
Analysis of miRNAs in Osteogenesis imperfecta Caused by Mutations in COL1A1 and COL1A2: Insights into Molecular Mechanisms and Potential Therapeutic Targets

 , , , , , , ,
, , , , , , , 
Abstract
:1. Introduction
2. Results
2.1. Mutation Analysis in the COL1A1 and COL1A2 Genes from OI Type I
2.2. Identification of miRNAs in OI Type I
2.3. Expression Analysis of miRNAs in OI
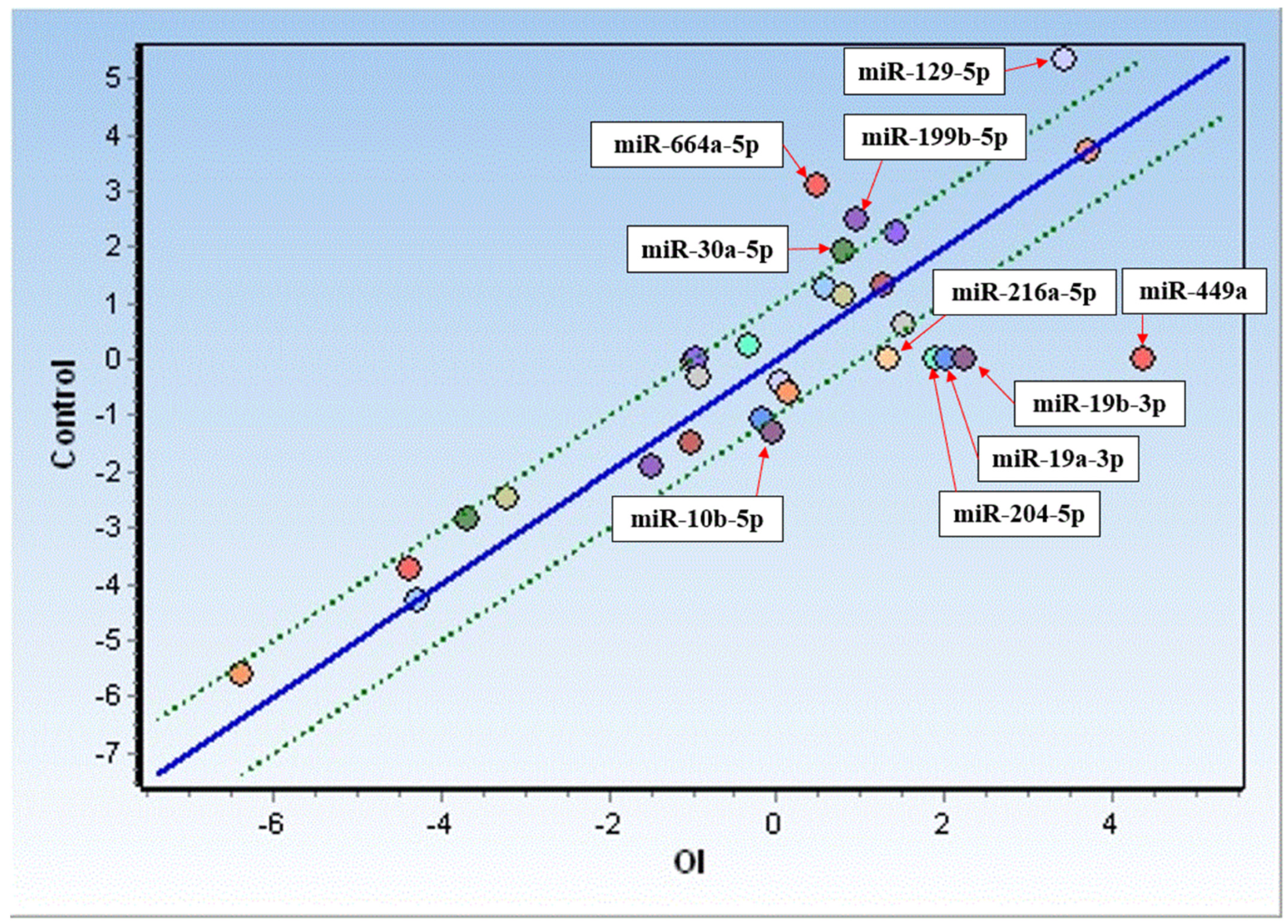
2.4. miRNA Correlation Analysis in OI
2.5. Association of Identified miRNAs with Mutations in COL1A1 and COL1A2
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Participants and Control Material
5.2. Isolation Procedure of Skin Fibroblasts from OI Donors
5.3. Fibroblast Cultures
5.4. Fibroblast DNA Isolation and Amplification
5.5. Sequence Analyses of COL1A1 and COL1A2 Genes from OI Donors
5.6. Fibroblast RNA Isolation
5.7. Real-Time Quantitative Polymerase Chain Reaction
5.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sillence, D.O.; Senn, A.; Danks, D.M. Genetic heterogeneity in osteogenesis imperfecta. J. Med. Genet. 1979, 16, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Byers, P.H.; Cole, W.G. Osteogenesis Imperfecta. In Connective Tissue and Its Heritable Disorders: Molecular, Genetic, and Medical Aspects; Wiley: Hoboken, NJ, USA; pp. 385–430. [CrossRef]
- Van Dijk, F.S.; Pals, G.; Van Rijn, R.R.; Nikkels, P.G.J.; Cobben, J.M. Classification of Osteogenesis Imperfecta revisited. Eur. J. Med. Genet. 2010, 53, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.M.; Chang, W.; Morello, R.; Cabral, W.A.; Weis, M.; Eyre, D.R.; Leikin, S.; Makareeva, E.; Kuznetsova, N.; Uveges, T.E.; et al. Deficiency of Cartilage-Associated Protein in Recessive Lethal Osteogenesis Imperfecta. N. Engl. J. Med. 2006, 355, 2757–2764. [Google Scholar] [CrossRef] [PubMed]
- Cabral, W.A.; Chang, W.; Barnes, A.M.; Weis, M.; Scott, M.A.; Leikin, S.; Makareeva, E.; Kuznetsova, N.V.; Rosenbaum, K.N.; Tifft, C.J.; et al. Prolyl 3-hydroxylase 1 deficiency causes a recessive metabolic bone disorder resembling lethal/severe osteogenesis imperfecta. Nat Genet. 2007, 39, 359–365. [Google Scholar] [CrossRef]
- Christiansen, H.E.; Schwarze, U.; Pyott, S.M.; AlSwaid, A.; Al Balwi, M.; Alrasheed, S.; Pepin, M.G.; Weis, M.A.; Eyre, D.R.; Byers, P.H. Homozygosity for a Missense Mutation in SERPINH1, which Encodes the Collagen Chaperone Protein HSP47, Results in Severe Recessive Osteogenesis Imperfecta. Am. J. Hum. Genet. 2010, 86, 389. [Google Scholar] [CrossRef] [PubMed]
- Asharani, P.V.; Keupp, K.; Semler, O.; Wang, W.; Li, Y.; Thiele, H.; Yigit, G.; Pohl, E.; Becker, J.; Frommolt, P.; et al. Attenuated BMP1 function compromises osteogenesis, leading to bone fragility in humans and zebrafish. Am. J. Hum. Genet. 2012, 90, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Roughley, P.J.; Rauch, F.; Glorieux, F.H. Osteogenesis imperfecta—clinical and molecular diversity. Eur. Cell. Mater. 2003, 5, 41–47; discussion 47. [Google Scholar] [CrossRef]
- Augusciak-Duma, A.; Witecka, J.; Sieron, A.L.; Janeczko, M.; Pietrzyk, J.J.; Ochman, K.; Galicka, A.; Borszewska-Kornacka, M.K.; Pilch, J.; Jakubowska-Pietkiewicz, E.; et al. Mutations in the COL1A1 and COL1A2 genes associated with osteogenesis imperfecta (OI) types I or III. Acta Biochim. Pol. 2018, 65, 79–86. [Google Scholar] [CrossRef]
- Botor, M.; Fus-Kujawa, A.; Uroczynska, M.; Stepien, K.L.; Galicka, A.; Gawron, K.; Sieron, A.L. Osteogenesis Imperfecta: Current and Prospective Therapies. Biomolecules 2021, 11, 1493. [Google Scholar] [CrossRef]
- Trejo, P.; Rauch, F. Osteogenesis imperfecta in children and adolescents—New developments in diagnosis and treatment. Osteoporos. Int. 2016, 27, 3427–3437. [Google Scholar] [CrossRef]
