


Potential Chemopreventive Effects of Dietary Combination of Phytochemicals against Cancer Development



{kind=link}
Abstract
1. Introduction
2. Human Carcinogens
3. Metabolic Activation of Carcinogens
4. Phytochemicals
5. Combined Chemoprevention Strategy
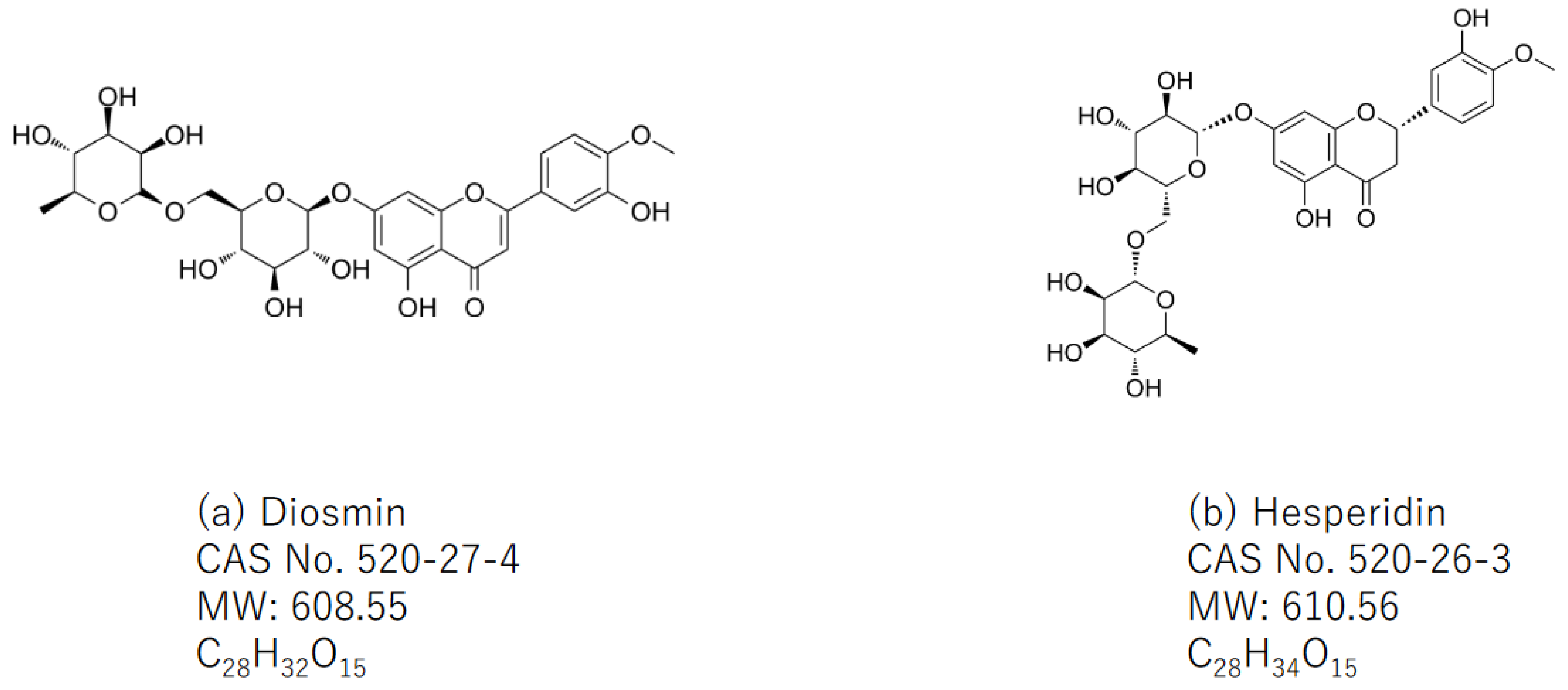
5.1. The Natural Sources of Diosmin and Hesperidin
5.2. Combination Treatment with Two Natural Compounds, Diosmin and Hesperidin
5.3. The Effects of a Single Administration of Diosmin, Hesperidin, or the Combination of Both Compounds
5.4. The Molecular Targets of Diosmin and Hesperidin
5.5. Combination Treatment with a Natural Compound and Synthetic Chemical (Drug)
5.6. Combination Treatment with Synthetic Chemicals (Drugs) with Chemopreventive Effects
6. Cellular Senescence and Chemoprevention
7. Clinical Trials of Chemoprevention Studies Using Phytochemicals and Limitations
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parkin, D.M. Global cancer statistics in the year 2000. Lancet Oncol. 2001, 2, 533–543. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Macias, M.; Canada-Higueras, E.; Alegre, E.; Bielsa, A.; Gracia, J.; Patino-Garcia, A.; Ferrer-Costa, R.; Sendino, T.; Andueza, M.P.; Mateos, B.; et al. Performance comparison of two next-generation sequencing panels to detect actionable mutations in cell-free DNA in cancer patients. Clin. Chem. Lab. Med. 2020, 58, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Sun, S.Q.; Huang, K.L.; Oh, C.; Bailey, M.H.; Varghese, R.; Wyczalkowski, M.A.; Ning, J.; Tripathi, P.; McMichael, J.F.; et al. Integrative omics analyses broaden treatment targets in human cancer. Genome Med. 2018, 10, 60. [Google Scholar] [CrossRef]
- Sunami, K.; Ichikawa, H.; Kubo, T.; Kato, M.; Fujiwara, Y.; Shimomura, A.; Koyama, T.; Kakishima, H.; Kitami, M.; Matsushita, H.; et al. Feasibility and utility of a panel testing for 114 cancer-associated genes in a clinical setting: A hospital-based study. Cancer Sci. 2019, 110, 1480–1490. [Google Scholar] [CrossRef]
- Sporn, M.B.; Dunlop, N.M.; Newton, D.L.; Smith, J.M. Prevention of chemical carcinogenesis by vitamin A and its synthetic analogs (retinoids). Fed. Proc. 1976, 35, 1332–1338. [Google Scholar]
- Tanaka, T. Chemoprevention of human cancer: Biology and therapy. Crit. Rev. Oncol. Hematol. 1997, 25, 139–174. [Google Scholar] [CrossRef]
- Tanaka, T. Effect of diet on human carcinogenesis. Crit. Rev. Oncol. Hematol. 1997, 25, 73–95. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Kamendulis, L.M. The role of oxidative stress in carcinogenesis. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 239–267. [Google Scholar] [CrossRef]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef] [PubMed]
- George, B.P.; Chandran, R.; Abrahamse, H. Role of Phytochemicals in Cancer Chemoprevention: Insights. Antioxidants 2021, 10, 1455. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. Animal models of carcinogenesis in inflamed colorectum: Potential use in chemoprevention study. Curr. Drug Targets 2012, 13, 1689–1697. [Google Scholar] [CrossRef]
- Tanaka, T.; Ishikawa, H. Mast cells and inflammation-associated colorectal carcinogenesis. Semin. Immunopathol. 2013, 35, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Yamada, Y.; Sugie, S.; Mori, H. A novel inflammation-related mouse colon carcinogenesis model induced by azoxymethane and dextran sodium sulfate. Cancer Sci. 2003, 94, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kohno, H.; Murakami, M.; Shimada, R.; Kagami, S. Colitis-related rat colon carcinogenesis induced by 1-hydroxy-anthraquinone and methylazoxymethanol acetate (Review). Oncol. Rep. 2000, 7, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yasui, Y.; Ishigamori-Suzuki, R.; Oyama, T. Citrus compounds inhibit inflammation- and obesity-related colon carcinogenesis in mice. Nutr. Cancer 2008, 60 (Suppl. S1), 70–80. [Google Scholar] [CrossRef]
- Tanaka, T.; Tanaka, T.; Tanaka, M.; Kuno, T. Cancer chemoprevention by citrus pulp and juices containing high amounts of beta-cryptoxanthin and hesperidin. J. Biomed. Biotechnol. 2012, 2012, 516981. [Google Scholar] [CrossRef]
