
Antiviral Effects of Artemisinin and Its Derivatives against SARS-CoV-2 Main Protease: Computational Evidences and Interactions with ACE2 Allelic Variants

 ,
,  ,
,  , ,
, ,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
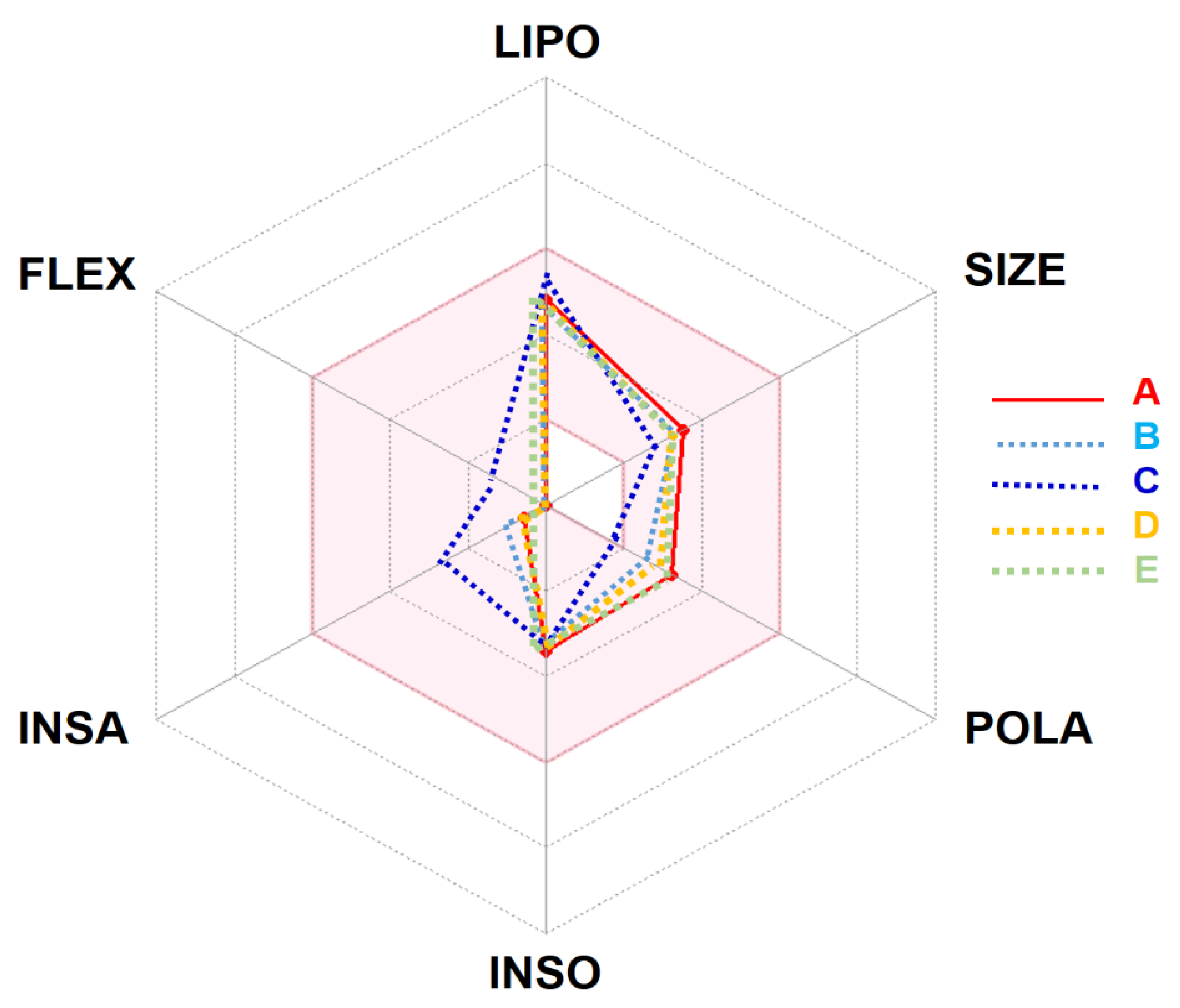
4.1. In Silico ADMET and Pharmacokinetic Profiles
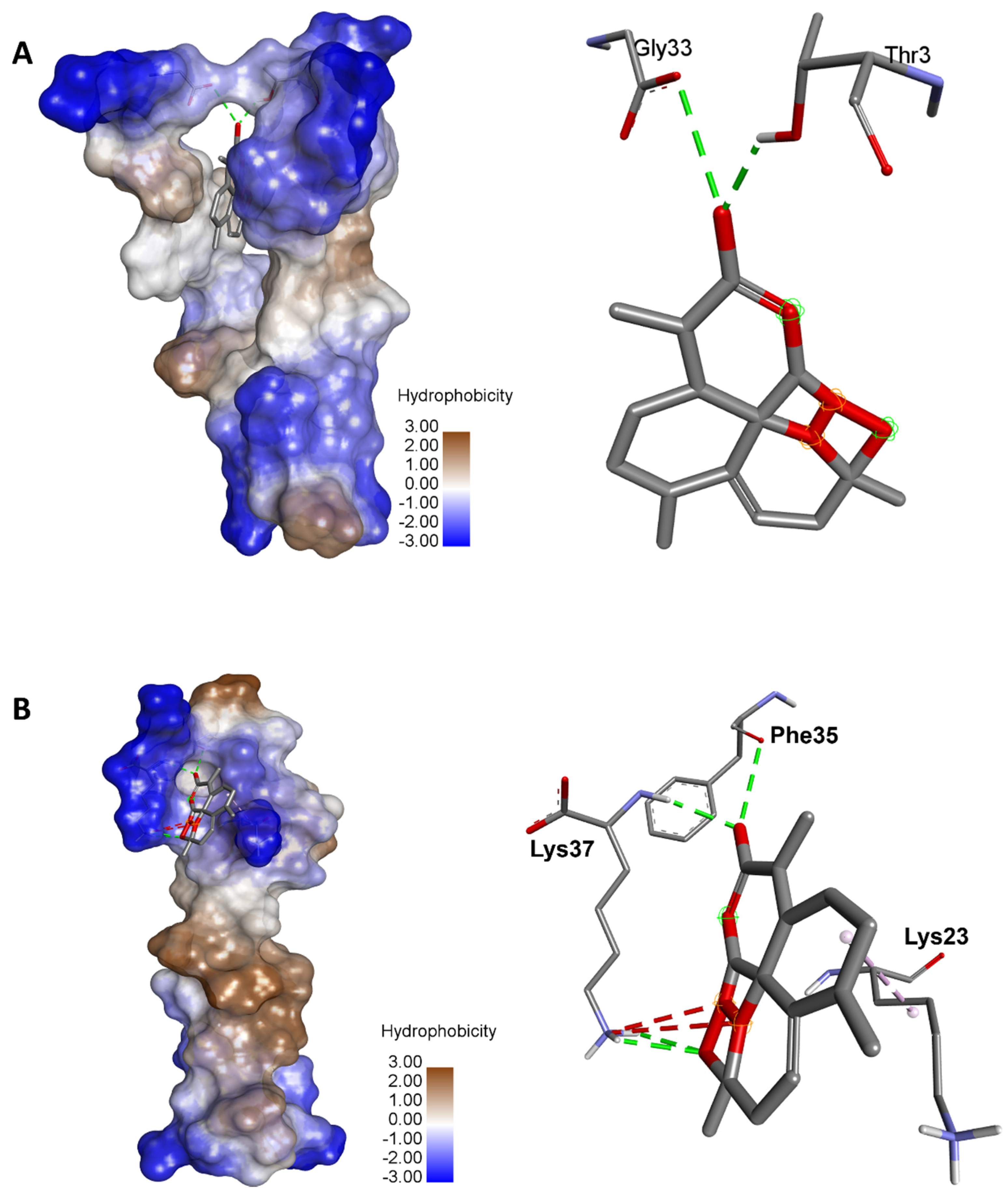
4.2. Genetic Variants of Human ACE2
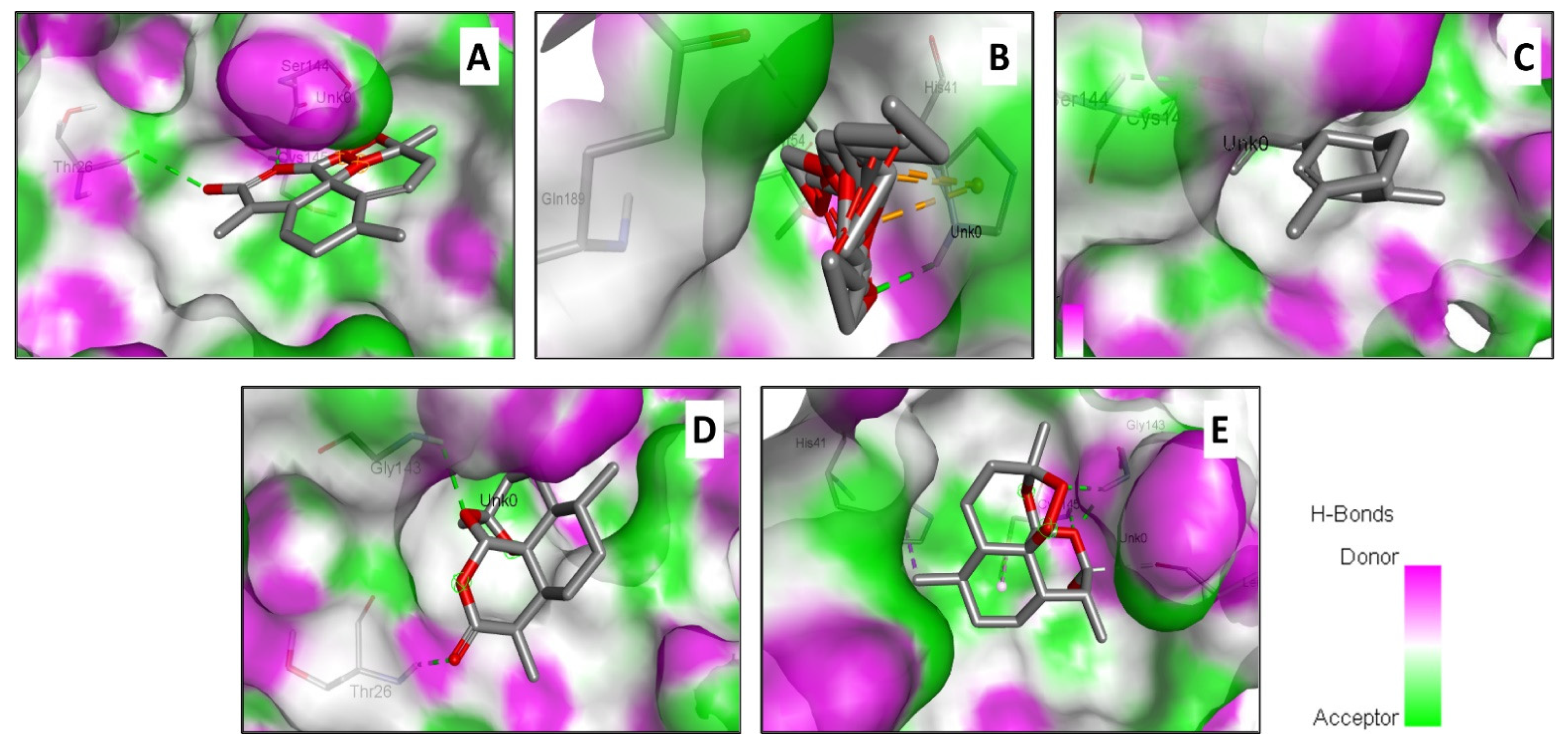
4.3. Molecular Binding and Interactions
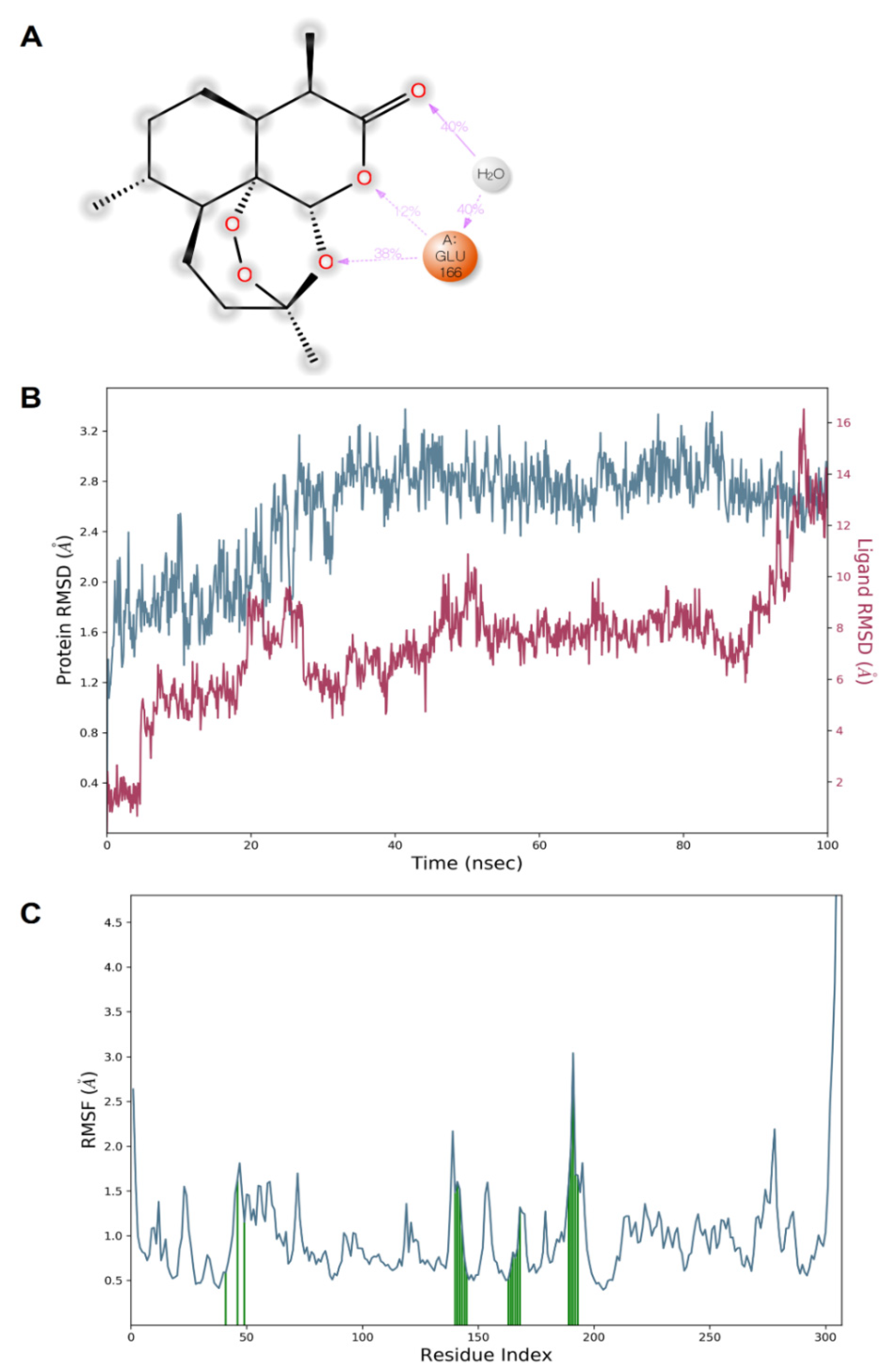
4.4. Molecular Dynamics (MD) Simulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Badraoui, R.; Alrashedi, M.M.; El-May, M.V.; Bardakci, F. Acute respiratory distress syndrome: A life threatening associated complication of SARS-CoV-2 infection inducing COVID-19. J. Biomol. Struct. Dyn. 2021, 39, 6842–6851. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Novel coronavirus (COVID-19) outbreak: Hope and search for effective therapeutic agent. Lett. Drug Des. Discov. 2020, 17, 940–942. [Google Scholar] [CrossRef]
- Sivaraman, D.; Pradeep, P.; Manoharan, S.S.; Bhat, C.R.; Leela, K.; Venugopal, V. Current Strategies and Approaches in Combating SARS-CoV-2 Virus that Causes COVID-19. Lett. Drug Des. Discov. 2020, 17, 672–674. [Google Scholar] [CrossRef]
- van Riel, D.; de Wit, E. Next-generation vaccine platforms for COVID-19. Nat. Mater. 2020, 19, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Le, T.T.; Andreadakis, Z.; Kumar, A.; Román, R.G.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef] [PubMed]
- Galloway, S.E. Emergence of SARS-CoV-2b.1.1.7 lineage—United States, December 29, 2020–January 12, 2021. MMWR-Morb. Mortal. Wkly. Rep. 2021, 70, 95–99. [Google Scholar] [CrossRef]
- Lauring, A.S.; Hodcroft, E.B. Genetic Variants of SARS-CoV-2—What Do They Mean? JAMA 2021, 325, 529–531. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, A.; Srivastava, J.K.; Siddiqui, M.H.; Uddin, M.S.; Aleya, L. Hydroxychloroquine in COVID-19: Therapeutic promises, current status, and environmental implications. Environ. Sci. Pollut. Res. 2021, 28, 40431–40444. [Google Scholar] [CrossRef] [PubMed]
- Badraoui, R.; Adnan, M.; Bardakci, F.; Alreshidi, M.M. Chloroquine and Hydroxychloroquine Interact Differently with ACE2 Domains Reported to Bind with the Coronavirus Spike Protein: Mediation by ACE2 Polymorphism. Molecules 2021, 26, 673. [Google Scholar] [CrossRef]
- Colson, P.; Rolain, J.M.; Lagier, J.C.; Brouqui, P.; Raoult, D. Chloroquine and hydroxychloroquine as available weapons to fight COVID-19. Int. J. Antimicrob. Agents 2020, 55, 105932. [Google Scholar] [CrossRef]
