
Ginsenoside Rh3 Inhibits Lung Cancer Metastasis by Targeting Extracellular Signal-Regulated Kinase: A Network Pharmacology Study

Abstract
:1. Introduction
2. Results
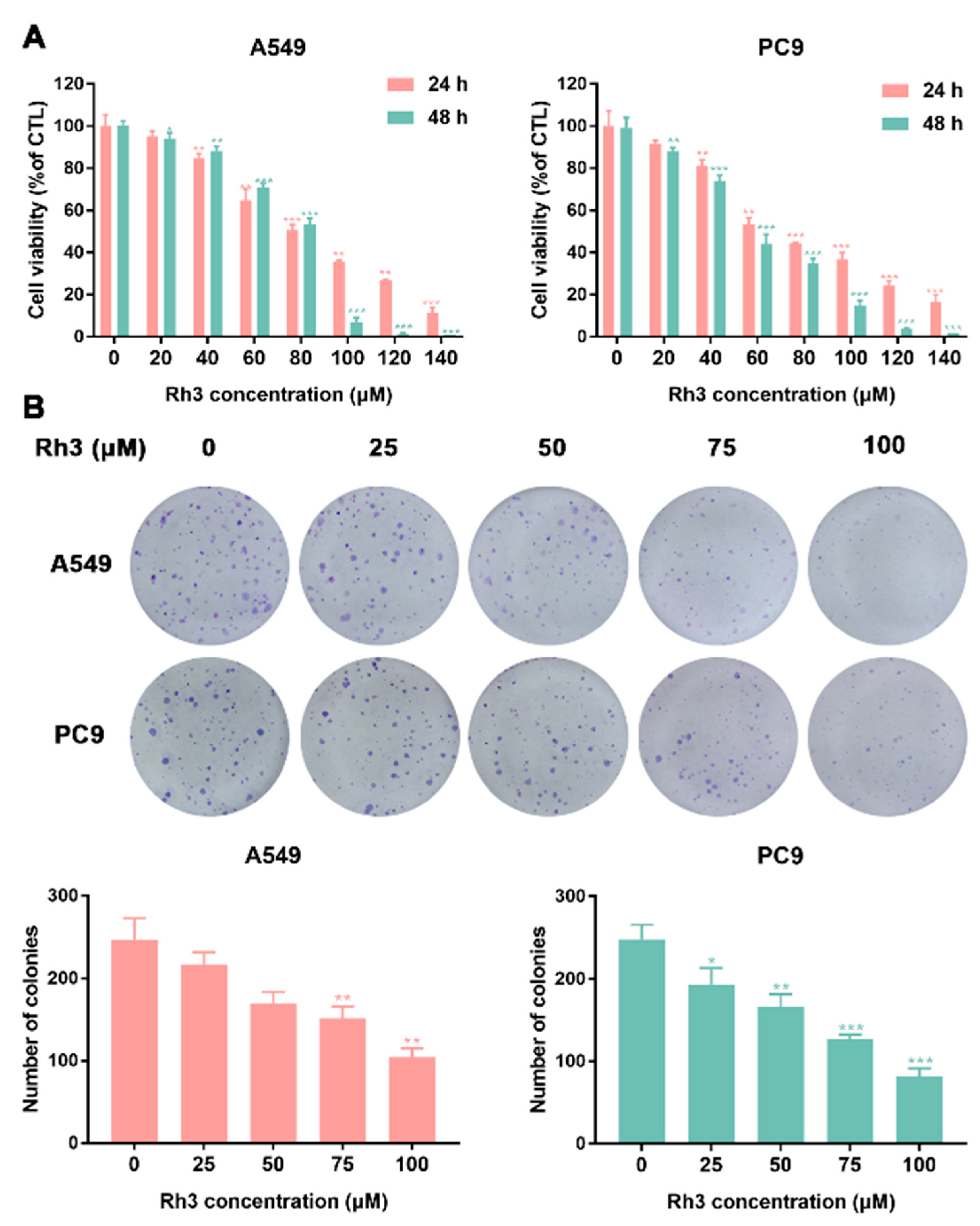
2.1. Inhibition of Lung Cancer Growth by Ginsenoside Rh3 In Vitro
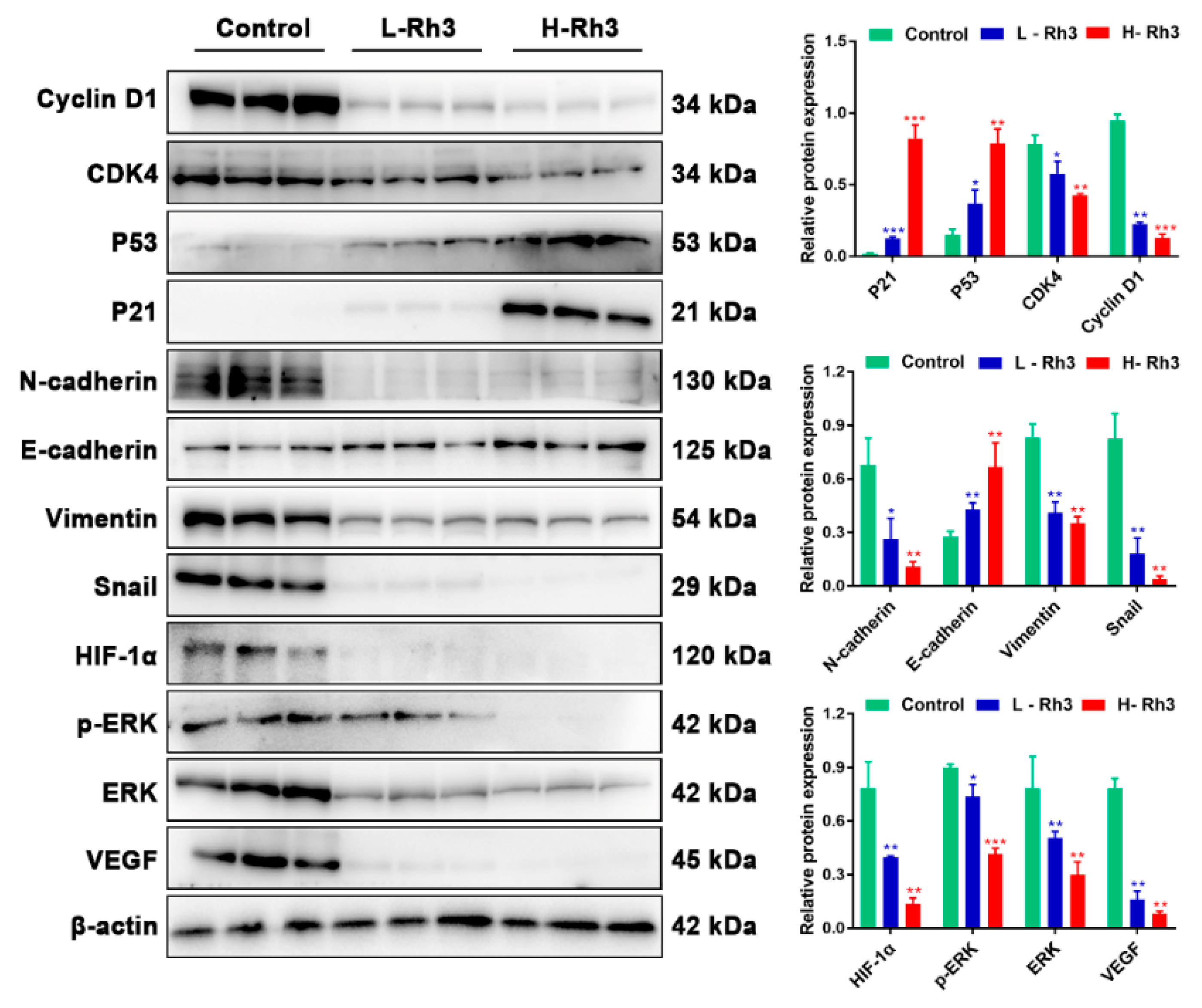
2.2. Ginsenoside Rh3 Triggers G1 Phase Arrest In Vitro
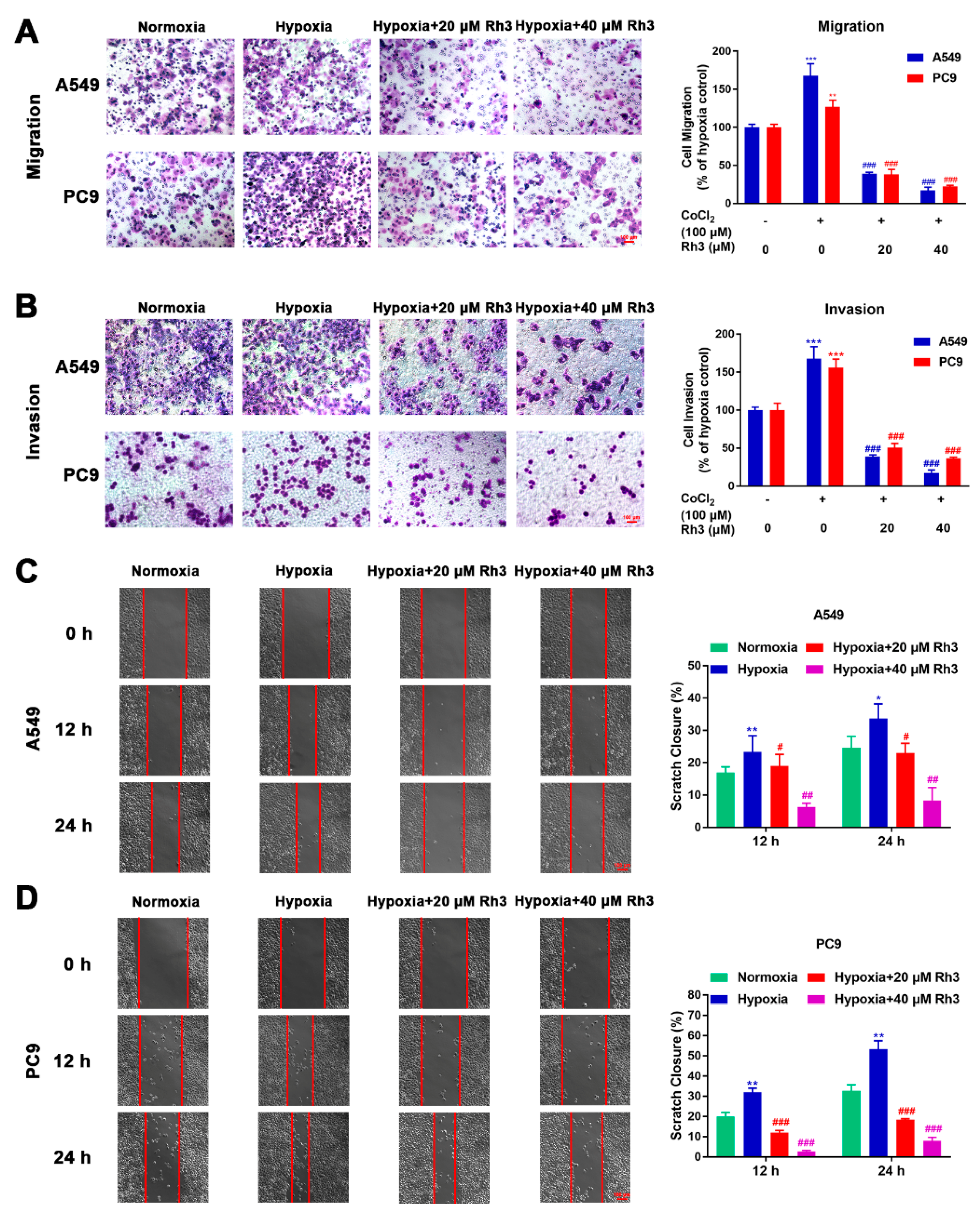
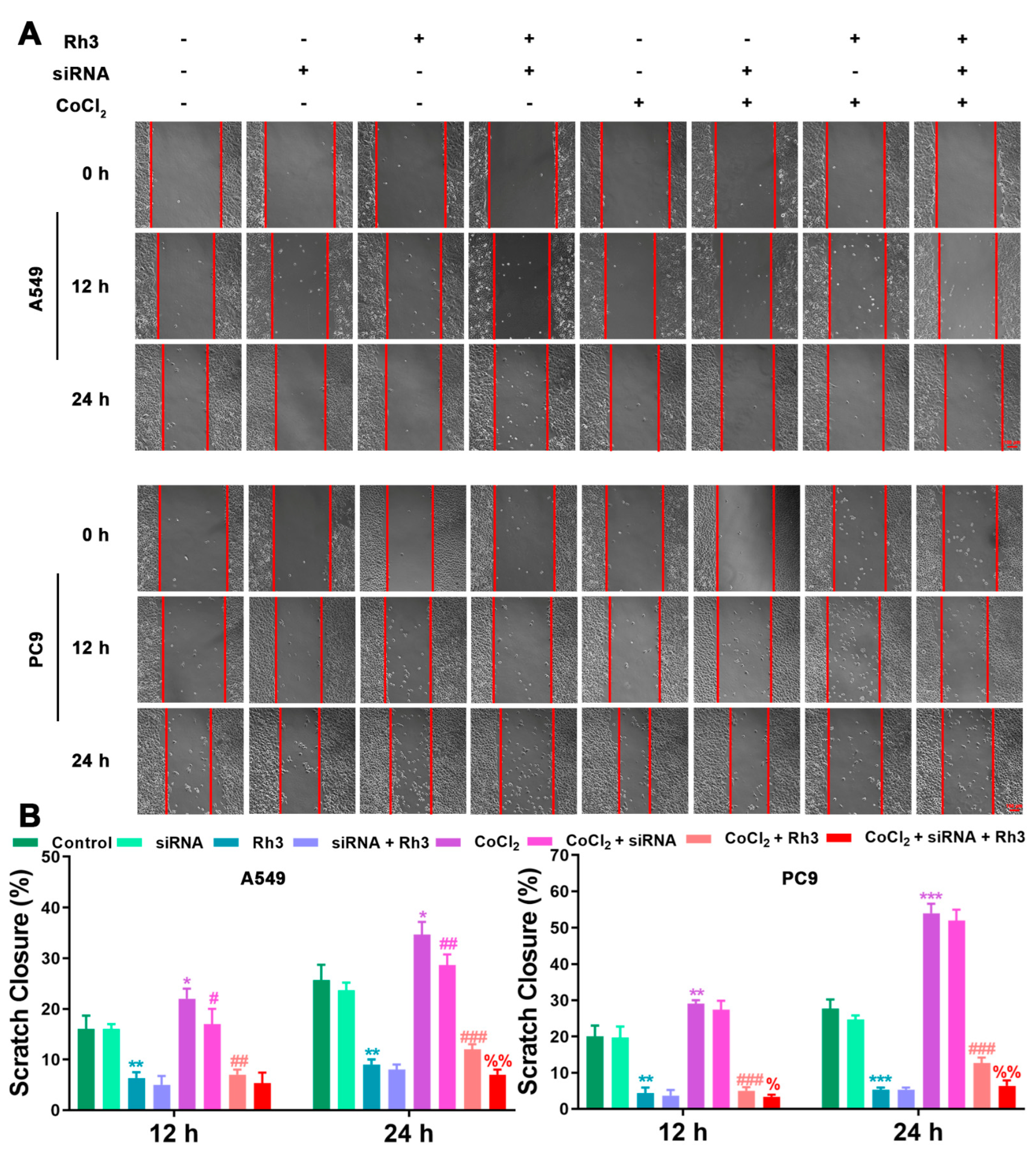
2.3. Ginsenoside Rh3 Inhibits Hypoxia-Induced Migration and Invasion in Lung Cancer Cells
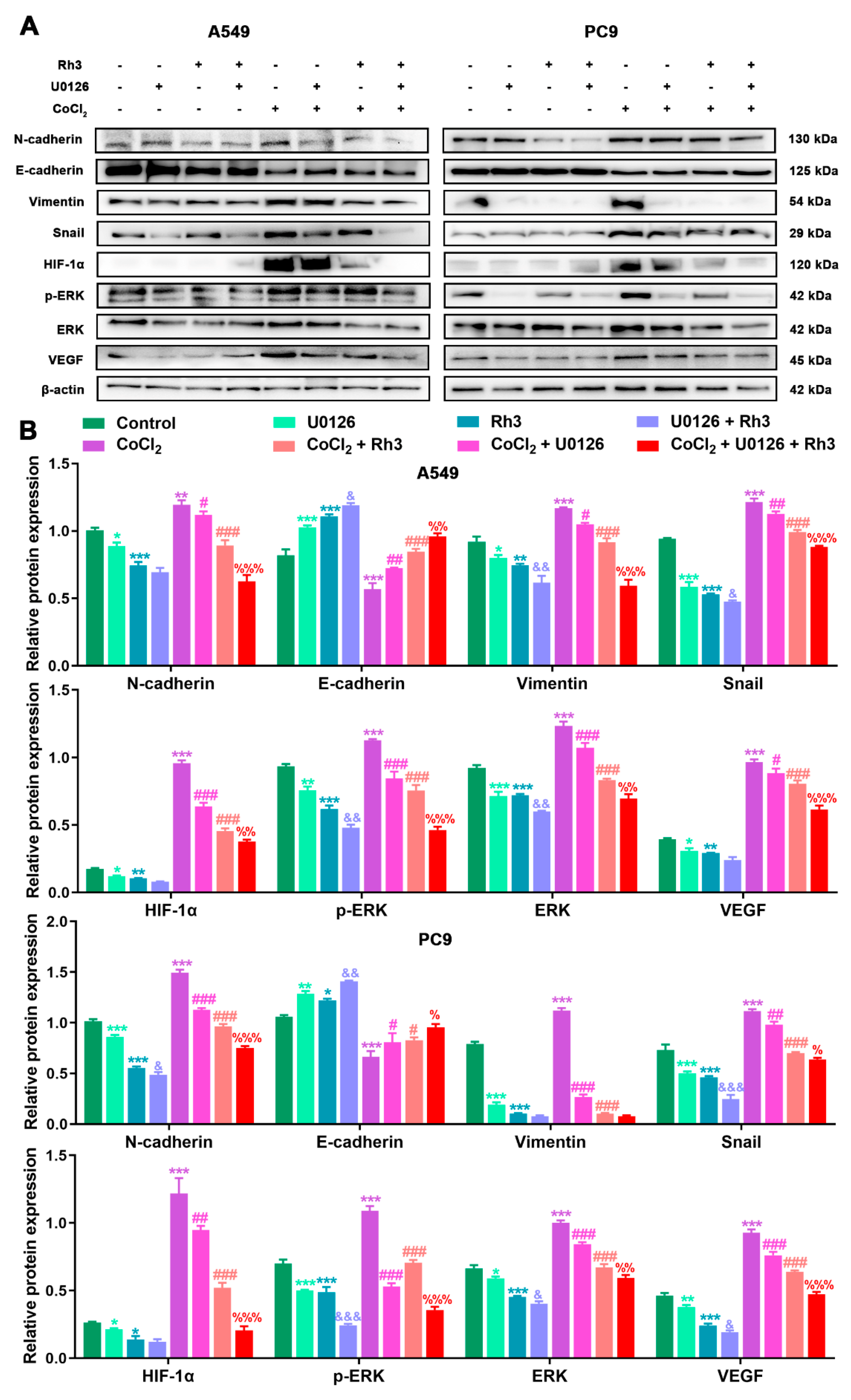
2.4. Regulation of Lung Cancer Metastasis-Associated Proteins by Rh3
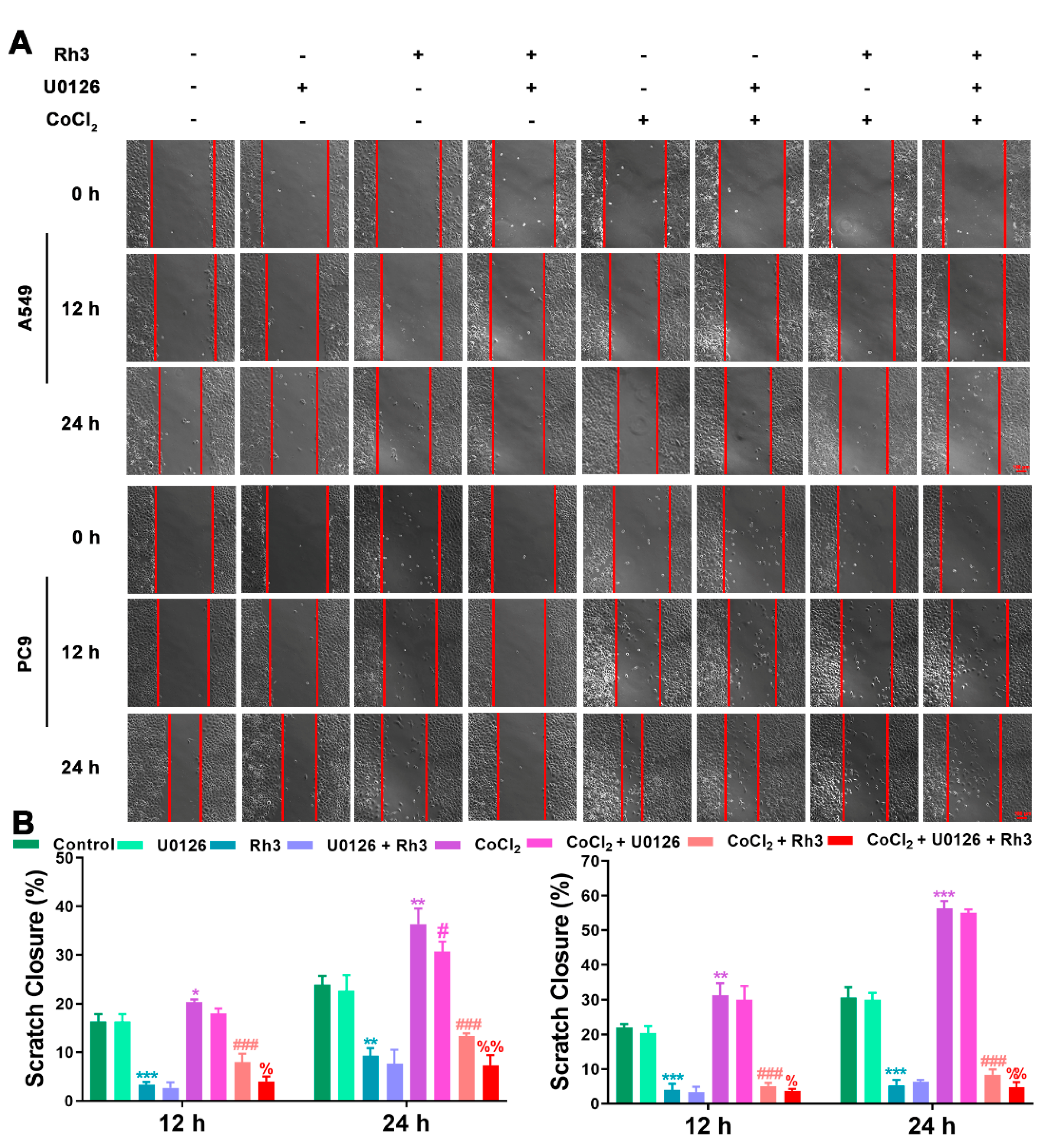
2.5. Ginsenoside Rh3 Targeting ERK Inhibits Metastasis Capability in Lung Cancer Cells
2.6. Rh3 Inhibits Metastasis of Lung Cancer and Triggers G1 Phase Arrest In Vivo
2.7. Ginsenoside Rh3 induces Low Toxicity In Vivo
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. In Vitro Cytotoxicity Assay
4.3. Cell Migration and Invasion Assays
4.4. Flow Cytometry Analysis
4.5. Western Blotting
4.6. RNA Isolation and RT-qPCR
4.7. siRNA Transfection Analysis
4.8. Analysis of Metastasis in Nude Mice
4.9. Network Construction and Molecular Docking
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Inamura, K. Lung cancer: Understanding its molecular pathology and the 2015 WHO classification. Front. Oncol. 2017, 7, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.-B.; He, X.; Huang, Y.-F.; Wu, Q.-N.; Zhou, Y.-C.; Hao, D.-J. Long Noncoding RNA CRNDE Promotes Multiple Myeloma Cell Growth by Suppressing miR-451. Oncol. Res. 2017, 25, 1207–1214. [Google Scholar] [CrossRef]
