
Carajurin Induces Apoptosis in Leishmania amazonensis Promastigotes through Reactive Oxygen Species Production and Mitochondrial Dysfunction

 , , ,
, , ,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
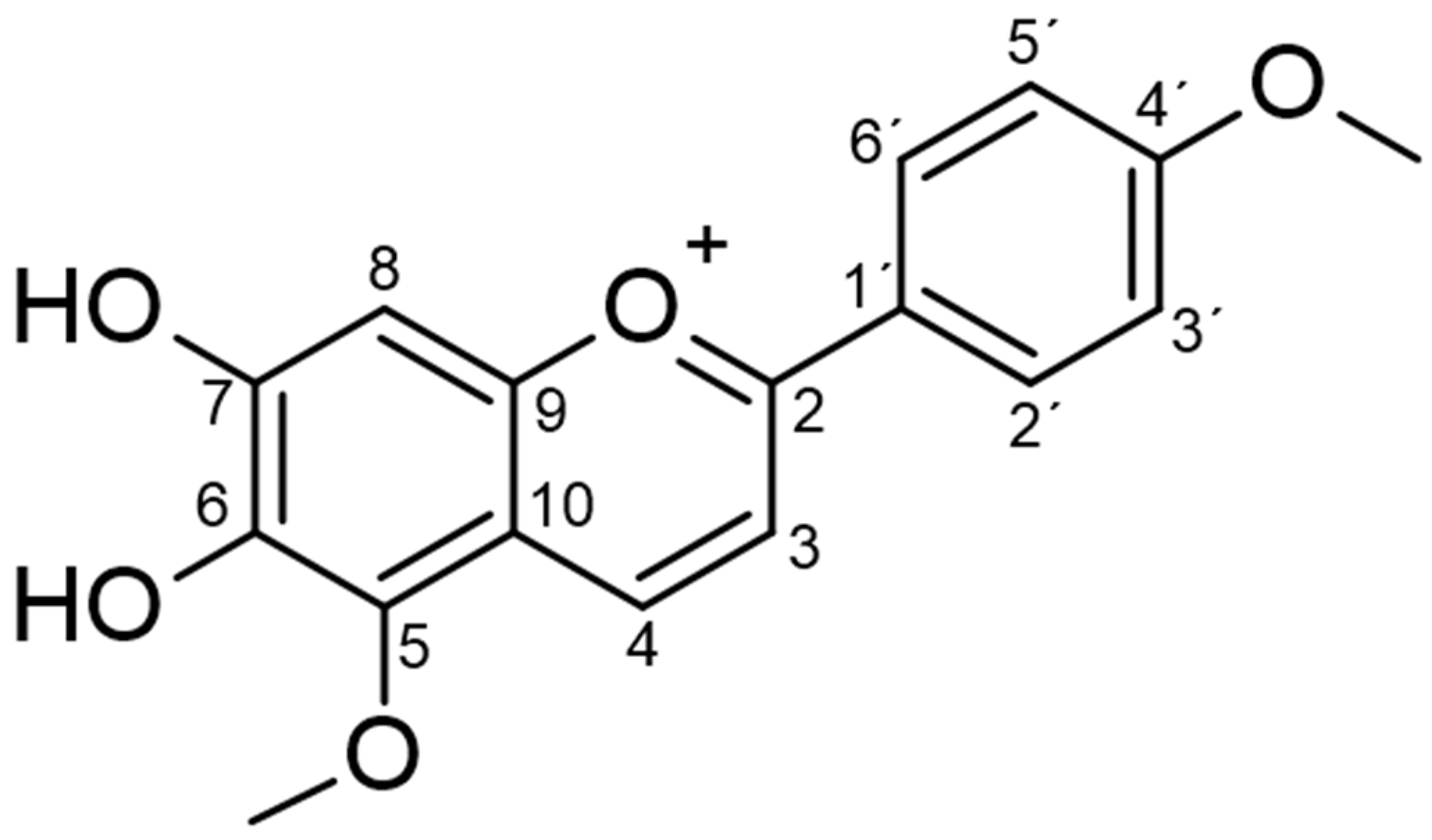
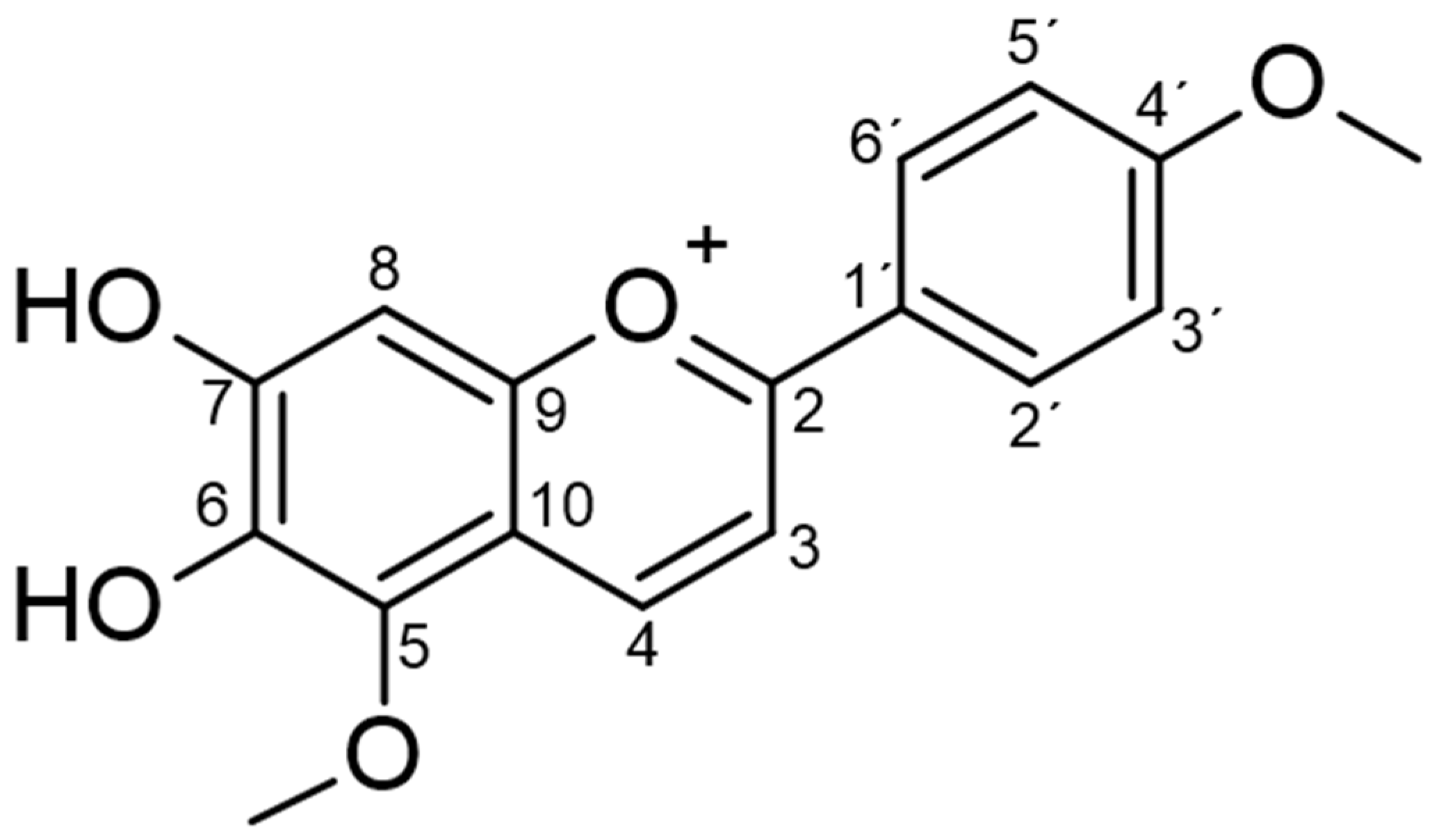
2.1. Phytochemical Analysis
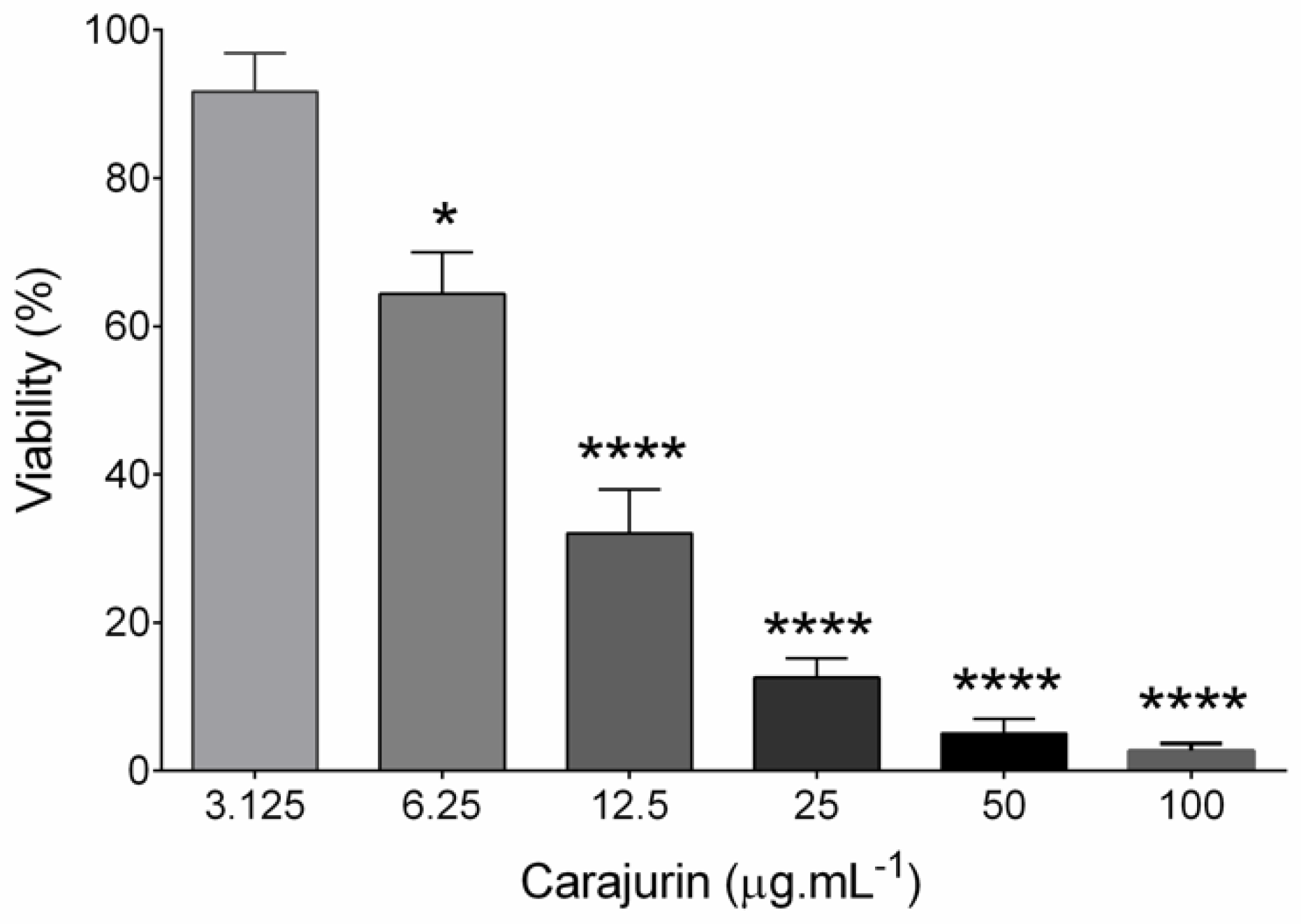
