
Resveratrol and Curcumin for Chagas Disease Treatment—A Systematic Review

 , , ,
, , ,  and
and
Abstract
:1. Introduction
- ⟹
- Are RVT and CUR trypanocide agents?
- ⟹
- Are there any RVT and CUR benefits in in vivo-infected animals with T. cruzi which can support the clinical study?
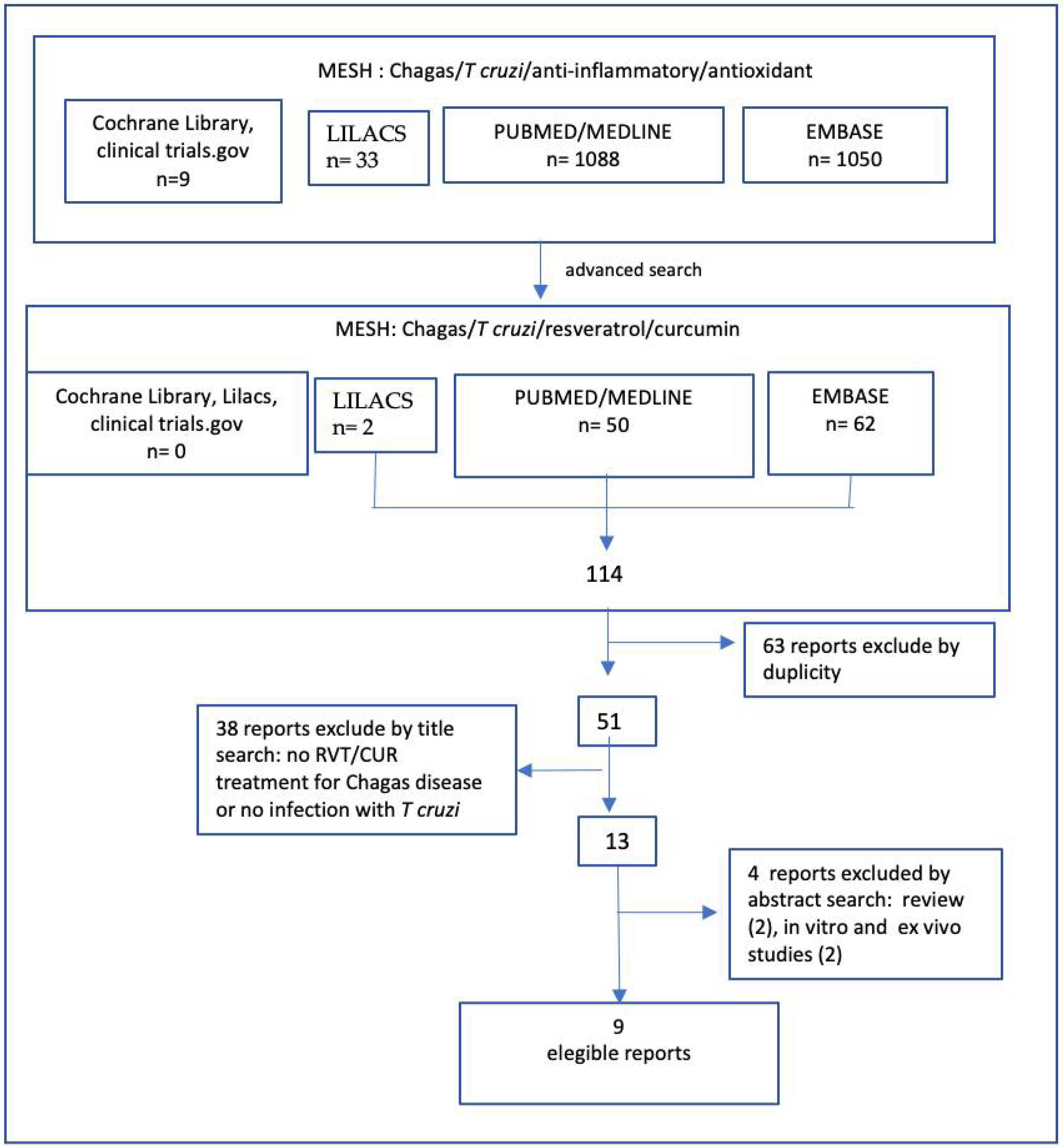
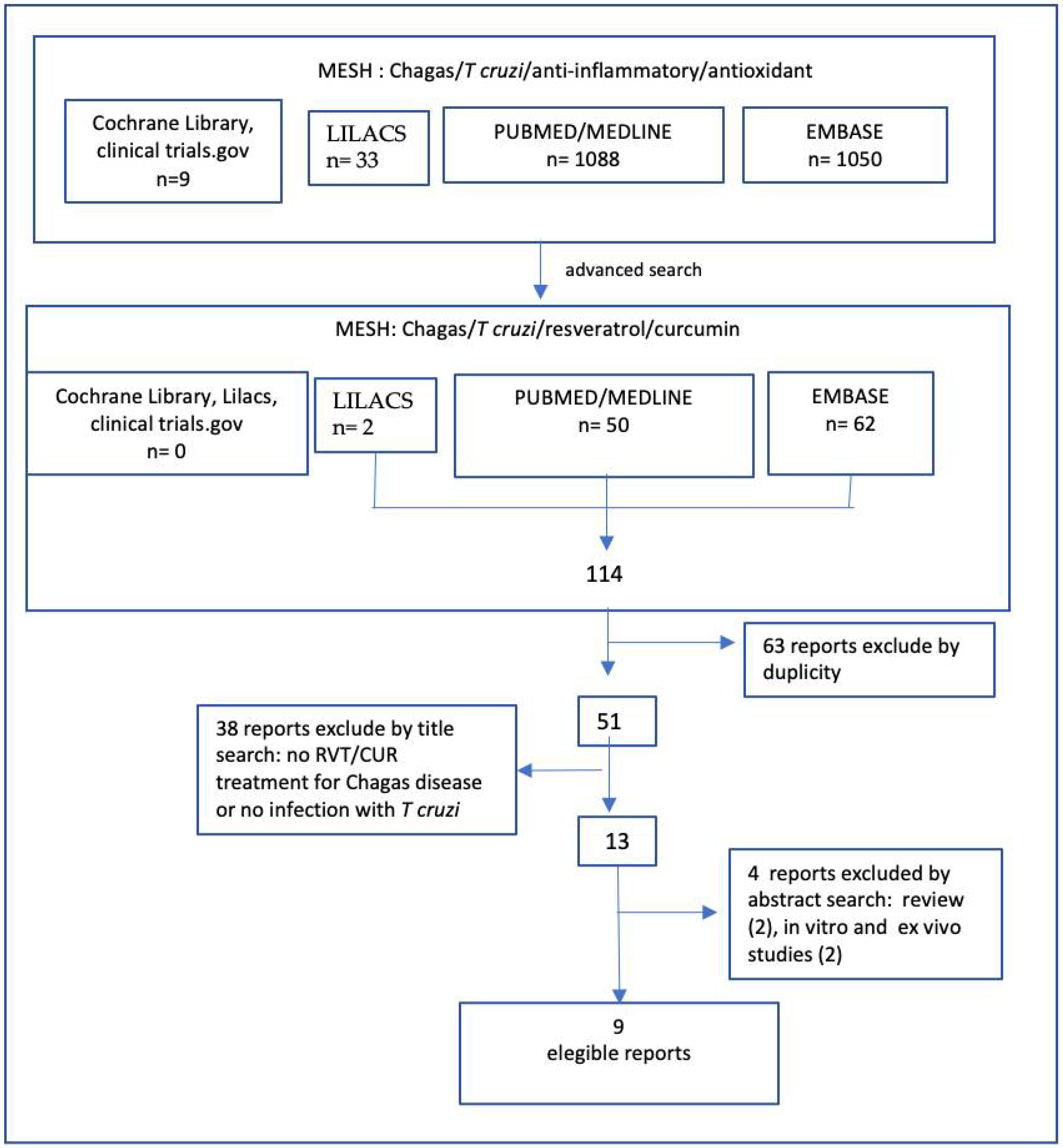
2. Methodology
Literature Search
3. Results
Literature Search and Study Selection
4. Discussion