- Li, J.G.; Xu, H. Chinese Medicine in Fighting against Covid-19: Role and Inspiration. Chin. J. Integr. Med. 2021, 27, 3–6. [Google Scholar] [CrossRef]
- Wang, J.; Xu, C.; Wong, Y.K.; He, Y.; Adegnika, A.A.; Kremsner, P.G.; Agnandji, S.T.; A Sall, A.; Liang, Z.; Qiu, C.; et al. Preparedness is essential for malaria-endemic regions during the COVID-19 pandemic. Lancet 2020, 395, 1094–1096. [Google Scholar] [CrossRef]
- Estrada, E. Topological analysis of SARS CoV-2 main protease. Chaos 2020, 30, 061102. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.V.; Teli, D.M.; Kanhed, A.M.; Patel, N.R.; Shah, B.S.; Vora, A.K.; Chhabria, M.T.; Yadav, M.R. Identification of Potential Mpro Inhibitors for the Treatment of COVID-19 by Targeted Covalent Inhibition: An In Silico Approach. Int. J. Quant. Struct. Prop. Relatsh. (IJQSPR) 2021, 6, 58–77. [Google Scholar]
- Kanhed, A.M.; Patel, D.V.; Teli, D.M.; Patel, N.R.; Chhabria, M.T.; Yadav, M.R. Identification of potential Mpro inhibitors for the treatment of COVID-19 by using systematic virtual screening approach. Mol. Divers. 2020, 25, 383–401. [Google Scholar] [CrossRef]
- Świderek, K.; Moliner, V. Revealing the molecular mechanisms of proteolysis of SARS-CoV-2 M pro by QM/MM computational methods. Chem. Sci. 2020, 11, 10626–10630. [Google Scholar] [CrossRef]
- Biembengut, Í.V.; De Souza, T.D.A.C. Coagulation modifiers targeting SARS-CoV-2 main protease Mpro for COVID-19 treatment: An in silico approach. Mem. Inst. Oswaldo Cruz 2020, 115, e200179. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Zhang, B.; Jiang, X.M.; Su, H.; Li, J.; Zhao, Y.; Xie, X.; Jin, Z.; Peng, J.; Liu, F.; et al. Structure-based design of antiviral drug candidates targeting the SARS-CoV-2 main protease. Science 2020, 368, 1331–1335. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef]
- da Silva, J.K.R.; Figueiredo, P.L.B.; Byler, K.G.; Setzer, W.N. Essential oils as antiviral agents, potential of essential oils to treat sars-cov-2 infection: An in-silico investigation. Int. J. Mol. Sci. 2020, 21, 3426. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzym. Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef]