- Tang, C.; Liao, Z.; Hess, K.; Chance, W.W.; Zhuang, Y.; Jensen, G.; Xu, T.; Komaki, R.; Gomez, D.R. Prognosis and predictors of site of first metastasis after definitive radiation therapy for non-small cell lung cancer. Acta Oncol. 2016, 55, 1022–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, T.; Ibaragi, S.; Hu, G.-F. Epithelial-Mesenchymal Transition and Cell Cooperativity in Metastasis. Cancer Res. 2009, 69, 7135–7139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sipos, F. Epithelial-to-mesenchymal and mesenchymal-to-epithelial transitions in the colon. World J. Gastroenterol. 2012, 18, 601–608. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, P.; Huang, H.; Ye, X.; Chen, W.; Xu, G.; Zhang, F. Androgen receptor regulates eIF5A2 expression and promotes prostate cancer metastasis via EMT. Cell Death Discov. 2021, 7, 373. [Google Scholar] [CrossRef]
- de Araújo, R.S.A.; Carmo, J.D.O.D.S.; Silva, S.L.d.O.; da Silva, C.R.A.C.; Souza, T.P.M.; de Mélo, N.B.; Bourguignon, J.-J.; Schmitt, M.; de Aquino, T.M.; Rodarte, R.S.; et al. Coumarin Derivatives Exert Anti-Lung Cancer Activity by Inhibition of Epithelial–Mesenchymal Transition and Migration in A549 Cells. Pharmaceuticals 2022, 15, 104. [Google Scholar] [CrossRef]
- Umezu, T.; Tadokoro, H.; Azuma, K.; Yoshizawa, S.; Ohyashiki, K.; Ohyashiki, J.H. Exosomal miR-135b shed from hypoxic multiple myeloma cells enhances angiogenesis by targeting factor-inhibiting HIF-1. Blood 2014, 124, 3748–3757. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Ahn, G.-O.; Seita, J.; Hong, B.-J.; Kim, Y.-E.; Bok, S.; Lee, C.-J.; Kim, K.S.; Lee, J.C.; Leeper, N.J.; Cooke, J.P.; et al. Transcriptional activation of hypoxia-inducible factor-1 (HIF-1) in myeloid cells promotes angiogenesis through VEGF and S100A8. Proc. Natl. Acad. Sci. USA 2014, 111, 2698–2703. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Li, J.; Peng, Y.; Shen, B.; Li, Y.; Liu, L.; Wang, C.; Xu, Y.; Lin, S.; Zhang, S.; et al. Ginsenoside Rb1 attenuates methamphetamine (METH)-induced neurotoxicity through the NR2B/ERK/CREB/BDNF signalings In Vitro and In Vivo models. J. Ginseng Res. 2022, 46, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Sulzmaier, F.J.; Ramos, J.W. RSK Isoforms in Cancer Cell Invasion and Metastasis. Cancer Res. 2013, 73, 6099–6105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.