- Wattenberg, L.W. Inhibition of carcinogenesis by minor dietary constituents. Cancer Res. 1992, 52, 2085s–2091s. [Google Scholar]
- Gullett, N.P.; Ruhul Amin, A.R.; Bayraktar, S.; Pezzuto, J.M.; Shin, D.M.; Khuri, F.R.; Aggarwal, B.B.; Surh, Y.J.; Kucuk, O. Cancer prevention with natural compounds. Semin. Oncol. 2010, 37, 258–281. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Khor, T.O.; Shu, L.; Su, Z.Y.; Fuentes, F.; Lee, J.H.; Kong, A.N. Plants vs. cancer: A review on natural phytochemicals in preventing and treating cancers and their druggability. Anticancer Agents Med. Chem. 2012, 12, 1281–1305. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kohno, H.; Mori, H. Chemoprevention of Colon Carcinogenesis by Dietary Non-nutritive Compounds. Asian Pac. J. Cancer Prev. 2001, 2, 165–177. [Google Scholar] [PubMed]
- Nainu, F.; Frediansyah, A.; Mamada, S.S.; Permana, A.D.; Salampe, M.; Chandran, D.; Emran, T.B.; Simal-Gandara, J. Natural products targeting inflammation-related metabolic disorders: A comprehensive review. Heliyon 2023, 9, e16919. [Google Scholar] [CrossRef] [PubMed]
- Banudevi, S.; Swaminathan, S.; Maheswari, K.U. Pleiotropic Role of Dietary Phytochemicals in Cancer: Emerging Perspectives for Combinational Therapy. Nutr. Cancer 2015, 67, 1021–1048. [Google Scholar] [CrossRef] [PubMed]
- Shankar, G.M.; Swetha, M.; Keerthana, C.K.; Rayginia, T.P.; Anto, R.J. Cancer Chemoprevention: A Strategic Approach Using Phytochemicals. Front. Pharmacol. 2021, 12, 809308. [Google Scholar] [CrossRef]
- Wattenberg, L.W. Chemoprevention of cancer. Cancer Res. 1985, 45, 1–8. [Google Scholar] [CrossRef]
- Wattenberg, L.W. Chemoprevention of cancer. Prev. Med. 1996, 25, 44–45. [Google Scholar] [CrossRef]
- Wattenberg, L.W. An overview of chemoprevention: Current status and future prospects. Proc. Soc. Exp. Biol. Med. 1997, 216, 133–141. [Google Scholar] [CrossRef]
- Sporn, M.B. Combination chemoprevention of cancer. Nature 1980, 287, 107–108. [Google Scholar] [CrossRef]
- Anzano, M.A.; Byers, S.W.; Smith, J.M.; Peer, C.W.; Mullen, L.T.; Brown, C.C.; Roberts, A.B.; Sporn, M.B. Prevention of breast cancer in the rat with 9-cis-retinoic acid as a single agent and in combination with tamoxifen. Cancer Res. 1994, 54, 4614–4617. [Google Scholar] [PubMed]
- Sporn, M.B.; Hong, W.K. Concomitant DFMO and sulindac chemoprevention of colorectal adenomas: A major clinical advance. Nat. Clin. Pract. Oncol. 2008, 5, 628–629. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.; Risingsong, R.; Royce, D.B.; Williams, C.R.; Sporn, M.B.; Pioli, P.A.; Gediya, L.K.; Njar, V.C.; Liby, K.T. The combination of the histone deacetylase inhibitor vorinostat and synthetic triterpenoids reduces tumorigenesis in mouse models of cancer. Carcinogenesis 2013, 34, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y. Overview of genotoxic carcinogens and non-genotoxic carcinogens. Exp. Toxicol. Pathol. 1992, 44, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Wilde, E.C.; Chapman, K.E.; Stannard, L.M.; Seager, A.L.; Brusehafer, K.; Shah, U.K.; Tonkin, J.A.; Brown, M.R.; Verma, J.R.; Doherty, A.T.; et al. A novel, integrated in vitro carcinogenicity test to identify genotoxic and non-genotoxic carcinogens using human lymphoblastoid cells. Arch. Toxicol. 2018, 92, 935–951. [Google Scholar] [CrossRef]
- Rosenberg, D.W.; Giardina, C.; Tanaka, T. Mouse models for the study of colon carcinogenesis. Carcinogenesis 2009, 30, 183–196. [Google Scholar] [CrossRef]
- Harris, C.C. Interindividual variation among humans in carcinogen metabolism, DNA adduct formation and DNA repair. Carcinogenesis 1989, 10, 1563–1566. [Google Scholar] [CrossRef][Green Version]
- Miller, J.A. Recent studies on the metabolic activation of chemical carcinogens. Cancer Res. 1994, 54, 1879s–1881s. [Google Scholar]
- Miller, J.A.; Miller, E.C. The metabolic activation and nucleic acid adducts of naturally-occurring carcinogens: Recent results with ethyl carbamate and the spice flavors safrole and estragole. Br. J. Cancer 1983, 48, 1–15. [Google Scholar] [CrossRef][Green Version]
