

Novel Role of Molecular Hydrogen: The End of Ophthalmic Diseases?

Abstract
1. Introduction
2. Advantages and Potentials of Molecular Hydrogen in Treating Ocular Diseases
3. Mechanisms Underlying the Therapeutic Effects of Molecular Hydrogen
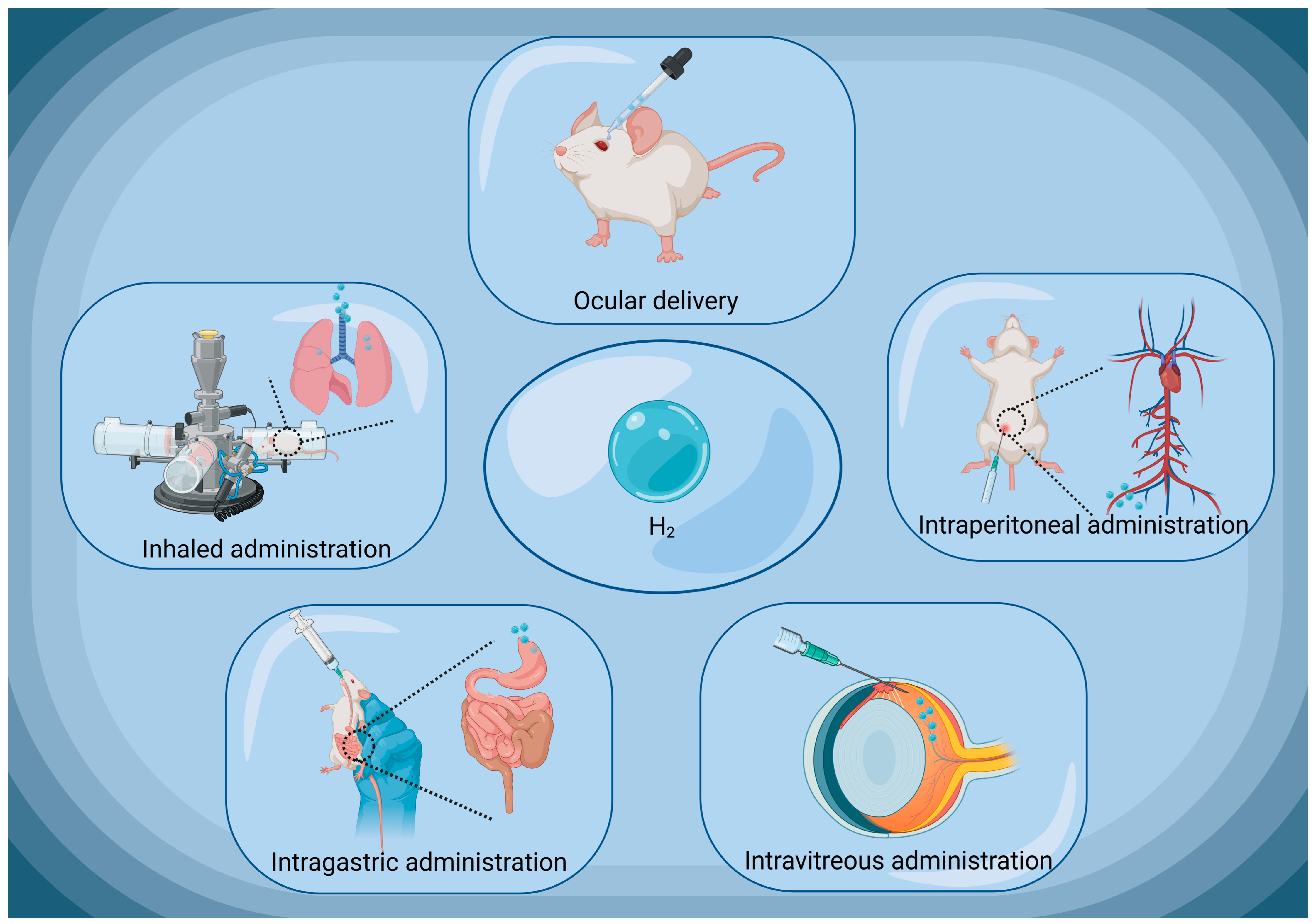
4. Current Administration Approaches of Molecular Hydrogen
4.1. Hydrogen Gas
4.2. Hydrogen-Rich Water/Hydrogen-Rich Saline
4.3. Molecular Hydrogen Produced by Intestinal Bacteria
5. Molecular Hydrogen-Mediated Therapeutic Effects on Ocular Diseases
5.1. Molecular Hydrogen-Mediated Therapeutic Effects on Dry Eye Disease (DED)
5.2. Molecular Hydrogen Protects the Cornea from Alkali Burn
5.3. Molecular Hydrogen-Mediated Therapeutic Effects on Ultraviolet B Ray-Induced Corneal Damage
5.4. The Potential Therapeutic Effects on Corneal Endothelial Dysfunction
5.5. Molecular Hydrogen-Mediated Therapeutic Effects on Cataract
5.6. The Potential Therapeutic Effects against Uveitis
5.7. Molecular Hydrogen-Mediated Therapeutic Effects on Retinitis Pigmentosa
5.8. Molecular Hydrogen-Mediated Therapeutic Effects on Diabetic Retinopathy
5.9. Therapeutic Potential of Molecular Hydrogen in Glaucoma
5.10. Therapeutic Potential of Molecular Hydrogen in Age-Related Macular Degeneration
5.11. Therapeutic Potential of Molecular Hydrogen in Optic Nerve Crush
6. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ohta, S. Recent progress toward hydrogen medicine: Potential of molecular hydrogen for preventive and therapeutic applications. Curr. Pharm. Des. 2011, 17, 2241–2252. [Google Scholar] [CrossRef]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar]
- Cejka, C.; Kubinova, S.; Cejkova, J. The preventive and therapeutic effects of molecular hydrogen in ocular diseases and injuries where oxidative stress is involved. Free Radic. Res. 2019, 53, 237–247. [Google Scholar]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 2016, 1–23. [Google Scholar]
- Karkucinska-Wieckowska, A.; Simoes, I.C.M.; Kalinowski, P.; Lebiedzinska-Arciszewska, M.; Zieniewicz, K.; Milkiewicz, P.; Górska-Ponikowska, M.; Pinton, P.; Malik, A.N.; Krawczyk, M.; et al. Mitochondria, oxidative stress and Non-Alcoholic Fatty Liver Disease: A complex relationship. Eur. J. Clin. Investig. 2022, 52, e13622. [Google Scholar]
- Cui, H.; Kong, Y.; Zhang, H. Oxidative stress, mitochondrial dysfunction, and aging. J. Signal. Transduct. 2012, 2012, 646354. [Google Scholar]
- Franceschelli, S.; Gatta, D.M.P.; Pesce, M.; Ferrone, A.; Di Martino, G.; Di Nicola, M.; De Lutiis, M.A.; Vitacolonna, E.; Patruno, A.; Grilli, A.; et al. Modulation of the oxidative plasmatic state in gastroesophageal reflux disease with the addition of rich water molecular hydrogen: A new biological vision. J. Cell. Mol. Med. 2018, 22, 2750–2759. [Google Scholar]
- Abad-Jiménez, Z.; López-Domènech, S.; Gómez-Abril, S.Á.; Periañez-Gómez, D.; de Marañón, A.M.; Bañuls, C.; Morillas, C.; Víctor, V.M.; Rocha, M. Effect of Roux-en-Y Bariatric Bypass Surgery on Subclinical Atherosclerosis and Oxidative Stress Markers in Leukocytes of Obese Patients: A One-Year Follow-Up Study. Antioxidants 2020, 9, 734. [Google Scholar]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar]
- Shokolenko, I.; Venediktova, N.; Bochkareva, A.; Wilson, G.L.; Alexeyev, M.F. Oxidative stress induces degradation of mitochondrial DNA. Nucleic Acids Res. 2009, 37, 2539–2548. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell. Biol. Vol. 2011, 194, 7–15. [Google Scholar]
- Evans, M.D.; Dizdaroglu, M.; Cooke, M.S. Oxidative DNA damage and disease: Induction, repair and significance. Mutat. Res./Rev. Mutat. Res. 2004, 567, 1–61. [Google Scholar]
- Janssens, S.; Tschopp, J. Signals from within: The DNA-damage-induced NF-kappaB response. Cell Death Differ. 2006, 13, 773–784. [Google Scholar] [PubMed]
- Krishnamoorthy, R.R.; Crawford, M.J.; Chaturvedi, M.M.; Jain, S.K.; Aggarwal, B.B.; Al-Ubaidi, M.R.; Agarwal, N. Photo-oxidative stress down-modulates the activity of nuclear factor-kappaB via involvement of caspase-1, leading to apoptosis of photoreceptor cells. J. Biol. Chem. 1999, 274, 3734–3743. [Google Scholar] [PubMed]
- Bahr, T.; Welburn, K.; Donnelly, J.; Bai, Y. Emerging model systems and treatment approaches for Leber’s hereditary optic neuropathy: Challenges and opportunities. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1866, 165743. [Google Scholar]
- Blasiak, J.; Petrovski, G.; Veréb, Z.; Facskó, A.; Kaarniranta, K. Oxidative Stress, Hypoxia, and Autophagy in the Neovascular Processes of Age-Related Macular Degeneration. Biomed. Res. Int. 2014, 2014, 1–7. [Google Scholar]
- Nam, T. Lipid Peroxidation and Its Toxicological Implications. Toxicol. Res. 2011, 27, 1–6. [Google Scholar] [PubMed]
- Su, L.; Zhang, J.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 1–13. [Google Scholar]
- Hsueh, Y.J.; Chen, Y.N.; Tsao, Y.T.; Cheng, C.M.; Wu, W.C.; Chen, H.C. The Pathomechanism, Antioxidant Biomarkers, and Treatment of Oxidative Stress-Related Eye Diseases. Int. J. Mol. Sci. 2022, 23, 1255. [Google Scholar]
- Sadowska-Bartosz, I.; Bartosz, G.; Grune, T.; Sereikaite, J. Role of Oxidative, Nitrative, and Chlorinative Protein Modifications in Aging and Age-Related Diseases. Oxid. Med. Cell. Longev. 2018, 2018, 1–2. [Google Scholar] [CrossRef]
- Hull, D.S.; Green, K. Oxygen free radicals and corneal endothelium. Lens Eye Toxic. Res. 1989, 6, 87–91. [Google Scholar] [PubMed]