-J.; Diao, L.-T.; Cai, N.; Zhang, L.-T.; Xiang, S.; Jia, C.-C.; Qiu, D.-B.; Liu, C.; Sun, Y.-J.; Lei, H.; et al. mascRNA and its parent lncRNA MALAT1 promote proliferation and metastasis of hepatocellular carcinoma cells by activating ERK/MAPK signaling pathway. Cell Death Discov. 2021, 7, 110. [Google Scholar] [CrossRef]
- Savoia, P.; Fava, P.; Casoni, F.; Cremona, O. Targeting the ERK Signaling Pathway in Melanoma. Int. J. Mol. Sci. 2019, 20, 1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonis, I.; Chachami, G.; Samiotaki, M.; Panayotou, G.; Paraskeva, E.; Kalousi, A.; Georgatsou, E.; Bonanou, S.; Simos, G. Identification of MAPK Phosphorylation Sites and Their Role in the Localization and Activity of Hypoxia-inducible Factor-1α. J. Biol. Chem. 2006, 281, 33095–33106. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Moreno, O.; Lecanda, J.; Green, J.E.; Segura, V.; Catena, R.; Serrano, D.; Calvo, A. VEGF elicits epithelial-mesenchymal transition (EMT) in prostate intraepithelial neoplasia (PIN)-like cells via an autocrine loop. Exp. Cell Res. 2010, 316, 554–567. [Google Scholar] [CrossRef]
- Luo, M.; Hou, L.; Li, J.; Shao, S.; Huang, S.; Meng, D.; Liu, L.; Feng, L.; Xia, P.; Qin, T.; et al. VEGF/NRP-1 axis promotes progression of breast cancer via enhancement of epithelial-mesenchymal transition and activation of NF-κB and β-catenin. Cancer Lett. 2016, 373, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Gao, Y.; Liu, Q.; Wan, L.; Liu, H.; Bian, W.; Du, Y.; Huang, C. Zoledronic acid re-sensitises gefitinib-resistant lung cancer cells by inhibiting the JAK/STAT3 signalling pathway and reversing epithelial-mesenchymal transition. Oncol. Rep. 2020, 45, 459–468. [Google Scholar] [CrossRef]