- Narkhede, R.R.; Pise, A.V.; Cheke, R.S.; Shinde, S.D. Recognition of natural products as potential inhibitors of COVID-19 main protease (Mpro): In-silico evidences. Nat. Prod. Bioprospect. 2020, 10, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.H.; Woo, H.J.; Kang, H.K.; Kim, Y.M.; Kim, D.W.; Ahn, S.A.; Xia, Y.; Kim, D. Flavonoid-mediated inhibition of SARS coronavirus 3C-like protease expressed in Pichia pastoris. Biotechnol. Lett. 2012, 34, 831–838. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, J.Y.; Kim, D.; Nguyen, T.T.; Park, S.J.; Chang, J.S.; Park, K.H.; et al. Biflavonoids from Torreya nucifera displaying SARS-CoV 3CLpro inhibition. Bioorg. Med. Chem. 2010, 18, 7940–7947. [Google Scholar] [CrossRef] [PubMed]
- Arun, P.; Hussain, R.; Akhter, Y.; Rao, M.V.B.; Shyam, P.; Yellaboina, S. Drug Re-purposing from SARS-CoV Led the Identification of Potential Candidate Drug Target and Alternate Drug Molecules against SARSCoV-2. Lett. Drug Des. Discov. 2020, 17, 1325–1327. [Google Scholar] [CrossRef]
- Kandeel, M.; Altaher, A.; Alnazawi, M. Molecular dynamics and inhibition of MERS CoV papain-like protease by small molecule imidazole and aminopurine derivatives. Lett. Drug Des. Discov. 2019, 16, 584–591. [Google Scholar] [CrossRef]
- Kandeel, M.; Abdelrahman, A.H.; Oh-Hashi, K.; Ibrahim, A.; Venugopala, K.N.; Morsy, M.A.; Ibrahim, M.A.A. Repurposing of FDA-approved antivirals, antibiotics, anthelmintics, antioxidants, and cell protectives against SARS-CoV-2 papain-like protease. J. Biomol. Struct. Dyn. 2020, 39, 5129–5136. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, B.; Huang, Y.; Chen, Z.; Liu, P.; Huang, J.; Lao, L. Effects of moxa (Folium Artemisiae argyi) smoke exposure on heart rate and heart rate variability in healthy young adults: A randomized, controlled human study. Evid. Based Complement Altern. Med. 2013, 2013, 510318. [Google Scholar] [CrossRef]
- Munyangi, J.; Cornet-Vernet, L.; Idumbo, M.; Lu, C.; Lutgen, P.; Perronne, C.; Ngombe, N.; Bianga, J.; Mupenda, B.; Lalukala, P.; et al. Effect of Artemisia annua and Artemisia afra tea infusions on schistosomiasis in a large clinical trial. Phytomedicine 2018, 51, 233–240. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, N.; Zhu, W.; Zhang, Y.; Yang, J.; Wang, Y.; Song, X.; Hu, L.; Wu, Z.; Liu, Q.; et al. Does Moxa Smoke Have Significant Effect on the Acupuncturist’s Respiratory System? A Population-Based Study. Evid. Based Complement Altern. Med. 2019, 2019, 4873235. [Google Scholar] [CrossRef]
- Wheeler, J.; Coppock, B.; Chen, C. Does the burning of moxa (Artemisia vulgaris) in traditional Chinese medicine constitute a health hazard? Acupunct. Med. 2009, 27, 16–20. [Google Scholar] [CrossRef]
- Kapepula, P.M.; Kabengele, J.K.; Kingombe, M.; Van Bambeke, F.; Tulkens, P.M.; Kishabongo, A.S.; Decloedt, E.; Zumla, A.; Tiberi, S.; Suleman, F.; et al. Artemisia spp. derivatives for COVID-19 treatment: Anecdotal use, political hype, treatment potential, challenges, and road map to randomized clinical trials. Am. J. Trop. Med. Hyg. 2020, 103, 960–964. [Google Scholar] [CrossRef]