- Kowluru, R.A.; Chan, P. Oxidative Stress and Diabetic Retinopathy. Exp. Diabetes Res. 2007, 2007, 1–12. [Google Scholar]
- Beatty, S.; Koh, H.H.; Phil, M.; Henson, D.; Boulton, M. The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [PubMed]
- Tian, L.; Wen, Y.; Li, S.; Zhang, P.; Wang, Y.; Wang, J.; Cao, K.; Du, L.; Wang, N.; Jie, Y. Benefits and Safety of Astaxanthin in the Treatment of Mild-To-Moderate Dry Eye Disease. Front. Nutr. 2022, 8, 796951. [Google Scholar] [PubMed]
- Totsuka, K.; Ueta, T.; Uchida, T.; Roggia, M.F.; Nakagawa, S.; Vavvas, D.G.; Honjo, M.; Aihara, M. Oxidative stress induces ferroptotic cell death in retinal pigment epithelial cells. Exp. Eye Res. 2019, 181, 316–324. [Google Scholar]
- Fu, Z.; Zhang, J.; Zhang, Y. Role of Molecular Hydrogen in Ageing and Ageing-Related Diseases. Oxid. Med. Cell. Longev. 2022, 2022, 2249749. [Google Scholar]
- Yamamoto, H.; Ichikawa, Y.; Hirano, S.; Sato, B.; Takefuji, Y.; Satoh, F. Molecular Hydrogen as a Novel Protective Agent against Pre-Symptomatic Diseases. Int. J. Mol. Sci. 2021, 22, 7211. [Google Scholar] [CrossRef]
- Gote, V.; Sikder, S.; Sicotte, J.; Pal, D. Ocular Drug Delivery: Present Innovations and Future Challenges. J. Pharmacol. Exp. Ther. 2019, 370, 602–624. [Google Scholar]
- Lanier, O.L.; Manfre, M.G.; Bailey, C.; Liu, Z.; Sparks, Z.; Kulkarni, S.; Chauhan, A. Review of Approaches for Increasing Ophthalmic Bioavailability for Eye Drop Formulations. AAPS Pharmscitech. 2021, 22, 107. [Google Scholar]
- Kulkarni, S.S.; Kompella, U.B. Potential Drug Delivery Approaches for XFS-associated and XFS-associated Glaucoma. J. Glaucoma 2014, 23, S77–S79. [Google Scholar] [CrossRef]
- Yannuzzi, N.A.; Si, N.; Relhan, N.; Kuriyan, A.E.; Albini, T.A.; Berrocal, A.M.; Davis, J.L.; Smiddy, W.E.; Townsend, J.; Miller, D.; et al. Endophthalmitis After Clear Corneal Cataract Surgery: Outcomes Over Two Decades. Am. J. Ophthalmol. 2017, 174, 155–159. [Google Scholar] [CrossRef]
- Uang, H.; Schoenwald, R.D.; Lach, J.L. Corneal Penetration Behavior of P-Blocking Agents 111: In Vitro-In Vivo Correlations. J. Pharm. Sci. 1983, 72, 1279–1281. [Google Scholar]
- Hornof, M.; Toropainen, E.; Urtti, A. Cell culture models of the ocular barriers. Eur. J. Pharm. Biopharm. 2005, 60, 207–225. [Google Scholar]
- Gote, V.; Ansong, M.; Pal, D. Prodrugs and nanomicelles to overcome ocular barriers for drug penetration. Expert Opin. Drug Met. 2020, 16, 885–906. [Google Scholar]
- Nguyen, H.; Eng, S.; Ngo, T.; Dass, C.R. Delivery of therapeutics for deep-seated ocular conditions—Status quo. J. Pharm. Pharmacol. 2018, 70, 994–1001. [Google Scholar]
- Chu, Y.Y.; Hua, N.; Ru, Y.S.; Zhao, S.Z. The protection of hydrogen-rich saline on a rat dry eye model induced by scopolamine hydrobromide. Zhonghua Yan Ke Za Zhi 2017, 11, 363–371. [Google Scholar]
- Kubota, M.; Shimmura, S.; Kubota, S.; Miyashita, H.; Kato, N.; Noda, K.; Ozawa, Y.; Usui, T.; Ishida, S.; Umezawa, K.; et al. Hydrogen and N-acetyl-L-cysteine rescue oxidative stress-induced angiogenesis in a mouse corneal alkali-burn model. Investig. Ophth. Vis. Sci. 2011, 52, 427–433. [Google Scholar] [CrossRef]
- Kasamatsu, M.; Arima, T.; Ikebukuro, T.; Nakano, Y.; Tobita, Y.; Uchiyama, M.; Shimizu, A.; Takahashi, H. Prophylactic Instillation of Hydrogen-Rich Water Decreases Corneal Inflammation and Promotes Wound Healing by Activating Antioxidant Activity in a Rat Alkali Burn Model. Int. J. Mol. Sci. 2022, 23, 9774. [Google Scholar] [PubMed]
- Li, R.; Qu, Y.; Li, X.; Tao, Y.; Yang, Q.; Wang, J.; Diao, Y.; Li, Q.; Fang, Y.; Huang, Y.; et al. Molecular Hydrogen Attenuated N-methyl-N-Nitrosourea Induced Corneal Endothelial Injury by Upregulating Anti-Apoptotic Pathway. Investig. Ophth. Vis. Sci. 2021, 62, 2. [Google Scholar]
- Igarashi, T.; Ohsawa, I.; Kobayashi, M.; Igarashi, T.; Suzuki, H.; Iketani, M.; Takahashi, H. Hydrogen prevents corneal endothelial damage in phacoemulsification cataract surgery. Sci. Rep. 2016, 6, 31190. [Google Scholar]
- Yang, C.; Yan, H.; Ding, T. Hydrogen saline prevents selenite-induced cataract in rats. Mol. Vis. 2013, 19, 1684–1693. [Google Scholar]
- Yan, W.; Chen, T.; Long, P.; Zhang, Z.; Liu, Q.; Wang, X.; An, J.; Zhang, Z. Effects of Post-Treatment Hydrogen Gas Inhalation on Uveitis Induced by Endotoxin in Rats. Med. Sci. Monit. 2018, 24, 3840–3847. [Google Scholar] [CrossRef]
- Yan, W.M.; Zhang, L.; Chen, T.; Zhao, G.H.; Long, P.; An, J.; Zhang, Z.M. Effects of hydrogen-rich saline on endotoxin-induced uveitis. Med. Gas Res. 2017, 7, 9–18. [Google Scholar]
- Igarashi, T.; Ohsawa, I.; Kobayashi, M.; Miyazaki, K.; Igarashi, T.; Kameya, S.; Shiozawa, A.L.; Ikeda, Y.; Miyagawa, Y.; Sakai, M.; et al. Drinking hydrogen water improves photoreceptor structure and function in retinal degeneration 6 mice. Sci. Rep. 2022, 12, 13610. [Google Scholar]