- Alharbi, S.A. A Systematic Overview of Osteogenesis Imperfecta. Mol. Biol. 2015, 5, 150. [Google Scholar] [CrossRef]
- Deguchi, M.; Tsuji, S.; Katsura, D.; Kasahara, K.; Kimura, F.; Murakami, T. Current Overview of Osteogenesis Imperfecta. Medicina 2021, 57, 464. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, F.S.; Huizer, M.; Kariminejad, A.; Marcelis, C.L.; Plomp, A.S.; Terhal, P.A.; Meijers-Heijboer, H.; Weiss, M.M.; Van Rijn, R.R.; Cobben, J.M.; et al. Complete COL1A1 allele deletions in osteogenesis imperfecta. Genet. Med. 2010, 12, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Mortier, G.R.; Cohn, D.H.; Cormier-Daire, V.; Hall, C.; Krakow, D.; Mundlos, S.; Nishimura, G.; Robertson, S.; Sangiorgi, L.; Savarirayan, R.; et al. Nosology and classification of genetic skeletal disorders: 2019 revision. Am. J. Med. Genet. A 2019, 179, 2393–2419. [Google Scholar] [CrossRef]
- Cai, Y.; Yu, X.; Hu, S.; Yu, J. A brief review on the mechanisms of miRNA regulation, Genomics. Proteom. Bioinform. 2009, 7, 147–154. [Google Scholar] [CrossRef]
- Kaneto, C.M.; Lima, P.S.P.; Zanette, D.L.; Prata, K.L.; Neto, J.M.P.; de Paula, F.J.A.; Silva, W.A. COL1A1 and miR-29b show lower expression levels during osteoblast differentiation of bone marrow stromal cells from Osteogenesis Imperfecta patients. BMC Med. Genet. 2014, 15, 45. [Google Scholar] [CrossRef]
- Wang, Z.; Lu, Y.; Zhang, X.; Xu, C.; Han, J.; Ren, X.; Wang, Y.; Li, Z. Serum microRNA is a promising biomarker for osteogenesis imperfecta. Intractable Rare Dis. Res. 2012, 1, 81. [Google Scholar] [CrossRef]
- Kocijan, R.; Muschitz, C.; Geiger, E.; Skalicky, S.; Baierl, A.; Dormann, R.; Plachel, F.; Feichtinger, X.; Heimel, P.; Fahrleitner-Pammer, A.; et al. Circulating microRNA Signatures in Patients With Idiopathic and Postmenopausal Osteoporosis and Fragility Fractures. J. Clin. Endocrinol. Metab. 2016, 101, 4125–4134. [Google Scholar] [CrossRef]
- Yang, G.; Wang, J.; Huang, B. Loss of miR-217 promotes osteosarcoma cell proliferation through targeting SETD8. Pharmazie 2018, 73, 711–714. [Google Scholar] [CrossRef]
- Dai, Z.; Jin, Y.; Zheng, J.; Liu, K.; Zhao, J.; Zhang, S.; Wu, F.; Sun, Z. MiR-217 promotes cell proliferation and osteogenic differentiation of BMSCs by targeting DKK1 in steroid-associated osteonecrosis. Biomed. Pharmacother. 2019, 109, 1112–1119. [Google Scholar] [CrossRef]
- Chen, R.; Wu, J.C.; Liu, T.; Qu, Y.; Lu, L.G.; Xu, M.Y. MicroRNA profile analysis in the liver fibrotic tissues of chronic hepatitis B patients. J. Dig. Dis. 2017, 18, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Castoldi, G.; di Gioia, C.R.T.; Bombardi, C.; Catalucci, D.; Corradi, B.; Gualazzi, M.G.; Leopizzi, M.; Mancini, M.; Zerbini, G.; Condorelli, G.; et al. MiR-133a regulates collagen 1A1: Potential role of miR-133a in myocardial fibrosis in angiotensin II-dependent hypertension. J. Cell. Physiol. 2012, 227, 850–856. [Google Scholar] [CrossRef]
- Roderburg, C.; Luedde, M.; Cardenas, D.V.; Vucur, M.; Mollnow, T.; Zimmermann, H.W.; Koch, A.; Hellerbrand, C.; Weiskirchen, R.; Frey, N.; et al. miR-133a mediates TGF-β-dependent derepression of collagen synthesis in hepatic stellate cells during liver fibrosis. J. Hepatol. 2013, 58, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Tavallaee, G. Role of microRNA-27b-3p in the Regulation of Key Extracellular Matrix Markers in the Synovium During Osteoarthritis. Ph.D. Thesis, University of Toronto, Toronto, ON, Canada, 2020; p. 172. Available online: http://proxy.library.vcu.edu/login?url=https://www.proquest.com/dissertations-theses/role-microrna-27b-3p-regulation-key-extracellular/docview/2425107599/se-2?accountid=14780%0Ahttps://libkey.io/libraries/468/openurl?genre=dissertations&au=Tavallaee%2C+Gh (accessed on 31 August 2023).