- Younis, W.; Asif, H.; Sharif, A.; Riaz, H.; Bukhari, I.A.; Assiri, A.M. Traditional medicinal plants used for respiratory disorders in Pakistan: A review of the ethno-medicinal and pharmacological evidence. Chin. Med. 2018, 13, 48. [Google Scholar]
- Yang, C.; Hu, D.H.; Feng, Y. Essential oil of Artemisia vestita exhibits potent in vitro and in vivo antibacterial activity: Investigation of the effect of oil on biofilm formation, leakage of potassium ions and survival curve measurement. Mol. Med. Rep. 2015, 12, 5762–5770. [Google Scholar] [CrossRef][Green Version]
- Efferth, T.; Romero, M.R.; Wolf, D.G.; Stamminger, T.; Marin, J.J.; Marschall, M. The antiviral activities of artemisinin and artesunate. Clin. Infect. Dis. 2008, 47, 804–811. [Google Scholar] [CrossRef]
- Sehailia, M.; Chemat, S. Antimalarial-agent artemisinin and derivatives portray more potent binding to Lys353 and Lys31-binding hotspots of SARS-CoV-2 spike protein than hydroxychloroquine: Potential repurposing of artenimol for COVID-19. J. Biomol. Struct. Dyn. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüuger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.E.; Kadioglu, O.; Seo, E.J.; Greten, H.J.; Brenk, R.; Efferth, T. Quantitative Structure–Activity Relationship and Molecular Docking of Artemisinin Derivatives to Vascular Endothelial Growth Factor Receptor 1. Anticancer Res. 2015, 35, 1929–1934. [Google Scholar] [PubMed]
- Roy, K.; Ojha, P.K. Advances in quantitative structure–activity relationship models of antimalarials. Expert Opin. Drug Discov. 2010, 5, 751–778. [Google Scholar] [CrossRef]
- Ginosyan, S.; Grabski, H.; Tiratsuyan, S. In vitro and in silico Determination of the Interaction of Artemisinin with Human Serum Albumin. Mol. Biol. 2020, 54, 653–666. [Google Scholar] [CrossRef]
- Rolta, R.; Salaria, D.; Kumar, V.; Sourirajan, A.; Dev, K. Phytocompounds of Rheum emodi, Thymus serpyllum and Artemisia annua inhibit COVID-19 binding to ACE2 receptor: In silico approach. Curr. Pharmacol. Rep. 2021, 7, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Omar, S.; Bouziane, I.; Bouslama, Z.; Djemel, A. In-silico identification of potent inhibitors of COVID-19 main protease (Mpro) and angiotensin converting enzyme 2 (ACE2) from natural products: Quercetin, hispidulin, and cirsimaritin exhibited better potential inhibition than hydroxy-chloroquine against COVID-19 main protease active site and ACE2. Biol. Med. Chem. 2020. [Google Scholar] [CrossRef]
