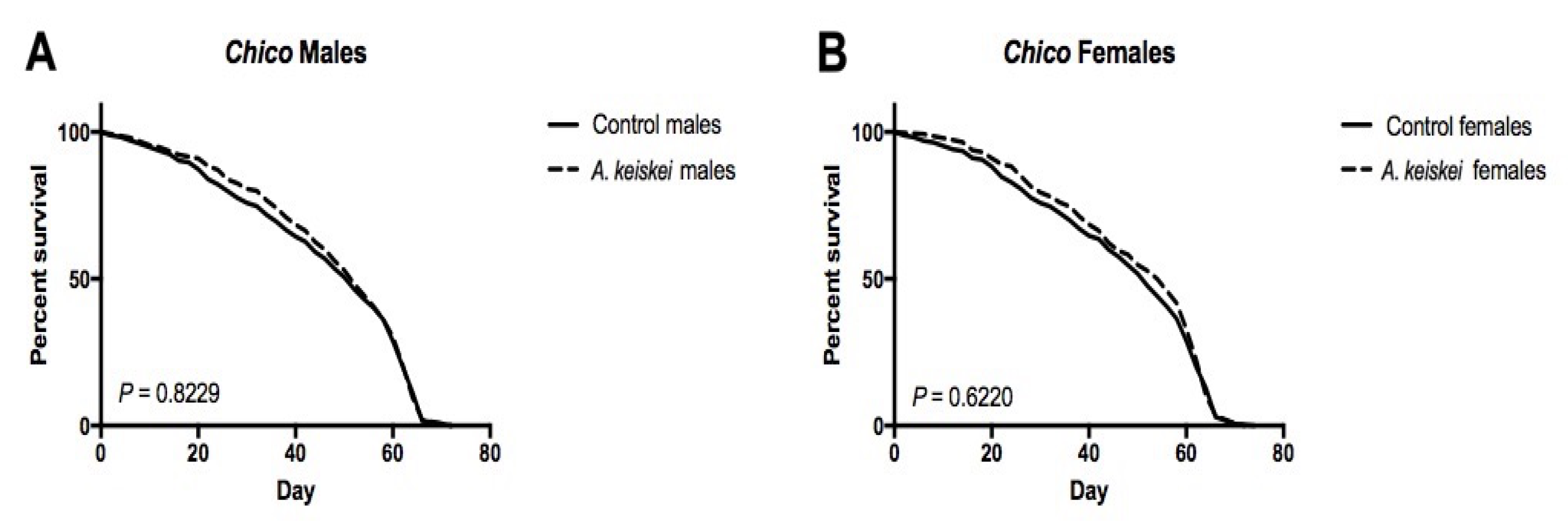
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Preparation of Ethanol Extract of A. keiskei
4.3. Drosophila melanogaster Strains and Culture
4.4. Lifespan Assays
4.5. Stress Resistance Assays
4.6. Climbing Assay
4.7. Fecundity Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amalia, R.; Aulifa, D.L.; Zain, D.N.; Pebiansyah, A.; Levita, J. The Cytotoxicity and Nephroprotective Activity of the Ethanol Extracts of Angelica keiskei Koidzumi Stems and Leaves against the NAPQI-Induced Human Embryonic Kidney (HEK293) Cell Line. Evid.-Based Complement. Altern. Med. 2021, 2021, 1–6. [Google Scholar] [CrossRef]
- Bae, U.-J.; Ryu, J.-H.; Park, B.-H.; Bae, E.J. Angelica keiskei Root Extract Attenuates Bile Duct Ligation-Induced Liver Injury in Mice. J. Med. Food 2022, 25, 435–442. [Google Scholar] [CrossRef]
- Noh, S.; Go, A.; Kim, D.B.; Park, M.; Jeon, H.W.; Kim, B. Role of Antioxidant Natural Products in Management of Infertility: A Review of Their Medicinal Potential. Antioxidants 2020, 9, 957. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Samukawa, Y.; Yamashita, Y.; Ashida, H. 4-Hydroxyderricin and Xanthoangelol Isolated from Angelica keiskei Prevent Dexamethasone-Induced Muscle Loss. Food Funct. 2020, 11, 5498–5512. [Google Scholar] [CrossRef]
- Kil, Y.-S.; Pham, S.T.; Seo, E.K.; Jafari, M. Angelica keiskei, an Emerging Medicinal Herb with Various Bioactive Constituents and Biological Activities. Arch. Pharm. Res. 2017, 40, 655–675. [Google Scholar] [CrossRef]
- Kweon, M.; Lee, H.; Park, C.; Choi, Y.H.; Ryu, J.-H. A Chalcone from Ashitaba (Angelica keiskei) Stimulates Myoblast Differentiation and Inhibits Dexamethasone-Induced Muscle Atrophy. Nutrients 2019, 11, 2419. [Google Scholar] [CrossRef]
- Tu, L.; Wang, R.; Fang, Z.; Sun, M.; Sun, X.; Wu, J.; Dang, Y.; Liu, J. Assessment of the Hypoglycemic and Hypolipidemic Activity of Flavonoid-Rich Extract from Angelica keiskei. Molecules 2022, 27, 6625. [Google Scholar] [CrossRef]
- Zhang, W.; Jin, Q.; Luo, J.; Wu, J.; Wang, Z. Phytonutrient and Anti-Diabetic Functional Properties of Flavonoid-Rich Ethanol Extract from Angelica keiskei Leaves. J. Food Sci. Technol. 2018, 55, 4406–4412. [Google Scholar] [CrossRef]
- Park, C.; Kim, D.H.; Kim, T.H.; Jeong, S.U.; Yoon, J.H.; Moon, S.-K.; Kwon, C.-Y.; Park, S.-H.; Hong, S.H.; Shim, J.-H.; et al. Improvement of Oxidative Stress-Induced Cytotoxicity of Angelica keiskei (Miq.) Koidz. Leaves Extract through Activation of Heme Oxygenase-1 in C2C12 Murine Myoblasts. Biotechnol. Bioproc. E 2023, 28, 51–62. [Google Scholar] [CrossRef]
- Aulifa, D.L.; Adnyana, I.K.; Sukrasno, S.; Levita, J. Inhibitory Activity of Xanthoangelol Isolated from Ashitaba (Angelica keiskei Koidzumi) towards α-Glucosidase and Dipeptidyl Peptidase-IV: In Silico and In Vitro Studies. Heliyon 2022, 8, e09501. [Google Scholar] [CrossRef]