- Straub, K.M.; Burlingame, A.L. Carcinogen binding to DNA. Biomed. Mass Spectrom. 1981, 8, 431–435. [Google Scholar] [CrossRef]
- Van Duuren, B.L. Prediction of carcinogenicity based on structure, chemical reactivity and possible metabolic pathways. J. Environ. Pathol. Toxicol. 1980, 3, 11–34. [Google Scholar] [PubMed]
- Barnes, J.L.; Zubair, M.; John, K.; Poirier, M.C.; Martin, F.L. Carcinogens and DNA damage. Biochem. Soc. Trans. 2018, 46, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Rendic, S.; Guengerich, F.P. Contributions of human enzymes in carcinogen metabolism. Chem. Res. Toxicol. 2012, 25, 1316–1383. [Google Scholar] [CrossRef] [PubMed]
- Cheung, E.C.; Vousden, K.H. The role of ROS in tumour development and progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef]
- Harris, I.S.; DeNicola, G.M. The Complex Interplay between Antioxidants and ROS in Cancer. Trends Cell Biol. 2020, 30, 440–451. [Google Scholar] [CrossRef]
- Kakehashi, A.; Wei, M.; Fukushima, S.; Wanibuchi, H. Oxidative stress in the carcinogenicity of chemical carcinogens. Cancers 2013, 5, 1332–1354. [Google Scholar] [CrossRef]
- Goetz, M.E.; Luch, A. Reactive species: A cell damaging rout assisting to chemical carcinogens. Cancer Lett. 2008, 266, 73–83. [Google Scholar] [CrossRef]
- Ferguson, L.R. Chronic inflammation and mutagenesis. Mutat. Res. 2010, 690, 3–11. [Google Scholar] [CrossRef]
- Yu, Y.; Cui, Y.; Niedernhofer, L.J.; Wang, Y. Occurrence, Biological Consequences, and Human Health Relevance of Oxidative Stress-Induced DNA Damage. Chem. Res. Toxicol. 2016, 29, 2008–2039. [Google Scholar] [CrossRef]
- Linhart, K.; Bartsch, H.; Seitz, H.K. The role of reactive oxygen species (ROS) and cytochrome P-450 2E1 in the generation of carcinogenic etheno-DNA adducts. Redox Biol. 2014, 3, 56–62. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Talalay, P.; Benson, A.M. Elevation of quinone reductase activity by anticarcinogenic antioxidants. Adv. Enzym. Regul. 1982, 20, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Wogan, G.N.; Hecht, S.S.; Felton, J.S.; Conney, A.H.; Loeb, L.A. Environmental and chemical carcinogenesis. Semin. Cancer Biol. 2004, 14, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Sheweita, S.A. Drug-metabolizing enzymes: Mechanisms and functions. Curr. Drug Metab. 2000, 1, 107–132. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P.; Shimada, T. Activation of procarcinogens by human cytochrome P450 enzymes. Mutat. Res. 1998, 400, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T. Inhibition of Carcinogen-Activating Cytochrome P450 Enzymes by Xenobiotic Chemicals in Relation to Antimutagenicity and Anticarcinogenicity. Toxicol. Res. 2017, 33, 79–96. [Google Scholar] [CrossRef]
- Nagao, M.; Sugimura, T.; Matsushima, T. Environmental mutagens and carcinogens. Annu. Rev. Genet. 1978, 12, 117–159. [Google Scholar] [CrossRef]
- Sugimura, T. Nutrition and dietary carcinogens. Carcinogenesis 2000, 21, 387–395. [Google Scholar] [CrossRef]
- Sugimura, T.; Nagao, M.; Wakabayashi, K. Carcinogenicity of food mutagens. Environ. Health Perspect. 1996, 104 (Suppl. S3), 429–433. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Nagao, M.; Esumi, H.; Sugimura, T. Food-derived mutagens and carcinogens. Cancer Res. 1992, 52, 2092s–2098s. [Google Scholar] [PubMed]
- Nebert, D.W.; Dalton, T.P.; Okey, A.B.; Gonzalez, F.J. Role of aryl hydrocarbon receptor-mediated induction of the CYP1 enzymes in environmental toxicity and cancer. J. Biol. Chem. 2004, 279, 23847–23850. [Google Scholar] [CrossRef] [PubMed]
- Stading, R.; Gastelum, G.; Chu, C.; Jiang, W.; Moorthy, B. Molecular mechanisms of pulmonary carcinogenesis by polycyclic aromatic hydrocarbons (PAHs): Implications for human lung cancer. Semin. Cancer Biol. 2021, 76, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.; Desai, D.; Hecht, S.S.; Hoffmann, D. Synthesis of tobacco-specific N-nitrosamines and their metabolites and results of related bioassays. Crit. Rev. Toxicol. 1996, 26, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Hecht, S.S. DNA adduct formation from tobacco-specific N-nitrosamines. Mutat. Res. 1999, 424, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of Phytochemicals in Cancer Prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef]