- Rak, B.; Mehlich, D.; Garbicz, F.; Domosud, Z.; Paskal, W.; Marczewska, J.M.; Włodarski, P.K. Post-transcriptional Regulation of MMP16 and TIMP2 Expression via miR-382, miR-410 and miR-200b in Endometrial Cancer. Cancer Genom. Proteom. 2017, 14, 389–401. [Google Scholar] [CrossRef]
- Lin, C.A.; Duan, K.Y.; Wang, X.W.; Zhang, Z.S. Study on the role of Hsa-miR-382-5p in epidural fibrosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3663–3668. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, L.; Yu, Q.; Zhang, Y.; Yan, L.; Chen, Z.J. The estrogen-regulated lncRNA H19/miR-216a-5p axis alters stromal cell invasion and migration via ACTA2 in endometriosis. Mol. Hum. Reprod. 2019, 25, 550–561. [Google Scholar] [CrossRef]
- Rong, Y.; Zhang, J.; Jiang, D.; Ji, C.; Wang, J.; Ge, X.; Tang, P.; Yu, S.; Cui, W.; Cai, W. Hypoxic pretreatment of small extracellular vesicles mediates cartilage repair in osteoarthritis by delivering miR-216a-5p. Acta Biomater. 2021, 122, 325–342. [Google Scholar] [CrossRef]
- Genz, B.; Coleman, M.A.; Irvine, K.M.; Kutasovic, J.R.; Miranda, M.; Gratte, F.D.; Tirnitz-Parker, J.E.E.; Olynyk, J.K.; Calvopina, D.A.; Weis, A.; et al. Overexpression of miRNA-25-3p inhibits Notch1 signaling and TGF-β-induced collagen expression in hepatic stellate cells. Sci. Rep. 2019, 9, 8541. [Google Scholar] [CrossRef]
- He, X.; Deng, L. Potential of miR-25-3p in protection of chondrocytes: Emphasis on osteoarthritis. Folia Histochem. Cytobiol. 2021, 59, 30–39. [Google Scholar] [CrossRef]
- Zou, M.; Wang, F.; Gao, R.; Wu, J.; Ou, Y.; Chen, X.; Wang, T.; Zhou, X.; Zhu, W.; Li, P.; et al. Autophagy inhibition of hsa-miR-19a-3p/19b-3p by targeting TGF-β R II during TGF-β1-induced fibrogenesis in human cardiac fibroblasts. Sci. Rep. 2016, 6, 24747. [Google Scholar] [CrossRef]
- Duca, R.B.; Massillo, C.; Dalton, G.N.; Farré, P.L.; Graña, K.D.; Gardner, K.; De Siervi, A. MiR-19b-3p and miR-101-3p as potential biomarkers for prostate cancer diagnosis and prognosis. Am. J. Cancer Res. 2021, 11, 2802. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8263646/ (accessed on 27 April 2023). [PubMed]
- Nikulin, S.V.; Knyazev, E.N.; Gerasimenko, T.N.; Shilin, S.A.; Gazizov, I.N.; Zakharova, G.S.; Poloznikov, A.A.; Shkurnikov, M.Y. Non-Invasive Evaluation of Extracellular Matrix Formation in the Intestinal Epithelium. Bull. Exp. Biol. Med. 2018, 166, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.P.; Zhang, C.L.; Ma, X.L.; Hu, J.P.; Cai, T.; Zhang, L. Analyzing the Interactions of mRNAs and ncRNAs to Predict Competing Endogenous RNA Networks in Osteosarcoma Chemo-Resistance. Mol. Ther. 2019, 27, 518. [Google Scholar] [CrossRef]
- Luan, W.; Qian, Y.; Ni, X.; Bu, X.; Xia, Y.; Wang, J.; Ruan, H.; Ma, S.; Xu, B. miR-204-5p acts as a tumor suppressor by targeting matrix metalloproteinases-9 and B-cell lymphoma-2 in malignant melanoma. Onco. Targets. Ther. 2017, 10, 1237. [Google Scholar] [CrossRef]
- Paik, S.; Jung, H.S.; Lee, S.; Yoon, D.S.; Park, M.S.; Lee, J.W. miR-449a Regulates the Chondrogenesis of Human Mesenchymal Stem Cells Through Direct Targeting of Lymphoid Enhancer-Binding Factor-1. Stem Cells Dev. 2012, 21, 3298. [Google Scholar] [CrossRef]
- Martinez-Anton, A.; Sokolowska, M.; Kern, S.; Davis, A.S.; Alsaaty, S.; Taubenberger, J.K.; Sun, J.; Cai, R.; Danner, R.L.; Eberlein, M.; et al. Changes in microRNA and mRNA expression with differentiation of human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 2013, 49, 384–395. [Google Scholar] [CrossRef]
- Masè, M.; Grasso, M.; Avogaro, L.; Giacomaz, M.N.; D’Amato, E.; Tessarolo, F.; Graffigna, A.; Denti, M.A.; Ravelli, F. Upregulation of mir-133b and mir-328 in patients with atrial dilatation: Implications for stretch-induced atrial fibrillation. Front. Physiol. 2019, 10, 1133. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Dou, X.; Li, S.; Jia, Q.; Ling, P.; Liu, H.; Han, Q.; Sun, S. miR-590-5p affects chondrocyte proliferation, apoptosis, and inflammation by targeting FGF18 in osteoarthritis. Am. J. Transl. Res. 2021, 13, 8728. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8430182/ (accessed on 27 April 2023).