- Mottin, M.; Caesar, L.K.; Brodsky, D.; Mesquita, N.C.M.R.; de Oliveira, K.Z.; Noske, G.D.; Sousa, B.K.P.; Ramos, P.R.P.S.; Jarmer, H.; Loh, B.; et al. Chalcones from Angelica keiskei (Ashitaba) Inhibit Key Zika Virus Replication Proteins. Bioorganic Chem. 2022, 120, 105649. [Google Scholar] [CrossRef]
- Caesar, L.K.; Kellogg, J.J.; Kvalheim, O.M.; Cech, R.A.; Cech, N.B. Integration of Biochemometrics and Molecular Networking to Identify Antimicrobials in Angelica keiskei. Planta Med. 2018, 84, 721–728. [Google Scholar] [CrossRef]
- Karimi-Sales, E.; Mohaddes, G.; Alipour, M.R. Chalcones as Putative Hepatoprotective Agents: Preclinical Evidence and Molecular Mechanisms. Pharmacol. Res. 2018, 129, 177–187. [Google Scholar] [CrossRef]
- Rocha, S.; Ribeiro, D.; Fernandes, E.; Freitas, M. A Systematic Review on Anti-Diabetic Properties of Chalcones. CMC 2020, 27, 2257–2321. [Google Scholar] [CrossRef]
- Martins, T.; Fonseca, B.M.; Rebelo, I. Antioxidant Effects of Chalcones during the Inflammatory Response: An Overall Review. Curr. Med. Chem. 2021, 28, 7658–7713. [Google Scholar] [CrossRef]
- Aoki, N.; Muko, M.; Ohta, E.; Ohta, S. C-Geranylated Chalcones from the Stems of Angelica keiskei with Superoxide-Scavenging Activity. J. Nat. Prod. 2008, 71, 1308–1310. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Q.; Fredimoses, M.; Gao, G.; Wang, K.; Chen, H.; Wang, T.; Oi, N.; Zykova, T.A.; Reddy, K.; et al. The Ashitaba (Angelica keiskei) Chalcones 4-Hydroxyderricin and Xanthoangelol Suppress Melanomagenesis by Targeting BRAF and PI3K. Cancer Prev. Res. 2018, 11, 607–620. [Google Scholar] [CrossRef]
- Ogienko, A.A.; Omelina, E.S.; Bylino, O.V.; Batin, M.A.; Georgiev, P.G.; Pindyurin, A.V. Drosophila as a Model Organism to Study Basic Mechanisms of Longevity. Int. J. Mol. Sci. 2022, 23, 11244. [Google Scholar] [CrossRef]
- Bass, T.M.; Weinkove, D.; Houthoofd, K.; Gems, D.; Partridge, L. Effects of Resveratrol on Lifespan in Drosophila melanogaster and Caenorhabditis Elegans. Mech. Ageing Dev. 2007, 128, 546–552. [Google Scholar] [CrossRef]
- Islam, M.S.; Jin, Y.Y.; Chung, H.-J.; Kim, H.-J.; Baek, S.-H.; Hong, S.-T. Effect of the Resveratrol Rice DJ526 on Longevity. Nutrients 2019, 11, 1804. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Adedara, A.O.; Adie, M.A.; Vicente-Crespo, M.; Farombi, E.O. Resveratrol Prolongs Lifespan and Improves 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Oxidative Damage and Behavioural Deficits in Drosophila melanogaster. Biochem. Biophys Res. Commun. 2018, 503, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Staats, S.; Wagner, A.E.; Kowalewski, B.; Rieck, F.T.; Soukup, S.T.; Kulling, S.E.; Rimbach, G. Dietary Resveratrol Does Not Affect Life Span, Body Composition, Stress Response, and Longevity-Related Gene Expression in Drosophila melanogaster. Int. J. Mol. Sci. 2018, 19, 223. [Google Scholar] [CrossRef] [PubMed]