- Tanaka, T. Cancer chemoprevention by natural-products (review). Oncol. Rep. 1994, 1, 1139–1155. [Google Scholar] [CrossRef]
- Tanaka, T.; Hosokawa, M.; Yasui, Y.; Ishigamori, R.; Miyashita, K. Cancer chemopreventive ability of conjugated linolenic acids. Int. J. Mol. Sci. 2011, 12, 7495–7509. [Google Scholar] [CrossRef]
- Lee, J.H.; Khor, T.O.; Shu, L.; Su, Z.Y.; Fuentes, F.; Kong, A.N. Dietary phytochemicals and cancer prevention: Nrf2 signaling, epigenetics, and cell death mechanisms in blocking cancer initiation and progression. Pharmacol. Ther. 2013, 137, 153–171. [Google Scholar] [CrossRef]
- Baer-Dubowska, W.; Szaefer, H. Modulation of carcinogen-metabolizing cytochromes P450 by phytochemicals in humans. Expert. Opin. Drug Metab. Toxicol. 2013, 9, 927–941. [Google Scholar] [CrossRef]
- Henkler, F.; Brinkmann, J.; Luch, A. The role of oxidative stress in carcinogenesis induced by metals and xenobiotics. Cancers 2010, 2, 376–396. [Google Scholar] [CrossRef] [PubMed]
- Prakobwong, S.; Khoontawad, J.; Yongvanit, P.; Pairojkul, C.; Hiraku, Y.; Sithithaworn, P.; Pinlaor, P.; Aggarwal, B.B.; Pinlaor, S. Curcumin decreases cholangiocarcinogenesis in hamsters by suppressing inflammation-mediated molecular events related to multistep carcinogenesis. Int. J. Cancer 2011, 129, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Yasui, Y.; Kim, M.; Tanaka, T. PPAR Ligands for Cancer Chemoprevention. PPAR Res. 2008, 2008, 548919. [Google Scholar] [CrossRef] [PubMed]
- Tiffon, C. The Impact of Nutrition and Environmental Epigenetics on Human Health and Disease. Int. J. Mol. Sci. 2018, 19, 3425. [Google Scholar] [CrossRef]
- Bayat, S.; Shekari Khaniani, M.; Choupani, J.; Alivand, M.R.; Mansoori Derakhshan, S. HDACis (class I), cancer stem cell, and phytochemicals: Cancer therapy and prevention implications. Biomed. Pharmacother. 2018, 97, 1445–1453. [Google Scholar] [CrossRef]
- Surh, Y.J.; Na, H.K.; Lee, S.S. Transcription factors and mitogen-activated protein kinases as molecular targets for chemoprevention with anti-inflammatory phytochemicals. Biofactors 2004, 21, 103–108. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Y.F. Natural compounds as anticancer agents: Experimental evidence. World J. Exp. Med. 2012, 2, 45–57. [Google Scholar] [CrossRef]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front. Pharmacol. 2019, 10, 1614. [Google Scholar] [CrossRef]
- Kucuk, O. New opportunities in chemoprevention research. Cancer Investig. 2002, 20, 237–245. [Google Scholar] [CrossRef]
- Jorvig, J.E.; Chakraborty, A. Zerumbone inhibits growth of hormone refractory prostate cancer cells by inhibiting JAK2/STAT3 pathway and increases paclitaxel sensitivity. Anticancer Drugs 2015, 26, 160–166. [Google Scholar] [CrossRef]
- Momtazi, A.A.; Sahebkar, A. Difluorinated Curcumin: A Promising Curcumin Analogue with Improved Anti-Tumor Activity and Pharmacokinetic Profile. Curr. Pharm. Des. 2016, 22, 4386–4397. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.Y.; Ediriweera, M.K.; Ryu, J.Y.; Kim, H.Y.; Cho, S.K. Catechol enhances chemo- and radio-sensitivity by targeting AMPK/Hippo signaling in pancreatic cancer cells. Oncol. Rep. 2021, 45, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Islam, J.; Shree, A.; Khan, H.A.; Sultana, S. Chemopreventive potential of Diosmin against benzo[a]pyrene induced lung carcinogenesis in Swiss Albino mice. J. Biochem. Mol. Toxicol. 2022, 36, e23187. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Mohammed, H.M.; Khadrawy, S.M.; Galaly, S.R. Hesperidin protects against chemically induced hepatocarcinogenesis via modulation of Nrf2/ARE/HO-1, PPARgamma and TGF-beta1/Smad3 signaling, and amelioration of oxidative stress and inflammation. Chem. Biol. Interact. 2017, 277, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Roohbakhsh, A.; Parhiz, H.; Soltani, F.; Rezaee, R.; Iranshahi, M. Molecular mechanisms behind the biological effects of hesperidin and hesperetin for the prevention of cancer and cardiovascular diseases. Life Sci. 2015, 124, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.; Rehman, M.U.; Lateef, A.; Khan, R.; Khan, A.Q.; Qamar, W.; Ali, F.; O’Hamiza, O.; Sultana, S. Diosmin protects against ethanol-induced hepatic injury via alleviation of inflammation and regulation of TNF-α and NF-κB activation. Alcohol 2013, 47, 131–139. [Google Scholar] [CrossRef]