- Tao, Y.; Chen, T.; Fang, W.; Yan, Z.; Yang, Q.; Huang, Y.; Yu, L.; Fan, L. The Comparative Efficiency of Intraperitoneal and Intravitreous Injection of Hydrogen Rich Saline against N-Methyl-N-Nitrosourea Induced Retinal Degeneration: A Topographic Study. Front. Pharmacol. 2017, 8, 587. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, R.; Xu, J.; Sun, J.; Xu, T.; Gu, Q.; Wu, X. Hydrogen-Rich Saline Prevents Early Neurovascular Dysfunction Resulting from Inhibition of Oxidative Stress in STZ-Diabetic Rats. Curr. Eye Res. 2013, 38, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhao, S.; Zhang, J.H.; Sun, X. Hydrogen Saline Treatment Attenuates Hyperoxia-Induced Retinopathy by Inhibition of Oxidative Stress and Reduction of VEGF Expression. Ophthalmic Res. 2012, 47, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hua, N.; Xie, K.; Zhao, T.; Yu, Y. Hydrogen-rich saline reduces cell death through inhibition of DNA oxidative stress and overactivation of poly (ADP-ribose) polymerase-1 in retinal ischemia-reperfusion injury. Mol. Med. Rep. 2015, 12, 2495–2502. [Google Scholar]
- Wang, R.; Wu, J.; Chen, Z.; Xia, F.; Sun, Q.; Liu, L. Postconditioning with inhaled hydrogen promotes survival of retinal ganglion cells in a rat model of retinal ischemia/reperfusion injury. Brain Res. 2016, 1632, 82–90. [Google Scholar]
- Li, R.; Liu, Y.; Xie, J.; Huang, X.; Zhang, L.; Liu, H.; Li, L. Sirt3 mediates the protective effect of hydrogen in inhibiting ROS-induced retinal senescence. Free. Radic. Biol. Med. 2019, 135, 116–124. [Google Scholar]
- Liu, Y.; Li, R.; Xie, J.; Hu, J.; Huang, X.; Ren, F.; Li, L. Protective Effect of Hydrogen on Sodium Iodate-Induced Age-Related Macular Degeneration in Mice. Front. Aging Neurosci. 2018, 10, 389. [Google Scholar] [PubMed]
- Liang, I.; Ko, W.; Hsu, Y.; Lin, Y.; Chang, Y.; Zong, X.; Lai, P.; Chang, D.; Hung, C. The Anti-Inflammatory Effect of Hydrogen Gas Inhalation and Its Influence on Laser-Induced Choroidal Neovascularization in a Mouse Model of Neovascular Age-Related Macular Degeneration. Int. J. Mol. Sci. 2021, 22, 12049. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Wang, X.; Yang, X.; Xiao, Q.; Jiang, F. Protective effect of saturated hydrogen saline against blue light-induced retinal damage in rats. Int. J. Ophthalmol.-Chi 2012, 5, 151–157. [Google Scholar]
- Tian, L.; Zhang, L.; Xia, F.; An, J.; Sugita, Y.; Zhang, Z. Hydrogen-rich saline ameliorates the retina against light-induced damage in rats. Med. Gas Res. 2013, 3, 19. [Google Scholar] [CrossRef][Green Version]
- Sun, J.; Xu, T.; Zuo, Q.; Wang, R.; Qi, A.; Cao, W.; Sun, A.; Sun, X.; Xu, J.; Agudo-Barriuso, M. Hydrogen-rich saline promotes survival of retinal ganglion cells in a rat model of optic nerve crush. PLoS ONE 2014, 9, e99299. [Google Scholar]
- Wei, L.; Ge, L.; Qin, S.; Shi, Y.; Du, C.; Du, H.; Liu, L.; Yu, Y.; Sun, X. Hydrogen-rich saline protects retina against glutamate-induced excitotoxic injury in guinea pig. Exp. Eye Res. 2012, 94, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Kamimura, N.; Igarashi, T.; Takahashi, H.; Ohta, S.; Oharazawa, H. Protective effect of molecular hydrogen against oxidative stress caused by peroxynitrite derived from nitric oxide in rat retina. Clin. Exp. Ophthalmol. 2015, 43, 568–577. [Google Scholar] [CrossRef]
- Penders, J.; Kissner, R.; Koppenol, W.H. ONOOH does not react with H2: Potential beneficial effects of H2 as an antioxidant by selective reaction with hydroxyl radicals and peroxynitrite. Free Radic. Biol. Med. 2014, 75, 191–194. [Google Scholar] [CrossRef]
- Zgorzynska, E.; Dziedzic, B.; Walczewska, A. An Overview of the Nrf2/ARE Pathway and Its Role in Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 9592. [Google Scholar]
- Fu, Z.; Zhang, J. Molecular hydrogen is a promising therapeutic agent for pulmonary disease. J. Zhejiang Univ.-Sci. B 2022, 23, 102–122. [Google Scholar]
- Nogueira, J.E.; Branco, L.G.S. Recent Advances in Molecular Hydrogen Research Reducing Exercise-Induced Oxidative Stress and Inflammation. Curr. Pharm. Des. 2021, 27, 731. [Google Scholar] [PubMed]
- Yao, W.; Guo, A.; Han, X.; Wu, S.; Chen, C.; Luo, C.; Li, H.; Li, S.; Hei, Z. Aerosol inhalation of a hydrogen-rich solution restored septic renal function. Aging 2019, 11, 12097–12113. [Google Scholar]
- Maurer, M.; von Stebut, E. Macrophage inflammatory protein-1. Int. J. Biochem. Cell Biol. 2004, 36, 1882–1886. [Google Scholar] [CrossRef] [PubMed]
- Nickells, R.W.; Zack, D.J. Apoptosis in ocular disease: A molecular overview. Ophthalmic Genet. 1996, 17, 145. [Google Scholar]
- Wang, D.; Wang, L.; Zhang, Y.; Zhao, Y.; Chen, G. Hydrogen gas inhibits lung cancer progression through targeting SMC3. Biomed. Pharmacother. 2018, 104, 788–797. [Google Scholar] [PubMed]
- Yang, M.; So, K.; Lam, W.C.; Lo, A.C.Y. Novel Programmed Cell Death as Therapeutic Targets in Age-Related Macular Degeneration? Int. J. Mol. Sci. 2020, 21, 7279. [Google Scholar]
- Zhao, T.; Guo, X.; Sun, Y. Iron Accumulation and Lipid Peroxidation in the Aging Retina: Implication of Ferroptosis in Age-Related Macular Degeneration. Aging Dis. 2021, 12, 529. [Google Scholar]
- Zanini, D.; Todorovic, N.; Korovljev, D.; Stajer, V.; Ostojic, J.; Purac, J.; Kojic, D.; Vukasinovic, E.; Djordjievski, S.; Sopic, M.; et al. The effects of 6-month hydrogen-rich water intake on molecular and phenotypic biomarkers of aging in older adults aged 70 years and over: A randomized controlled pilot trial. Exp. Gerontol. 2021, 155, 111574. [Google Scholar]
- Milkovic, L.; Zarkovic, N.; Marusic, Z.; Zarkovic, K.; Jaganjac, M. The 4-Hydroxynonenal–Protein Adducts and Their Biological Relevance: Are Some Proteins Preferred Targets? Antioxidants 2023, 12, 856. [Google Scholar]
- Yang, M.; Dong, Y.; He, Q.; Zhu, P.; Zhuang, Q.; Shen, J.; Zhang, X.; Zhao, M. Hydrogen: A Novel Option in Human Disease Treatment. Oxid. Med. Cell. Longev. 2020, 2020, 1–17. [Google Scholar]