- Arabit, J.; Elhaj, R.; Schriner, S.E.; Sevrioukov, E.A.; Jafari, M. Rhodiola rosea improves lifespan, locomotion, and neurodegeneration in a Drosophila melanogaster model of Huntington’s disease. Biomed. Res. Int. 2019, 85, 6726874. [Google Scholar]
- Kil, Y.-S.; Nam, J.-W.; Lee, J.; Seo, E.K. Separation of Two Major Chalcones from Angelica keiskei by High-Speed Counter-Current Chromatography. Arch. Pharm. Res. 2015, 38, 1506–1511. [Google Scholar] [CrossRef]
- Rose, M.R.; Charlesworth, B. Genetics of Life History in Drosophila melanogaster. I. Sib analysis of adult females. Genetics 1981, 97, 173–186. [Google Scholar] [CrossRef]
- Rose, M.R.; Drapeau, M.D.; Yazdi, P.G.; Shah, K.H.; Moise, D.B.; Thakar, R.R.; Rauser, C.L.; Mueller, L.D. Evolution of Late-Life Mortality in Drosophila melanogaster. Evolution 2002, 56, 1982–1991. [Google Scholar] [CrossRef] [PubMed]
- Schriner, S.E.; Katoozi, N.S.; Pham, K.Q.; Gazarian, M.; Zarban, A.; Jafari, M. Extension of Drosophila Lifespan by Rosa Damascena Associated with an Increased Sensitivity to Heat. Biogerontology 2012, 13, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Gargano, J.W.; Martin, I.; Bhandari, P.; Grotewiel, M.S. Rapid Iterative Negative Geotaxis (RING): A New Method for Assessing Age-Related Locomotor Decline in Drosophila. Exp. Gerontol. 2005, 40, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Lee, B.-S.; Semnani, S.; Avanesian, A.; Um, C.-Y.; Jeon, H.-J.; Seong, K.-M.; Yu, K.; Min, K.-J.; Jafari, M. Curcumin Extends Life Span, Improves Health Span, and Modulates the Expression of Age-Associated Aging Genes in Drosophila Melanogaster. Rejuvenation Res. 2010, 13, 561–570. [Google Scholar] [CrossRef]
- Zanco, B.; Mirth, C.K.; Sgrò, C.M.; Piper, M.D. A Dietary Sterol trade-off Determines Lifespan Responses to Dietary Restriction in Drosophila melanogaster Females. eLife 2021, 10, e62335. [Google Scholar] [CrossRef]
- Yang, F.; Xiu, M.; Yang, S.; Li, X.; Tuo, W.; Su, Y.; He, J.; Liu, Y. Extension of Drosophila Lifespan by Astragalus Polysaccharide through a Mechanism Dependent on Antioxidant and Insulin/IGF-1 Signaling. Evid. Based Complement. Altern. Med. 2021, 2021, e6686748. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Xie, M.; Luo, L.; Tian, Y.; Yu, G.; Wu, Q.; Fan, X.; Yang, D.; Mao, X.; Gaur, U.; et al. Inhibitor GSK690693 Extends Drosophila Lifespan via Reduce AKT Signaling Pathway. Mech. Ageing Dev. 2022, 202, 111633. [Google Scholar] [CrossRef] [PubMed]