4.1. Limitations of the Study
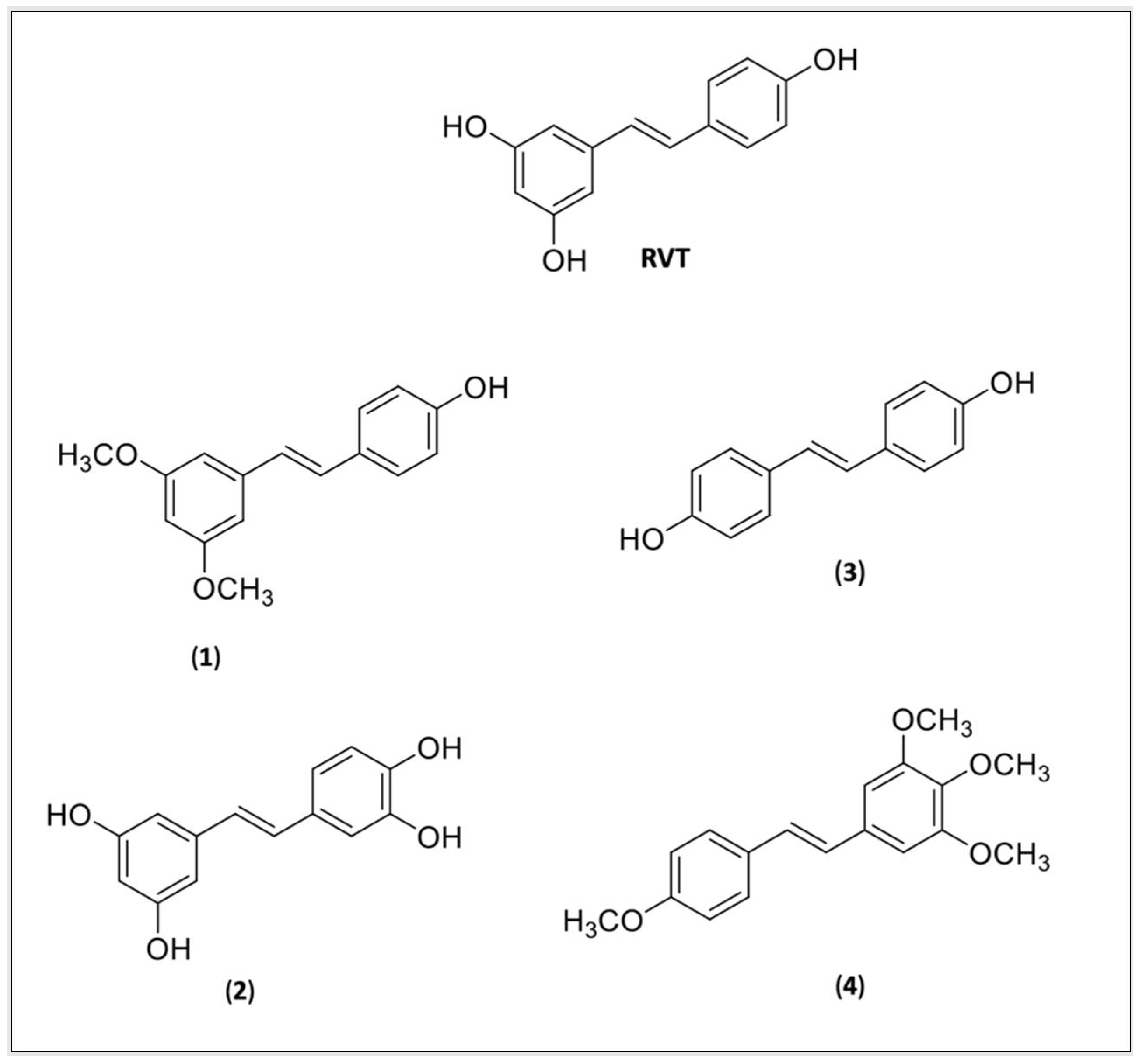
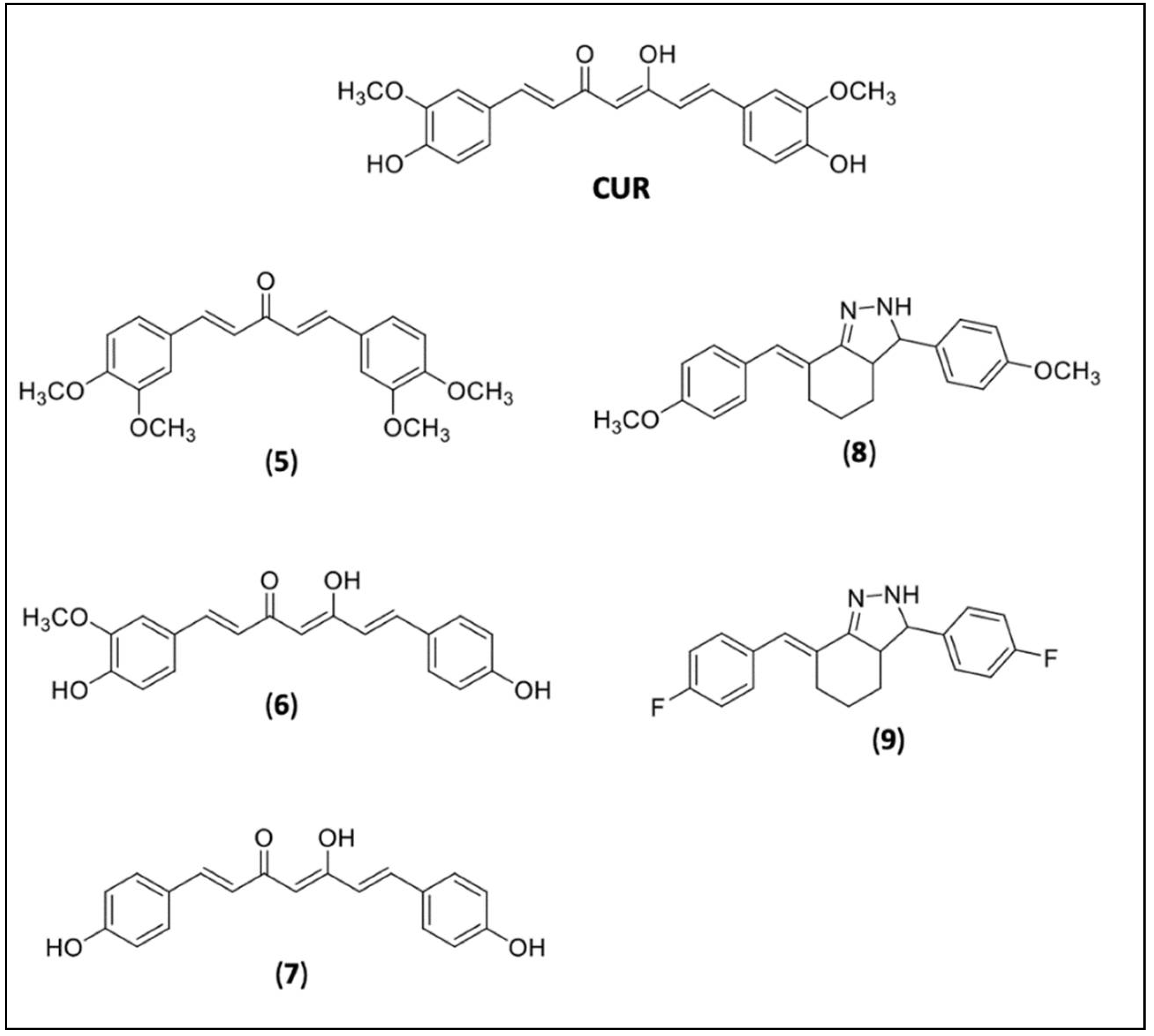
4.2. RVT and CUR Derivatives
5. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mills, R.M. Chagas Disease: Epidemiology and Barriers to Treatment. Am. J. Med. 2020, 11, 1262–1265. [Google Scholar] [CrossRef] [PubMed]
- DNDi, Drug for Neglected Diseases Initiative, América Latina. Available online: https://www.dndial.org/doencas/doenca-chagas/ (accessed on 3 December 2021).