- Martínez-González, E.; Brochado-Kith, Ó.; Gómez-Sanz, A.; Martín-Carbonero, L.; Jimenez-Sousa, M.Á.; Martínez-Román, P.; Resino, S.; Briz, V.; Fernández-Rodríguez, A. Comparison of methods and characterization of small RNAs from plasma extracellular vesicles of HIV/HCV coinfected patients. Sci. Rep. 2020, 10, 11140. [Google Scholar] [CrossRef]
- Chen, Y.N.; Ren, C.C.; Yang, L.; Nai, M.M.; Xu, Y.M.; Zhang, F.; Liu, Y. MicroRNA let-7d-5p rescues ovarian cancer cell apoptosis and restores chemosensitivity by regulating the p53 signaling pathway via HMGA1. Int. J. Oncol. 2019, 54, 1771–1784. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Z.; Shao, Z.; Li, C.; Li, Y.; Liu, Q.; Zhang, Y.; Tan, B.; Liu, Y. Identifying multiple collagen gene family members as potential gastric cancer biomarkers using integrated bioinformatics analysis. PeerJ 2020, 8, e9123. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhu, H.-Y.; Bai, W.-D.; Su, L.-L.; Liu, J.-Q.; Cai, W.-X.; Zhao, B.; Gao, J.-X.; Han, S.-C.; Li, J.; et al. MiR-10a and miR-181c regulate collagen type I generation in hypertrophic scars by targeting PAI-1 and uPA. FEBS Lett. 2015, 589, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ye, Z.; Zhao, Z.; Li, L. Clinicopathologic significance of miR-10b expression in gastric carcinoma. Hum. Pathol. 2013, 44, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Cai, Y.; Rong, X.; Chen, J.; Zheng, D.; Chen, L.; Zhang, J. MiR-661 promotes tumor invasion and metastasis by directly inhibiting RB1 in non small cell lung cancer. Mol. Cancer 2017, 16, 122. [Google Scholar] [CrossRef]
- Wang, W.; Dong, R.; Guo, Y.; He, J.; Shao, C.; Yi, P.; Yu, F.; Gu, D.; Zheng, J. CircMTO1 inhibits liver fibrosis via regulation of miR-17-5p and Smad7. J. Cell. Mol. Med. 2019, 23, 5486. [Google Scholar] [CrossRef]
- Liu, E.; Lv, L.; Zhan, Y.; Ma, Y.; Feng, J.; He, Y.; Wen, Y.; Zhang, Y.; Pu, Q.; Ji, F.; et al. METTL3/N6-methyladenosine/ miR-21-5p promotes obstructive renal fibrosis by regulating inflammation through SPRY1/ERK/NF-κB pathway activation. J. Cell. Mol. Med. 2021, 25, 7660–7674. [Google Scholar] [CrossRef]
- Fukumoto, I.; Kikkawa, N.; Matsushita, R.; Kato, M.; Kurozumi, A.; Nishikawa, R.; Goto, Y.; Koshizuka, K.; Hanazawa, T.; Enokida, H.; et al. Tumor-suppressive microRNAs (miR-26a/b, miR-29a/b/c and miR-218) concertedly suppressed metastasis-promoting LOXL2 in head and neck squamous cell carcinoma. J. Hum. Genet. 2016, 61, 109–118. [Google Scholar] [CrossRef]
- Wu, L.; Wang, Q.; Guo, F.; Ma, X.; Wang, J.; Zhao, Y.; Yan, Y.; Qin, G. Involvement of miR-27a-3p in diabetic nephropathy via affecting renal fibrosis, mitochondrial dysfunction, and endoplasmic reticulum stress. J. Cell. Physiol. 2021, 236, 1454–1468. [Google Scholar] [CrossRef]
- Xu, Y.; Li, D.; Zhu, Z.; Li, L.; Jin, Y.; Ma, C.; Zhang, W. miR-27a-3p negatively regulates osteogenic differentiation of MC3T3-E1 preosteoblasts by targeting osterix. Mol. Med. Rep. 2020, 22, 1717–1726. [Google Scholar] [CrossRef]
- Yan, Y.; Du, C.; Duan, X.; Yao, X.; Wan, J.; Jiang, Z.; Qin, Z.; Li, W.; Pan, L.; Gu, Z.; et al. Inhibiting collagen I production and tumor cell colonization in the lung via miR-29a-3p loading of exosome-/liposome-based nanovesicles. Acta Pharm. Sin. B 2022, 12, 939. [Google Scholar] [CrossRef]
- Shen, H.; Wang, L.; Chen, Q.; Xu, J.; Zhang, J.; Fang, L.; Wang, J.; Fan, W. The prognostic value of COL3A1/FBN1/COL5A2/SPARC-mir-29a-3p-H19 associated ceRNA network in Gastric Cancer through bioinformatic exploration. J. Cancer 2020, 11, 4933. [Google Scholar] [CrossRef]
- Cao, B.; Dai, X. Platelet lysate induces chondrogenic differentiation of umbilical cord-derived mesenchymal stem cells by regulating the lncRNA H19/miR-29b-3p/SOX9 axis. FEBS Open Bio. 2020, 10, 2656. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Wang, X.; Zhou, F. Liver microRNA-29b-3p positively correlates with relative enhancement values of magnetic resonance imaging and represses liver fibrosis. J. Biochem. 2020, 168, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jin, J.; Tian, X.; Wu, L. hsa-miR-29c-3p regulates biological function of colorectal cancer by targeting SPARC. Oncotarget 2017, 8, 104508. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Jiang, T.; Hu, X.; Liu, Z.; Zhao, L.; Liu, H.; Liu, Z.; Ma, L. Downregulation of microRNA-30a in bronchoalveolar lavage fluid from idiopathic pulmonary fibrosis patients. Mol. Med. Rep. 2018, 18, 5799–5806. [Google Scholar] [CrossRef] [PubMed]
- Haghi, M.; Taha, M.F.; Javeri, A. Suppressive effect of exogenous miR-16 and miR-34a on tumorigenesis of breast cancer cells. J. Cell. Biochem. 2019, 120, 13342–13353. [Google Scholar] [CrossRef]