- Huang, L.; Fu, L. Mechanisms of resistance to EGFR tyrosine kinase inhibitors. Acta Pharm. Sin. B 2015, 5, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-C.S.H.; Ko, P.-J.; Pan, Y.-S.; Lin, H.-Y.; Whang-Peng, J.; Davis, P.J.; Wang, K. Role of thyroid hormone-integrin αvβ3-signal and therapeutic strategies in colorectal cancers. J. Biomed. Sci. 2021, 28, 24. [Google Scholar] [CrossRef]
- Zhu, N.; Hou, J.; Ma, G.; Liu, J. Network Pharmacology Identifies the Mechanisms of Action of Shaoyao Gancao Decoction in the Treatment of Osteoarthritis. Med. Sci. Monit. 2019, 25, 6051–6073. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, J.; Zhang, D.; Wang, K.; Duan, X.; Meng, Z.; Zhang, X. Network Pharmacology-Based Approach to Investigate the Mechanisms of Hedyotis diffusa Willd. in the Treatment of Gastric Cancer. Evid. Based Complement. Altern. Med. 2018, 2018, 7802639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Zhao, K. Herb–Drug Interactions of Commonly Used Chinese Medicinal Herbs. Int. Rev. Neurobiol. 2017, 135, 197–232. [Google Scholar] [CrossRef] [PubMed]
- Rengarajan, T.; Nandakumar, N.; Rajendran, P.; Haribabu, L.; Nishigaki, I.; Balasubramanian, M.P. D-Pinitol Promotes Apoptosis in MCF-7 Cells via Induction of p53 and Bax and Inhibition of Bcl-2 and NF-κB. Asian Pac. J. Cancer Prev. 2014, 15, 1757–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Li, H.T.; Wei, S.Z.; Cai, H.D.; Zhu, Y.; Liu, H.H.; Li, Y.Z.; Wang, J.P.; Zou, W.J.; Zhao, Y.L. Use of Network Pharmacology and Molecular Docking to Investigate the Mechanism by Which Ginseng Ameliorates Hypoxia. Biomed. Environ. Sci. 2018, 31, 855–858. [Google Scholar] [PubMed]
- Panossian, A.G.; Efferth, T.; Shikov, A.N.; Pozharitskaya, O.N.; Kuchta, K.; Mukherjee, P.K.; Banerjee, S.; Heinrich, M.; Wu, W.; Guo, D.; et al. Evolution of the adaptogenic concept from traditional use to medical systems: Pharmacology of stress- and aging-related diseases. Med. Res. Rev. 2021, 41, 630–703. [Google Scholar] [CrossRef] [PubMed]
