
Castasterone, a Plant Steroid Hormone, Affects Human Small-Cell Lung Cancer Cells and Reverses Multi-Drug Resistance


Abstract
1. Introduction
2. Results
2.1. Cytotoxicity of Castasterone
2.2. Reversal of Drug Resistance by CAS
2.3. Combinations of CAS and Chemotherapeutic Drugs
2.4. CAS Induction of Apoptosis: DNA Fragmentation
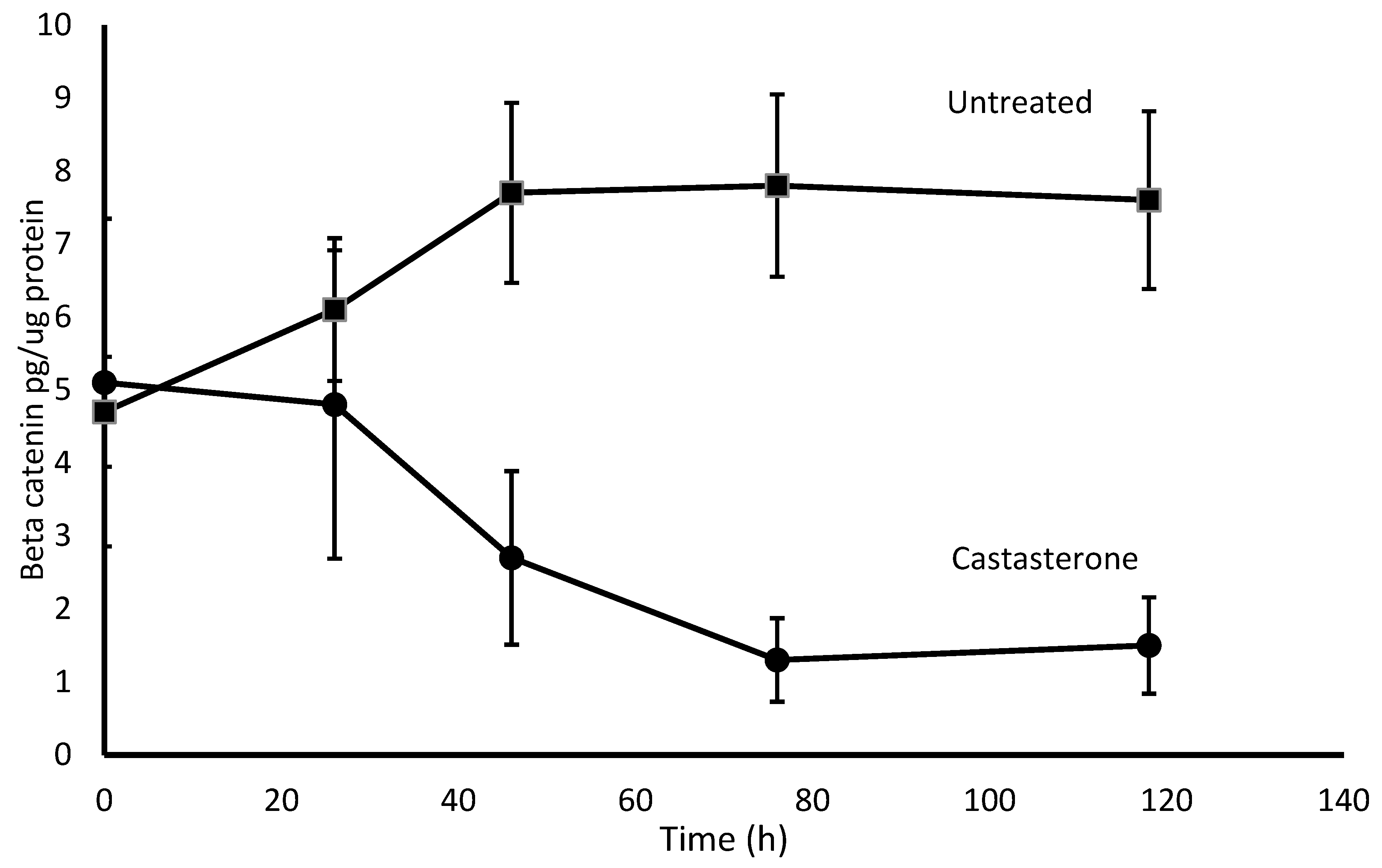
2.5. CAS Effect on β-Catenin Concentration
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Cytotoxicity and Reversal of Resistance
4.3. Combination Studies with CAS, EB, and Chemotherapeutic Drugs
4.4. Apoptosis Analysis by DNA Breaks
4.5. β-Catenin Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parikh, M.; Reiss, J.; Lara, P.N. New and emerging developments in extensive-stage small cell lung cancer therapeutics. Curr. Opin. Oncol. 2016, 28, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Kelly, K. Treatment of refractory and relapsed small cell lung cancer. UpToDate 2019, 1–13. [Google Scholar]
- Rudin, M.M.; Brambilla, E.; Faivre-Finn, C.; Sage, J. Small-cell lung cancer. Nature Rev. Dis. Prim. 2021, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Schulze, A.B.; Evers, G.; Kerckhoff, A.; Mohr, M.; Schliemann, C.; Berdel, W.; Schmidt, L.H. Future options of molecular-targeted therapy in small-cell lung cancer. Cancers 2019, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.; Mandava, N.; Worley, J.; Plimmer, J.; Smith, M. Brassins, a new family of plant hormone from rape pollen. Nature 1970, 252, 1065–1066. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Xing, K.; Zhamg, J.; Liu, H.; Ali, K.; Li, W.; Bai, Q.; Ren, H. Evolutionary analysis and functional identification of ancient brassinosteroid receptors in Ceratopteris richardii. Int. J. Mol. Sci. 2022, 23, 6795. [Google Scholar] [CrossRef] [PubMed]
- Manghwar, H.; Hussain, A.; Ali, Q.; Liu, F. Brassinosteroids (BRs) role in plant development and coping with different stresses. Int. J. Mol. Sci. 2022, 23, 1012. [Google Scholar] [CrossRef] [PubMed]
- Hategan, L.; Godza, B.; Szekersm, S. Regulation of brassinosteroids signaling. In Brassinosteroids: A Class of Plant Hormone; Hyatt, S., Ahmad, A., Eds.; Springer Science & Business Media: Berlin, Germany, 2010; pp. 57–82. [Google Scholar]
- Belkhadr, Y.; Jailais, Y. The molecular circuitry of brassinosteroid signaling. New Phytol. 2015, 206, 522–540. [Google Scholar] [CrossRef] [PubMed]
- Sadava, D.; Kane, S.E. The effect of brassinolide, a plant steroid hormone, on drug-resistant small-cell ling carcinoma cells. Biochem. Biophys. Res. Commun. 2017, 493, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Yokata, T.; Ogino, Y.; Takahashi, N.; Saimoto, H.; Fujioka, S.; Sakurai, A. Brassinolide is biosynthesixed from casasterone in Catharanthus roseus crown gall cells. Agric. Biol. Chem. 1990, 54, 1107–1108. [Google Scholar]