- WHO. World Health Organization. Chagas Disease (American Tripanosomiasis). Available online: https://www.who.int/health-topics/chagas-disease#tab=tab_1 (accessed on 5 January 2022).
- Bern, C. Chagas Disease. N. Engl. J. Med. 2015, 5, 456–466. [Google Scholar] [CrossRef]
- Pérez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 10115, 82–94. [Google Scholar] [CrossRef]
- Echeverria, L.E.; Morillo, C.A. American Trypanosomiasis (Chagas Disease). Infect. Dis. Clin. N. Am. 2019, 1, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Velasco, A.; Morillo, C.A. Chagas heart disease: A contemporary review. J. Nucl. Cardiol. 2020, 2, 445–451. [Google Scholar] [CrossRef]
- Shikanai-Yasuda, M.A.; Carvalho, N.B. Oral transmission of Chagas disease. Clin. Infect. Dis. 2012, 54, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Pereira, K.S.; Schmidt, F.L.; Barbosa, R.L.; Guaraldo, A.M.; Franco, R.M.; Dias, V.L.; Passos, L.A. Transmission of chagas disease (American trypanosomiasis) by food. Adv. Food Nutr. Res. 2010, 59, 63–85. [Google Scholar] [CrossRef]
- Filigheddu, M.T.; Górgolas, M.; Ramos, J.M. Orally-transmitted Chagas disease. Med. Clin. 2017, 3, 125–131. [Google Scholar] [CrossRef]
- Dantas, R.O. Management of Esophageal Dysphagia in Chagas Disease. Dysphagia 2021, 3, 517–522. [Google Scholar] [CrossRef]
- Farias, G.F.A.; Moura, D.T.H.; Moura, E.T.H.; Rezende, D.T.; Hathorn, K.E.; Nasi, A.; Queiroz, N.S.F.; Moura, E.G.H. Peroral endoscopic myotomy (POEM): A comparative study between chagasic and idiopathic achalasia. Endosc. Int. Open 2020, 8, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, A.M.; Thapar, N.; Karunaratne, T.N.; De Giorgio, R. Clinical aspects of neurointestinal disease: Pathophysiology, diagnosis and treatment. Dev. Biol. 2016, 417, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Lannes-Vieira, J.; Silverio, J.C.; Pereira, I.R.; Vinagre, N.F.; Carvalho, C.M.; Paiva, C.N.; da Silva, A.A. Chronic Trypanosoma cruzi-elicited cardiomyopathy: From the discovery to the proposal of rational therapeutic interventions targeting cell adhesion molecules and chemokine receptors—How to make a dream come true. Mem. Inst. Oswaldo Cruz 2009, 1, 226–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonney, K.M.; Luthringer, D.J.; Kim, S.A.; Garg, N.J.; Engman, D.M. Pathology and Pathogenesis of Chagas Heart Disease. Annu. Rev. Pathol. 2019, 14, 421–447. [Google Scholar] [CrossRef] [PubMed]
- Groom, Z.C.; Zochios, V.; Protopapas, A.D. Translating Chagasic dilating cardiomyopathy to surgical therapies: An under published global challenge. SAGE Open Med. 2019, 18, 21–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lidani, K.C.F.; Sandri, T.L.; Castillo-Neyra, R.; Andrade, F.A.; Guimarães, C.M.; Marques, E.N.; Beltrame, M.H.; Gilman, R.H.; Messias-Reason, I. Clinical and epidemiological aspects of chronic Chagas disease from Southern Brazil. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200225. [Google Scholar] [CrossRef] [PubMed]
- Guarner, J. Chagas disease as example of a reemerging parasite. Semin Diagn Pathol. 2019, 3, 164–169. [Google Scholar] [CrossRef]
- Thakare, R.; Dasgupta, A.; Chopra, S. Update on nifurtimox for treatment of Chagas disease. Drugs Today 2021, 4, 251–263. [Google Scholar] [CrossRef]
- Talvani, A.; Teixeira, M.M. Inflammation and Chagas disease some mechanisms and relevance. Adv. Parasitol. 2011, 76, 171–194. [Google Scholar] [CrossRef]
- Dutra, W.O.; Menezes, C.A.; Magalhães, L.M.; Gollob, K.J. Immunoregulatory networks in human Chagas disease. Parasite Immunol. 2014, 8, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.S.; Silva, D.K.C.; Dos Reis, B.P.Z.C.; Barreto, B.C.; Cardoso, C.M.A.; Ribeiro Dos Santos, R.; Meira, C.S.; Soares, M.B.P. Immunomodulation for the Treatment of Chronic Chagas Disease Cardiomyopathy: A New Approach to an Old Enemy. Front. Cell. Infect. Microbiol. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Marin-Neto, J.A.; Simões, M.V.; Rassi Junior, A. Pathogenesis of chronic Chagas cardiomyopathy: The role of coronary microvascular derangements. Rev. Soc. Bras. Med. Trop. 2013, 46, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Santos, É.; Menezes Falcão, L. Chagas cardiomyopathy and heart failure: From epidemiology to treatment. Rev. Port. Cardiol. 2020, 39, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Cuba, M.B.; Machado, M.P.; Farnesi, T.S.; Alves, A.C.; Martins, L.A.; de Oliveira, L.F.; Capitelli, C.S.; Leite, C.F.; Vinícius Silva, M.; Machado, J.R.; et al. Effects of cholinergic stimulation with pyridostigmine bromide on chronic chagasic cardiomyopathic mice. Mediat. Inflamm. 2014, 2014, 475946. [Google Scholar] [CrossRef] [PubMed]
- Nunes, J.P.S.; Andrieux, P.; Brochet, P.; Almeida, R.R.; Kitano, E.; Honda, A.K.; Iwai, L.K.; Andrade-Silva, D.; Goudenège, D.; Alcântara, K.S.; et al. Co-Exposure of Cardiomyocytes to IFN-γ and TNF-α Induces Mitochondrial Dysfunction and Nitro-Oxidative Stress: Implications for the Pathogenesis of Chronic Chagas Disease Cardiomyopathy. Front. Immunol. 2021, 12, 755862. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.R.; Shrestha-Bajracharya, D.; Almeida-Leite, C.M.; Leite, R.; Bahia, M.T.; Talvani, A. Short-term therapy with simvastatin reduces inflammatory mediators and heart inflammation during the acute phase of experimental Chagas disease. Mem. Inst. Oswaldo Cruz 2012, 4, 513–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, L.; Caldas, I.S.; Azevedo, M.A.; Gonçalves, K.R.; da Silva do Nascimento, A.F.; Figueiredo, V.P.; de Figueiredo, D.L.; de Lima, W.G.; Torres, R.M.; Bahia, M.T.; et al. Low doses of simvastatin therapy ameliorate cardiac inflammatory remodeling in Trypanosoma cruzi-infected dogs. Am. J. Trop. Med. Hyg. 2011, 2, 325–331. [Google Scholar] [CrossRef] [Green Version]
- González-Herrera, F.; Cramer, A.; Pimentel, P.; Castillo, C.; Liempi, A.; Kemmerling, U.; Machado, F.S.; Maya, J.D. Simvastatin Attenuates Endothelial Activation through 15-Epi-Lipoxin A4 Production in Murine Chronic Chagas Cardiomyopathy. Antimicrob. Agents Chemother. 2017, 61, e02137-16, Erratum in Antimicrob Agents Chemother. 2017, 5.. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Rivera, D.; Liempi, A.; González-Herrera, F.; Fuentes-Retamal, S.; Carrillo, I.; Abarca, P.; Castillo, C.; Kemmerling, U.; Pesce, B.; Maya, J.D. Simvastatin Improves Cardiac Function through Notch 1 Activation in BALB/c Mice with Chronic Chagas Cardiomyopathy. Antimicrob. Agents Chemother. 2020, 64, e02141-19. [Google Scholar] [CrossRef]
- NCT04984616 Atorvastatin on Inflammation and Cardiac Function in Chronic Chagas Disease (ATOCHA). Available online: https://www.clinicaltrials.gov/ct2/show/NCT04984616?cond=NCT04984616&draw=2 (accessed on 10 January 2022).