- Shikov, A.N.; Narkevich, I.A.; Flisyuk, E.V.; Luzhanin, V.G.; Pozharitskaya, O.N. Medicinal plants from the 14th edition of the Russian Pharmacopoeia, recent updates. J. Ethnopharmacol. 2021, 268, 113685. [Google Scholar] [CrossRef]
- Yi, Y.-S. Roles of ginsenosides in inflammasome activation. J. Ginseng Res. 2019, 43, 172–178. [Google Scholar] [CrossRef]
- Li, B.; Zhao, J.; Wang, C.-Z.; Searle, J.; He, T.-C.; Yuan, C.-S.; Du, W. Ginsenoside Rh2 induces apoptosis and paraptosis-like cell death in colorectal cancer cells through activation of p53. Cancer Lett. 2011, 301, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Yayeh, T.; Jung, K.-H.; Jeong, H.-Y.; Park, J.-H.; Song, Y.-B.; Kwak, Y.-S.; Kang, H.-S.; Cho, J.-Y.; Oh, J.-W.; Kim, S.-K.; et al. Korean Red Ginseng Saponin Fraction Downregulates Proinflammatory Mediators in LPS Stimulated RAW264.7 Cells and Protects Mice against Endotoxic Shock. J. Ginseng Res. 2012, 36, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.S.; Ham, J.; Kim, Y.-J.; Park, J.H.; Cho, E.-J.; Yamabe, N. Heat-processed Panax ginseng and diabetic renal damage: Active components and action mechanism. J. Ginseng Res. 2013, 37, 379–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, Y.; Zhang, Y.-L.; Ma, C.-J.; Li, M.-Q.; Tang, C.-Y.; Yang, Y.-F.; Zeng, J.-H.; Huang, X.-Y.; Yi, J.; Wang, X.-M.; et al. Neuroprotective Effects of Ginsenoside Rg1 against Hyperphosphorylated Tau-Induced Diabetic Retinal Neurodegeneration via Activation of IRS-1/Akt/GSK3β Signaling. J. Agric. Food Chem. 2019, 67, 8348–8360. [Google Scholar] [CrossRef] [PubMed]