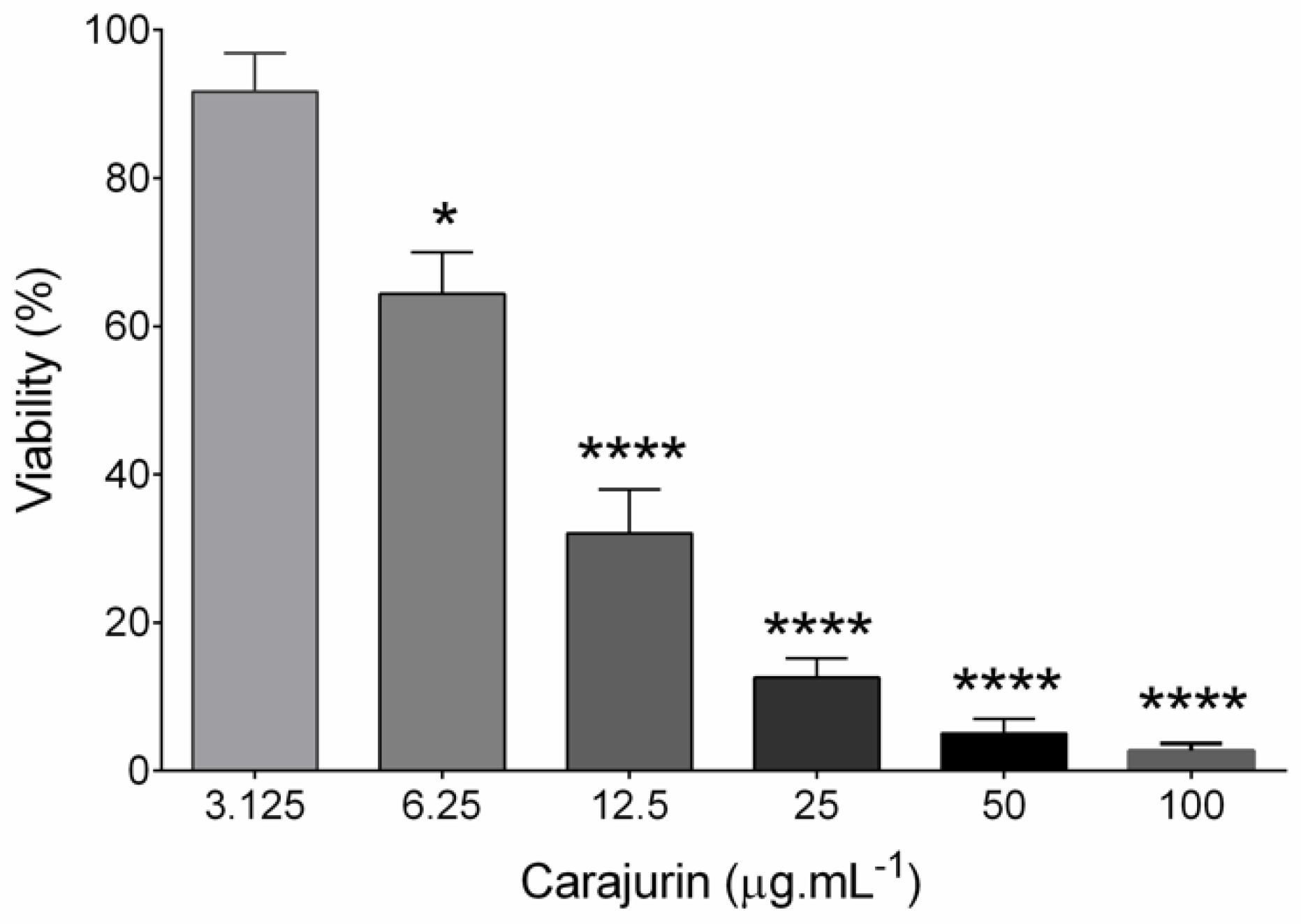
2.2. Anti-leishmanial Activity and Cytotoxicity
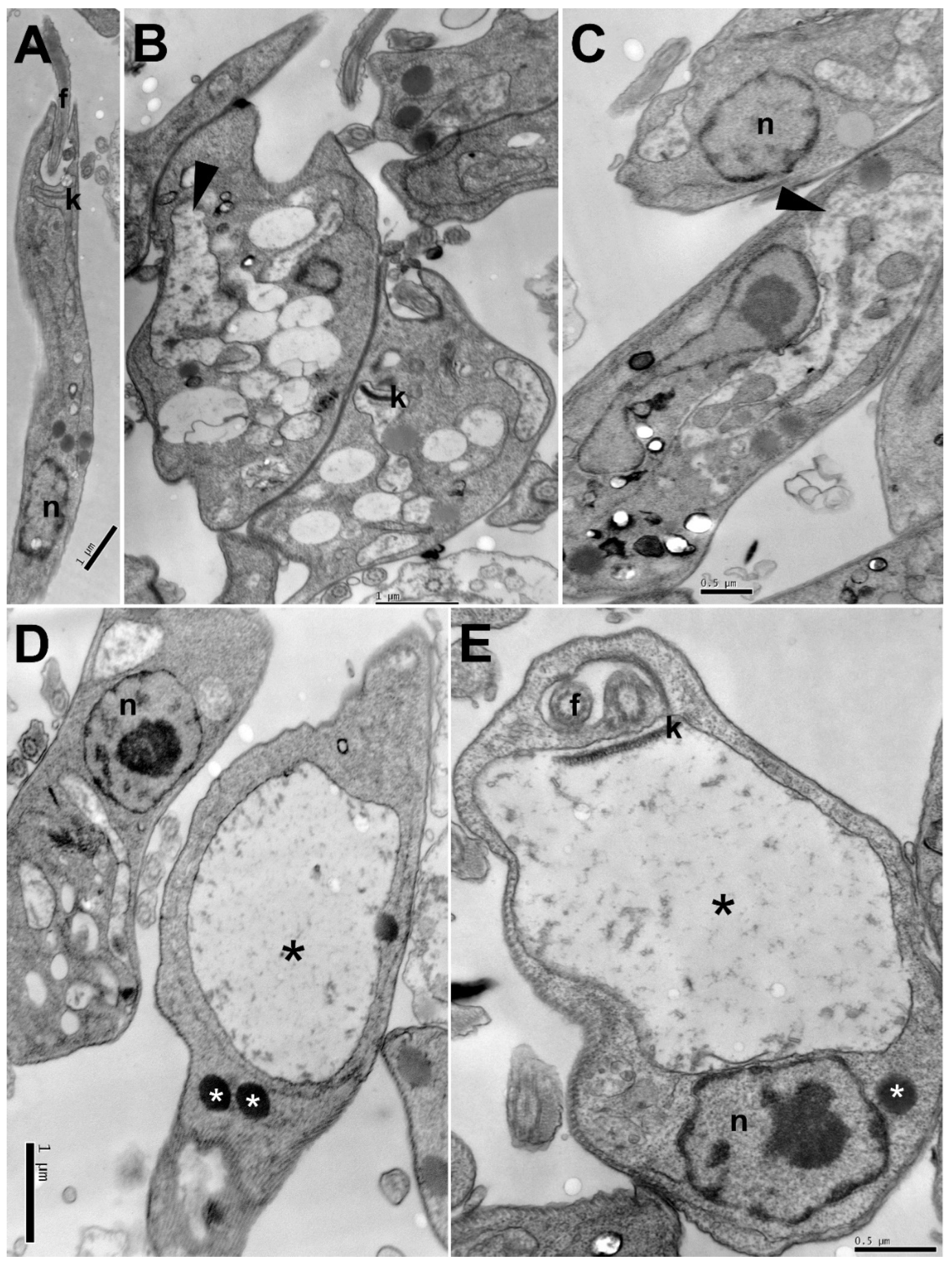
2.3. Ultrastructural Changes
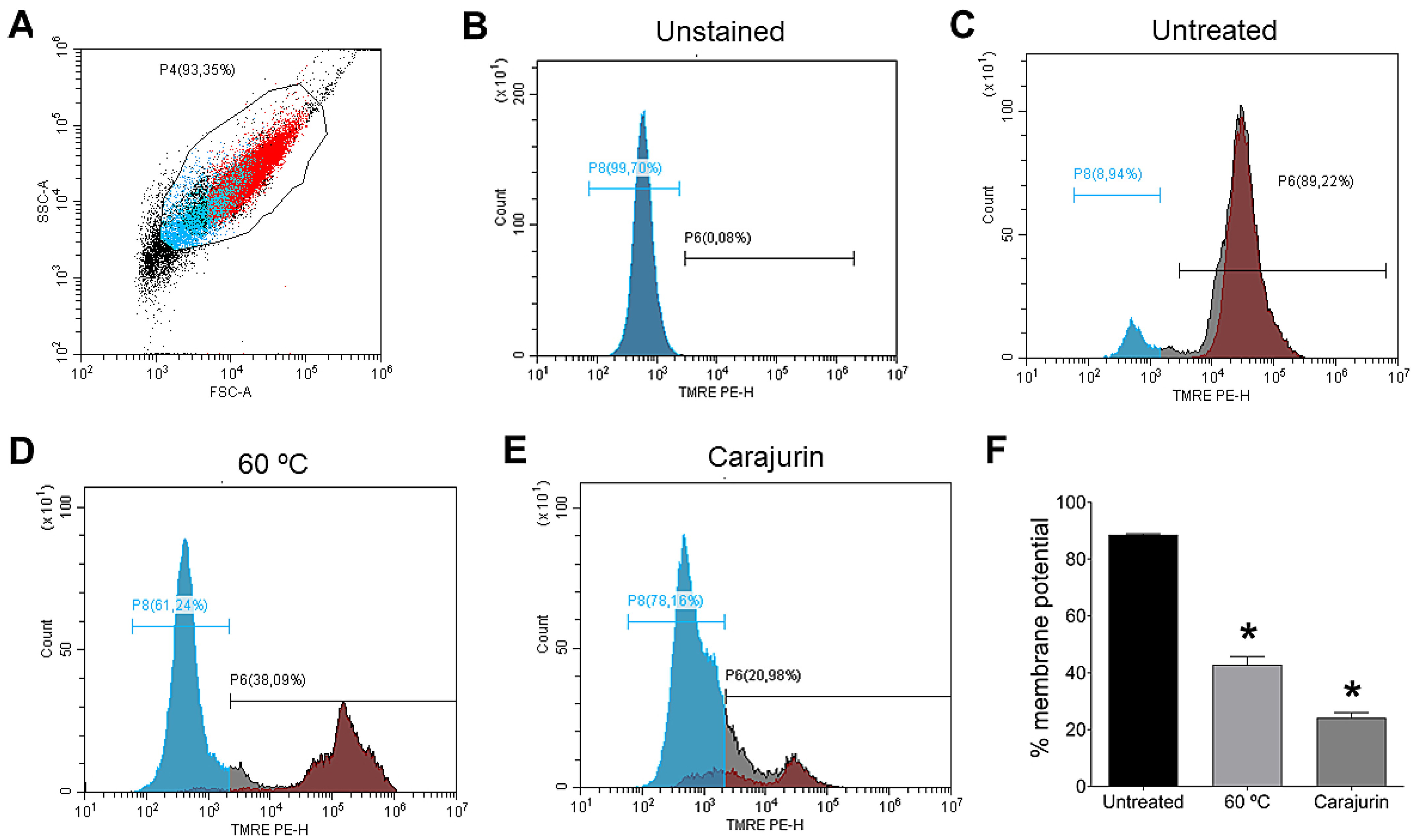
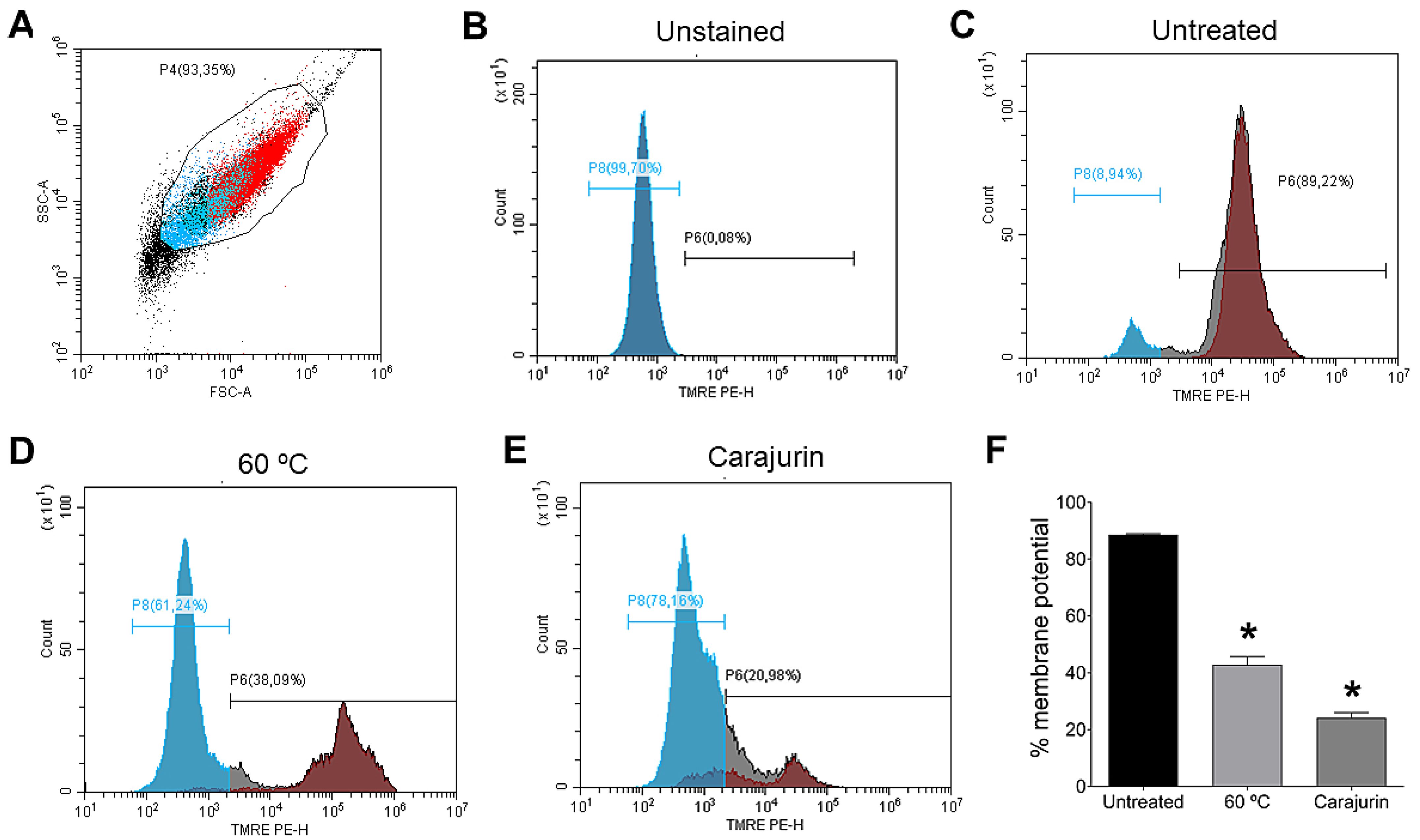
2.4. Mitochondrial Membrane Potential (Δψm)
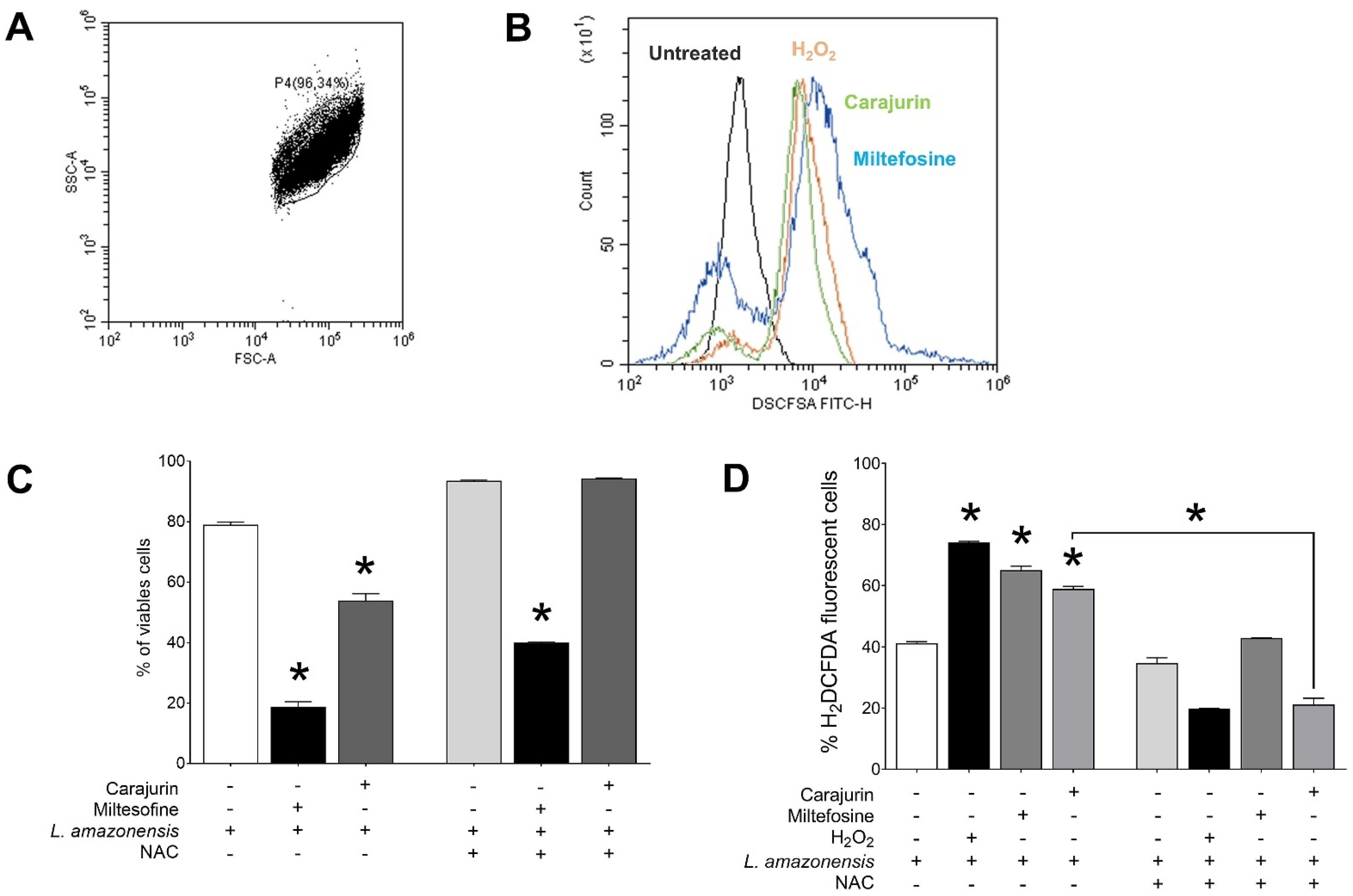