- Chou, K.Y.U.; Chang, A.N.C.; Tsai, T.E.F.U.; Lin, Y.I.C.; Chen, H.E.N.; Ho, C.Y.E.N.; Chen, P.O.C.; Hwang, T.I.S. MicroRNA-34a-5p serves as a tumor suppressor by regulating the cell motility of bladder cancer cells through matrix metalloproteinase-2 silencing. Oncol. Rep. 2021, 45, 911–920. [Google Scholar] [CrossRef]
- Mao, G.; Zhang, Z.; Huang, Z.; Chen, W.; Huang, G.; Meng, F.; Zhang, Z.; Kang, Y. MicroRNA-92a-3p regulates the expression of cartilage-speci fi c genes by directly targeting histone deacetylase 2 in chondrogenesis and degradation. Osteoarthr. Cartil. 2017, 25, 521–532. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, J.; Zhang, D.; Wang, L.; Zhao, W.; Lin, D.Y.T.; Chen, R.; Xie, H.; Hu, X.; Fang, X.; et al. MicroRNA expression profiles of scar and normal tissue from patients with posterior urethral stricture caused by pelvic fracture urethral distraction defects. Int. J. Mol. Med. 2018, 41, 2733–2743. [Google Scholar] [CrossRef]
- Zhou, X.; Luo, D.; Sun, H.; Qi, Y.; Xu, W.; Jin, X.; Li, C.; Lin, Z.; Li, G. MiR-132-3p regulates ADAMTS-5 expression and promotes chondrogenic differentiation of rat mesenchymal stem cells. J. Cell. Biochem. 2018, 119, 2579–2587. [Google Scholar] [CrossRef]
- Zhou, L.L.; Zhu, Y.M.; Qian, F.Y.; Yuan, C.C.; Yuan, D.P.; Zhou, X.P. MicroRNA-143-3p contributes to the regulation of pain responses in collagen-induced arthritis. Mol. Med. Rep. 2018, 18, 3219–3228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, X.; Tang, Y.; Chen, C.; Jing, R.; Liu, T. miR-155-5p Implicates in the Pathogenesis of Renal Fibrosis via Targeting SOCS1 and SOCS6. Oxidative Med. Cell. Longev. 2020, 2020, 6263921. [Google Scholar] [CrossRef]
- Chen, G.; Wang, D.; Zhao, X.; Cao, J.; Zhao, Y.; Wang, F.; Bai, J. miR-155-5p modulates malignant behaviors of hepatocellular carcinoma by directly targeting CTHRC1 and indirectly regulating GSK-3β-involved Wnt/β-catenin signaling. Cancer Cell Int. 2017, 17, 118. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Lu, L.; Liang, J.; Guo, X.; Zhang, P.H.; Luo, S. Keloid microRNA expression analysis and the influence of miR-199a-5p on the proliferation of keloid fibroblasts. Genet. Mol. Res. 2014, 13, 2727–2738. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gu, S.; Chen, B.F.; Shen, W.L.; Yin, Z.; Xu, G.W.; Hu, J.J.; Zhu, T.; Li, G.; Wan, C.; et al. Nanoparticle delivery of stable miR-199a-5p agomir improves the osteogenesis of human mesenchymal stem cells via the HIF1a pathway. Biomaterials 2015, 53, 239–250. [Google Scholar] [CrossRef]
- Wu, A.; Chen, Y.; Liu, Y.; Lai, Y.; Liu, D. miR-199b-5p inhibits triple negative breast cancer cell proliferation, migration and invasion by targeting DDR1. Oncol. Lett. 2018, 16, 4889–4896. [Google Scholar] [CrossRef]
- Winship, A.; Ton, A.; Van Sinderen, M.; Menkhorst, E.; Rainczuk, K.; Griffiths, M.; Cuman, C.; Dimitriadis, E. Mouse double minute homologue 2 (MDM2) downregulation by miR-661 impairs human endometrial epithelial cell adhesive capacity. Reprod. Fertil. Dev. 2018, 30, 477–486. [Google Scholar] [CrossRef]
- Sahin, Y.; Altan, Z.; Arman, K.; Bozgeyik, E.; Ozer, M.K.; Arslan, A. Inhibition of miR-664a interferes with the migration of osteosarcoma cells via modulation of MEG3. Biochem. Biophys. Res. Commun. 2017, 490, 1100–1105. [Google Scholar] [CrossRef]
- Ding, R.; Liu, X.; Zhang, J.; Yuan, J.; Zheng, S.; Cheng, X.; Jia, J. Downregulation of miR-1-3p expression inhibits the hypertrophy and mineralization of chondrocytes in DDH. J. Orthop. Surg. Res. 2021, 16, 512. [Google Scholar] [CrossRef]
- Guan, G.; Niu, X.; Qiao, X.; Wang, X.; Liu, J.; Zhong, M. Upregulation of Neural Cell Adhesion Molecule 1 (NCAM1) by hsa-miR-141-3p Suppresses Ameloblastoma Cell Migration. Med. Sci. Monit. 2020, 26, e923491-1–e923491-8. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, C.; Hou, L.; Wu, Y. Silencing of the lncRNA TUG1 attenuates the epithelial-mesenchymal transition of renal tubular epithelial cells by sponging miR-141-3p via regulating β-catenin. Am. J. Physiol. Renal Physiol. 2020, 319, F1125–F1134. [Google Scholar] [CrossRef] [PubMed]
- Makino, K.; Jinnin, M.; Kajihara, I.; Honda, N.; Sakai, K.; Masuguchi, S.; Fukushima, S.; Inoue, Y.; Ihn, H. Circulating miR-142-3p levels in patients with systemic sclerosis. Clin. Exp. Dermatology Exp. Dermatol. 2011, 37, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Börschel, C.S.; Stejskalova, A.; Schäfer, S.D.; Kiesel, L.; Götte, M. miR-142-3p Reduces the Size, Migration, and Contractility of Endometrial and Endometriotic Stromal Cells by Targeting Integrin- and Rho GTPase-Related Pathways That Regulate Cytoskeletal Function. Biomedicines 2020, 8, 291. [Google Scholar] [CrossRef] [PubMed]
- Sillence, D.O.; Rimoin, D.L. Classification of osteogenesis imperfect. Lancet 1978, 1, 1041–1042. [Google Scholar] [CrossRef] [PubMed]
- Kuivaniemi, H.; Tromp, G.; Prockop, D.J. Mutations in collagen genes: Causes of rare and some common diseases in humans. FASEB J. 1991, 5, 2052–2060. [Google Scholar] [CrossRef]