- Abraini, J.H.; Gardette-Chauffour, M.C.; Martinez, E.; Rostain, J.C.; Lemaire, C. Psychophysiological reactions in humans during an open sea dive to 500 m with a hydrogen-helium-oxygen mixture. J. Appl. Physiol. 1994, 76, 1113–1118. [Google Scholar] [PubMed]
- Buchholz, B.M.; Kaczorowski, D.J.; Sugimoto, R.; Yang, R.; Wang, Y.; Billiar, T.R.; McCurry, K.R.; Bauer, A.J.; Nakao, A. Hydrogen Inhalation Ameliorates Oxidative Stress in Transplantation Induced Intestinal Graft Injury. Am. J. Transpl. 2008, 8, 2015–2024. [Google Scholar]
- Roberts, B.J.; Fife, W.P.; Corbett, T.H.; Schabel, F.M., Jr. Response of five established solid transplantable mouse tumors and one mouse leukemia to hyperbaric hydrogen. Cancer Treat. Rep. 1978, 62, 1077–1079. [Google Scholar]
- Hong, Y.; Chen, S.; Zhang, J.M. Hydrogen as a selective antioxidant: A review of clinical and experimental studies. J. Int. Med. Res. 2010, 38, 1893–1903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, P.; Hong, N.; Liu, F.; Shan, Y.; Wu, Y.; An, B.; Sang, H.; Kong, Q. Effect and mechanism of hydrogen-rich bath on mice with imiquimod-induced psoriasis. Exp. Derm. 2023, 1–8. [Google Scholar] [CrossRef]
- Chen, T.; Tao, Y.; Yan, W.; Yang, G.; Chen, X.; Cao, R.; Zhang, L.; Xue, J.; Zhang, Z. Protective effects of hydrogen-rich saline against N-methyl-N-nitrosourea-induced photoreceptor degeneration. Exp. Eye Res. 2016, 148, 65–73. [Google Scholar]
- Oharazawa, H.; Igarashi, T.; Yokota, T.; Fujii, H.; Suzuki, H.; Machide, M.; Takahashi, H.; Ohta, S.; Ohsawa, I. Protection of the retina by rapid diffusion of hydrogen: Administration of hydrogen-loaded eye drops in retinal ischemia-reperfusion injury. Investig. Ophth. Vis. Sci. 2010, 51, 487–492. [Google Scholar] [CrossRef]
- Cai, J.; Kang, Z.; Liu, K.; Liu, W.; Li, R.; Zhang, J.H.; Luo, X.; Sun, X. Neuroprotective effects of hydrogen saline in neonatal hypoxia–ischemia rat model. Brain Res. 2009, 1256, 129–137. [Google Scholar]
- Nagata, K.; Nakashima-Kamimura, N.; Mikami, T.; Ohsawa, I.; Ohta, S. Consumption of molecular hydrogen prevents the stress-induced impairments in hippocampus-dependent learning tasks during chronic physical restraint in mice. Neuropsychopharmacology 2009, 34, 501–508. [Google Scholar]
- Nakashima-Kamimura, N.; Mori, T.; Ohsawa, I.; Asoh, S.; Ohta, S. Molecular hydrogen alleviates nephrotoxicity induced by an anti-cancer drug cisplatin without compromising anti-tumor activity in mice. Cancer Chemother. Pharm. 2009, 64, 753–761. [Google Scholar]
- Shimouchi, A.; Nose, K.; Yamaguchi, M.; Ishiguro, H.; Kondo, T. Breath hydrogen produced by ingestion of commercial hydrogen water and milk. Biomark. Insights 2009, 4, 27–32. [Google Scholar]
- Flint, H.J.; Bayer, E.A.; Rincon, M.T.; Lamed, R.; White, B.A. Polysaccharide utilization by gut bacteria: Potential for new insights from genomic analysis. Nat. Rev. Microbiol. 2008, 6, 121–131. [Google Scholar] [PubMed]
- Zhai, X.; Chen, X.; Shi, J.; Shi, D.; Ye, Z.; Liu, W.; Li, M.; Wang, Q.; Kang, Z.; Bi, H.; et al. Lactulose ameliorates cerebral ischemia–reperfusion injury in rats by inducing hydrogen by activating Nrf2 expression. Free Radic. Biol. Med. 2013, 665, 731–741. [Google Scholar]
- Nishimura, N.; Tanabe, H.; Komori, E.; Sasaki, Y.; Inoue, R.; Yamamoto, T. Transplantation of High Hydrogen-Producing Microbiota Leads to Generation of Large Amounts of Colonic Hydrogen in Recipient Rats Fed High Amylose Maize Starch. Nutrients 2018, 10, 144. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, M.; Tsuno, S.; Matsumoto, M.; Tsubota, K. Hydrogen-producing milk to prevent reduction in tear stability in persons using visual display terminals. Ocul. Surf. 2019, 17, 714–721. [Google Scholar] [PubMed]
- Nishimura, N.; Tanabe, H.; Sasaki, Y.; Makita, Y.; Ohata, M.; Yokoyama, S.; Asano, M.; Yamamoto, T.; Kiriyama, S. Pectin and high-amylose maize starch increase caecal hydrogen production and relieve hepatic ischaemia–reperfusion injury in rats. Br. J. Nutr. 2012, 107, 485–492. [Google Scholar]
- Shimouchi, A.; Nose, K.; Takaoka, M.; Hayashi, H.; Kondo, T. Effect of Dietary Turmeric on Breath Hydrogen. Dig. Dis. Sci. 2009, 54, 1725–1729. [Google Scholar] [CrossRef]
- Wolf, P.G.; Biswas, A.; Morales, S.E.; Greening, C.; Gaskins, H.R. H2 metabolism is widespread and diverse among human colonic microbes. Gut Microbes 2016, 7, 235–245. [Google Scholar] [CrossRef]
- GR, G. Physiology and ecology of the sulphate-reducing bacteria. J. Appl. Bacteriol. 1990, 69, 769–797. [Google Scholar]
- Lajoie, S.F.; Bank, S.; Miller, T.L.; Wolin, M.J. Acetate production from hydrogen and [13C]carbon dioxide by the microflora of human feces. Appl. Environ. Microb 1988, 54, 2723–2727. [Google Scholar] [CrossRef]
- Gibson, G.R.; Cummings, J.H.; Macfarlane, G.T. Competition for hydrogen between sulphate-reducing bacteria and methanogenic bacteria from the human large intestine. J. Appl. Bacteriol. 1988, 65, 241–247. [Google Scholar]
- Strocchi, A.; Levitt, M.D. Maintaining intestinal H2 balance: Credit the colonic bacteria. Gastroenterology 1992, 102, 1424–1426. [Google Scholar]
- Fan, D.; Hu, H.; Sun, X.; Meng, X.; Zhang, Y.; Pan, S. Oral administration of lactulose: A novel therapy for acute carbon monoxide poisoning via increasing intestinal hydrogen production. Undersea Hyperb. Med. 2016, 43, 45–48. [Google Scholar]