- NCT00875173 Selenium Treatment and Chagasic Cardiopathy (STCC). Available online: https://www.clinicaltrials.gov/ct2/show/NCT01863576?cond=NCT01863576&draw=2&rank= (accessed on 10 January 2022).
- NCT01863576 Effects of Omega-3 Supplementation on the Cytokine and Lipid Profiles in Patients with Chronic Chagas Cardiomyopathy. Available online: https://www.clinicaltrials.gov/ct2/show/NCT01863576?cond=NCT01863576&draw=2&rank=1 (accessed on 10 January 2022).
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 5, 1851–1891. [Google Scholar] [CrossRef]
- Galiniak, S.; Aebisher, D.; Bartusik-Aebisher, D. Health benefits of resveratrol administration. Acta Biochim. Pol. 2019, 1, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-Inflammatory Action and Mechanisms of Resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef] [PubMed]
- Chalons, P.; Amor, S.; Courtaut, F.; Cantos-Villar, E.; Richard, T.; Auger, C.; Chabert, P.; Schni-Kerth, V.; Aires, V.; Delmas, D. Study of Potential Anti-Inflammatory Effects of Red Wine Extract and Resveratrol through a Modulation of Interleukin-1-Beta in Macrophages. Nutrients 2018, 10, 1856. [Google Scholar] [CrossRef] [Green Version]
- Bononi, I.; Tedeschi, P.; Mantovani, V.; Maietti, A.; Mazzoni, E.; Pancaldi, C.; Brandolini, V.; Tognon, M. Antioxidant Activity of Resveratrol Diastereomeric Forms Assayed in Fluorescent-Engineered Human Keratinocytes. Antioxidants 2022, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Daiber, A.; Förstermann, U.; Li, H. Antioxidant effects of resveratrol in the cardiovascular system. Br. J. Pharmacol. 2017, 12, 1633–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyck, G.J.B.; Raj, P.; Zieroth, S.; Dyck, J.R.B.; Ezekowitz, J.A. The Effects of Resveratrol in Patients with Cardiovascular Disease and Heart Failure: A Narrative Review. Int. J. Mol. Sci. 2019, 20, 904. [Google Scholar] [CrossRef] [Green Version]
- Gal, R.; Deres, L.; Toth, K.; Halmosi, R.; Habon, T. The Effect of Resveratrol on the Cardiovascular System from Molecular Mechanisms to Clinical Results. Int. J. Mol. Sci. 2021, 22, 10152. [Google Scholar] [CrossRef] [PubMed]
- Mirhadi, E.; Roufogalis, B.D.; Banach, M.; Barati, M.; Sahebkar, A. Resveratrol: Mechanistic and therapeutic perspectives in pulmonary arterial hypertension. Pharmacol. Res. 2021, 163, 105287. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.J.; Fariana, M.; Moni, A.; Hossain, K.S.; Hannan, M.A.; Ha, H. Prospective Pharmacological Potential of Resveratrol in Delaying Kidney Aging. Int. J. Mol. Sci. 2021, 22, 8258. [Google Scholar] [CrossRef]
- Huang, D.D.; Shi, G.; Jiang, Y.; Yao, C.; Zhu, C. A review on the potential of Resveratrol in prevention and therapy of diabetes and diabetic complications. Biomed. Pharmacother. 2020, 125, 109767. [Google Scholar] [CrossRef]
- Springer, M.; Moco, S. Resveratrol and Its Human Metabolites-Effects on Metabolic Health and Obesity. Nutrients 2019, 11, 143. [Google Scholar] [CrossRef] [Green Version]
- Su, C.F.; Jiang, L.; Zhang, X.-W.; Iyaswamy, A.; Li, M. Resveratrol in Rodent Models of Parkinson’s Disease: A Systematic Review of Experimental Studies. Front. Pharmacol. 2021, 12, 879. [Google Scholar] [CrossRef] [PubMed]
- Manjula, R.; Anuja, K.; Alcain, F.J. SIRT1 and SIRT2 Activity Control in Neurodegenerative Diseases. Front. Pharmacol. 2021, 11, 1899. [Google Scholar] [CrossRef] [PubMed]
- Sawda, C.; Moussa, C.; Turner, R.S. Resveratrol for Alzheimer’s Disease. Ann. N. Y. Academ. Sci. 2017, 1403, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.A.Q.; Silva, J.P.B.; Romeiro, C.F.R.; dos Santos, S.M.; Rodrigues, C.A.; Gonçalves, P.R.; Sakai, J.T.; Mendes, P.F.S.; Varela, E.L.P.; Monteiro, M.C. Neuroprotective Mechanisms of Resveratrol in Alzheimer’s Disease: Role of SIRT1. Oxi. Medic. Cell. Longe. 2018, 2018, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jardim, F.R.; de Rossi, F.T.; Nascimento, M.X.; da Silva Barros, R.G.; Borges, P.A.; Prescilio, I.C.; de Oliveira, M.R. Resveratrol and Brain Mitochondria: A Review. Mol. Neurobio. 2017, 55, 2085–2101. [Google Scholar] [CrossRef]
- Griñán-Ferré, C.; Bellver-Sanchis, A.; Izquierdo, V.; Corpas, R.; Roig-Soriano, J.; Chillón, M.; Andres-Lacueva, C.; Somogyvári, M.; Sőti, C.; Sanfeliu, C.; et al. The Pleiotropic Neuroprotective Effects of Resveratrol in Cognitive Decline and Alzheimer’s Disease Pathology: From Antioxidant to Epigenetic Therapy. Ageing Res. Rev. 2021, 67, 101271. [Google Scholar] [CrossRef]
- Pavan, A.; Silva, G.; Jornada, D.; Chiba, D.; Fernandes, G.; Man Chin, C.; dos Santos, J. Unraveling the Anticancer Effect of Curcumin and Resveratrol. Nutrients 2016, 8, 628. [Google Scholar] [CrossRef] [Green Version]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an Anti-Cancer Agent: A Review. Crit. Rev. Food Sci. Nutr. 2017, 58, 1428–1447. [Google Scholar] [CrossRef]
- Huang, X.; Li, X.; Xie, M.; Huang, Z.; Huang, Y.; Wu, G.; Peng, Z.; Sun, Y.; Ming, Q.; Liu, Y.; et al. Resveratrol: Review on Its Discovery, Anti-Leukemia Effects and Pharmacokinetics. Chem. Bio. Interact. 2019, 306, 29–38. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, Q.M.; Lu, Y.Y.; Zhang, H.; Chen, Q.L.; Zhao, M.; Su, S.B. Resveratrol Inhibits the Migration and Metastasis of MDA-MB-231 Human Breast Cancer by Reversing TGF-β1-Induced Epithelial-Mesenchymal Transition. Molecules 2019, 24, 1131. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Chen, L.; Zhu, F.; Han, X.; Sun, L.; Chen, K. The Cytotoxicity Effect of Resveratrol: Cell Cycle Arrest and Induced Apoptosis of Breast Cancer 4T1 Cells. Toxins 2019, 11, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vervandier-Fasseur, D.; Latruffe, N. The Potential Use of Resveratrol for Cancer Prevention. Molecules 2019, 24, 4506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Wang, G.; Jin, G.; Yao, K.; Zhao, Z.; Bie, L.; Guo, Y.; Li, N.; Deng, W.; Chen, X.; et al. Resveratrol Suppresses Colon Cancer Growth by Targeting the AKT/STAT3 Signaling Pathway. Int. J. Mol. Med. 2018, 43, 630–640. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Zhou, M.; Huang, D.; Wasan, H.; Zhang, K.; Sun, L.; Huang, H.; Ma, S.; Shen, M.; Ruan, S. Resveratrol Inhibits the Invasion and Metastasis of Colon Cancer through Reversal of Epithelial-Mesenchymal Transition via the AKT/GSK-3β/Snail Signaling Pathway. Mol. Med. Rep. 2019, 20, 2783–2795. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Yu, D.; Peng, Y.; Yi, H.; Wang, Y.; Cheng, T.; Shi, B.; Yang, G.; Lai, W.; Wu, X.; et al. Resveratrol Induces AMPK and MTOR Signaling Inhibition-Mediated Autophagy and Apoptosis in Multiple Myeloma Cells. Acta Biochim. Biophys. Sini. 2021, 53, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Sorrenti, V.; Buriani, A.; Fortinguerra, S.; Scapagnini, G.; Zella, D. Resveratrol, Rapamycin and Metformin as Modulators of Antiviral Pathways. Viruses 2020, 12, 1458. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liao, D.; Zhou, G.; Zhu, Z.; Cui, Y.; Pu, R. Antiviral Activities of Resveratrol against Rotavirus In Vitro and In Vivo. Phytomedicine 2020, 77, 153230. [Google Scholar] [CrossRef]
- Filardo, S.; Di Pietro, M.; Mastromarino, P.; Sessa, R. Therapeutic Potential of Resveratrol against Emerging Respiratory Viral Infections. Pharmacol. Ther. 2020, 214, 107613. [Google Scholar] [CrossRef]
- Vestergaard, M.; Ingmer, H. Antibacterial and Antifungal Properties of Resveratrol. Int. J. Antimicro. Agents 2019, 53, 716–723. [Google Scholar] [CrossRef]
- Ma, D.S.L.; Tan, L.T.-H.; Chan, K.-G.; Yap, W.H.; Pusparajah, P.; Chuah, L.-H.; Ming, L.C.; Khan, T.M.; Lee, L.-H.; Goh, B.-H. Resveratrol—Potential Antibacterial Agent against Foodborne Pathogens. Front. Pharma. 2018, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol Improves Health and Survival of Mice on a High-Calorie Diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Price, N.L.; Gomes, A.P.; Ling, A.J.Y.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 Is Required for AMPK Activation and the Beneficial Effects of Resveratrol on Mitochondrial Function. Cell Metabol. 2012, 15, 675–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H.; Lee, J.-H.; Lee, H.-Y.; Min, K.-J. Sirtuin Signaling in Cellular Senescence and Aging. BMB Rep. 2019, 52, 24–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.R.; Li, S.; Lin, C.C. Effect of Resveratrol and Pterostilbene on Aging and Longevity. BioFactors 2017, 44, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Bradamante, S.; Barenghi, L.; Piccinini, F.; Bertelli, A.A.E.; De Jonge, R.; Beemster, P.; De Jong, J.W. Resveratrol Provides Late-Phase Cardioprotection by Means of a Nitric Oxide- and Adenosine-Mediated Mechanism. Euro. J. Pharma. 2003, 465, 115–123. [Google Scholar] [CrossRef]
- Hung, L.M.; Su, M.J.; Chen, J.K. Resveratrol Protects Myocardial Ischemia–Reperfusion Injury through Both NO-Dependent and NO-Independent Mechanisms. Free Rad. Bio. Med. 2004, 36, 774–781. [Google Scholar] [CrossRef]
- Lee, Y.; Im, E. Regulation of MiRNAs by Natural Antioxidants in Cardiovascular Diseases: Focus on SIRT1 and ENOS. Antioxidants 2021, 10, 377. [Google Scholar] [CrossRef]
- Xia, N.; Förstermann, U.; Li, H. Resveratrol and Endothelial Nitric Oxide. Molecules 2014, 19, 16102–16121. [Google Scholar] [CrossRef]
- Chaplin, A.; Carpéné, C.; Mercader, J. Resveratrol, Metabolic Syndrome, and Gut Microbiota. Nutrients 2018, 10, 1651. [Google Scholar] [CrossRef] [Green Version]
- Koh, Y.; Lee, P.; Kuo, Y.; Nagabhushanam, K.; Ho, C.; Pan, M. Dietary Pterostilbene and Resveratrol Modulate the Gut Microbiota Influenced by Circadian Rhythm Dysregulation. Mol. Nutr. Food Res. 2021, 65, 2100434. [Google Scholar] [CrossRef]
- Xu, T.; Lu, B. The Effects of Phytochemicals on Circadian Rhythm and Related Diseases. Crit. Rev. Food Sci. Nutr. 2018, 59, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Ramirez-Tortosa, M. Curcumin and Health. Molecules 2016, 21, 264. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Wang, L.; Yin, R.; Hudlikar, R.; Li, S.; Kuo, H.D.; Peter, R.; Sargsyan, D.; Guo, Y.; Liu, X.; et al. Epigenetics/Epigenomics and Prevention by Curcumin of Early Stages of Inflammatory-Driven Colon Cancer. Mol. Carci. 2019, 59, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanyam, K.; Varier, R.A.; Altaf, M.; Swaminathan, V.; Siddappa, N.B.; Ranga, U.; Kundu, T.K. Curcumin, a Novel P300/CREB-Binding Protein-Specific Inhibitor of Acetyltransferase, Represses the Acetylation of Histone/Nonhistone Proteins and Histone Acetyltransferase-Dependent Chromatin Transcription. J. Biol. Chem. 2004, 279, 51163–51171. [Google Scholar] [CrossRef] [Green Version]
- Soflaei, S.S.; Momtazi-Borojeni, A.A.; Majeed, M.; Derosa, G.; Maffioli, P.; Sahebkar, A. Curcumin: A Natural Pan-HDAC Inhibitor in Cancer. Curr. Pharma. Des. 2018, 24, 123–129. [Google Scholar] [CrossRef]
- Venturelli, S.; Berger, A.; Böcker, A.; Busch, C.; Weiland, T.; Noor, S.; Leischner, C.; Schleicher, S.; Mayer, M.; Weiss, T.S.; et al. Resveratrol as a Pan-HDAC Inhibitor Alters the Acetylation Status of Histone [Corrected] Proteins in Human-Derived Hepatoblastoma Cells. PLoS ONE 2013, 8, e73097. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, H.B.; Yang, J.; Wang, J.R.; Liu, J.X.; Li, C.L. Curcumin Alleviates Isoproterenol-Induced Cardiac Hypertrophy and Fibrosis through Inhibition of Autophagy and Activation of MTOR. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7500–7508. [Google Scholar] [CrossRef]
- Li, H.; Sureda, A.; Devkota, H.P.; Pittalà, V.; Barreca, D.; Silva, A.S.; Tewari, D.; Xu, S.; Nabavi, S.M. Curcumin, the Golden Spice in Treating Cardiovascular Diseases. Biotech. Adv. 2020, 38, 107343. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s Risk of Bias Tool for Animal Studies. BMC Med. Res. Method. 2014, 14, 43. [Google Scholar] [CrossRef] [Green Version]
- The ARRIVE Guidelines 2.0. Available online: https://arriveguidelines.org/arrive-guidelines (accessed on 15 January 2022).