- Roh, J.; Moon, J.; Youn, J.-H.; Seo, C.; Park, Y.; Kim, S.-K. Establishment of biosynthetic pathways to generate castasterone as the biologically active brassinosteroid in Brachypodium distachyon. J. Agric. Food Chem. 2020, 68, 3912–3923. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; An, Z.; Yue, Q.; Zhao, C.; Zhang, S.; Sun, X.; Li, K.; Zhao, L.; Su, L. Effects of brassinosteroid on cancer cells: A review. J. Biochem. Mol. Toxicol. 2022, 36, e23026. [Google Scholar] [CrossRef] [PubMed]
- Lorca, M.; Cabezas, D.; Araque, I.; Teran, A.; Hernandez, S.; Mellado, M.; Espinoza, L.; Mella, J. Cancer and brassinosteroids: Mechanisms of action, SAR and future perspectives. Steroids 2023, 190, 109153. [Google Scholar] [CrossRef] [PubMed]
- Chou, T. Drug combination studies and their synergy quantification using the Chou-Talay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Miller, K.; Fuchs, H.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Ahmad, A. Brassinosteroids, A Class of Plant Hormone; Springer: New York, NY, USA, 2011. [Google Scholar]
- Nelson, W.J.; Nusse, R. Convergence of Wnt, β-catenin and cadherin pathways. Science 2004, 303, 1483–1487. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Devarakonda, S.; Skidmore, Z.; Krysiak, K.; Ramu, A.; Triani, L.; Kunisaki, J.; Masood, A.; Waqar, S.; Spies, N.; et al. Recurrent WNT pathway alterations are frequent in relapsed small cell lung cancer. Nat. Commun. 2018, 9, 3787. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Condition | H69 (IC50, μM) | VPA16 (IC50, μM) |
|---|---|---|
| Etoposide, no preinc. | 0.65 ± 0.35 | 6.70 ± 2.44 a |
| Etoposide, CAS preinc. | 0.57 ± 0.30 | 1.59 ± 0.98 b |
| Doxorubicin, no preinc. | 0.04 ± 0.02 | 0.32 ± 0.20 a |
| Doxorubicin, CAS preinc. | 0.04 ± 0.03 | 0.12 ± 0.03 b |
| Condition | ED75 |
|---|---|
| CAS and Etoposide | 0.77 |
| CAS and Doxorubicin | 0.85 |
| CAS and Epibrassinolide | 1.07 |
| Condition | Single-Strand DNA Breaks, OD405 |
|---|---|
| Untreated | 0.54 ± 0.19 |
| Castasterone | 1.29 ± 0.47 a |
| Epibrassinolide | 1.38 ± 0.21 a |
| Etoposide | 1.35 ± 0.30 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadava, D.; Chen, S. Castasterone, a Plant Steroid Hormone, Affects Human Small-Cell Lung Cancer Cells and Reverses Multi-Drug Resistance. Pharmaceuticals 2023, 16, 170. https://doi.org/10.3390/ph16020170
Sadava D, Chen S. Castasterone, a Plant Steroid Hormone, Affects Human Small-Cell Lung Cancer Cells and Reverses Multi-Drug Resistance. Pharmaceuticals. 2023; 16(2):170. https://doi.org/10.3390/ph16020170
Chicago/Turabian StyleSadava, David, and Shiuan Chen. 2023. "Castasterone, a Plant Steroid Hormone, Affects Human Small-Cell Lung Cancer Cells and Reverses Multi-Drug Resistance" Pharmaceuticals 16, no. 2: 170. https://doi.org/10.3390/ph16020170
APA StyleSadava, D., & Chen, S. (2023). Castasterone, a Plant Steroid Hormone, Affects Human Small-Cell Lung Cancer Cells and Reverses Multi-Drug Resistance. Pharmaceuticals, 16(2), 170. https://doi.org/10.3390/ph16020170