- Schiffman, R.M. Reliability and Validity of the Ocular Surface Disease Index. Arch. Ophthalmol. 2000, 118, 615. [Google Scholar] [CrossRef] [PubMed]
- Kristin, W. What Is Dry Eye Disease? JAMA-J. Am. Med. Assoc. 2022, 328, 84. [Google Scholar]
- Sophia, S.; Tong, L. Dry eye disease and oxidative stress. Acta Ophthalmol. 2018, 96, e412–e420. [Google Scholar]
- Levitt, M.D. Production and excretion of hydrogen gas in man. N. Engl. J. Med. 1969, 281, 122. [Google Scholar] [CrossRef]
- Hammer, H.F. Colonic hydrogen absorption: Quantification of its effect on hydrogen accumulation caused by bacterial fermentation of carbohydrates. Gut 1993, 34, 818–822. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar]
- Viatour, P.; Merville, M.; Bours, V.; Chariot, A. Phosphorylation of NF-κB and IκB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar]
- Baldwin, A.J. The NF-kappa B and I kappa B proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef] [PubMed]
- Rowe, G.C.; Jiang, A.; Arany, Z. PGC-1 Coactivators in Cardiac Development and Disease. Circ. Res. 2010, 107, 825–838. [Google Scholar] [PubMed]
- Han, B.; Jiang, W.; Liu, H.; Wang, J.; Zheng, K.; Cui, P.; Feng, Y.; Dang, C.; Bu, Y.; Wang, Q.M.; et al. Upregulation of neuronal PGC-1α ameliorates cognitive impairment induced by chronic cerebral hypoperfusion. Theranostics 2020, 10, 2832–2848. [Google Scholar] [CrossRef] [PubMed]
- Crapo, J.D.; Oury, T.; Rabouille, C.; Slot, J.W.; Chang, L.Y. Copper, zinc superoxide dismutase is primarily a cytosolic protein in human cells. Proc. Natl. Acad. Sci. USA 1992, 89, 10405–10409. [Google Scholar] [CrossRef]
- Izadi, M.; Jonaidi-Jafari, N.; Pourazizi, M.; Alemzadeh-Ansari, M.H.; Hoseinpourfard, M.J. Photokeratitis induced by ultraviolet radiation in travelers: A major health problem. J. Postgrad. Med. 2018, 64, 40. [Google Scholar]
- Zhao, C.; Li, W.; Duan, H.; Li, Z.; Jia, Y.; Zhang, S.; Wang, X.; Zhou, Q.; Shi, W. NAD(+) precursors protect corneal endothelial cells from UVB-induced apoptosis. Am. J. Physiol.-Cell Physiol. 2020, 318, C796–C805. [Google Scholar] [CrossRef]
- Čejková, J.; Lojda, Z. Histochemical study on xanthine oxidase activity in the normal rabbit cornea and lens and after repeated irradiation of the eye with UVB rays. Acta Histochem. 1996, 98, 47–52. [Google Scholar] [CrossRef]
- Cejkova, J.; Stipek, S.; Crkovska, J.; Ardan, T. Changes of superoxide dismutase, catalase and glutathione peroxidase in the corneal epithelium after UVB rays. Histochemical and biochemical study. Histol. Histopathol. 2000, 15, 1043–1050. [Google Scholar]
- Gupta, P.K.; Berdahl, J.P.; Chan, C.C.; Rocha, K.M.; Yeu, E.; Ayres, B.; Farid, M.; Lee, W.B.; Beckman, K.A.; Kim, T.; et al. The corneal endothelium: Clinical review of endothelial cell health and function. J. Cataract. Refract. Surg. 2021, 47, 1218–1226. [Google Scholar]
- Parekh, M.; Ferrari, S.; Sheridan, C.; Kaye, S.; Ahmad, S. Concise Review: An Update on the Culture of Human Corneal Endothelial Cells for Transplantation. Stem Cell Transl. Med. 2016, 5, 258–264. [Google Scholar]
- Feizi, S. Corneal endothelial cell dysfunction: Etiologies and management. Adv. Ophthalmol. 2018, 10, 2137551084. [Google Scholar]
- Perone, J.M.; Boiche, M.; Lhuillier, L.; Ameloot, F.; Premy, S.; Jeancolas, A.; Goetz, C.; Neiter, E. Correlation Between Postoperative Central Corneal Thickness and Endothelial Damage After Cataract Surgery by Phacoemulsification. Cornea 2018, 37, 587–590. [Google Scholar]
- Holst, A.; Rolfsen, W.; Svensson, B.; Ollinger, K.; Lundgren, B. Formation of free radicals during phacoemulsification. Curr. Eye Res. 1993, 12, 359. [Google Scholar] [CrossRef]
- Takahashi, H. Corneal Endothelium and Phacoemulsification. Cornea 2016, 35, 3–7. [Google Scholar]
- Price, M.O.; Mehta, J.S.; Jurkunas, U.V.; Price, F.W. Corneal endothelial dysfunction: Evolving understanding and treatment options. Prog. Retin. Eye Res. 2021, 82, 100904. [Google Scholar] [PubMed]
- Igarashi, T.; Ohsawa, I.; Kobayashi, M.; Umemoto, Y.; Arima, T.; Suzuki, H.; Igarashi, T.; Otsuka, T.; Takahashi, H. Effects of Hydrogen in Prevention of Corneal Endothelial Damage During Phacoemulsification: A Prospective Randomized Clinical Trial. Am. J. Ophthalmol. 2019, 207, 10–17. [Google Scholar] [PubMed]
- Moreau, K.L.; King, J.A. Protein misfolding and aggregation in cataract disease and prospects for prevention. Trends Mol. Med. 2012, 18, 273–282. [Google Scholar]
- Cicinelli, M.V.; Buchan, J.C.; Nicholson, M.; Varadaraj, V.; Khanna, R.C. Cataracts. Lancet 2023, 401, 377–389. [Google Scholar]
- Sunkireddy, P.; Jha, S.N.; Kanwar, J.R.; Yadav, S.C. Natural antioxidant biomolecules promises future nanomedicine based therapy for cataract. Colloids Surf. B Biointerfaces 2013, 112, 554–562. [Google Scholar]
- Varma, S.D.; Chand, D.; Sharma, Y.R.; Kuck, J.J.F.; Richards, R.D. Oxidative stress on lens and cataract formation: Role of light and oxygen. Curr. Eye Res. 1984, 3, 35. [Google Scholar]
- Peng, J.; Zheng, T.; Liang, Y.; Duan, L.; Zhang, Y.; Wang, L.; He, G.; Xiao, H.P. Coumaric Acid Protects Human Lens Epithelial Cells against Oxidative Stress-Induced Apoptosis by MAPK Signaling. Oxid. Med. Cell. Longev. 2018, 2018, 1–7. [Google Scholar]
- Krishna, U.; Ajanaku, D.; Denniston, A.K.; Gkika, T. Uveitis: A sight-threatening disease which can impact all systems. Postgrad. Med. J. 2017, 93, 766–773. [Google Scholar]