- Saoudi, M.; Badraoui, R.; Bouhajja, H.; Ncir, M.; Rahmouni, F.; Grati, M.; Jamoussi, K.; El Feki, A. Deltamethrin induced oxidative stress in kidney and brain of rats: Protective effect of Artemisia campestris essential oil. Biomed. Pharmacother. 2017, 94, 955–963. [Google Scholar] [CrossRef]
- Tu, Y. The discovery of artemisinin (qinghaosu) and gifts from Chinese medicine. Nat. Med. 2011, 17, 1217–1220. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Yuan, W.; Li, S.; Gupta, S.C. Curcumin-free turmeric exhibits anti-inflammatory and anticancer activities: Identification of novel components of turmeric. Mol. Nutr. Food Res. 2013, 57, 1529–1542. [Google Scholar] [CrossRef]
- Badraoui, R.; Rebai, T.; Elkahoui, S.; Alreshidi, M.; Veettil, V.N.; Noumi, E.; Al-Motair, K.A.; Aouadi, K.; Kadri, A.; De Feo, V.; et al. Allium subhirsutum L. as a Potential Source of Antioxidant and Anticancer Bioactive Molecules: HR-LCMS Phytochemical Profiling, In Vitro and In Vivo Pharmacological Study. Antioxidants 2020, 9, 1003. [Google Scholar] [CrossRef] [PubMed]
- Akacha, A.; Badraoui, R.; Rebai, T.; Zourgui, L. Effect of Opuntia ficus indica extract on methotrexate-induced testicular injury: A biochemical, docking and histological study. J. Biomol. Struct. Dyn. 2020, 1–11. [Google Scholar] [CrossRef]
- Zammel, N.; Saeed, M.; Bouali, N.; Elkahoui, S.; Alam, J.M.; Rebai, T.; Kausar, M.A.; Adnan, M.; Siddiqui, A.J.; Badraoui, R. Antioxidant and anti-Inflammatory effects of Zingiber officinale roscoe and Allium subhirsutum: In silico, biochemical and histological Study. Foods 2021, 10, 1383. [Google Scholar] [CrossRef]
- Mzid, M.; Badraoui, R.; Kedir, S.B.; Sahnoun, Z.; Rebai, T. Protective effects of ethanolic extract of Urtica urens L. against toxicity of imidacloprid on bone remodeling in rats and antioxidant activities. Biomed. Pharmacother. 2017, 91, 1022–1041. [Google Scholar] [CrossRef]
- De Olivera, V.M.; da Rocha, M.N.; Magalhães, E.P.; da Silva Mendes, F.R.; Marinho, M.M.; de Menezes, R.R.P.P.B.; Sampaio, T.L.; Dos Santos, H.S.; Martins, A.M.C.; Marinho, E.S. Computational approach towards the design of artemisinin-thymoquinone hybrids against main protease of SARS-CoV-2. Future J. Pharm. Sci. 2021, 7, 185. [Google Scholar] [CrossRef]
- Bakchiche, B.; Gherib, A.; Bronze, M.R.; Ghareeb, M.A. Identification, quantification, and antioxidant activity of hydroalcoholic extract of Artemisia campestris from Algeria. Turk. J. Pharm. Sci. 2019, 16, 234. [Google Scholar] [CrossRef] [PubMed]