- Lopez, T.; Schriner, S.E.; Okoro, M.; Lu, D.; Chiang, B.T.; Huey, J.; Jafari, M. Green Tea Polyphenols Extend the Lifespan of Male Drosophila melanogaster while Impairing Reproductive Fitness. J. Med. Food 2014, 17, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-N.; Shim, Y.-J.; Kang, B.-H.; Park, J.-J.; Min, B.-H. Over-Expression of Human Clusterin Increases Stress Resistance and Extends Lifespan in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2012, 420, 851–856. [Google Scholar] [CrossRef]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of Life-Span by Loss of CHICO, a Drosophila Insulin Receptor Substrate Protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef]
- Kim, D.W.; Curtis-Long, M.J.; Yuk, H.J.; Wang, Y.; Song, Y.H.; Jeong, S.H.; Park, K.H. Quantitative Analysis of Phenolic Metabolites from Different Parts of Angelica keiskei by HPLC–ESI MS/MS and Their Xanthine Oxidase Inhibition. Food Chem. 2014, 153, 20–27. [Google Scholar] [CrossRef]
- Li, L.; Aldini, G.; Carini, M.; Chen, C.-Y.O.; Chun, H.-K.; Cho, S.-M.; Park, K.-M.; Correa, C.R.; Russell, R.M.; Blumberg, J.B.; et al. Characterisation, Extraction Efficiency, Stability and Antioxidant Activity of Phytonutrients in Angelica keiskei. Food Chem. 2009, 115, 227–232. [Google Scholar] [CrossRef]
- Yan, R.; Yan, J.; Chen, X.; Yu, Y.; Sun, T. Xanthoangelol Prevents Ox-LDL–Induced Endothelial Cell Injury by Activating Nrf2/ARE Signaling. J. Cardiovasc. Pharmacol. 2019, 74, 162. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, S.; Zhai, L.; Sun, L.; Zhao, D.; Wang, Z.; Li, X. Ginsenoside Extract from Ginseng Extends Lifespan and Health Span in Caenorhabditis Elegans. Food Funct. 2021, 12, 6793–6808. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, X.; Zhuang, C.; Lin, Y.; Cao, Y.; Chen, Y. Healthspan Improvements in Caenorhabditis Elegans with Traditional Chinese Herbal Tea. Oxidative Med. Cell. Longev. 2020, 2020, e4057841. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.; Zhu, L.J.; Yen, K.; Tissenbaum, H.A. Uncoupling Lifespan and Healthspan in Caenorhabditis Elegans Longevity Mutants. Proc. Natl. Acad. Sci. USA 2015, 112, E277–E286. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yolitz, J.; Alberico, T.; Laslo, M.; Sun, Y.; Wheeler, C.T.; Sun, X.; Zou, S. Cranberry Interacts with Dietary Macronutrients to Promote Healthy Aging in Drosophila. J. Gerontol. Ser. A 2014, 69, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C. The Plasticity of Aging: Insights from Long-Lived Mutants. Cell 2005, 120, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Leiser, S.F.; Miller, R.A. Nrf2 Signaling, a Mechanism for Cellular Stress Resistance in Long-Lived Mice. Mol. Cell Biol. 2010, 30, 871–884. [Google Scholar] [CrossRef]