- Ung, L.; Pattamatta, U.; Carnt, N.; Wilkinson-Berka, J.L.; Liew, G.; White, A.J.R. Oxidative stress and reactive oxygen species: A review of their role in ocular disease. Clin. Sci. 2017, 131, 2865–2883. [Google Scholar]
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [PubMed]
- Campochiaro, P.A.T.A. The mechanism of cone cell death in Retinitis Pigmentosa. Prog. Retin. Eye Res. 2018, 62, 24–37. [Google Scholar]
- Zheng, Z.; Chen, H.; Ke, G.; Fan, Y.; Zou, H.; Sun, X.; Gu, Q.; Xu, X.; Ho, P.C.P. Protective Effect of Perindopril on Diabetic Retinopathy Is Associated with Decreased Vascular Endothelial Growth Factor–to–Pigment Epithelium–Derived Factor Ratio. Diabetes 2009, 58, 954–964. [Google Scholar]
- Forbes, J.M.; Coughlan, M.T.; Cooper, M.E. Oxidative Stress as a Major Culprit in Kidney Disease in Diabetes. Diabetes 2008, 57, 1446–1454. [Google Scholar]
- Guma, M.; Rius, J.; Duong-Polk, K.X.; Haddad, G.G.; Lindsey, J.D.; Karin, M. Genetic and pharmacological inhibition of JNK ameliorates hypoxia-induced retinopathy through interference with VEGF expression. Proc. Natl. Acad. Sci. USA 2009, 106, 8760–8765. [Google Scholar] [CrossRef]
- Dong, A.; Xie, B.; Shen, J.; Yoshida, T.; Yokoi, K.; Hackett, S.F.; Campochiaro, P.A. Oxidative stress promotes ocular neovascularization. J. Cell Physiol. 2009, 219, 544–552. [Google Scholar]
- Anderson, R.E.; Rapp, L.M.; Wiegand, R.D. Lipid peroxidation and retinal degeneration. Curr. Eye Res. 1984, 3, 223–227. [Google Scholar]
- Kowluru, R.A.; Tang, J.; Kern, T.S. Abnormalities of retinal metabolism in diabetes and experimental galactosemia. VII. Effect of long-term administration of antioxidants on the development of retinopathy. Diabetes 2001, 50, 1938–1942. [Google Scholar] [CrossRef] [PubMed]
- Kern, T.S.; Kowluru, R.A.; Engerman, R.L. Abnormalities of retinal metabolism in diabetes or galactosemia: ATPases and glutathione. Investig. Ophthalmol. Vis. Sci. 1994, 35, 2962–2967. [Google Scholar]
- Kowluru, R.A.; Abbas, S.N. Diabetes-induced mitochondrial dysfunction in the retina. Investig. Ophth. Vis. Sci. 2003, 44, 5327. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.A.; Guberski, D.L.; Somogyi-Mann, M.; Grant, M.B. Increased H2O2, vascular endothelial growth factor and receptors in the retina of the BBZ/WOR diabetic rat. Free Radic. Biol. Med. 2000, 28, 91–101. [Google Scholar] [CrossRef]
- Wohaieb, S.A.; Godin, D.V. Alterations in Free Radical Tissue-Defense Mechanisms in Streptozocin-Induced Diabetes in Rat: Effects of Insulin Treatment. Diabetes 1987, 36, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The Pathophysiology and Treatment of Glaucoma: A Review. JAMA J. Am. Med. Assoc. 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N. Glaucoma neuroprotection: What is it? Why is it needed? Can. J. Ophthalmol. 2007, 42, 396–398. [Google Scholar] [CrossRef]
- Dong, Z.; Shinmei, Y.; Dong, Y.; Inafuku, S.; Fukuhara, J.; Ando, R.; Kitaichi, N.; Kanda, A.; Tanaka, K.; Noda, K.; et al. Effect of geranylgeranylacetone on the protection of retinal ganglion cells in a mouse model of normal tension glaucoma. Heliyon 2016, 2, e00191. [Google Scholar] [PubMed]
- Manassakorn, A.; Aupapong, S. Retinal nerve fiber layer defect patterns in primary angle-closure and open-angle glaucoma: A comparison using optical coherence tomography. Jpn. J. Ophthalmol. 2011, 55, 28–34. [Google Scholar] [CrossRef]
- Malishevskaya, T.N.; Yusupov, A.R.; Shatskikh, S.V.; Antipina, N.A.; Klindyuk, T.S.; Bogdanova, D.S. Efficacy and safety of neuroprotection in patients with primary open-angle glaucoma. Vestn. Oftal’mol. 2019, 135, 83. [Google Scholar] [CrossRef]
- Liu, M.; Li, H.; Yang, R.; Ji, D.; Xia, X. GSK872 and necrostatin-1 protect retinal ganglion cells against necroptosis through inhibition of RIP1/RIP3/MLKL pathway in glutamate-induced retinal excitotoxic model of glaucoma. J. Neuroinflamm. 2022, 19, 262. [Google Scholar]
- Izzotti, A.; Bagnis, A.; Sacca, S. The role of oxidative stress in glaucoma. Mutat. Res./Rev. Mutat. Res. 2006, 612, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Geng, L.; Xu, W.; Qin, L.; Peng, G.; Huang, Y. The potential utilizations of hydrogen as a promising therapeutic strategy against ocular diseases. Clin. Risk Manag. 2016, 12, 799–806. [Google Scholar]
- Li, G.; Osborne, N.N. Oxidative-induced apoptosis to an immortalized ganglion cell line is caspase independent but involves the activation of poly(ADP-ribose)polymerase and apoptosis-inducing factor. Brain Res. 2008, 1188, 35–43. [Google Scholar] [CrossRef]
- Tan, W.; Zou, J.; Yoshida, S.; Jiang, B.; Zhou, Y. The Role of Inflammation in Age-Related Macular Degeneration. Int. J. Biol. Sci. 2020, 16, 2989–3001. [Google Scholar] [CrossRef]
- Feher, J.; Kovacs, I.; Artico, M.; Cavallotti, C.; Papale, A.; Balacco Gabrieli, C. Mitochondrial alterations of retinal pigment epithelium in age-related macular degeneration. Neurobiol. Aging 2006, 27, 983–993. [Google Scholar]
- Plafker, S.M.; O’Mealey, G.B.; Szweda, L.I. Mechanisms for countering oxidative stress and damage in retinal pigment epithelium. Int. Rev. Cell Mol. Biol. 2012, 298, 135–177. [Google Scholar] [PubMed]
- Tonade, D.; Kern, T.S. Photoreceptor cells and RPE contribute to the development of diabetic retinopathy. Prog. Retin. Eye Res. 2021, 83, 100919. [Google Scholar]