- Saoudi, M.; Badraoui, R.; Rahmouni, F.; Jamoussi, K.; El Feki, A. Antioxidant and protective effects of Artemisia campestris essential oil against chlorpyrifos-induced kidney and liver injuries in rats. Front. Physiol. 2021, 12, 194. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.G.; Barreira, L.; Bijttebier, S.; Pieters, L.; Marques, C.; Santos, T.F.; Rodrigues, M.J.; Varela, J.; Custodio, L. Health promoting potential of herbal teas and tinctures from Artemisia campestris subsp. maritima: From traditional remedies to prospective products. Sci. Rep. 2018, 8, 4689. [Google Scholar]
- Ben Nasr, H.; Badraoui, R. Approach of utilizing Artemisia herbs to treat covid-19. Braz. J. Pharm. Sci. 2022, in press. [Google Scholar]
- Sudeep, H.V.; Gouthamchandra, K.; Shyamprasad, K. Molecular docking analysis of Withaferin A from Withania somnifera with the Glucose regulated protein 78 (GRP78) receptor and the SARS-CoV-2 main protease. Bioinformation 2020, 16, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Hchicha, K.; Korb, M.; Badraoui, R.; Naïli, H. A novel sulfate-bridged binuclear copper (II) complex: Structure, optical, ADMET and in vivo approach in a murine model of bone metastasis. New J. Chem. 2021, 45, 13775–13784. [Google Scholar] [CrossRef]
- Li, F.; Li, W.; Farazan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complex with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Hussain, M.; Jabeen, N.; Raza, F.; Shabbir, S.; Baig, A.A.; Amanullah, A.; Aziz, B. Structural variations in human ACE2 may influence its binding with SARS-CoV-2 spike protein. J. Med. Virol. 2020, 92, 1580–1586. [Google Scholar] [CrossRef]
- Othman, I.M.M.; Gad-Elkareem, M.A.M.; Radwan, H.A.; Badraoui, R.; Aouadi, K.; Snoussi, M.; Kadri, A. Synthesis, Structure-Activity Relationship and in silico Studies of Novel Pyrazolothiazole and Thiazolopyridine Derivatives as Prospective Antimicrobial and Anticancer Agents. ChemistrySelect 2021, 6, 7860–7872. [Google Scholar] [CrossRef]
- Badraoui, R.; Saeed, M.; Bouali, N.; Hamadou, W.S.; Elkahoui, S.; Alam, M.J.; Siddiqui, A.J.; Adnan, M.; Saoudi, M.; Rebai, T. Expression profiling of selected immune genes and trabecular microarchitecture in breast cancer skeletal metastases model; effect of α–tocopherol acetate supplementation. Calcif. Tissue Int. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.; Shoaib, A.; Tasleem, M.; Alabdallah, N.M.; Alam, M.J.; El Asmar, Z.; Jamal, Q.M.S.; Bardakci, F.; Alqahtani, S.S.; Ansari, I.A.; et al. Assessment of antidiabetic activity of the shikonin by the allosteric inhibition of protein-tyrosine phosphate 1B (PTP1B) using state of art: An in silico and in vitro tactics. Molecules 2021, 26, 3996. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.K.; Kumar, S.; Saloni; Misra, S.; Yadav, L.; Teli, M.; Sharma, P.; Chaudhary, S.; Kumar, N.; Choi, E.H.; et al. Molecular Insights into the Interaction of RONS and Thieno[3,2-c]pyran Analogs with SIRT6/COX-2: A Molecular Dynamics Study. Sci. Rep. 2018, 8, 4777. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
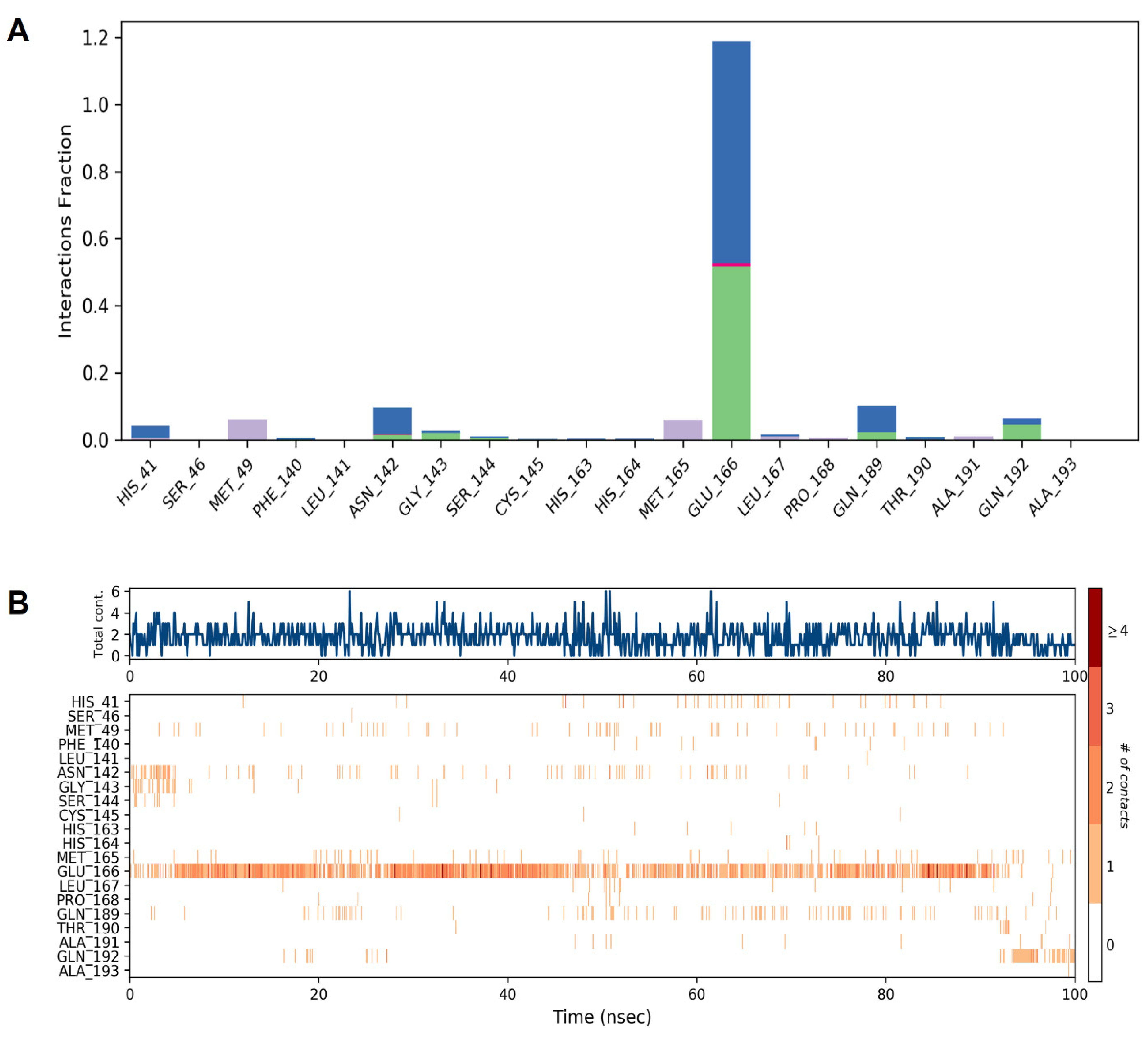{kind=link}
| Entry | Compounds | ||||
|---|---|---|---|---|---|
| A | B | C | D | E | |
| Properties/Lipophilicity/Drug-likeness | |||||
| Molecular weight (g × mol−1) | 282.33 | 266.33 | 218.33 | 266.33 | 284.35 |
| Num. heavy atoms | 20 | 19 | 16 | 19 | 20 |
| Num. arom. heavy atoms | 0 | 0 | 0 | 0 | 0 |
| Fraction Csp3 | 0.93 | 0.87 | 0.67 | 0.93 | 1.00 |
| Num. rotatable bonds | 0 | 0 | 2 | 0 | 0 |
| Num. H-bond acceptors | 5 | 4 | 1 | 4 | 5 |
| Num. H-bond donors | 0 | 0 | 0 | 0 | 1 |
| Molar Refractivity | 70.38 | 69.71 | 69.24 | 69.29 | 71.34 |
| TPSA (Ų) | 53.99 | 36.92 | 17.07 | 44.76 | 57.15 |
| Consensus Log Po/w | 2.49 | 2.72 | 3.44 | 2.58 | 2.25 |
| Lipinski’s Rule. | Yes | Yes | Yes | Yes | Yes |
| Bioavailability Score | 0.55 | 0.55 | 0.55 | 0.55 | 0.55 |
| PAINS | 0 alert | 0 alert | 0 alert | 0 alert | 0 alert |
| Pharmacokinetics | |||||
| GI absorption | High | High | High | High | High |
| BBB permeant | Yes | Yes | Yes | Yes | Yes |
| P-gp substrate | No | No | No | No | No |
| CYP1A2 inhibitor | Yes | Yes | No | Yes | Yes |
| CYP2C19 inhibitor | No | No | Yes | No | No |
| CYP2C9 inhibitor | No | No | Yes | No | No |
| CYP2D6 inhibitor | No | No | No | No | No |
| CYP3A4 inhibitor | No | No | No | No | No |
| Log Kp (cm/s) | −5.96 | −6.05 | −5.01 | −5.90 | −5.91 |
| Compound Name | Binding Affinity (kcal/mol) | Intermolecular Interactions | ||
|---|---|---|---|---|
| Conventional Hydrogen Bonds | Interacting Residues in the Pocket Region of SARS-CoV-2 Mpro | Closest Residue (Distance, Å) | ||
| Artemisinin | −7.2 | 3 | Thr26, Ser144, Cys145 | Ser144 (2.436) |
| Acetate of dihydroartemisinin | −8.1 | 2 | His41, Tyr54, Gln189 | Tyr54 (2.743) |
| Artemisinic Aldehyde | −5.2 | 2 | Ser144, Cys145 | Cys145 (2.374) |
| Deoxyartemisinin | −6.5 | 2 | Thr26, Gly143 | Thr26 (2.206) |
| Dihydroartemisinin | −6.6 | 3 | His41, Leu141, Gly143, Cys145 | Gly143 (1.980) |
| Chloroquine (Reference) | −5.3 | 3 | Leu27, His41, Leu141, Ser144, Cys145, Met165, Glu166 | (2.815) |
| ACE2 Allelic Variant | Intermolecular Interactions | |