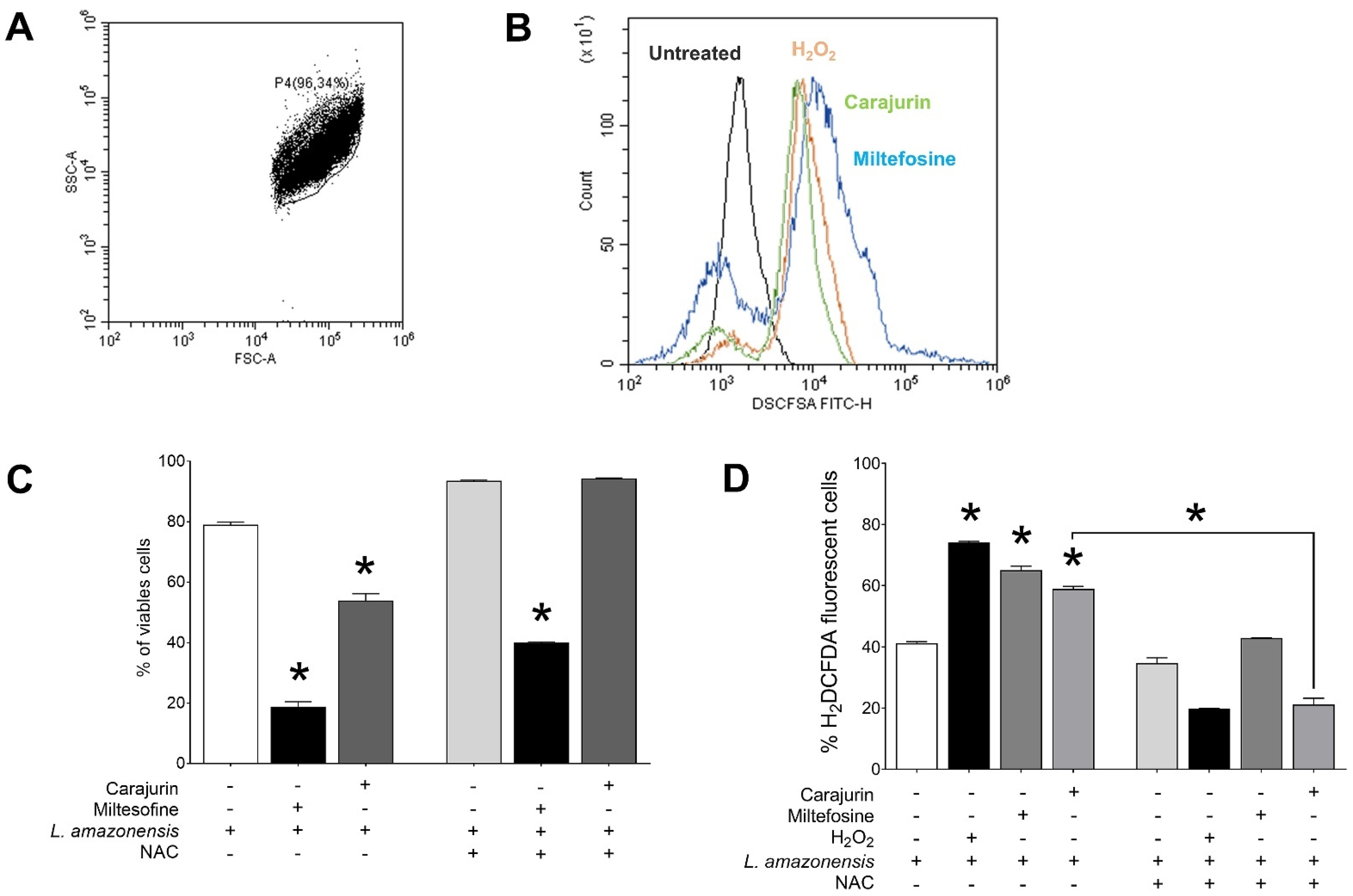
2.5. Measurement of ROS
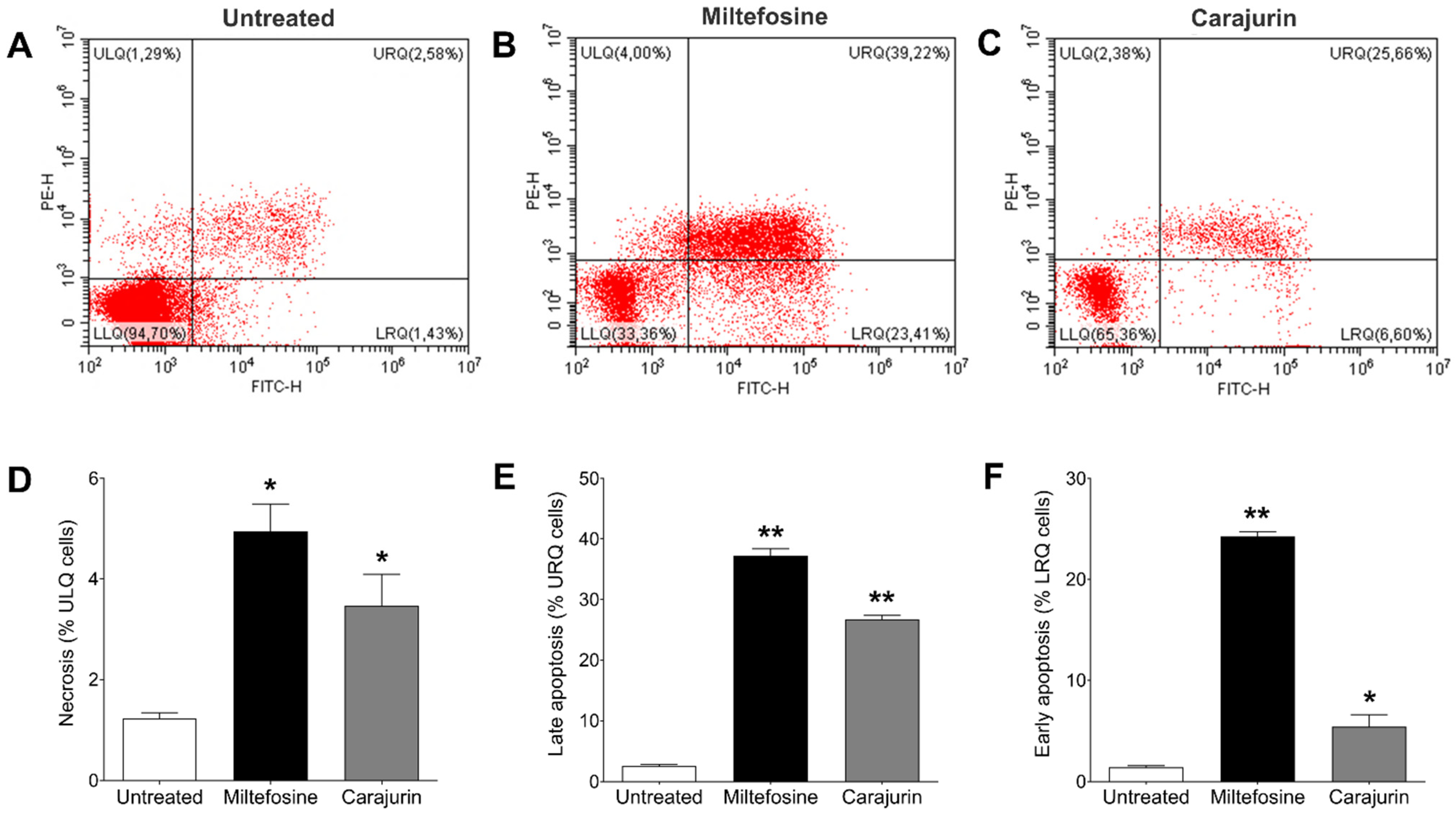
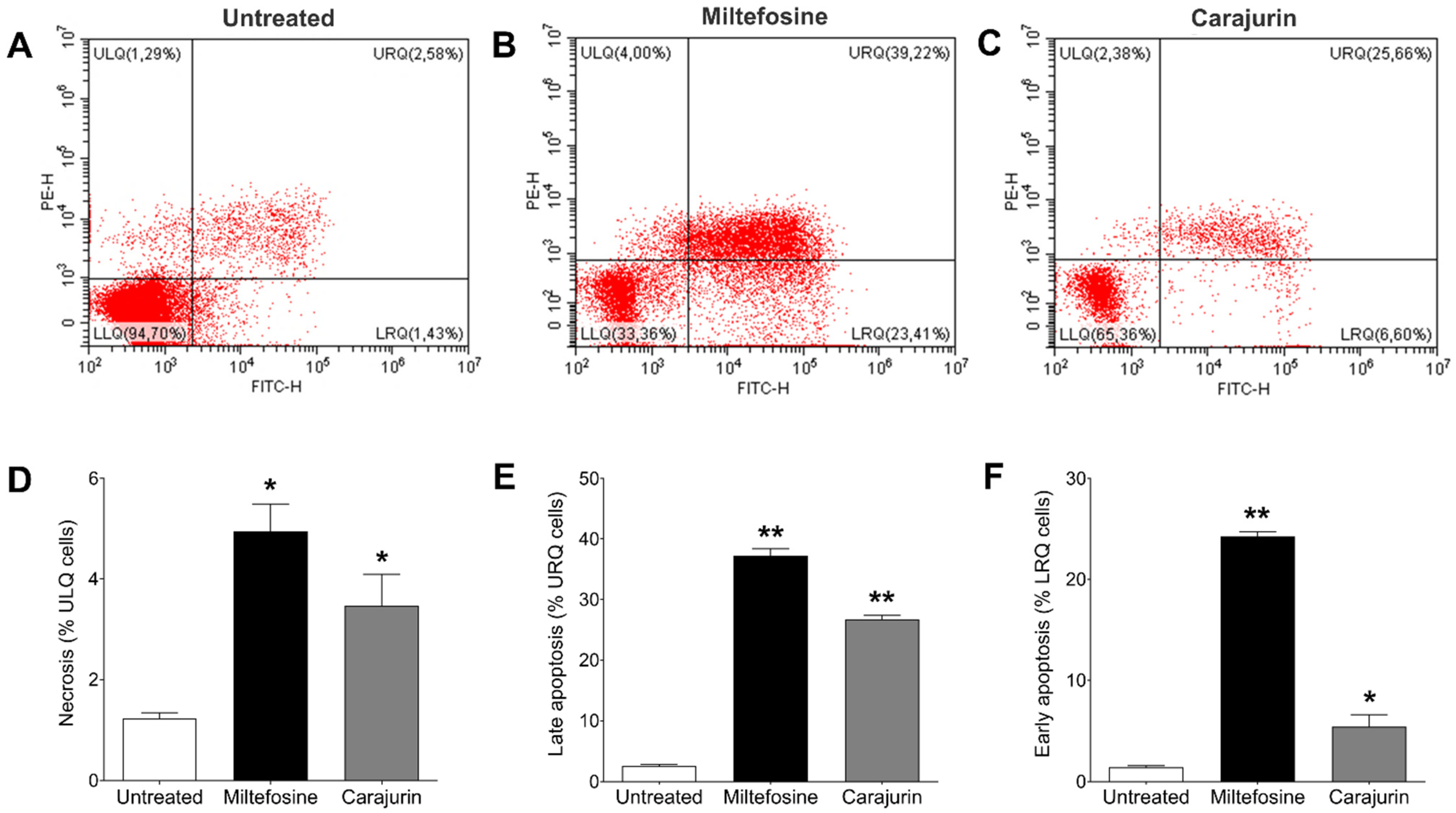
2.6. Evaluation of Phosphatidylserine Exposure and Cell Membrane Integrity
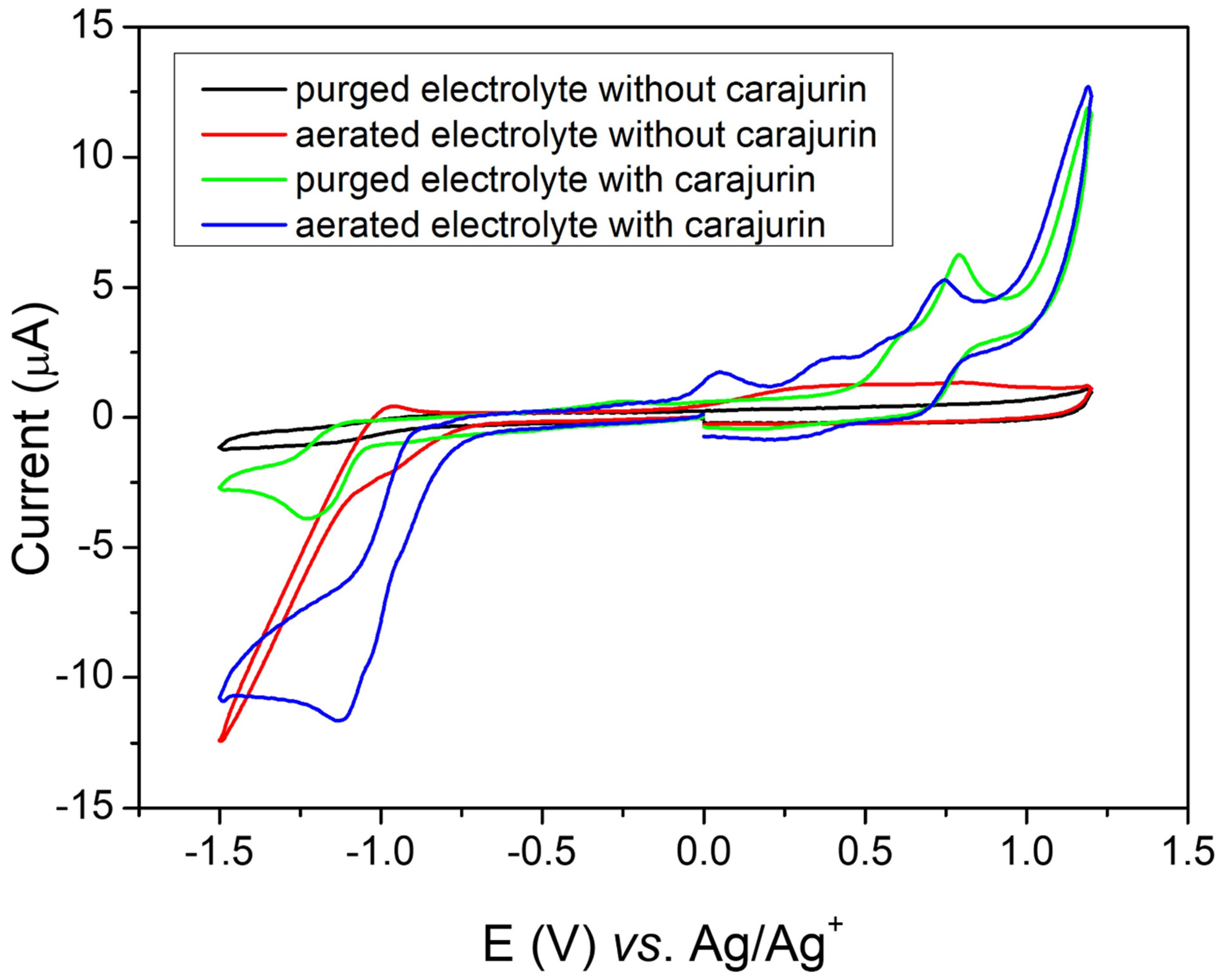
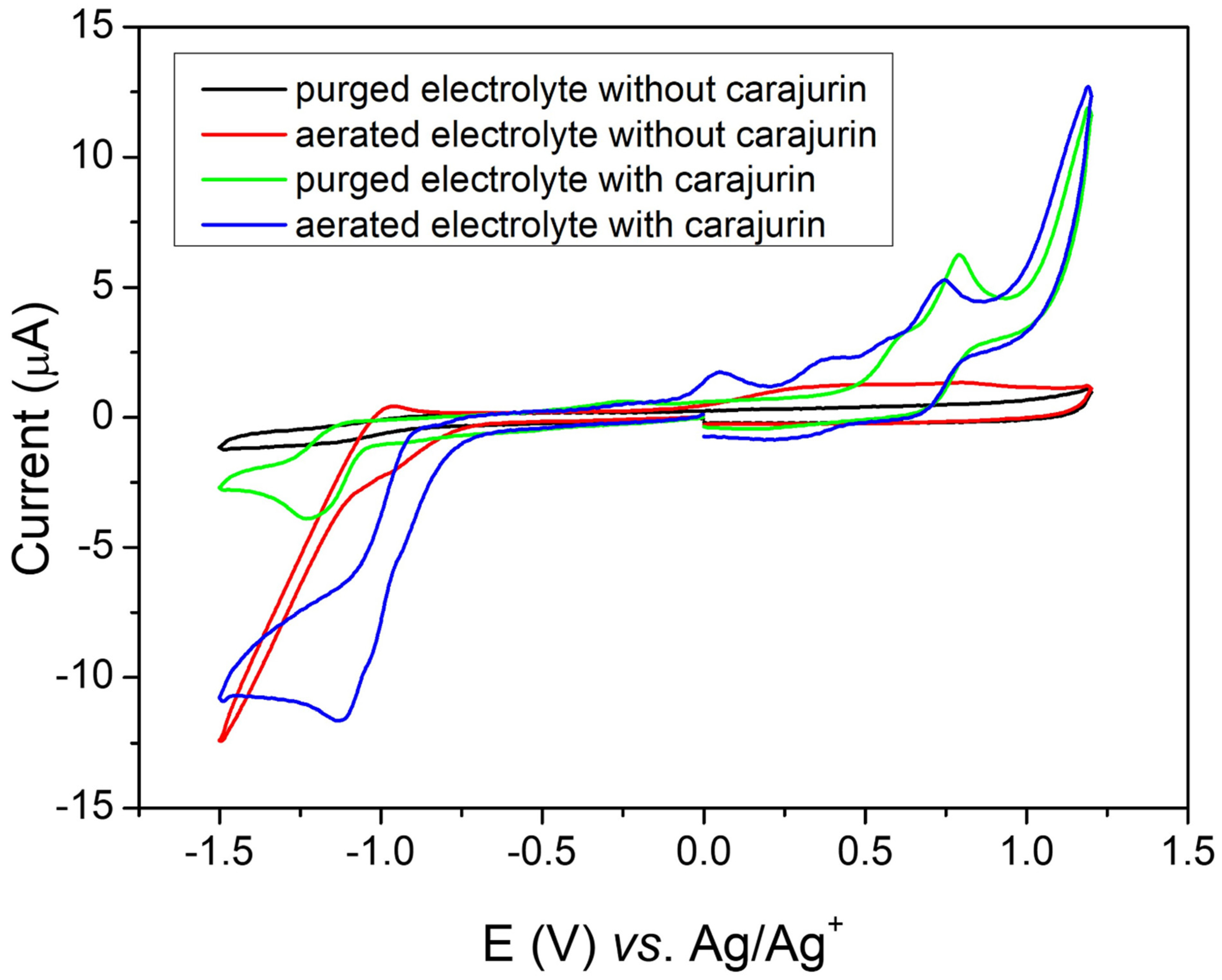
2.7. Electrochemical Tests/Cyclic Voltammetry