- Kaarniranta, K.; Uusitalo, H.; Blasiak, J.; Felszeghy, S.; Kannan, R.; Kauppinen, A.; Salminen, A.; Sinha, D.; Ferrington, D. Mechanisms of mitochondrial dysfunction and their impact on age-related macular degeneration. Prog. Retin. Eye Res. 2020, 79, 100858. [Google Scholar]
- Tao, Y.; Jiang, P.; Wei, Y.; Wang, P.; Sun, X.; Wang, H. α-Lipoic Acid Treatment Improves Vision-Related Quality of Life in Patients with Dry Age-Related Macular Degeneration. Tohoku J. Exp. Med. 2016, 240, 209–214. [Google Scholar] [CrossRef]
- Marie, M.; Bigot, K.; Angebault, C.; Barrau, C.; Gondouin, P.; Pagan, D.; Fouquet, S.; Villette, T.; Sahel, J.; Lenaers, G.; et al. Light action spectrum on oxidative stress and mitochondrial damage in A2E-loaded retinal pigment epithelium cells. Cell Death Dis. 2018, 9, 287. [Google Scholar]
- Alaimo, A.; Liñares, G.G.; Bujjamer, J.M.; Gorojod, R.M.; Alcon, S.P.; Martínez, J.H.; Baldessari, A.; Grecco, H.E.; Kotler, M.L. Toxicity of blue led light and A2E is associated to mitochondrial dynamics impairment in ARPE-19 cells: Implications for age-related macular degeneration. Arch. Toxicol. 2019, 93, 1401–1415. [Google Scholar] [PubMed]
- Park, S.; Jang, Y.P. The Protective Effect of Brown-, Gray-, and Blue-Tinted Lenses against Blue LED Light-Induced Cell Death in A2E-Laden Human Retinal Pigment Epithelial Cells. Ophthalmic Res. 2017, 57, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Zha, X.; Wu, G.; Zhang, H.; Yang, Y.; Zhang, Y.; Ma, L. PRDX6 regulates the H2O2 and blue light-induced APRE-19 cell apoptosis via down-regulating and interacting with RARA. Anim. Cells Syst. 2019, 23, 241–245. [Google Scholar] [CrossRef]
- Bien, A.; Seidenbecher, C.I.; Bockers, T.M.; Sabel, B.A.; Kreutz, M.R. Apoptotic versus necrotic characteristics of retinal ganglion cell death after partial optic nerve injury. J. Neurotraum 1999, 16, 153–163. [Google Scholar] [CrossRef]
- Kreutz, M.R.; Seidenbecher, C.I.; Sabel, A.B.A. Molecular plasticity of retinal ganglion cells after partial optic nerve injury. Restor. Neurol. Neurosci. 1999, 14, 127–134. [Google Scholar] [PubMed]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef]
- Li, H.; Luo, Y.; Yang, P.; Liu, J. Hydrogen as a complementary therapy against ischemic stroke: A review of the evidence. J. Neurol. Sci. 2019, 396, 240–246. [Google Scholar] [CrossRef]
- Yoritaka, A.; Kobayashi, Y.; Hayashi, T.; Saiki, S.; Hattori, N. Randomized double-blind placebo-controlled trial of hydrogen inhalation for Parkinson’s disease: A pilot study. Neurol. Sci. 2021, 42, 4767–4770. [Google Scholar] [CrossRef]
- Nie, C.; Ding, X.; Rong, A.; Zheng, M.; Li, Z.; Pan, S.; Yang, W. Hydrogen gas inhalation alleviates myocardial ischemia-reperfusion injury by the inhibition of oxidative stress and NLRP3-mediated pyroptosis in rats. Life Sci. 2021, 272, 119248. [Google Scholar] [CrossRef]
- Lazar, H.L. Molecular hydrogen: A novel therapy for the treatment of pulmonary hypertension. J. Thorac. Cardiovasc. Surg. 2015, 150, 654–655. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.; Dressler, N.; Ben-Shushan, R.S.; Meerson, A.; LeBaron, T.W.; Tamir, S. Effects of alkaline-electrolyzed and hydrogen-rich water, in a high-fat-diet nonalcoholic fatty liver disease mouse model. World J. Gastroenterol. WJG 2018, 24, 5095–5108. [Google Scholar] [PubMed]
- Koyama, Y.; Taura, K.; Hatano, E.; Tanabe, K.; Yamamoto, G.; Nakamura, K.; Yamanaka, K.; Kitamura, K.; Narita, M.; Nagata, H.; et al. Effects of oral intake of hydrogen water on liver fibrogenesis in mice. Hepatol. Res. 2014, 44, 663–677. [Google Scholar] [PubMed]
- Najmanová, E.; Manethová, H.; Botek, M.; Pluháček, F. Vliv Rychlého Příjmu Vodíkem Sycené Vody na Nitrooční Tlak u Zdravých Osob. Cesk. Slov. Oftalmol. 2023, 79, 180–184. [Google Scholar]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ocular Diseases | Experimental Objects | Disease Models | Methods of H2 Uptake | Effect of H2 | Refs. |
|---|---|---|---|---|---|
| Dry eye disease | Wistar rats | Scopolamine-induced DED | Intraperitoneal injection of HRS (0.6 mmol/L) at a dose of 5 mL/kg BW daily for 28 days; dropping HRS into the eye 1 time per hour, 9 times per day | Inhibiting the activity of NF-kB to reduce inflammation | [36] |
| Alkali burn of cornea | SOD-1−/− mice or WT mice | Alkali burn | Irrigated with HRW (0.5–0.6 ppm) onto the cornea for 30 min | Reducing oxidative stress; inhibiting angiogenesis in cornea | [37] |
| Wistar rats | Irrigated with HRW (1.2–1.6 ppm) onto the cornea for 5 min | Upregulating the expression of antioxidants | [38] | ||
| UVB | SOD-1−/− mice or WT mice | Ultraviolet B ray-induced corneal damage model | Irrigated with HRW (0.5–0.6 ppm) onto the cornea | Reducing oxidative stress; inhibiting of angiogenesis in cornea | [37] |
| Corneal endothelial dysfunction | Rabbits | MNU-induced corneal endothelial cell injury | Irrigated with HRS (1.2 ppm) 3 times a day for 3 min and 3 drops per second for 7 days | Anti-apoptotic effect through the NF-κB/NLRP3 and FOXO3a/p53/p21 pathway | [39] |
| Rabbits | Corneal endothelial dysfunction induced by phacoemulsification | Ultrasound oscillation with irrigation solution at almost 61% H2 dissolved concentration for 30 s | Reducing oxidative stress | [40] | |
| Cataract | Rats | Selenite-induced cataract | Intraperitoneal injection of HRS (0.6 mmol/L) at a dose of 5 mL/kg BW daily from postnatal day 8 to postnatal day 17 | Maintaining the activity of antioxidant enzymes, inhibiting lipid peroxidation | [41] |