- Vafa, A.; Afzal, S.M.; Barnwal, P.; Rashid, S.; Shahid, A.; Alpashree; Islam, J.; Sultana, S. Protective role of diosmin against testosterone propionate-induced prostatic hyperplasia in Wistar rats: Plausible role of oxidative stress and inflammation. Hum. Exp. Toxicol. 2020, 39, 1133–1146. [Google Scholar] [CrossRef]
- Mustafa, S.; Akbar, M.; Khan, M.A.; Sunita, K.; Parveen, S.; Pawar, J.S.; Massey, S.; Agarwal, N.R.; Husain, S.A. Plant metabolite diosmin as the therapeutic agent in human diseases. Curr. Res. Pharmacol. Drug Discov. 2022, 3, 100122. [Google Scholar] [CrossRef]
- Tanaka, T.; Makita, H.; Kawabata, K.; Mori, H.; Kakumoto, M.; Satoh, K.; Hara, A.; Sumida, T.; Tanaka, T.; Ogawa, H. Chemoprevention of azoxymethane-induced rat colon carcinogenesis by the naturally occurring flavonoids, diosmin and hesperidin. Carcinogenesis 1997, 18, 957–965. [Google Scholar] [CrossRef]
- Tanaka, T.; Makita, H.; Ohnishi, M.; Mori, H.; Satoh, K.; Hara, A.; Sumida, T.; Fukutani, K.; Tanaka, T.; Ogawa, H. Chemoprevention of 4-nitroquinoline 1-oxide-induced oral carcinogenesis in rats by flavonoids diosmin and hesperidin, each alone and in combination. Cancer Res. 1997, 57, 246–252. [Google Scholar]
- Yang, M.; Tanaka, T.; Hirose, Y.; Deguchi, T.; Mori, H.; Kawada, Y. Chemopreventive effects of diosmin and hesperidin on N-butyl-N-(4-hydroxybutyl)nitrosamine-induced urinary-bladder carcinogenesis in male ICR mice. Int. J. Cancer 1997, 73, 719–724. [Google Scholar] [CrossRef]
- El-Deek, S.E.M.; Abd-Elghaffar, S.K.H.; Hna, R.S.; Mohamed, H.G.; El-Deek, H.E.M. Effect of Hesperidin against Induced Colon Cancer in Rats: Impact of Smad4 and Activin A Signaling Pathway. Nutr. Cancer 2022, 74, 697–714. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Bedmar, Z.; Anter, J.; Alonso-Moraga, A.; Martin de Las Mulas, J.; Millan-Ruiz, Y.; Guil-Luna, S. Demethylating and anti-hepatocarcinogenic potential of hesperidin, a natural polyphenol of Citrus juices. Mol. Carcinog. 2017, 56, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Kamaraj, S.; Ramakrishnan, G.; Anandakumar, P.; Jagan, S.; Devaki, T. Antioxidant and anticancer efficacy of hesperidin in benzo(a)pyrene induced lung carcinogenesis in mice. Investig. New Drugs 2009, 27, 214–222. [Google Scholar] [CrossRef]
- Saiprasad, G.; Chitra, P.; Manikandan, R.; Sudhandiran, G. Hesperidin alleviates oxidative stress and downregulates the expressions of proliferative and inflammatory markers in azoxymethane-induced experimental colon carcinogenesis in mice. Inflamm. Res. 2013, 62, 425–440. [Google Scholar] [CrossRef]
- Saiprasad, G.; Chitra, P.; Manikandan, R.; Sudhandiran, G. Hesperidin induces apoptosis and triggers autophagic markers through inhibition of Aurora-A mediated phosphoinositide-3-kinase/Akt/mammalian target of rapamycin and glycogen synthase kinase-3 beta signalling cascades in experimental colon carcinogenesis. Eur. J. Cancer 2014, 50, 2489–2507. [Google Scholar] [CrossRef]
- Siddiqi, A.; Saidullah, B.; Sultana, S. Anti-carcinogenic effect of hesperidin against renal cell carcinoma by targeting COX-2/PGE2 pathway in Wistar rats. Environ. Toxicol. 2018, 33, 1069–1077. [Google Scholar] [CrossRef]
- Vabeiryureilai, M.; Lalrinzuali, K.; Jagetia, G.C. Chemopreventive effect of hesperidin, a citrus bioflavonoid in two stage skin carcinogenesis in Swiss albino mice. Heliyon 2019, 5, e02521. [Google Scholar] [CrossRef]
- Tanaka, T.; Makita, H.; Kawabata, K.; Mori, H.; Kakumoto, M.; Satoh, K.; Hara, A.; Sumida, T.; Fukutani, K.; Tanaka, T.; et al. Modulation of N-methyl-N-amylnitrosamine-induced rat oesophageal tumourigenesis by dietary feeding of diosmin and hesperidin, both alone and in combination. Carcinogenesis 1997, 18, 761–769. [Google Scholar] [CrossRef]