- Forlino, A.; Cabral, W.A.; Barnes, A.M.; Marini, J.C. New perspectives on osteogenesis imperfecta. Nat. Rev. Endocrinol. 2011, 7, 540–557. [Google Scholar] [CrossRef]
- Prockop, D.J.; Colige, A.; Helminen, H.; Khillan, J.S.; Pereira, R.; Vandenberg, P. Mutations in Type 1 Procollagen That Cause Osteogenesis Imperfecta: Effects of the Mutations on the Assembly of Collagen into Fibrils, the Basis of Phenotypic Variations, and Potential Antisense Therapies. J. Bone Miner. Res. 1993, 8, S489–S492. [Google Scholar] [CrossRef]
- Sun, K.; Wang, J.; Liu, F.; Ji, Z.; Guo, Z.; Zhang, C.; Yao, M. Ossotide promotes cell differentiation of human osteoblasts from osteogenesis imperfecta patients by up-regulating miR-145. Biomed. Pharmacother. 2016, 83, 1105–1110. [Google Scholar] [CrossRef]
- Adams, B.D.; Parsons, C.; Walker, L.; Zhang, W.C.; Slack, F.J. Targeting noncoding RNAs in disease. J. Clin. Invest. 2017, 127, 761–771. [Google Scholar] [CrossRef]
- Leimena, C.; Qiu, H. Non-Coding RNA in the Pathogenesis, Progression and Treatment of Hypertension. Int. J. Mol. Sci. 2018, 19, 927. [Google Scholar] [CrossRef]
- Fus-Kujawa, A.; Mendrek, B.; Bajdak-Rusinek, K.; Diak, N.; Strzelec, K.; Gutmajster, E.; Janelt, K.; Kowalczuk, A.; Trybus, A.; Rozwadowska, P.; et al. Gene-repaired iPS cells as novel approach for patient with Osteogenesis imperfecta. Front. Bioeng. Biotechnol. 2023, 11, 1205122. [Google Scholar] [CrossRef] [PubMed]
- Witecka, J.; Auguściak-Duma, A.M.; Kruczek, A.; Szydło, A.; Lesiak, M.; Krzak, M.; Pietrzyk, J.J.; Männikkö, M.; Sieroń, A.L. Two novel COL1A1 mutations in patients with osteogenesis imperfecta (OI) affect the stability of the collagen type I triple-helix. J. Appl. Genet. 2008, 49, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Majka, M.; Janeczko, M.; Goździk, J.; Jarocha, D.; Auguściak-Duma, A.; Witecka, J.; Lesiak, M.; Koryciak-Komarska, H.; Sieroń, A.L.; Józef, J.J. Cell therapy of a patient with type III osteogenesis imperfecta caused by mutation in COL1A2 gene and unstable collagen type I*. Open J. Genet. 2013, 3, 49–60. [Google Scholar] [CrossRef]

{kind=link}
| Patient No. | Age (Yrs) | Sex (F/M) | RefSNP (rs) Number or Mutation Position | Type of Mutation | Brief Clinical Description |
|---|---|---|---|---|---|
| OI donors harboring mutations in the COL1A1 gene | |||||
| 1 | 10 | M | rs1907512918 | Missense | About 10 bone fractures, osteoporosis (densitometry). |
| 2 | 0.5 | M | g.8874 G/T; c.662 G/T; p.Gly221Val; exon 9 * | Missense | Right femur fracture in early infancy, motor development delay, joint hypermobility, funnel chest, submucosal cleft palate, blue sclera, negative family history. |
| 3 | 8 | F | rs72645362 | Missense | About 11 bone fractures, normal hearing, positive family history. |
| 4 | 4 | F | g.20199 A/T, g.202000 delGACTGGTGAG; c.3881 A/T, c.3882-3891 del; p.Glu1294ValfsX32; exon 50 * | Missense and 10-nucleotide deletion (frameshift) | About 9 bone fractures, normal hearing, speech defect, normal bone density, lack of bone deformities, blue sclera, negative family history, no dentinogenesis. |
| 5 | 20 | F | g.2113 delG, frameshift; c.231 delG; p.Thr78Profs; exon 2 * | Single-nucleotide deletion (frameshift) | Blue sclera, negative family history. |
| 6 | 10 | F | rs72651634 | Single-nucleotide deletion (frameshift) | About 12 bone fractures, normal hearing. |
| 7 | 5 | M | rs72645370 | Single-nucleotide deletion (frameshift) | About 2 bone fractures per year (at the observation period), blue sclera, negative family history. |
| 8 | 29 | M | rs72653156 | Exon skipping | About 20 bone fractures, normal hearing, osteoporosis (densitometry). |
| OI donors with mutations in the COL1A2 gene | |||||
| 9 | 13 | M | g.26908 G/A; c.1828 G/A; p.Gly610Ser; exon 31 * | Missense | N.D. |
| 10 | 15 | M | rs72658185 | Missense | N.D. |
| miRNAs | Percentage of OI Donors with a Specific miRNA Detection | Target Gene(s) | Effect(s) | Location and/or Characteristics of miRNAsin ECM-Related Diseases | Ref. |
|---|---|---|---|---|---|
| PART A | miRNAs detected in OI but not in control fibroblasts | ||||
| hsa-miR-19a-3p | 70% | TGFβRII | Not found |
| [32] |
| hsa-miR-19b-3p | 70% | TGFβRII | Not found |
| [33,34] |
| hsa-miR-133a-3p | 30% | COL1A1 | Negative |
| [23,24] |
| hsa-miR-200b-3p | 30% | TIMP2 | Negative |
| [26,35] |
| hsa-miR-204-5p | 50% | MMP9 | Not found |
| [36] |
| hsa-miR-216a-5p | 50% | JAK2 | Negative |
| [28,29] |
| hsa-miR-377-3p | 30% | MMP2 and MMP16 | Negative |
| [26] |
| hsa-miR-449a | 70% | LEF1 | Not found |
| [37,38] |
| hsa-miR-590-5p | 30% | FGF18 | Negative |
| [39,40] |
| PART B | miRNAs detected in OI and control cells | ||||
| hsa-let-7a-5p | 100% | Not found | Not found |
| [41] |