| Uveitis | Rats | Endotoxin-induced uveitis | Inhaling mixed gas that consisted of 67% H2 and 33% O2 for once a day for 3 weeks | Suppressing the microglia activation | [42] |
| Rats | Intraperitoneal injection of HRS (0.6 mM) at a dose of 10 mL/kg BW once a day for 1 week | Maintaining the integrity of the blood–aqueous barrier | [43] | ||
| RP | Rats | Rd6 rats | Drinking HRW (1.2–1.6 ppm) 3.42 ± 0.14 mL/day for 1 week | Neuroprotective effect | [44] |
| Rats | MNU-induced RP | Intraperitoneal (10 mL/kg BW) and intravitreous (8 μL) injections of HRS (0.6 mmol/L) | Increasing the level of SOD, modulating the expressions of apoptosis-related genes | [45] | |
| DR | Male rats | Rats with streptozotocin-induced diabetes mellitus | Intraperitoneal injections of HRS (0.86 mmol/L) at a dose of 5 mL/kg BW daily for 1 month | Reducing oxidative stress; preserving synaptophysin and BDNF levels | [46] |
| C57BL/6J mice | Rats with diabetes mellitus | Intraperitoneal injections of HRS | Reducing the retinal neovascularization, and the expression of VEGF and MDA | [47] | |
| Glaucoma | Rats | Retinal ischemia/reperfusion | Consecutive peritoneally injected of HRS (0.6 mM) at a dose of 5 mL/kg BW until the rats were sacrificed | Alleviating apoptosis of RGCs by overactivating PARP-1 | [48] |
| Inhaling mixed gas that consisted of 67% H2 and 33% O2 for 1 h daily for 7 days | Lessening RGCs loss; reducing the levels of IL1-β, TNF-α and 4-HNE | [49] | |||
| AMD | Mice | NaIO3-induced AMD | Intragastric administration of HRS (4.0 mg/L) at a dose of 10 mL/kg BW for 12 days. | Inhibiting cellular senescence; maintaining DNA homeostasis | [50] |
| Intragastric administration of HRW (0.55~0.65 mM) at a dose of 1 mL/g three times daily for 12 days | Inhibiting oxidative stress and apoptosis | [51] | |||
| Laser-induced CNV mouse | Inhaling mixed gas that consisted of 21% oxygen, 42% H2 and 37% nitrogen gas for 2/5 h daily for 15 days | Alleviating CNV leakage | [52] | ||
| Light-induced retinal damage | Rats | Blue light-induced damage model | Intraperitoneal injection of saturated HRS (0.6 mmol/L) at a dose of 1 mL/100 g BW once a day before and during the exposure session | Suppressing photo-oxidative stress | [53] |
| Intraperitoneal injection of saturated HRS(5 mL/kg) before and within 5 days after light exposure | [54] | ||||
| Optic nerve injury | Rats | Establishing the optic nerve crush model via surgery | Intraperitoneal injection of saturated HRS (5 mL/kg) at 6:00 and 18:00 lasting for 2 weeks | Reducing the lipid peroxidation and apoptosis of RGCs | [55] |
| Guinea pigs | Glutamate-induced retinal injury model | Intravitreous (almost 0.6 mmol/L) and/or peritoneal injection (5 mL/kg) of HRS | Clearing glutamate by increasing EAAT-1; reducing RGCs apoptosis of by upregulating GRP78 | [56] | |
| RGCs cells | S-nitroso-N-acetylpenicillamine-induced oxidative stress model | Culturing cell in medium consisting of 5% O2 and 95% H2 (v/v) for 24–72 h | Suppressing ONOO−-mediated oxidative stress by clearing peroxynitrit | [57] |
| Method | Advantages | Disadvantages | Preparation Method | Administration Method | Ref. |
|---|---|---|---|---|---|
| H2 gas | Rapid and reliable | Risk of explosion, requires strict management and monitoring | Prepared using H2 generators | Direct inhalation of H2 gas under professional guidance | [2,74] |
| Insignificant effect on blood pressure | Inconveniences during usage | ||||
| HRW/HRS | Simple administration through drinking or injection, easily accepted by patients | Limited storage and duration in the body, limited treatment effect | H2 gas injection under high pressure | Oral ingestion or drinking, controlled dosage and frequency as per medical advice | [36,38,56,75,76,77,78,79,81] |
| Partly mitigates the risk of direct H2 gas usage | Reaction between metals and water | Injection under professional healthcare personnel | |||
| Electrolysis to produce HRW/HRS | Through drip infusion, controlled drip rate and dosage | ||||
| Local eye drops | |||||
| H2 produced by intestinal bacteria | Production of H2 by intestinal bacteria, providing longer duration of effect | Variability due to individual differences in gut microbiota affecting H2 production | Induced by ingesting non-digestible substances | Through normal dietary intake of non-digestible components | [82,84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.-Y.; Xue, R.-Y.; Wu, H.; Pu, N.; Wei, D.; Zhao, N.; Song, Z.-M.; Tao, Y. Novel Role of Molecular Hydrogen: The End of Ophthalmic Diseases? Pharmaceuticals 2023, 16, 1567. https://doi.org/10.3390/ph16111567
Li S-Y, Xue R-Y, Wu H, Pu N, Wei D, Zhao N, Song Z-M, Tao Y. Novel Role of Molecular Hydrogen: The End of Ophthalmic Diseases? Pharmaceuticals. 2023; 16(11):1567. https://doi.org/10.3390/ph16111567
Chicago/Turabian StyleLi, Si-Yu, Rong-Yue Xue, Hao Wu, Ning Pu, Dong Wei, Na Zhao, Zong-Ming Song, and Ye Tao. 2023. "Novel Role of Molecular Hydrogen: The End of Ophthalmic Diseases?" Pharmaceuticals 16, no. 11: 1567. https://doi.org/10.3390/ph16111567
APA StyleLi, S.-Y., Xue, R.-Y., Wu, H., Pu, N., Wei, D., Zhao, N., Song, Z.-M., & Tao, Y. (2023). Novel Role of Molecular Hydrogen: The End of Ophthalmic Diseases? Pharmaceuticals, 16(11), 1567. https://doi.org/10.3390/ph16111567