- Kohno, H.; Taima, M.; Sumida, T.; Azuma, Y.; Ogawa, H.; Tanaka, T. Inhibitory effect of mandarin juice rich in beta-cryptoxanthin and hesperidin on 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced pulmonary tumorigenesis in mice. Cancer Lett. 2001, 174, 141–150. [Google Scholar] [CrossRef]
- Tanaka, T.; Kohno, H.; Murakami, M.; Shimada, R.; Kagami, S.; Sumida, T.; Azuma, Y.; Ogawa, H. Suppression of azoxymethane-induced colon carcinogenesis in male F344 rats by mandarin juices rich in β-cryptoxanthin and hesperidin. Int. J. Cancer 2000, 88, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Makita, H.; Ohnishi, M.; Hirose, Y.; Wang, A.; Mori, H.; Satoh, K.; Hara, A.; Ogawa, H. Chemoprevention of 4-nitroquinoline 1-oxide-induced oral carcinogenesis by dietary curcumin and hesperidin: Comparison with the protective effect of β-carotene. Cancer Res. 1994, 54, 4653–4659. [Google Scholar] [PubMed]
- Li, L.; Yuan, L.; Chen, X.; Wang, Q.; Tian, J.; Yang, K.; Zhou, E. Current Treatments for Breast Cancer-Related Lymphoedema: A Systematic Review. Asian Pac. J. Cancer Prev. 2016, 17, 4875–4883. [Google Scholar] [CrossRef] [PubMed]
- Pecking, A.P. Evaluation by lymphoscintigraphy of the effect of a micronized flavonoid fraction (Daflon 500 mg) in the treatment of upper limb lymphedema. Int. Angiol. 1995, 14, 39–43. [Google Scholar]
- Pecking, A.P.; Fevrier, B.; Wargon, C.; Pillion, G. Efficacy of Daflon 500 mg in the treatment of lymphedema (secondary to conventional therapy of breast cancer). Angiology 1997, 48, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Sezer, A.; Usta, U.; Kocak, Z.; Yagci, M.A. The effect of a flavonoid fractions diosmin + hesperidin on radiation-induced acute proctitis in a rat model. J. Cancer Res. Ther. 2011, 7, 152–156. [Google Scholar] [CrossRef]
- Jain, A.; Madu, C.O.; Lu, Y. Phytochemicals in Chemoprevention: A Cost-Effective Complementary Approach. J. Cancer 2021, 12, 3686–3700. [Google Scholar] [CrossRef]
- Khuanphram, N.; Taya, S.; Kongtawelert, P.; Wongpoomchai, R. Sesame Extract Promotes Chemopreventive Effect of Hesperidin on Early Phase of Diethylnitrosamine-Initiated Hepatocarcinogenesis in Rats. Pharmaceutics 2021, 13, 1687. [Google Scholar] [CrossRef]
- Huwait, E.; Mobashir, M. Potential and Therapeutic Roles of Diosmin in Human Diseases. Biomedicines 2022, 10, 1076. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F. A mechanistic review of the anticancer potential of hesperidin, a natural flavonoid from citrus fruits. Nutr. Res. 2021, 92, 21–31. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Babiker, A.Y.; Anwar, S. Hesperidin, a Bioflavonoid in Cancer Therapy: A Review for a Mechanism of Action through the Modulation of Cell Signaling Pathways. Molecules 2023, 28, 5152. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, R.; Shi, W.; Li, L.; Liu, H.; Chen, Z.; Wu, L. Metabolism and pharmacological activities of the natural health-benefiting compound diosmin. Food Funct. 2020, 11, 8472–8492. [Google Scholar] [CrossRef]
- Sauter, E.R. Cancer prevention and treatment using combination therapy with natural compounds. Expert. Rev. Clin. Pharmacol. 2020, 13, 265–285. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.S.; Burgeiro, A.; Garcia, R.; Moreno, A.J.; Carvalho, R.A.; Oliveira, P.J. Doxorubicin-induced cardiotoxicity: From bioenergetic failure and cell death to cardiomyopathy. Med. Res. Rev. 2014, 34, 106–135. [Google Scholar] [CrossRef] [PubMed]
- Nonnekens, J.; Hoeijmakers, J.H. After surviving cancer, what about late life effects of the cure? EMBO Mol. Med. 2017, 9, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Wigmore, P.M.; Mustafa, S.; El-Beltagy, M.; Lyons, L.; Umka, J.; Bennett, G. Effects of 5-FU. Adv. Exp. Med. Biol. 2010, 678, 157–164. [Google Scholar] [CrossRef]
- Min, H.Y.; Lee, H.Y. Molecular targeted therapy for anticancer treatment. Exp. Mol. Med. 2022, 54, 1670–1694. [Google Scholar] [CrossRef]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25 (Suppl. S2), 41–59. [Google Scholar] [CrossRef]