- Coon, J.T.; Ernst, E. Panax ginseng: A systematic review of adverse effects and drug interactions. Drug Saf. 2002, 25, 323–344. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.-W.; Bae, E.-A.; Kim, D.-H. Inhibitory effect of ginsenoside Rg5 and its metabolite ginsenoside Rh3 in an oxazolone-induced mouse chronic dermatitis model. Arch. Pharmacal. Res. 2006, 29, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.; Lee, J.; Park, Y.S.; Lim, Y.; Chang, D.H.; Park, J.; Hwang, J.S. Inhibitory mechanism of Korean Red Ginseng on GM-CSF expression in UVB-irradiated keratinocytes. J. Ginseng Res. 2015, 39, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.-Z.; Li, K.-R.; Yu, Q.; Jiang, Q.; Yao, J.; Cao, C. Activation of Nrf2 by Ginsenoside Rh3 protects retinal pigment epithelium cells and retinal ganglion cells from UV. Free Radic. Biol. Med. 2018, 117, 238–246. [Google Scholar] [CrossRef]
- Lee, H.L.; Kang, K.S. Protective effect of ginsenoside Rh3 against anticancer drug-induced apoptosis in LLC-PK1 kidney cells. J. Ginseng Res. 2017, 41, 227–231. [Google Scholar] [CrossRef]
- Fukuoka, S.-I.; Nyaruhucha, C.M. Expression and functional analysis of rat P23, a gut hormone-inducible isoform of trypsin, reveals its resistance to proteinaceous trypsin inhibitors. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2002, 1588, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Ando, N.; Kato, H.; Igaki, H.; Shinoda, M.; Ozawa, S.; Shimizu, H.; Nakamura, T.; Yabusaki, H.; Aoyama, N.; Kurita, A.; et al. A Randomized Trial Comparing Postoperative Adjuvant Chemotherapy with Cisplatin and 5-Fluorouracil Versus Preoperative Chemotherapy for Localized Advanced Squamous Cell Carcinoma of the Thoracic Esophagus (JCOG9907). Ann. Surg. Oncol. 2012, 19, 68–74. [Google Scholar] [CrossRef]
- Fu, H.; Zhu, Y.; Wang, Y.; Liu, Z.; Zhang, J.; Xie, H.; Fu, Q.; Dai, B.; Ye, D.; Xu, J. Identification and Validation of Stromal Immunotype Predict Survival and Benefit from Adjuvant Chemotherapy in Patients with Muscle-Invasive Bladder Cancer. Clin. Cancer Res. 2018, 24, 3069–3078. [Google Scholar] [CrossRef] [Green Version]