- Percie du Sert, N.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting Animal Research: Explanation and Elaboration for the ARRIVE Guidelines 2.0. PLoS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef] [PubMed]
- Vilar-Pereira, G.; Carneiro, V.C.; Mata-Santos, H.; Vicentino, A.R.R.; Ramos, I.P.; Giarola, N.L.L.; Feijó, D.F.; Meyer-Fernandes, J.R.; Paula-Neto, H.A.; Medei, E.; et al. Resveratrol Reverses Functional Chagas Heart Disease in Mice. PLoS Pathog. 2016, 12, e1005947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, X.; Wen, J.; Koo, S.-J.; Liang, L.Y.; Garg, N.J. SIRT1-PGC1α-NFκB Pathway of Oxidative and Inflammatory Stress during Trypanosoma cruzi Infection: Benefits of SIRT1-Targeted Therapy in Improving Heart Function in Chagas Disease. PLoS Pathog. 2016, 12, e1005954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fracasso, M.; Bottari, N.B.; da Silva, A.D.; Grando, T.H.; Pillat, M.M.; Ulrich, H.; Vidal, T.; de Andrade, C.M.; Monteiro, S.G.; Nascimento, L.F.N.; et al. Effects of Resveratrol on the Differentiation Fate of Neural Progenitor Cells of Mouse Embryos Infected with Trypanosoma cruzi. Microb. Pathog. 2019, 132, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Fracasso, M.; Dutra da Silva, A.; Bottari, N.B.; Monteiro, S.G.; Garzon, L.R.; Farias de Souza, L.A.; Schetinger, M.R.C.; Da Silva, A.S. Resveratrol Impacts in Oxidative Stress in Liver during Trypanosoma cruzi Infection. Microb. Pathog. 2021, 153, 104800. [Google Scholar] [CrossRef] [PubMed]
- Nagajyothi, F.; Zhao, D.; Weiss, L.M.; Tanowitz, H.B. Curcumin Treatment Provides Protection against Trypanosoma cruzi Infection. Parasitol. Res. 2012, 110, 2491–2499. [Google Scholar] [CrossRef] [Green Version]
- Novaes, R.D.; Sartini, M.V.P.; Rodrigues, J.P.F.; Gonçalves, R.V.; Santos, E.C.; Souza, R.L.M.; Caldas, I.S. Curcumin Enhances the Anti-Trypanosoma cruzi Activity of Benznidazole-Based Chemotherapy in Acute Experimental Chagas Disease. Antimicro. Agents Chemother. 2016, 60, 3355–3364. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Wicz, S.; Corral, R.S. Cardioprotective Actions of Curcumin on the Pathogenic NFAT/COX-2/Prostaglandin E2 Pathway Induced during Trypanosoma cruzi Infection. Phytomedicine 2016, 23, 1392–1400. [Google Scholar] [CrossRef]
- Hernández, M.; Wicz, S.; Santamaría, M.H.; Corral, R.S. Curcumin Exerts Anti-Inflammatory and Vasoprotective Effects through Amelioration of NFAT-Dependent Endothelin-1 Production in Mice with Acute Chagas Cardiomyopathy. Mem. Inst. Oswaldo Cruz 2018, 113. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Wicz, S.; Pérez Caballero, E.; Santamaría, M.H.; Corral, R.S. Dual Chemotherapy with Benznidazole at Suboptimal Dose plus Curcumin Nanoparticles Mitigates Trypanosoma cruzi-Elicited Chronic Cardiomyopathy. Parasitol. Int. 2021, 81, 102248. [Google Scholar] [CrossRef]
- Shoaib, A.; Tabish, M.; Ali, S.; Arafah, A.; Wahab, S.; Almarshad, F.M.; Rashid, S.; Rehman, M.U. Dietary Phytochemicals in Cancer Signalling Pathways: Role of miRNA Targeting. Curr. Med. Chem. 2021, 28, 8036–8067. [Google Scholar] [CrossRef] [PubMed]
- Lazarin-Bidóia, D.; Garcia, F.P.; Ueda-Nakamura, T.; Silva, S.O.; Nakamura, C.V. Natural compounds based chemotherapeutic against Chagas disease and leishmaniasis: Mitochondrion as a strategic target. Mem. Inst. Oswaldo Cruz. 2022, 117, e220396. [Google Scholar] [CrossRef] [PubMed]
- Valera Vera, E.A.; Sayé, M.; Reigada, C.; Damasceno, F.S.; Silber, A.M.; Miranda, M.R.; Pereira, C.A. Resveratrol Inhibits Trypanosoma cruzi Arginine Kinase and Exerts a Trypanocidal Activity. Int. J. Biol. Macromol. 2016, 87, 498–503. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. PubChem. Resveratrol. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Resveratrol (accessed on 10 March 2022).
- Nagajyothi, F.; Weiss, L.M.; Silver, D.L.; Desruisseaux, M.S.; Scherer, P.E.; Herz, J.; Tanowitz, H.B. Trypanosoma cruzi Utilizes the Host Low Density Lipoprotein Receptor in Invasion. PLoS Negl. Trop. Dis. 2011, 5, e953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mythili, S.; Malathi, N. Diagnostic Markers of Acute Myocardial Infarction. Biomed. Rep. 2015, 3, 743–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walle, T. Bioavailability of Resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of Curcumin: Problems and Promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Chimento, A.; De Amicis, F.; Sirianni, R.; Sinicropi, M.; Puoci, F.; Casaburi, I.; Saturnino, C.; Pezzi, V. Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosuru, R.; Kandula, V.; Rai, U.; Prakash, S.; Xia, Z.; Singh, S. Pterostilbene Decreases Cardiac Oxidative Stress and Inflammation via Activation of AMPK/Nrf2/HO-1 Pathway in Fructose-Fed Diabetic Rats. Cardiovas. Drugs Ther. 2018, 32, 147–163. [Google Scholar] [CrossRef]