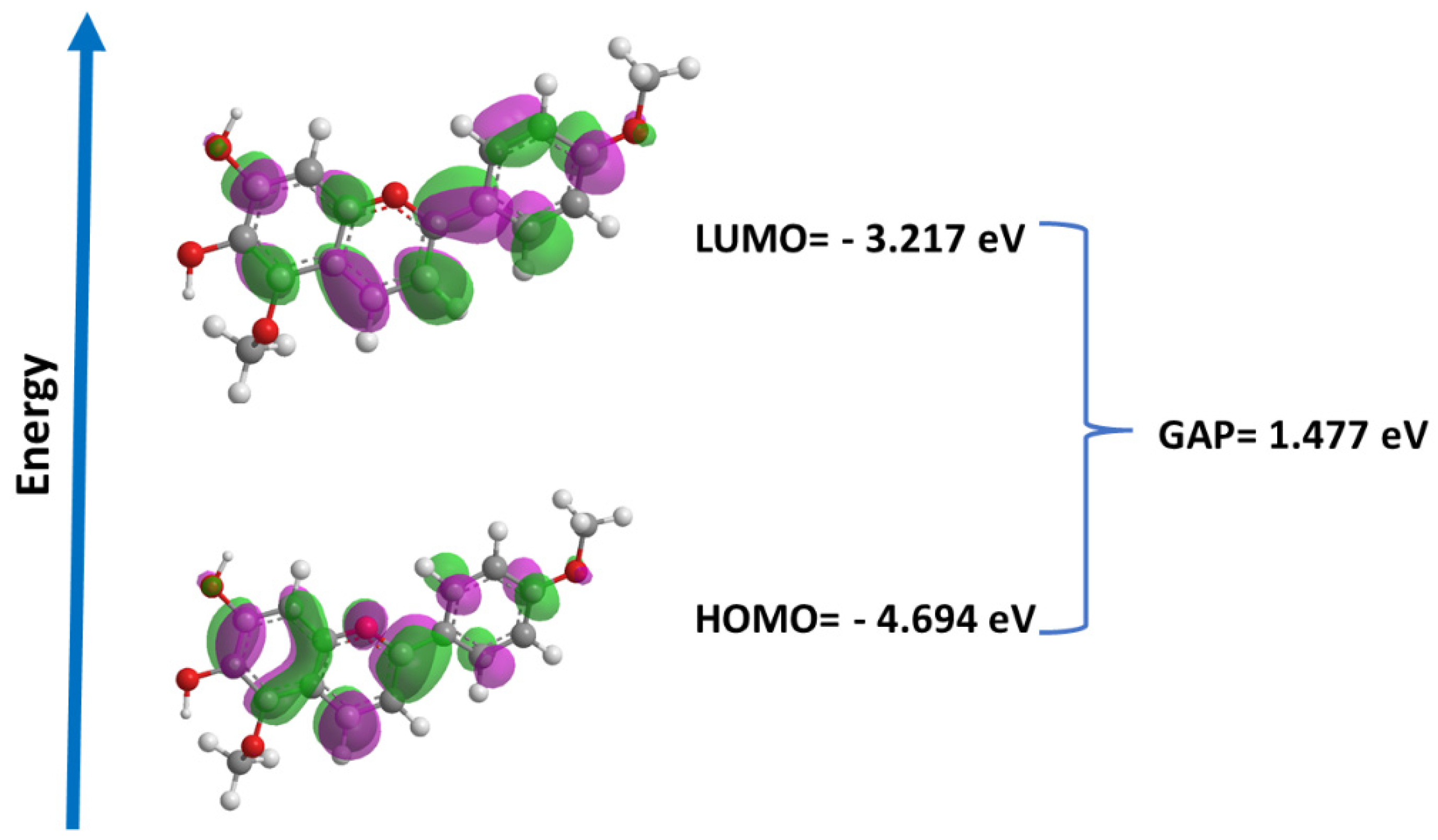
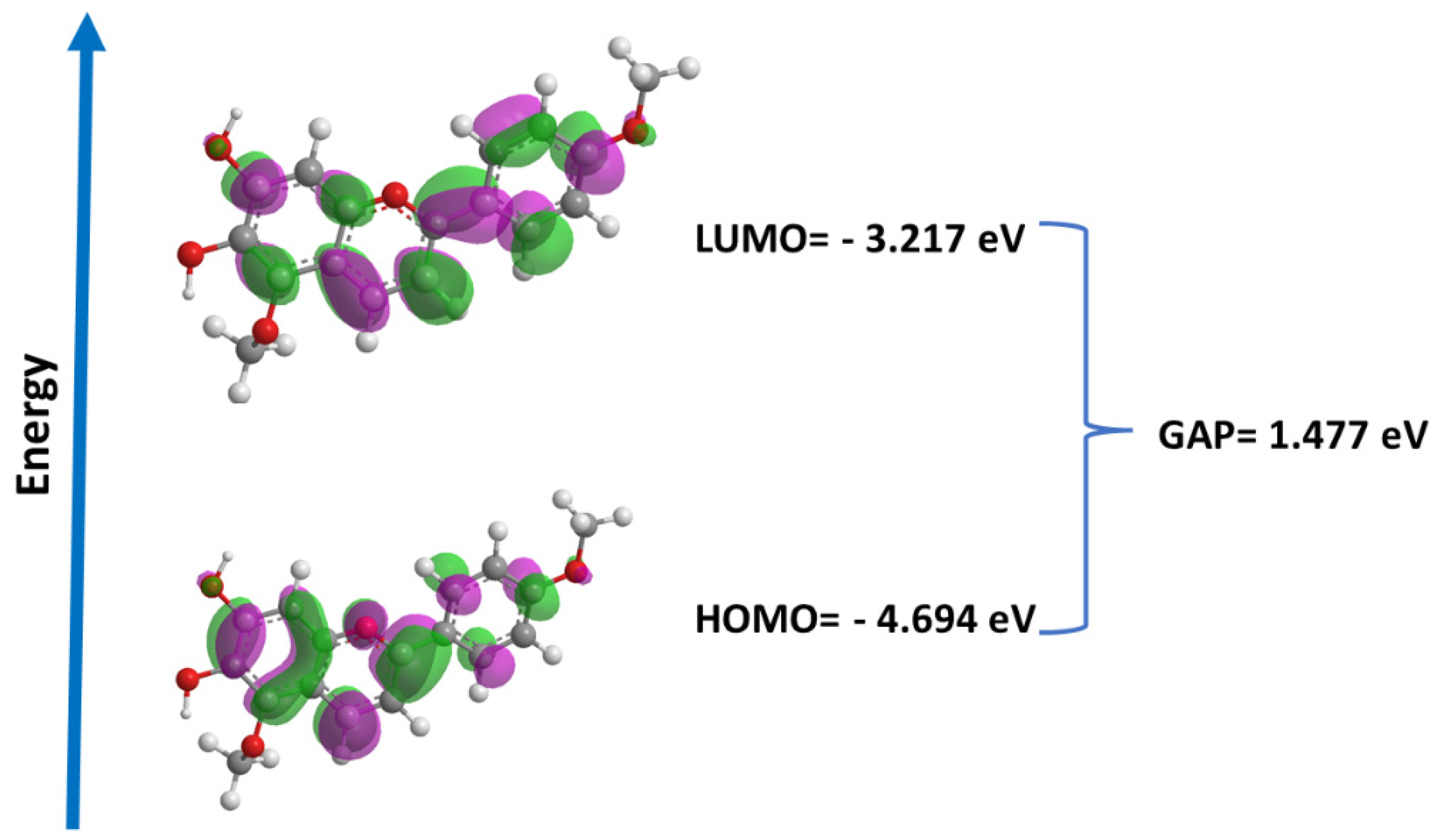
2.8. Quantum Studies
2.9. In Silico Prediction Physico-Chemical
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Plant Material
4.3. Isolation and Structural Characterization of Carajurin
4.4. Ethical Statements and Animals
4.5. Peritoneal Macrophage Isolation and Parasite Cultures
4.6. In Vitro Cytotoxicity Assay of Carajurin on Peritoneal Macrophages (CC50)
4.7. In Vitro Inhibition Assay of Carajurin on Promastigotes (IC50) and Selectivity Index
4.8. Transmission Electron Microscopy
4.9. Determination of Mitochondrial Membrane Potential (MMP)(ΔΨm)
4.10. Measurement of Reactive Oxygen Species (ROS)
4.11. Detection of L. amazonensis Apoptosis by Flow Cytometry
4.12. Electrochemical Tests/Cyclic Voltammetry
4.13. Quantum Studies
4.14. In Silico Prediction Physico-Chemical
4.15. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Leishmaniasis. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 9 February 2022).