- Nigg, E. Cyclin-dependent protein kinases: Key regulators of the eukaryotic cell cycle. BioEssays 1995, 17, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-H.; Wu, M.-Z.; Chiou, S.-H.; Chen, P.-M.; Chang, S.-Y.; Liu, C.-J.; Teng, S.-C.; Wu, K.-J. Direct regulation of TWIST by HIF-1α promotes metastasis. Nat. Cell Biol. 2008, 10, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-H.; Wu, K.-J. TWIST activation by hypoxia inducible factor-1 (HIF-1): Implications in metastasis and development. Cell Cycle 2008, 7, 2090–2096. [Google Scholar] [CrossRef] [PubMed]
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Graham, P.H.; Hao, J.; Bucci, J.; Cozzi, P.J.; Kearsley, J.H.; Li, Y. Emerging roles of radioresistance in prostate cancer metastasis and radiation therapy. Cancer Metastasis Rev. 2014, 33, 469–496. [Google Scholar] [CrossRef]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar] [CrossRef]
- Ralph, S.J.; Pritchard, R.; Rodríguez-Enríquez, S.; Moreno-Sánchez, R.; Ralph, R.K. Hitting the Bull’s-Eye in Metastatic Cancers—NSAIDs Elevate ROS in Mitochondria, Inducing Malignant Cell Death. Pharmaceuticals 2015, 8, 62–106. [Google Scholar] [CrossRef] [Green Version]
- Hagen, T.; Taylor, C.T.; Lam, F.; Moncada, S. Redistribution of Intracellular Oxygen in Hypoxia by Nitric Oxide: Effect on HIF1α. Science 2003, 302, 1975–1978. [Google Scholar] [CrossRef]
- Zhu, L.; Hu, S.; Chen, Q.; Zhang, H.; Fu, J.; Zhou, Y.; Bai, Y.; Pan, Y.; Shao, C. Macrophage contributes to radiation-induced anti-tumor abscopal effect on transplanted breast cancer by HMGB1/TNF-α signaling factors. Int. J. Biol. Sci. 2021, 17, 926–941. [Google Scholar] [CrossRef]
- Feng, W.; Liu, J.; Zhang, D.; Tan, Y.; Cheng, H.; Peng, C. Revealing the efficacy-toxicity relationship of Fuzi in treating rheumatoid arthritis by systems pharmacology. Sci. Rep. 2021, 11, 23083. [Google Scholar] [CrossRef]