- Harper, J.M.; Salmon, A.B.; Leiser, S.F.; Galecki, A.T.; Miller, R.A. Skin-Derived Fibroblasts from Long-Lived Species Are Resistant to Some, but Not All, Lethal Stresses and to the Mitochondrial Inhibitor Rotenone. Aging Cell 2007, 6, 1–13. [Google Scholar] [CrossRef]
- Carmona-Gutierrez, D.; Zimmermann, A.; Kainz, K.; Pietrocola, F.; Chen, G.; Maglioni, S.; Schiavi, A.; Nah, J.; Mertel, S.; Beuschel, C.B.; et al. The Flavonoid 4,4′-Dimethoxychalcone Promotes Autophagy-Dependent Longevity across Species. Nat. Commun. 2019, 10, 651. [Google Scholar] [CrossRef]
- Niraula, P.; Kim, M.S. N-Acetylcysteine Extends Lifespan of Drosophila via Modulating ROS Scavenger Gene Expression. Biogerontology 2019, 20, 533–543. [Google Scholar] [CrossRef]
- Cross, S.T.; Brehm, A.L.; Dunham, T.J.; Rodgers, C.P.; Keene, A.H.; Borlee, G.I.; Stenglein, M.D. Galbut Virus Infection Minimally Influences Drosophila melanogaster Fitness Traits in a Strain and Sex-Dependent Manner. Viruses 2023, 15, 539. [Google Scholar] [CrossRef]
- Ramnarine, T.J.S.; Grath, S.; Parsch, J. Natural Variation in the Transcriptional Response of Drosophila melanogaster to Oxidative Stress. G3 Bethesda 2021, 12, jkab366. [Google Scholar] [CrossRef]
- Malacrida, S.; De Lazzari, F.; Mrakic-Sposta, S.; Vezzoli, A.; Zordan, M.A.; Bisaglia, M.; Menti, G.M.; Meda, N.; Frighetto, G.; Bosco, G.; et al. Lifespan and ROS Levels in Different Drosophila melanogaster Strains after 24 h Hypoxia Exposure. Biol. Open 2022, 11, bio059386. [Google Scholar] [CrossRef]
- Bai, H.; Post, S.; Kang, P.; Tatar, M. Drosophila Longevity Assurance Conferred by Reduced Insulin Receptor Substrate Chico Partially Requires D4eBP. PLoS ONE 2015, 10, e0134415. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jafari, M.; Schriner, S.E.; Kil, Y.-S.; Pham, S.T.; Seo, E.K. Angelica keiskei Impacts the Lifespan and Healthspan of Drosophila melanogaster in a Sex and Strain-Dependent Manner. Pharmaceuticals 2023, 16, 738. https://doi.org/10.3390/ph16050738
Jafari M, Schriner SE, Kil Y-S, Pham ST, Seo EK. Angelica keiskei Impacts the Lifespan and Healthspan of Drosophila melanogaster in a Sex and Strain-Dependent Manner. Pharmaceuticals. 2023; 16(5):738. https://doi.org/10.3390/ph16050738
Chicago/Turabian StyleJafari, Mahtab, Samuel E. Schriner, Yun-Seo Kil, Sally T. Pham, and Eun Kyoung Seo. 2023. "Angelica keiskei Impacts the Lifespan and Healthspan of Drosophila melanogaster in a Sex and Strain-Dependent Manner" Pharmaceuticals 16, no. 5: 738. https://doi.org/10.3390/ph16050738
APA StyleJafari, M., Schriner, S. E., Kil, Y.-S., Pham, S. T., & Seo, E. K. (2023). Angelica keiskei Impacts the Lifespan and Healthspan of Drosophila melanogaster in a Sex and Strain-Dependent Manner. Pharmaceuticals, 16(5), 738. https://doi.org/10.3390/ph16050738