- Rezvan, H.; Moafi, M.; Sherkat, R.; Taleban, R. Leishmania vaccines entered in clinical trials: A review of literature. Int. J. Prev. Med. 2019, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Alcântara, L.M.; Ferreira, T.C.S.; Gadelha, F.R.; Miguel, D.C. Challenges in drug discovery targeting TriTryp diseases with an emphasis on leishmaniasis. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Mathew, N.S.; Negi, P.S. Plant-Derived Compounds against Protozoan Neglected Diseases: Toward Sustainable Drug De-velopment. In Discovery and Development of Therapeutics from Natural Products against Neglected Tropical Diseases; Elsevier: Amsterdam, The Netherlands, 2019; pp. 241–292. [Google Scholar]
- Raj, S.; Sasidharan, S.; Balaji, S.N.; Dubey, V.K.; Saudagar, P. Review on natural products as an alternative to contemporary anti-leishmanial therapeutics. J. Proteins Proteom. 2020, 11, 135–158. [Google Scholar] [CrossRef]
- Fonseca-Silva, F.; Inacio, J.D.F.; Canto-Cavalheiro, M.M.; Almeida-Amaral, E.E. Reactive Oxygen Species Production and Mitochondrial Dysfunction Contribute to Quercetin Induced Death in Leishmania amazonensis. PLoS ONE 2011, 6, e14666. [Google Scholar] [CrossRef] [PubMed]
- Menna-Barreto, R.F.S.; De Castro, S.L. The Double-Edged Sword in Pathogenic Trypanosomatids: The Pivotal Role of Mitochondria in Oxidative Stress and Bioenergetics. BioMed Res. Int. 2014, 2014, 614014. [Google Scholar] [CrossRef] [Green Version]
- Mondêgo-Oliveira, R.; de Sá Sousa, J.C.; Moragas-Tellis, C.J.; de Souza, P.V.R.; dos Santos Chagas, M.D.S.D.S.; Behrens, M.D.; Jesús Hardoim, D.D.J.; Taniwaki, N.N.; Chometon, T.Q.; Bertho, A.L.; et al. Vernonia brasiliana (L.) Druce induces ultrastructural changes and apoptosis-like death of Leishmania infantum promastigotes. Biomed. Pharmacother. 2021, 133, 111025. [Google Scholar] [CrossRef]
- Mafioleti, L.; da Silva Junior, I.F.; Colodel, E.M.; Flach, A.; Martins, D.T.D.O. Evaluation of the toxicity and antimicrobial activity of hydroethanolic extract of Arrabidaea chica (Humb. & Bonpl.) B. Verl. J. Ethnopharmacol. 2013, 150, 576–582. [Google Scholar] [CrossRef]
- Jorge, M.P.; Madjarof, C.; Ruiz, A.L.T.G.; Fernandes, A.T.; Rodrigues, R.A.F.; de Oliveira Sousa, I.M.; Foglio, M.A.; de Carvalho, J.E. Evaluation of wound healing properties of Arrabidaea chica Verlot extract. J. Ethnopharmacol. 2008, 118, 361–366. [Google Scholar] [CrossRef]
- Moragas-Tellis, C.J.; Almeida-Souza, F.; do Socorro dos Santos Chagas, M.; de Souza, P.V.R.; Silva-Silva, J.V.; Ramos, Y.J.; Moreira, D.; Calabrese, K.; Behrens, M. The Influence of Anthocyanidin Profile on Antileishmanial Activity of Arrabidaea chica Morphotypes. Molecules 2020, 25, 3547. [Google Scholar] [CrossRef]
- Silva-Silva, J.V.; Moragas-Tellis, C.J.; Chagas, M.S.S.; Souza, P.V.R.; Moreira, D.L.; de Souza, C.S.F.; Teixeira, K.F.; Cenci, A.R.; de Oliveira, A.S.; Almeida-Souza, F.; et al. Carajurin: A anthocyanidin from Arrabidaea chica as a potential biological marker of antileishmanial activity. Biomed. Pharmacother. 2021, 141, 111910. [Google Scholar] [CrossRef]
- Chapman, E.; Perkin, A.G.; Robinson, R. CCCCII.—The colouring matters of carajura. J. Chem. Soc. 1927, 3015–3041. [Google Scholar] [CrossRef]
- Zorn, B.; García-Piñeres, A.J.; Castro, V.; Murillo, R.; Mora, G.; Merfort, I. 3-Desoxyanthocyanidins from Arrabidaea chica. Phytochemistry 2001, 56, 831–835. [Google Scholar] [CrossRef]
- Devia, B.; Llabres, G.; Wouters, J.; Dupont, L.; Escribano-Bailon, M.T.; de Pascual-Teresa, S.; Angenot, L.; Tits, M. New 3-deoxyanthocyanidins from leaves ofArrabidaea chica. Phytochem. Anal. 2002, 13, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Rottini, M.M.; Amaral, A.C.F.; Ferreira, J.L.P.; Oliveira, E.S.C.; de Andrade Silva, J.R.; Taniwaki, N.N.; dos Santos, A.R.; Al-meida-Souza, F.; da Silva Freitas de Souza, C.; da Silva Calabrese, K. Endlicheria bracteolata (Meisn.) Essential Oil as a Weapon Against Leishmania amazonensis: In Vitro Assay. Molecules 2019, 24, 2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolić, M.D.; Pavlović, A.N.; Mitić, S.S.; Tošić, S.B.; Mitić, M.N.; Kaličanin, B.M.; Manojlović, D.D.; Stanković, D.M. Use of cyclic voltammetry to determine the antioxidant capacity of berry fruits: Correlation with spectrophotometric assays. Eur. J. Hortic. Sci. 2019, 84, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Silva, T.L.; Ferreira, F.R.; De Vasconcelos, C.C.; da Silva, R.C.; Lima, D.J.D.P.; Costa, P.R.R.; Netto, C.D.; Goulart, M.O.F. Reactive Oxygen Species Release, Alkylating Ability, and DNA Interactions of a Pterocarpanquinone: A Test Case for Electrochemistry. ChemElectroChem 2016, 3, 2252–2263. [Google Scholar] [CrossRef]
- Silva, T.; Da Silva, J.; Lima, D.; Ferreira, F.R.; De Vasconcelos, C.; Santos, D.; Netto, C.D.; Costa, P.; Goulart, M. Medicinal Electrochemistry of Halogenated and Nitrated Pterocarpanquinones. J. Braz. Chem. Soc. 2019, 30, 2438–2451. [Google Scholar] [CrossRef]
- Rodrigues, I.A.; Azevedo, M.M.B.; Chaves, F.C.M.; Alviano, C.S.; Alviano, D.S.; Vermelho, A.B. Arrabidaea chica Hexanic Extract Induces Mitochondrion Damage and Peptidase Inhibition on Leishmania spp. BioMed Res. Int. 2014, 2014, 985171. [Google Scholar] [CrossRef] [Green Version]
- Cortez De Sá, J.; Almeida-Souza, F.; Mondêgo-Oliveira, R.; Oliveira, I.D.S.D.S.; Lamarck, L.; Magalhães, I.D.F.B.; Ataídes-Lima, A.F.; Ferreira, H.D.S.; Abreu-Silva, A.L. Leishmanicidal, cytotoxicity and wound healing potential of Arrabidaea chica Verlot. BMC Complement. Altern. Med. 2015, 16, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Silva, J.V.; Moragas-Tellis, C.J.; Chagas, M.D.S.D.S.; de Souza, P.V.R.; Souza, C.D.S.F.D.; Hardoim, D.D.J.; Taniwaki, N.N.; Moreira, D.D.L.; Dutra Behrens, M.D.; Calabrese, K.D.S.; et al. Antileishmanial Activity of Flavones-Rich Fraction from Arrabidaea chica Verlot (Bignoniaceae). Front. Pharmacol. 2021, 12, 703985. [Google Scholar] [CrossRef]
- Weniger, B.; Robledo, S.; Arango, G.J.; Deharo, E.; Aragón, R.; Muñoz, V.; Callapa, J.; Lobstein, A.; Anton, R. Antiprotozoal activities of Colombian plants. J. Ethnopharmacol. 2001, 78, 193–200. [Google Scholar] [CrossRef]
- Kim, I.; Rodriguez-Enriquez, S.; Lemasters, J.J. Selective degradation of mitochondria by mitophagy. Arch. Biochem. Biophys. 2007, 462, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittra, B.; Saha, A.; Roy Chowdhury, A.; Pal, C.; Mandal, S.; Mukhopadhyay, S.; Bandyopadhyay, S.; Majumder, H.K. Luteolin, an Abundant Dietary Component is a Potent Anti-leishmanial Agent that Acts by Inducing Topoisomerase II-mediated Kinetoplast DNA Cleavage Leading to Apoptosis. Mol. Med. 2000, 6, 527–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, V.P.C.; Da Rocha, C.Q.; Queiroz, E.F.; Marcourt, L.; Vilegas, W.; Grimaldi, G.B.; Furrer, P.; Allémann, É.; Wolfender, J.-L.; Soares, M.B.P. Antileishmanial Activity of Dimeric Flavonoids Isolated from Arrabidaea brachypoda. Molecules 2018, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes Rodrigues, J.; Souza, W. Ultrastructural Alterations in Organelles of Parasitic Protozoa Induced by Different Classes of Metabolic Inhibitors. Curr. Pharm. Des. 2008, 14, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Majumder, H. Mitochondrion of Protozoan Parasite Emerges as Potent Therapeutic Target:Exciting Drugs are on the Horizon. Curr. Pharm. Des. 2008, 14, 839–846. [Google Scholar] [CrossRef]
- Fidalgo, L.M.; Gille, L. Mitochondria and Trypanosomatids: Targets and Drugs. Pharm. Res. 2011, 28, 2758–2770. [Google Scholar] [CrossRef] [PubMed]
- Miranda, N.; Gerola, A.P.; Novello, C.R.; Ueda-Nakamura, T.; de Oliveira Silva, S.; Dias-Filho, B.P.; Hioka, N.; de Mello, J.C.P.; Nakamura, C.V. Pheophorbide a, a compound isolated from the leaves of Arrabidaea chica, induces photodynamic inactivation of Trypanosoma cruzi. Photodiagn. Photodyn. Ther. 2017, 19, 256–265. [Google Scholar] [CrossRef]
- Gervazoni, L.F.O.; Barcellos, G.B.; Ferreira-Paes, T.; Almeida-Amaral, E.E. Use of Natural Products in Leishmaniasis Chemotherapy: An Overview. Front. Chem. 2020, 8, 8. [Google Scholar] [CrossRef]
- Roy, A.; Ganguly, A.; BoseDasgupta, S.; Das, B.B.; Pal, C.; Jaisankar, P.; Majumder, H.K. Mitochondria-Dependent Reactive Oxygen Species-Mediated Programmed Cell Death Induced by 3,3′-Diindolylmethane through Inhibition of F0F1-ATP Synthase in Unicellular Protozoan ParasiteLeishmania donovani. Mol. Pharmacol. 2008, 74, 1292–1307. [Google Scholar] [CrossRef]