- Samal, S.K.; Routray, S.; Veeramachaneni, G.K.; Dash, R.; Botlagunta, M. Ketorolac salt is a newly discovered DDX3 inhibitor to treat oral cancer. Sci. Rep. 2015, 5, 9982. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| β-catenin | GGGATTGGCTTTAGGCCTGT | GAAATTGCCGTAGCGGGTTC |
| E-cadherin | GACGCCATCAACACCGAGTT | AAATTGCCAGGCTCAATGAC |
| N-cadherin | GGTGGAGGAGAAGAAGACCAG | GGCATCAGGCTCCACAGT |
| Vimentin | GACGCCATCAACACCGAGTT | CTTTGTCGTTGGTTAGCTGGT |
| Snail | CTTCCAGCAGCCCTACGAC | CGGTGGGGTTGAGGATCT |
| HIF-1α | TTATCAACACCCCCTGCGACCT | GCCACATACTCCGTCAGAAAGC |
| VEGF | CTCATCAGTTGCCACTTCCACATA | AGCAATTCATCTGTGCTTTCATGTC |
| ERK | TCACAGGTACAGGGATGAGGACAC | CAAAGCACAGCAATGTCCTGAAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, X.; Liu, Y.; Qu, L.; Fan, C.; Ma, X.; Ouyang, P.; Fan, D. Ginsenoside Rh3 Inhibits Lung Cancer Metastasis by Targeting Extracellular Signal-Regulated Kinase: A Network Pharmacology Study. Pharmaceuticals 2022, 15, 758. https://doi.org/10.3390/ph15060758
Xue X, Liu Y, Qu L, Fan C, Ma X, Ouyang P, Fan D. Ginsenoside Rh3 Inhibits Lung Cancer Metastasis by Targeting Extracellular Signal-Regulated Kinase: A Network Pharmacology Study. Pharmaceuticals. 2022; 15(6):758. https://doi.org/10.3390/ph15060758
Chicago/Turabian StyleXue, Xiaodan, Yannan Liu, Linlin Qu, Cuiying Fan, Xiaoxuan Ma, Pingkai Ouyang, and Daidi Fan. 2022. "Ginsenoside Rh3 Inhibits Lung Cancer Metastasis by Targeting Extracellular Signal-Regulated Kinase: A Network Pharmacology Study" Pharmaceuticals 15, no. 6: 758. https://doi.org/10.3390/ph15060758
APA StyleXue, X., Liu, Y., Qu, L., Fan, C., Ma, X., Ouyang, P., & Fan, D. (2022). Ginsenoside Rh3 Inhibits Lung Cancer Metastasis by Targeting Extracellular Signal-Regulated Kinase: A Network Pharmacology Study. Pharmaceuticals, 15(6), 758. https://doi.org/10.3390/ph15060758