| hsa-let-7d-5p | 100% | HMGA1 | Negative |
| [42,43] |
| hsa-miR-10a-5p | 90% | Not found | Not found |
| [44] |
| hsa-miR-10b-5p | 90% | MDM2 and MDM4 | Negative |
| [45,46] |
| hsa-miR-17-5p | 70% | circMTO1, Smad7 | Negative |
| [47] |
| hsa-miR-21-5p | 100% | SPRY1/ERK/NF-kB pathway | Not found |
| [48] |
| hsa-miR-25-3p | 90% | TGFBR1, ADAM17, FKBP14, NOTCH1 pathway | Negative |
| [30,31] |
| hsa-miR-26a-5p | 100% | LOXL2 | Negative |
| [49] |
| hsa-miR-26b-5p | 100% | LOXL2 | Negative |
| [49] |
| hsa-miR-27a-3p | 100% | TMBIM6 | Negative |
| [50,51] |
| hsa-miR-27b-3p | 90% | COL1A1, Adamts8, THBS1, FN1, COL5A1, and COL14A1 | Positive |
| [25] |
| hsa-miR-29a-3p | 100% | Not found | Not found |
| [52,53] |
| hsa-miR-29b-3p | 80% | STAT3 Track H19/miR-29b-30/SOX9 pathway | Negative on STAT3 |
| [54,55] |
| hsa-miR-29c-3p | 100% | SPARC | Negative |
| [56] |
| hsa-miR-30a-5p | 90% | TGF-β-activated kinase 1/MAP3K7-binding protein 3 (TAB3) | Negative on TAB3, α-SMA, and fibronectin expression |
| [57] |
| hsa-miR-34a-5p | 90% | MMP2 | Negative |
| [58,59] |
| hsa-miR-92a-3p | 100% | HDAC2 | Not found |
| [60] |
| hsa-miR-129-5p | 70% | Not found | Not found |
| [61] |
| hsa-miR-132-3p | 90% | ACAN, SOX9, COL2A1, ADAMTS5 | Positive |
| [62] |
| hsa-miR-143-3p | 80% | PTGS2, MRGPRE, PGD2, TNF | Negative |
| [63] |
| hsa-miR-155-5p | 100% | CTHRC1 and regulation of Wnt/β-catenin signaling pathway by GSK-3β | Positive |
| [64,65] |
| hsa-miR-199a-5p | 100% | Acting by inhibiting the HIF1a pathway | Not found |
| [66,67] |
| hsa-miR-199b-5p | 70% | Cadherin N | Negative on EMT |
| [68] |
| hsa-miR-382-5p | 100% | COL1A1 | Positive |
| [26,27] |
| hsa-miR-661 | 100% | MDM2, CDC34 | Negative |
| [69] |
| hsa-miR-664a-5p | 80% | Likely impact on MEG3 and SNORA36 | Negative |
| [70] |
| PART C | miRNAs not detected in OI nor control cells | ||||
| hsa-miR-1-3p | 0% | SOX9, indirect effect on type X collagen | Negative |
| [71] |
| hsa-miR-141-3p | 0% | TUG1, β-catenin | Not found |
| [72,73] |
| hsa-miR-142-3p | 0% | ROCK2, CFL2, RAC1, WASL | Negative |
| [74,75] |
| hsa-miR-200a-3p | 0% | Not found | Not found |
| [22] |
| hsa-miR-217 | 0% | PTEN, DKK1 | Negative on PTEN and DKK1; positive on RUNX2 and COL1A1 |
| [20,21] |
| miRNA Type | Fold Change in Expression in OI vs. Control Cells |
|---|---|
| miR-449a * | −20.9119 |
| miR-19b-3p * | −4.74695 |
| miR-19a-3p * | −4.10178 |
| miR-204-5p * | −3.71922 |
| miR-216a-5p * | −2.52801 |
| miR-10b-5p * | −2.38378 |
| miR-17-5p | −1.91123 |
| miR-382-5p | −1.87937 |
| miR-10a-5p | −1.67714 |
| miR-199a-5p | −1.66411 |
| miR-155-5p | −1.40347 |
| miR-92a-3p | −1.36391 |
| let-7d-5p | −1.34304 |
| let-7a-5p | −1.01942 |
| miR-661 | −1.00696 |
| miR-143-3p | 1.01308 |
| miR−132-3p | 1.23114 |
| miR-34a-5p | 1.47401 |
| miR-29a-3p | 1.52705 |
| miR-27a-3p | 1.54248 |
| miR-26b-5p | 1.58722 |
| miR-26a-5p | 1.68471 |
| miR-21-5p | 1.73538 |
| miR-29c-3p | 1.76541 |
| miR-29b-3p | 1.78077 |
| miR-25-3p | 1.90594 |
| miR-27b-3p | 1.93489 |
| miR-30a-5p * | 2.20228 |
| miR-199b-5p * | 2.89487 |
| miR-129-5p * | 3.64773 |
| miR-664a-5p * | 6.02516 |
| miR-10a-5p | miR-10b-5p | miR-17-5p | miR-199a-5p | miR-19a-3p | miR-19b-3p | miR-204-5p | miR-216a-5p | miR-382-5p | miR-449a | |
|---|---|---|---|---|---|---|---|---|---|---|
| miR-10a-5p | 1 | 0.60 | 0.20 | 0.50 | −0.16 | −0.02 | 0.26 | 0.09 | 0.60 | −0.14 |
| miR-10b-5p | 0.60 | 1.00 | 0.10 | 0.67 | 0.06 | −0.05 | 0.18 | −0.06 | 0.53 | −0.01 |
| miR-17-5p | 0.20 | 0.1 | 1.00 | 0.49 | 0.08 | 0.27 | 0.15 | 0.09 | 0.26 | 0.31 |
| miR-199a-5p | 0.49 | 0.67 | 0.49 | 1.00 | −0.02 | −0.20 | −0.11 | −0.19 | 0.77 | 0.11 |
| miR-19a-3p | −0.16 | 0.06 | 0.08 | −0.02 | 1.00 | 0.63 | 0.26 | 0.34 | −0.03 | 0.74 |
| miR-19b-3p | −0.02 | −0.05 | 0.27 | −0.20 | 0.63 | 1.00 | 0.66 | 0.57 | −0.27 | 0.75 |
| miR-204-5p | 0.26 | 0.18 | 0.15 | −0.11 | 0.26 | 0.66 | 1.00 | 0.43 | −0.11 | 0.47 |
| miR-216a-5p | 0.09 | −0.06 | 0.09 | −0.19 | 0.34 | 0.57 | 0.43 | 1.00 | −0.15 | 0.31 |
| miR-382-5p | 0.60 | 0.53 | 0.26 | 0.77 | −0.03 | −0.27 | −0.11 | −0.15 | 1.00 | 0.02 |
| miR-449a | −0.14 | −0.01 | 0.31 | 0.11 | 0.74 | 0.75 | 0.47 | 0.31 | 0.02 | 1.00 |
| miR-199b-5p | miR-21-5p | miR-25-3p | miR-26a-5p | miR-26b-5p | miR-27a-3p | miR-27b-3p | miR-29a-3p | miR-29b-3p | miR-29c-3p | miR-30a-5p | miR-664a-5p | miR-129-5p | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| miR-199b-5p | 1.00 | 0.19 | 0.26 | 0.31 | 0.18 | 0.34 | 0.32 | 0.26 | 0.95 | 0.27 | 0.48 | 0.64 | 0.49 |
| miR-21-5p | 0.19 | 1.00 | 0.92 | 0.93 | 0.92 | 0.91 | 0.92 | 0.95 | 0.14 | 0.94 | 0.69 | 0.34 | −0.42 |
| miR-25-3p | 0.26 | 0.92 | 1.00 | 0.93 | 0.92 | 0.92 | 0.91 | 0.92 | 0.20 | 0.91 | 0.79 | 0.48 | −0.31 |
| miR-26a-5p | 0.31 | 0.93 | 0.93 | 1.00 | 0.95 | 0.94 | 0.94 | 0.97 | 0.23 | 0.96 | 0.81 | 0.42 | −0.38 |