- Smirlis, D.; Duszenko, M.; Ruiz, A.J.; Scoulica, E.; Bastien, P.; Fasel, N.; Soteriadou, K. Targeting essential pathways in trypanosomatids gives insights into protozoan mechanisms of cell death. Parasites Vectors 2010, 3, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, M.C.; Marques, F.C.S.; Blazius, R.D.; Santos Da Silva, O.; de Queiroz Cunha, F.; Bento, D.B.; Torres Romão, P.R. N-acetyl-l-cysteine reduces the parasitism of BALB/c mice infected with Leishmania amazonensis. Parasitol. Res. 2007, 102, 801–803. [Google Scholar] [CrossRef] [PubMed]
- Castro-Pinto, D.B.; Silva Lima, E.L.; Cunha, A.S.; Genestra, M.; De Léo, R.M.; Monteiro, F.; Leon, L.L. Leishmania amazonensis trypanothione reductase: Evaluation of the effect of glutathione analogs on parasite growth, infectivity and enzyme activity. J. Enzym. Inhib. Med. Chem. 2007, 22, 71–75. [Google Scholar] [CrossRef]
- Fonseca-Silva, F.; Canto-Cavalheiro, M.M.; Menna-Barreto, R.F.S.; Almeida-Amaral, E.E. Effect of Apigenin on Leishmania amazonensis Is Associated with Reactive Oxygen Species Production Followed by Mitochondrial Dysfunction. J. Nat. Prod. 2015, 78, 880–884. [Google Scholar] [CrossRef]
- Mehta, A.; Shaha, C. Apoptotic Death in Leishmania donovani Promastigotes in Response to Respiratory Chain Inhibition. J. Biol. Chem. 2004, 279, 11798–11813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera-Serra, M.G.; Lorenzo-Morales, J.; Romero, M.; Valladares, B.; Piñero, J.E. In vitro activity of perifosine: A novel alkylphospholipid against the promastigote stage of Leishmania species. Parasitol. Res. 2007, 100, 1155–1157. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.P.; Lazarin-Bidóia, D.; Ueda-Nakamura, T.; Silva, S.D.O.; Nakamura, C.V. Eupomatenoid-5 Isolated from Leaves ofPiper regnelliiInduces Apoptosis inLeishmania amazonensis. Evid. Based Complement. Altern. Med. 2013, 2013, 940531. [Google Scholar] [CrossRef] [Green Version]
- Zeouk, I.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; Bethencourt-Estrella, C.J.; Bazzocchi, I.L.; Bekhti, K.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Sesquiterpenoids and flavonoids from Inula viscosa induce programmed cell death in kinetoplastids. Biomed. Pharmacother. 2020, 130, 110518. [Google Scholar] [CrossRef]
- Manjolin, L.C.; dos Reis, M.B.G.; Maquiaveli, C.D.C.; Santos-Filho, O.A.; da Silva, E.R. Dietary flavonoids fisetin, luteolin and their derived compounds inhibit arginase, a central enzyme in Leishmania (Leishmania) amazonensis infection. Food Chem. 2013, 141, 2253–2262. [Google Scholar] [CrossRef] [Green Version]
- da Silva, E.R.; Brogi, S.; Lucon-Júnior, J.F.; Campiani, G.; Gemma, S.; Maquiaveli, C.D.C. Dietary polyphenols rutin, taxifolin and quercetin related compounds target Leishmania amazonensis arginase. Food Funct. 2019, 10, 3172–3180. [Google Scholar] [CrossRef]
- Mandal, A.; Das, S.; Roy, S.; Ghosh, A.K.; Sardar, A.H.; Verma, S.; Saini, S.; Singh, R.; Abhishek, K.; Kumar, A.; et al. Deprivation of L-Arginine Induces Oxidative Stress Mediated Apoptosis in Leishmania donovani Promastigotes: Contribution of the Polyamine Pathway. PLoS Negl. Trop. Dis. 2016, 10, e0004373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bombaça, A.C.S.; Viana, P.G.; Santos, A.C.C.; da Silva, T.L.; Rodrigues, A.B.M.; Guimarães, A.C.R.; Goulart, M.O.F.; da SilvaJúnior, E.N.; Menna-Barreto, R.F.S. Mitochondrial disfunction and ROS production are essential for anti-Trypanosoma cruzi activity of β-lapachone-derived naphthoimidazoles. Free Radic. Biol. Med. 2019, 130, 408–418. [Google Scholar] [CrossRef] [PubMed]
- de Paiva, Y.G.; Pinho Júnior, W.; de Souza, A.A.; Costa, C.O.; Silva, F.P.L.; Lima-Junior, C.G.; Vasconcellos, M.L.A.A.; Goulart, M.O.F. Electrochemical and computational studies, in protic medium, of Morita-Baylis-Hillman adducts and correlation with leishmanicidal activity. Electrochim. Acta 2014, 140, 557–563. [Google Scholar] [CrossRef]
- Justino, G.C.; Vieira, A.J.S.C. Antioxidant mechanisms of Quercetin and Myricetin in the gas phase and in solution—A comparison and validation of semi-empirical methods. J. Mol. Model. 2010, 16, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, I.D.S.D.S.; Moragas-Tellis, C.J.; Chagas, M.D.S.D.S.; Behrens, M.D.; Calabrese, K.D.S.; Abreu-Silva, A.L.; Almeida-Souza, F. Carapa guianensis Aublet (Andiroba) Seed Oil: Chemical Composition and Antileishmanial Activity of Limonoid-Rich Fractions. BioMed Res. Int. 2018, 2018, 5032816. [Google Scholar] [CrossRef] [Green Version]
- Cardona, C.M.; Li, W.; Kaifer, A.E.; Stockdale, D.; Bazan, G.C. Electrochemical Considerations for Determining Absolute Frontier Orbital Energy Levels of Conjugated Polymers for Solar Cell Applications. Adv. Mater. 2011, 23, 2367–2371. [Google Scholar] [CrossRef]
- Hückel, E. Quantentheoretische Beiträge zum Benzolproblem. Z. Physik 1931, 70, 204–286. [Google Scholar] [CrossRef]
- Roothaan, C.C.J. Self-Consistent Field Theory for Open Shells of Electronic Systems. Rev. Mod. Phys. 1960, 32, 179–185. [Google Scholar] [CrossRef]
- de Oliveira, A.S.; Llanes, L.C.; Nunes, R.J.; Nucci-Martins, C.; de Souza, A.S.; Palomino-Salcedo, D.L.; Dávila-Rodríguez, M.J.; Ferreira, L.L.G.; Santos, A.R.S.; Andricopulo, A.D. Antioxidant Activity, Molecular Docking, Quantum Studies and In Vivo Antinociceptive Activity of Sulfonamides Derived from Carvacrol. Front. Pharmacol. 2021, 12, 788850. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1H a,b 400 MHz (δ) in ppm (CDCl3) | Cosy (H/h Correlation) | 13C b 100 MHz (δ) in ppm (CDCl3) | DEPT | HSQC (H/C Correlation) | Hmbc (H/C Correlation) |
|---|---|---|---|---|---|---|
| 2 | - | 158.90 | Q | - | - | |
| 3 | 6.98 (d, J = 19.5 Hz) | H4 | 102.62 | CH | C3 | C10; C1′; C2 |
| 4 | 7.99 (d, J = 19.5 Hz) | H3 | 133.76 | CH | C4 | C5; C2; C9 |
| 5 | - | - | 135.02 | Q | - | - |
| 6 | - | - | 139.93 | Q | - | - |
| 7 | - | - | 176.82 | Q | - | - |
| 8 | 6.53 (s) | - | 98.61 | CH | C8 | C10; C6; C9 |
| 9 | - | - | 156.86 | Q | - | - |
| 10 | - | - | 118.16 | Q | - | - |
| 1′ | - | - | 123.43 | Q | - | - |
| 2′/6′ | 7.89 (d) J = 22.3 Hz | H3′e H5′ | 127.68 | CH | C2′/C6′ | C2′; C6′; C2, C4′ |
| 3′/5′ | 7.01 (d) J = 22.2 Hz | H2′e H6′ | 114.77 | CH | C3′/C5′ | C3′; C5′; C1′, C4′ |
| 4′ | - | - | 162.50 | Q | - | - |
| OCH3-5 | 4.10 (s) | - | 60.42 | CH3 | OCH3-5 | C5 |
| OCH3-4` | 3.90 (s) | - | 55.58 | CH3 | OCH3-4′ | C4′ |
| Compounds | Peritoneal Macrophages | L. amazonensis Promastigotes | |
|---|---|---|---|
| CC50 (μg mL−1) | IC50 (μg mL−1) | SI | |
| Carajurin | 258.2 ± 1.20 (856.9 µM) | 7.96 ± 1.23 (26.42 µM) | 32.4 |
| Amphotericin B | 8.740 ± 1.08 (9.458 µM) | 0.0299 ± 1.18 (0.03236 µM) | 292.3 |
| Property/Model Name | Carajurin |
|---|---|
| Physico-chemical | |
| Molecular weight | 301.31 |
| # Rotatable bonds | 3 |
| # H-bond acceptors | 5 |
| # H-bond donors | 2 |
| Surface area | 126.520 |
| TPSA (Å2) | 75.99 |
| Lipophilicity (log Po/w) | 0.68 |
| Drug-likeness | |
| Lipinski | Yes; 0 violation |
| Ghose | Yes |
| Veber | Yes |
| Egan | Yes |
| Muegge | Yes |
| Medicinal chemistry | |
| PAINS | 0 alert |
| Brenk | 1 alert: charged oxygen sulfur |
| Lead-likeness | Yes |
| Syntheticaccessibility | 4.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Silva, J.V.; Moragas-Tellis, C.J.; Chagas, M.S.S.; Souza, P.V.R.; Moreira, D.L.; Hardoim, D.J.; Taniwaki, N.N.; Costa, V.F.A.; Bertho, A.L.; Brondani, D.; et al. Carajurin Induces Apoptosis in Leishmania amazonensis Promastigotes through Reactive Oxygen Species Production and Mitochondrial Dysfunction. Pharmaceuticals 2022, 15, 331. https://doi.org/10.3390/ph15030331
Silva-Silva JV, Moragas-Tellis CJ, Chagas MSS, Souza PVR, Moreira DL, Hardoim DJ, Taniwaki NN, Costa VFA, Bertho AL, Brondani D, et al. Carajurin Induces Apoptosis in Leishmania amazonensis Promastigotes through Reactive Oxygen Species Production and Mitochondrial Dysfunction. Pharmaceuticals. 2022; 15(3):331. https://doi.org/10.3390/ph15030331
Chicago/Turabian StyleSilva-Silva, João Victor, Carla J. Moragas-Tellis, Maria S. S. Chagas, Paulo Victor R. Souza, Davyson L. Moreira, Daiana J. Hardoim, Noemi N. Taniwaki, Vanessa F. A. Costa, Alvaro L. Bertho, Daniela Brondani, and et al. 2022. "Carajurin Induces Apoptosis in Leishmania amazonensis Promastigotes through Reactive Oxygen Species Production and Mitochondrial Dysfunction" Pharmaceuticals 15, no. 3: 331. https://doi.org/10.3390/ph15030331
APA StyleSilva-Silva, J. V., Moragas-Tellis, C. J., Chagas, M. S. S., Souza, P. V. R., Moreira, D. L., Hardoim, D. J., Taniwaki, N. N., Costa, V. F. A., Bertho, A. L., Brondani, D., Zapp, E., de Oliveira, A. S., Calabrese, K. S., Behrens, M. D., & Almeida-Souza, F. (2022). Carajurin Induces Apoptosis in Leishmania amazonensis Promastigotes through Reactive Oxygen Species Production and Mitochondrial Dysfunction. Pharmaceuticals, 15(3), 331. https://doi.org/10.3390/ph15030331