- Kosuru, R.; Cai, Y.; Kandula, V.; Yan, D.; Wang, C.; Zheng, H.; Li, Y.; Irwin, M.G.; Singh, S.; Xia, Z. AMPK Contributes to Cardioprotective Effects of Pterostilbene against Myocardial Ischemia- Reperfusion Injury in Diabetic Rats by Suppressing Cardiac Oxidative Stress and Apoptosis. Cell. Phys. Biochem. 2018, 46, 1381–1397. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, L.; Li, F.; Hu, C.-P.; Zhang, Z. Restoration of Sirt1 Function by Pterostilbene Attenuates Hypoxia-Reoxygenation Injury in Cardiomyocytes. Eur. J. Pharm. 2016, 776, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Sun, Y.; Song, M.; Wu, X.; Xiao, H. Gastrointestinal Biotransformation of Resveratrol in Mice. FASEB J. 2016, 30. [Google Scholar] [CrossRef]
- Tang, Y.-L.; Chan, S.-W. A Review of the Pharmacological Effects of Piceatannol on Cardiovascular Diseases. Phytother. Res. 2014, 28, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-P.; Hung, L.-M.; Hsueh, C.-H.; Lai, L.-P.; Su, M.-J. Piceatannol, a Derivative of Resveratrol, Moderately Slows INa Inactivation and Exerts Antiarrhythmic Action in Ischaemia-Reperfused Rat Hearts. Brit. J. Pharm. 2009, 157, 381–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Zhang, Y.; Zhang, C.; Gao, L.; Li, J. Piceatannol Pretreatment Alleviates Acute Cardiac Injury via Regulating PI3K-Akt-ENOS Signaling in H9c2 Cells. Biomed. Pharm. 2019, 109, 886–891. [Google Scholar] [CrossRef]
- Coppa, T.; Lazzè, M.C.; Cazzalini, O.; Perucca, P.; Pizzala, R.; Bianchi, L.; Stivala, L.A.; Forti, L.; Maccario, C.; Vannini, V.; et al. Structure–Activity Relationship of Resveratrol and Its Analogue, 4,4′-Dihydroxy-Trans-Stilbene, toward the Endothelin Axis in Human Endothelial Cells. J. Med. Food 2011, 14, 1173–1180. [Google Scholar] [CrossRef]
- Liu, B.; Luo, X.-J.; Yang, Z.-B.; Zhang, J.-J.; Li, T.-B.; Zhang, X.-J.; Ma, Q.-L.; Zhang, G.-G.; Hu, C.-P.; Peng, J. Inhibition of NOX/VPO1 Pathway and Inflammatory Reaction by Trimethoxystilbene in Prevention of Cardiovascular Remodeling in Hypoxia-Induced Pulmonary Hypertensive Rats. J. Cardiovasc. Pharm. 2014, 63, 567–576. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef] [Green Version]
- Sueth-Santiago, V.; Moraes, J.D.B.; Sobral Alves, E.S.; Vannier-Santos, M.A.; Freire-de-Lima, C.G.; Castro, R.N.; Mendes-Silva, G.P.; Del Cistia, C.D.N.; Magalhães, L.G.; Andricopulo, A.D.; et al. The Effectiveness of Natural Diarylheptanoids against Trypanosoma cruzi: Cytotoxicity, Ultrastructural Alterations and Molecular Modeling Studies. PLoS ONE 2016, 11, e0162926. [Google Scholar] [CrossRef]
- Matiadis, D.; Saporiti, T.; Aguilera, E.; Robert, X.; Guillon, C.; Cabrera, N.; Pérez-Montfort, R.; Sagnou, M.; Alvarez, G. Pyrazol(In)E Derivatives of Curcumin Analogs as a New Class of Anti-Trypanosoma cruzi Agents. Future Med. Chem. 2021, 13, 701–714. [Google Scholar] [CrossRef]

{kind=link}
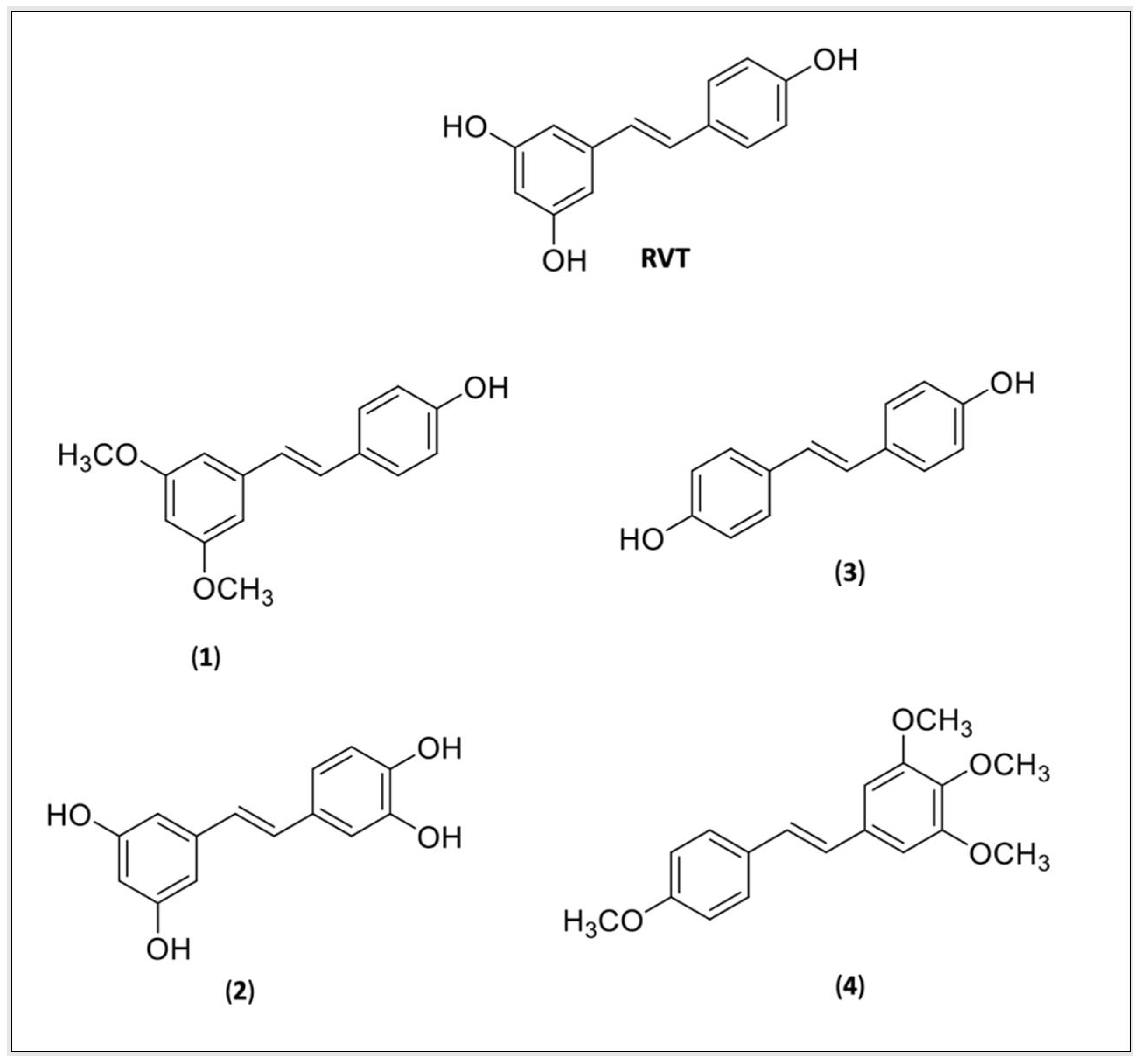{kind=link}
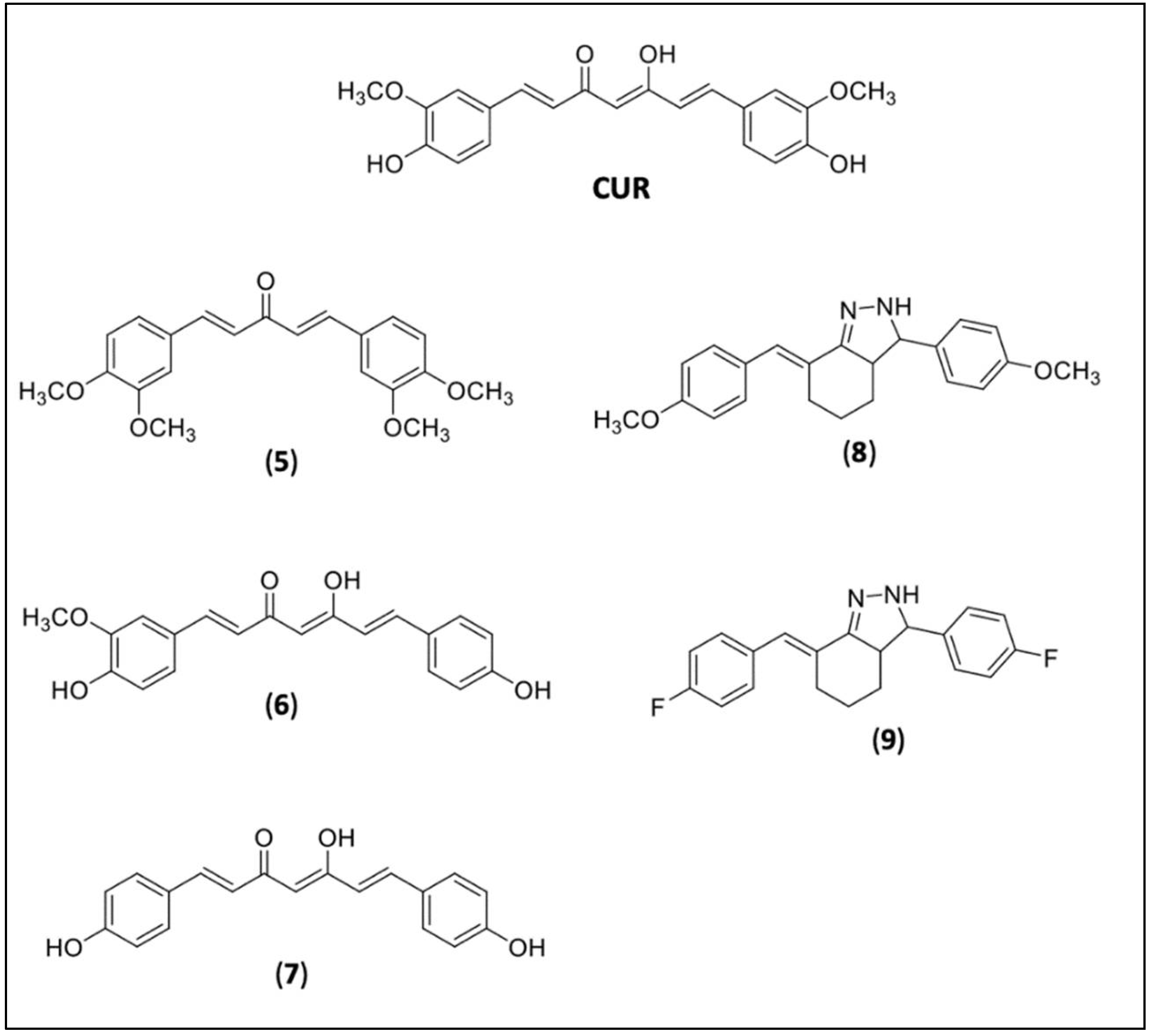{kind=link}
| Reference | Animal Specie | T. cruzi Strain | Treatment | Results | Q | B |
|---|---|---|---|---|---|---|
| Vilar-Pereira et al., 2016 [88] | BALB/c Mice female or male (5–7-week age) | 1 × 102 blood trypomastigote I Colombian strain | 30 days RVT 15 m/.kg/i.p, (vehicle = 10% ethanol/PBS) or 40 mg./Kg (10%ethano/PBS)/daily per os. for 30 days from the establishment of chronic chagasic cardiomyopathy at 60 dpi | At 90 dpi: Primary outcome: RVT decreases heart parasite burden. It was detected by quantitative PCR measured by the relative amount of parasites (TcS18) per host DNA (GADPH) in heart tissue. (Around 90%, p < 0.005 compared to vehicle.) Secondary outcome: The cardiac function:
|  |  |
| Wan et al., 2016 [89] | C57BL/6 mice Mice (6-weeks-old) | 1 × 104 trypomastigotes (SylvioX10 strain, ATCC 50823) were propagated by in vitro passage in C2C12 cells | RVT (20 mg/mL in drinking water) for three weeks, during days 90–111 pi. euthanasia: 150 dpi | Primary outcome: this study did not show parasitemia data. At 150 dpi: Secondary outcome:
|  |  |
| Fracasso et al., 2021 [90] | Swiss mice—female, age not informed | 1 × 104 trypomastigote Y strain | RSV (100 mg/kg) compared to BNZ (100 mg/kg) by gavage daily over 7 days. | Primary outcome: RVT has no effect on parasitemia over 8 dpi. Secondary outcome:
|  |  |
| Fracasso et al., 2021 [91] | Swiss mice—female, age not informed | 1 × 104 trypomastigotes Y strain | RSV (100 mg/kg (by gavage daily over 7 days. | Primary outcome: at 7 dpi: RVT has no influence on parasitemia (trypomastigote forms) Secondary outcomes:
|  |  |
| Animal | T. cruzi Strain | Treatment | Results | Q | B | |
|---|---|---|---|---|---|---|
| Nagajyothi et al., 2012 [92] | Six to 8-week-old male CD-1 mice | Brazil strain (maintained in C3H/He mice) | CUR (100 mg/kg/day orally) for 35 days day of treatment: not informed | Primary outcome: at 23 dpi: decrease of 30% parasitemia compared to infected without treatment heart parasitemia: reduction load in the heart (37% of control, p < 0.05) 100% survival against 60% rate in non-treated animals (p < 0.05) Secondary outcome: 15 dpi:
|  |  |
| Novaes et al., 2016 [93] | 8-week-old Swiss mice (weight, 30.17 ± 3.85 g) | 2 × 103 trypomastigotes Y strain | GROUP 1 CUR (100 mg/kg/day orally) CUR + BZN (50 or 100 mg/kg day orally) 20 days of treatment administered after 4 dpi GROUP 2 recrudescence group negative animals of group 1 treated with 50 mg/kg cyclophosphamide in three cycles of four consecutive days with 3 days between cycles | Primary outcome:
|  |  |
| Hernandez et al., 2016 [94] | Six- to eight-week-old female BALB/c mice | 50 trypomastigotes of RA strain | CUR, 25, 50, or 100 mg/kg body weight/day orally or BZN, 100 mg/kg body weight/day orally for 35 days. | Primary outcome: 21 dpi: cardiac parasitemia: it was not modified with CUR (all doses) against almost 100% decrease of heart parasitism load 35 dpi: 100% survival with 100 mg/kg (but not 25 or 50 mg/kg) and BZN 100 mg/kg against 55% of untreated infected mice Secondary outcome: 21 dpi inflammatory process attenuation in heart tissue with CUR 100 treatment, analyzed by leukocyte infiltration, cyclooxygenase-2 (COX-2), microsomal prostaglandin E synthase-1 (mPGES-1), and B-type natriuretic peptide (BNP) mRNA expression to normal (non-infected animal) level |  |  |
| Hernandez et al., 2018 [95] | C5BL/6 male mice (eight weeks old) | 103 blood trypomastigotes of Tulahuen strain | CUR (100 mg/kg) dissolved in corn oil oral (by gavage) from day 1–14 of infection BZN 100 mg/kg same period | Primary outcome: parasitemia 14 dpi: CUR very little effect on parasitemia profile compared to non-treated animals Secondary outcome:
|  |  |
| Hernandez et al., 2021 [96] | C57BL/6 mice female and male mice (eight weeks old) | 10,000 Brazil strain (DTU I, routinely maintained by serial subinoculation in C3HeJ mice at three-week intervals) | Nano formulated Cur preparations (size range, 250–300 nm) contained 0.15 mg CUR per mg of polymer, suspended in an aqueous solution of 1% wt/vol sodium carboxymethylcellulose and administered orally by gavage (0.15 mL) once a day. Treatment for 30 consecutive days, starting on day 60 of infection. | Primary outcome: at 130 dpi survival: heart tissue parasitemia: T. cruzi load was 4.39-fold (p < 0.01) higher in the myocardium from mice administered with PBS than in those treated with BZ Cur therapy had no significant effect on cardiac parasitism and did not limit BZ parasiticidal activity. Secondary outcome: at 130 dpi
|  |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imperador, C.H.L.; Scarim, C.B.; Bosquesi, P.L.; Lopes, J.R.; Cardinalli Neto, A.; Giarolla, J.; Ferreira, E.I.; dos Santos, J.L.; Chin, C.M. Resveratrol and Curcumin for Chagas Disease Treatment—A Systematic Review. Pharmaceuticals 2022, 15, 609. https://doi.org/10.3390/ph15050609
Imperador CHL, Scarim CB, Bosquesi PL, Lopes JR, Cardinalli Neto A, Giarolla J, Ferreira EI, dos Santos JL, Chin CM. Resveratrol and Curcumin for Chagas Disease Treatment—A Systematic Review. Pharmaceuticals. 2022; 15(5):609. https://doi.org/10.3390/ph15050609
Chicago/Turabian StyleImperador, Carlos Henrique Lima, Cauê Benito Scarim, Priscila Longhin Bosquesi, Juliana Romano Lopes, Augusto Cardinalli Neto, Jeanine Giarolla, Elizabeth Igne Ferreira, Jean Leandro dos Santos, and Chung Man Chin. 2022. "Resveratrol and Curcumin for Chagas Disease Treatment—A Systematic Review" Pharmaceuticals 15, no. 5: 609. https://doi.org/10.3390/ph15050609
APA StyleImperador, C. H. L., Scarim, C. B., Bosquesi, P. L., Lopes, J. R., Cardinalli Neto, A., Giarolla, J., Ferreira, E. I., dos Santos, J. L., & Chin, C. M. (2022). Resveratrol and Curcumin for Chagas Disease Treatment—A Systematic Review. Pharmaceuticals, 15(5), 609. https://doi.org/10.3390/ph15050609