| miR-26b-5p | 0.18 | 0.92 | 0.92 | 0.95 | 1.00 | 0.91 | 0.91 | 0.96 | 0.09 | 0.96 | 0.78 | 0.32 | −0.48 |
| miR-27a-3p | 0.34 | 0.91 | 0.92 | 0.94 | 0.91 | 1.00 | 0.90 | 0.92 | 0.27 | 0.90 | 0.86 | 0.53 | −0.27 |
| miR-27b-3p | 0.32 | 0.92 | 0.91 | 0.94 | 0.91 | 0.90 | 1.00 | 0.93 | 0.23 | 0.92 | 0.74 | 0.44 | −0.36 |
| miR-29a-3p | 0.26 | 0.95 | 0.92 | 0.97 | 0.96 | 0.92 | 0.93 | 1.00 | 0.16 | 0.99 | 0.76 | 0.35 | −0.50 |
| miR-29b-3p | 0.95 | 0.14 | 0.20 | 0.23 | 0.09 | 0.27 | 0.23 | 0.16 | 1.00 | 0.18 | 0.41 | 0.61 | 0.60 |
| miR-29c-3p | 0.27 | 0.94 | 0.91 | 0.96 | 0.96 | 0.90 | 0.92 | 0.99 | 0.18 | 1.00 | 0.73 | 0.31 | −0.49 |
| miR-30a-5p | 0.48 | 0.69 | 0.79 | 0.81 | 0.78 | 0.86 | 0.74 | 0.76 | 0.41 | 0.73 | 1.00 | 0.65 | −0.08 |
| miR-664a-5p | 0.64 | 0.34 | 0.48 | 0.42 | 0.32 | 0.53 | 0.44 | 0.35 | 0.61 | 0.31 | 0.65 | 1.00 | 0.41 |
| miR-129-5p | 0.49 | −0.42 | −0.31 | −0.38 | −0.48 | −0.27 | −0.36 | −0.50 | 0.60 | −0.49 | −0.08 | 0.41 | 1.00 |
| Patient Code | Downregulated miRNAs | Upregulated miRNAs |
|---|---|---|
| 1 | miR-10a-5p, miR-10b-5p, miR-19a-3p, miR-26b-5p, miR-34a-5p, miR-199a-5p, miR-382-5p | miR-17-5p, miR-29b-3p, miR-129-5p, miR-143-3p, miR-155-5p, miR-199b-5p, miR-664a |
| 2 | let-7a-5p, let-7d-5p, miR-10b-5p, miR-30a-5p, miR-155-5p, miR-199a-5p, miR-382-5p, miR-449a, miR-661 | miR-29b-3p, miR-34a-5p, miR-129-5p, miR-132-3p, miR-199b-5p, miR-664a-5p |
| 3 | let-7a-5p, miR-10a-5p, miR-10b-5p, miR-21-5p, miR-26b-5p, miR-27a-3p, miR-29a-3p, miR-29c-3p, miR-92a-3p, miR-382-5p | miR-17-5p, miR-29b-3p, miR-129-5p, miR-143-3p, miR-199b-5p, miR-664a-5p, miR-661 |
| 4 | miR-10a-5p, miR-10b-5p, miR-17-5p, miR-19a-3p, miR-19b-3p, miR-143-3p, miR-199a-5p, miR-449a, miR-661 | miR-26a-5p, miR-26b-5p, miR-29a-3p, miR-29c-3p |
| 5 | miR-17-5p, miR-19b-3p | let-7a-5p, let-7d-5p, miR-21-5p, miR-25-3p, miR-26a-5p, miR-26b-5p, miR-27a-3p, miR-27b-3p, miR-29a-3p, miR-29c-3p, miR-30a-5p, miR-34a-5p, miR-92a-3p, miR-129-5p, miR-132-3p, miR-155-5p, miR-199a-5p, miR-199b-5p, miR-382-5p, miR-661, miR-664a-5p |
| 6 | let-7d-5p, miR-10b-5p, miR-21-5p, miR-26b-5p, miR-27b-3p, miR-29a-3p, miR-34a-5p, miR-92a-3p, miR-132-3p, miR-143-3p, miR-199a-5p, miR-449a | miR-17-5p, miR-29b-3p, miR-30a-5p, miR-129-5p, miR-199b-5p, miR-661, miR-664a-5p |
| 7 | miR-10b-5p, miR-19a-3p, miR-19b-3p, miR-199a-5p, miR-449a | miR-10a-5p, miR-21-5p, miR-26a-5p, miR-26b-5p, miR-27a-3p, miR-27b-3p, miR-29a-3p, miR-29c-3p, miR-30a-5p, miR-664a-5p |
| 8 | miR-10b-5p, miR-17-5p, miR-19a-3p, miR-19b-3p, miR-129-5p, miR-155-5p, miR-199a-5p, miR-449a, miR-661 | let-7a-5p, miR-21-5p, miR-25-3p, miR-26a-5p, miR-26b-5p, miR-27a-3p, miR-27b-3p, miR-29a-3p, miR-29c-3p, miR-30a-5p, miR-34a-5p, miR-664a-5p |
| 9 | let-7a-5p, let-7d-5p, miR-10a-5p, miR-10b-5p, miR-19a-3p, miR-19b-3p, miR-132-3p, miR-129-5p, miR-155-5p, miR-199a-5p, miR-382-5p, miR-449a, miR-661 | miR-21-5p, miR-26b-5p, miR-27a-3p, miR-27b-3p, miR-29a-3p, miR-29c-3p, miR-30a-5p, miR-664a-5p |
| 10 | miR-19a-3p, miR-19b-3p, miR-25-3p, miR-382-5p, miR-449a | miR-27a-3p, miR-29b-3p, miR-30a-5p, miR-34a-5p, miR-129-5p, miR-132-3p, miR-199b-5p, miR-664a-5p |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botor, M.; Auguściak-Duma, A.; Lesiak, M.; Sieroń, Ł.; Dziedzic-Kowalska, A.; Witecka, J.; Asman, M.; Madetko-Talowska, A.; Bik-Multanowski, M.; Galicka, A.; et al. Analysis of miRNAs in Osteogenesis imperfecta Caused by Mutations in COL1A1 and COL1A2: Insights into Molecular Mechanisms and Potential Therapeutic Targets. Pharmaceuticals 2023, 16, 1414. https://doi.org/10.3390/ph16101414
Botor M, Auguściak-Duma A, Lesiak M, Sieroń Ł, Dziedzic-Kowalska A, Witecka J, Asman M, Madetko-Talowska A, Bik-Multanowski M, Galicka A, et al. Analysis of miRNAs in Osteogenesis imperfecta Caused by Mutations in COL1A1 and COL1A2: Insights into Molecular Mechanisms and Potential Therapeutic Targets. Pharmaceuticals. 2023; 16(10):1414. https://doi.org/10.3390/ph16101414
Chicago/Turabian StyleBotor, Malwina, Aleksandra Auguściak-Duma, Marta Lesiak, Łukasz Sieroń, Agata Dziedzic-Kowalska, Joanna Witecka, Marek Asman, Anna Madetko-Talowska, Mirosław Bik-Multanowski, Anna Galicka, and et al. 2023. "Analysis of miRNAs in Osteogenesis imperfecta Caused by Mutations in COL1A1 and COL1A2: Insights into Molecular Mechanisms and Potential Therapeutic Targets" Pharmaceuticals 16, no. 10: 1414. https://doi.org/10.3390/ph16101414
APA StyleBotor, M., Auguściak-Duma, A., Lesiak, M., Sieroń, Ł., Dziedzic-Kowalska, A., Witecka, J., Asman, M., Madetko-Talowska, A., Bik-Multanowski, M., Galicka, A., Sieroń, A. L., & Gawron, K. (2023). Analysis of miRNAs in Osteogenesis imperfecta Caused by Mutations in COL1A1 and COL1A2: Insights into Molecular Mechanisms and Potential Therapeutic Targets. Pharmaceuticals, 16(10), 1414. https://doi.org/10.3390/ph16101414