- Irshad, R.; Husain, M. Natural products in the reprogramming of cancer epigenetics. Toxicol. Appl. Pharmacol. 2021, 417, 115467. [Google Scholar] [CrossRef]
- Talib, W.H.; Alsayed, A.R.; Barakat, M.; Abu-Taha, M.I.; Mahmod, A.I. Targeting Drug Chemo-Resistance in Cancer Using Natural Products. Biomedicines 2021, 9, 1353. [Google Scholar] [CrossRef]
- Bayat Mokhtari, R.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed]
- Taylor, F.; Huffman, M.D.; Macedo, A.F.; Moore, T.H.; Burke, M.; Davey Smith, G.; Ward, K.; Ebrahim, S. Statins for the primary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2013, 2013, CD004816. [Google Scholar] [CrossRef] [PubMed]
- Joharatnam-Hogan, N.; Alexandre, L.; Yarmolinsky, J.; Lake, B.; Capps, N.; Martin, R.M.; Ring, A.; Cafferty, F.; Langley, R.E. Statins as Potential Chemoprevention or Therapeutic Agents in Cancer: A Model for Evaluating Repurposed Drugs. Curr. Oncol. Rep. 2021, 23, 29. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.S. Therapy insight: Potential of statins for cancer chemoprevention and therapy. Nat. Clin. Pract. Oncol. 2005, 2, 82–89. [Google Scholar] [CrossRef]
- Joo, M.K.; Park, J.J.; Chun, H.J. Additional Benefits of Routine Drugs on Gastrointestinal Cancer: Statins, Metformin, and Proton Pump Inhibitors. Dig. Dis. 2018, 36, 1–14. [Google Scholar] [CrossRef]
- Lochhead, P.; Chan, A.T. Statins and colorectal cancer. Clin. Gastroenterol. Hepatol. 2013, 11, 109–118; quiz e113–e104. [Google Scholar] [CrossRef]
- Poynter, J.N.; Gruber, S.B.; Higgins, P.D.; Almog, R.; Bonner, J.D.; Rennert, H.S.; Low, M.; Greenson, J.K.; Rennert, G. Statins and the risk of colorectal cancer. N. Engl. J. Med. 2005, 352, 2184–2192. [Google Scholar] [CrossRef]
- Rao, C.V.; Tokumo, K.; Rigotty, J.; Zang, E.; Kelloff, G.; Reddy, B.S. Chemoprevention of colon carcinogenesis by dietary administration of piroxicam, α-difluoromethylornithine, 16 alpha-fluoro-5-androsten-17-one, and ellagic acid individually and in combination. Cancer Res. 1991, 51, 4528–4534. [Google Scholar]
- Kojima, T.; Tanaka, T.; Kawamori, T.; Hara, A.; Mori, H. Chemopreventive effects of dietary D,L-α-difluoromethylornithine, an ornithine decarboxylase inhibitor, on initiation and postinitiation stages of diethylnitrosamine-induced rat hepatocarcinogenesis. Cancer Res. 1993, 53, 3903–3907. [Google Scholar]
- Tanaka, T.; Kojima, T.; Hara, A.; Sawada, H.; Mori, H. Chemoprevention of oral carcinogenesis by DL-α-difluoromethylornithine, an ornithine decarboxylase inhibitor: Dose-dependent reduction in 4-nitroquinoline 1-oxide-induced tongue neoplasms in rats. Cancer Res. 1993, 53, 772–776. [Google Scholar]
- Janakiram, N.B.; Mohammed, A.; Bryant, T.; Zhang, Y.; Brewer, M.; Duff, A.; Biddick, L.; Singh, A.; Lightfoot, S.; Steele, V.E.; et al. Potentiating NK cell activity by combination of Rosuvastatin and Difluoromethylornithine for effective chemopreventive efficacy against Colon Cancer. Sci. Rep. 2016, 6, 37046. [Google Scholar] [CrossRef] [PubMed]
- Kemp Bohan, P.M.; Mankaney, G.; Vreeland, T.J.; Chick, R.C.; Hale, D.F.; Cindass, J.L.; Hickerson, A.T.; Ensley, D.C.; Sohn, V.; Clifton, G.T.; et al. Chemoprevention in familial adenomatous polyposis: Past, present and future. Fam. Cancer 2021, 20, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Higurashi, T.; Arimoto, J.; Ashikari, K.; Takatsu, T.; Misawa, N.; Yoshihara, T.; Matsuura, T.; Fuyuki, A.; Ohkubo, H.; Nakajima, A. The efficacy of aspirin and metformin combination therapy in patients with rectal aberrant crypt foci: A double-blinded randomized controlled trial. BMC Cancer 2020, 20, 1043. [Google Scholar] [CrossRef] [PubMed]
- Valverde, A.; Penarando, J.; Canas, A.; Lopez-Sanchez, L.M.; Conde, F.; Hernandez, V.; Peralbo, E.; Lopez-Pedrera, C.; de la Haba-Rodriguez, J.; Aranda, E.; et al. Simultaneous inhibition of EGFR/VEGFR and cyclooxygenase-2 targets stemness-related pathways in colorectal cancer cells. PLoS ONE 2015, 10, e0131363. [Google Scholar] [CrossRef]
- Madka, V.; Patlolla, J.M.R.; Venkatachalam, K.; Zhang, Y.; Pathuri, G.; Stratton, N.; Lightfoot, S.; Janakiram, N.B.; Mohammed, A.; Rao, C.V. Chemoprevention of Colon Cancer by DFMO, Sulindac, and NO-Sulindac Administered Individually or in Combinations in F344 Rats. Cancers 2023, 15, 4001. [Google Scholar] [CrossRef]
- Nicastro, H.L.; Grubbs, C.J.; Margaret Juliana, M.; Bode, A.M.; Kim, M.S.; Lu, Y.; You, M.; Milne, G.L.; Boring, D.; Steele, V.E.; et al. Preventive effects of NSAIDs, NO-NSAIDs, and NSAIDs plus difluoromethylornithine in a chemically induced urinary bladder cancer model. Cancer Prev. Res. 2014, 7, 246–254. [Google Scholar] [CrossRef]
- Half, E.; Arber, N. Colon cancer: Preventive agents and the present status of chemoprevention. Expert. Opin. Pharmacother. 2009, 10, 211–219. [Google Scholar] [CrossRef]
- Carbone, P.P.; Douglas, J.A.; Larson, P.O.; Verma, A.K.; Blair, I.A.; Pomplun, M.; Tutsch, K.D. Phase I chemoprevention study of piroxicam and α-difluoromethylornithine. Cancer Epidemiol. Biomark. Prev. 1998, 7, 907–912. [Google Scholar]
- Jacoby, R.F.; Cole, C.E.; Tutsch, K.; Newton, M.A.; Kelloff, G.; Hawk, E.T.; Lubet, R.A. Chemopreventive efficacy of combined piroxicam and difluoromethylornithine treatment of Apc mutant Min mouse adenomas, and selective toxicity against Apc mutant embryos. Cancer Res. 2000, 60, 1864–1870. [Google Scholar]
- Gronich, N.; Rennert, G. Beyond aspirin-cancer prevention with statins, metformin and bisphosphonates. Nat. Rev. Clin. Oncol. 2013, 10, 625–642. [Google Scholar] [CrossRef]
- Guo, S.; Yu, C.; Feng, S.; Wei, J.; Tong, L.; Li, K.; Gao, Y.; Zhao, P.; Li, T.; Chen, M.; et al. Enabling the drug combination of celecoxib through a spherical co-agglomeration strategy with controllable and stable drug content and good powder properties. Int. J. Pharm. 2022, 626, 122180. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef] [PubMed]
- Niklander, S.E.; Lambert, D.W.; Hunter, K.D. Senescent Cells in Cancer: Wanted or Unwanted Citizens. Cells 2021, 10, 3315. [Google Scholar] [CrossRef] [PubMed]
- Sharpless, N.E.; Sherr, C.J. Forging a signature of in vivo senescence. Nat. Rev. Cancer 2015, 15, 397–408. [Google Scholar] [CrossRef]
- Birch, J.; Gil, J. Senescence and the SASP: Many therapeutic avenues. Genes Dev. 2020, 34, 1565–1576. [Google Scholar] [CrossRef]
- Takasugi, M.; Yoshida, Y.; Ohtani, N. Cellular senescence and the tumour microenvironment. Mol. Oncol. 2022, 16, 3333–3351. [Google Scholar] [CrossRef]
- Gonzalez-Meljem, J.M.; Apps, J.R.; Fraser, H.C.; Martinez-Barbera, J.P. Paracrine roles of cellular senescence in promoting tumourigenesis. Br. J. Cancer 2018, 118, 1283–1288. [Google Scholar] [CrossRef]
- Liu, H.; Xu, Q.; Wufuer, H.; Li, Z.; Sun, R.; Jiang, Z.; Dou, X.; Fu, Q.; Campisi, J.; Sun, Y. Rutin is a potent senomorphic agent to target senescent cells and can improve chemotherapeutic efficacy. Aging Cell 2023, e13921. [Google Scholar] [CrossRef]
- Rudzinska, A.; Juchaniuk, P.; Oberda, J.; Wisniewska, J.; Wojdan, W.; Szklener, K.; Mandziuk, S. Phytochemicals in Cancer Treatment and Cancer Prevention-Review on Epidemiological Data and Clinical Trials. Nutrients 2023, 15, 1896. [Google Scholar] [CrossRef]
- Hu, M.; Wu, B.; Liu, Z. Bioavailability of Polyphenols and Flavonoids in the Era of Precision Medicine. Mol. Pharm. 2017, 14, 2861–2863. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, T.; Aoki, R.; Terasaki, M. Potential Chemopreventive Effects of Dietary Combination of Phytochemicals against Cancer Development. Pharmaceuticals 2023, 16, 1591. https://doi.org/10.3390/ph16111591
Tanaka T, Aoki R, Terasaki M. Potential Chemopreventive Effects of Dietary Combination of Phytochemicals against Cancer Development. Pharmaceuticals. 2023; 16(11):1591. https://doi.org/10.3390/ph16111591
Chicago/Turabian StyleTanaka, Takuji, Ryogo Aoki, and Masaru Terasaki. 2023. "Potential Chemopreventive Effects of Dietary Combination of Phytochemicals against Cancer Development" Pharmaceuticals 16, no. 11: 1591. https://doi.org/10.3390/ph16111591
APA StyleTanaka, T., Aoki, R., & Terasaki, M. (2023). Potential Chemopreventive Effects of Dietary Combination of Phytochemicals against Cancer Development. Pharmaceuticals, 16(11), 1591. https://doi.org/10.3390/ph16111591