|---|---|---|
| Interacting Residues in the Pocket Region of ACE2 Variant (Distance, Å) | Binding Affinity (kcal/mol) | |
| rs4646116 | Conventional H-Bond: Thr41 (2.219) | −6.2 |
| Pi Cation: Trp37 (4.044) | ||
| Alkyl: Arg34 (4.897) | ||
| Pi-Alkyl: Trp37 (4.394) | ||
| rs73635825 | Conventional H-Bond: Ser13 (2.274), Ser13 (2.874) | −5.4 |
| Pi-Cation: Trp16 (4.328), Trp16 (4.514), Trp16 (4.105) | ||
| rs146676783 | Conventional H-Bond: Asn53 (2.258), Ala41 (3.359), Asn53 (3.104) | −5.2 |
| Carbon H-Bond: Ala41 (2.892) | ||
| rs76289235 | Conventional H-Bond: Tyr26 (2.163) | −5.7 |
| Pi-Cation: Tyr26 (4.375) | ||
| rs14393283 | Conventional H-Bond: Trp33 (2.921), Gln30 (3.372) | −5.8 |
| Attractive Charge: Glu17 (3.790), Glu17 (3.195) | ||
| Carbon H-Bond: Trp33 (3.338) | ||
| rs766996587 | Conventional H-Bond: Asn53 (2.254), Asn53 (2.917), Asn53 (3.017) | −4.9 |
| rs1348114695 | Conventional H-Bond: Arg21 (2.428) | −5.7 |
| Alkyl: Ile18 (4.943) | ||
| Pi-Alkyl: Phe12 (4.405) | ||
| rs961360700 | Conventional H-Bond: Thr3 (2.162), Gly33 (3.377) | −8.2 |
| rs755691167 | Conventional H-Bond: Gln54 (2.346) | −6.7 |
| Alkyl: Val58 (3.886) | ||
| rs1316056737 | Conventional H-Bond: Lys37 (2.363), Lys37 (2.588), Lys37 (2.632), Phe35 (3.267) | −5.1 |
| Alkyl: Lys23 (4.193) | ||
| rs781255386 | Conventional H-Bond: Arg25 (2.494), Lys30 (2.829) | −5.1 |
| rs1299103394 | Conventional H-Bond: Ser10 (2.222), Asp13 (3.259) | −5.4 |
| Carbon H-Bond: Ser10 (3.414) | ||
| Alkyl: Cys16 (4.680), Val24 (4.002) | ||
| rs759134032 | Conventional H-Bond: Ser57 (2.127) | −5.7 |
| Carbon H-Bond: Leu29 (3.680) | ||
| rs1238146879 | Conventional H-Bond: Tyr5 (3.056) | −5.4 |
| Pi-Cation: Tyr5 (3.898) | ||
| rs778500138 | Conventional H-Bond: Leu37 (2.078), Ser38 (2.958), Asn36 (3.219) | −5.6 |
| Alkyl: Leu35 (4.335) | ||
| rs1396769231 | Conventional H-Bond: Ser48 (2.586), Arg49 (2.670), Leu51 (2.417) | −5.5 |
| rs1016777825 | Conventional H-Bond: His34 (2.831) | −5.4 |
| Alkyl: Pro21 (4.570), Val27 (4.102), Ile29 (5.295) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badraoui, R.; Saoudi, M.; Hamadou, W.S.; Elkahoui, S.; Siddiqui, A.J.; Alam, J.M.; Jamal, A.; Adnan, M.; Suliemen, A.M.E.; Alreshidi, M.M.; et al. Antiviral Effects of Artemisinin and Its Derivatives against SARS-CoV-2 Main Protease: Computational Evidences and Interactions with ACE2 Allelic Variants. Pharmaceuticals 2022, 15, 129. https://doi.org/10.3390/ph15020129
Badraoui R, Saoudi M, Hamadou WS, Elkahoui S, Siddiqui AJ, Alam JM, Jamal A, Adnan M, Suliemen AME, Alreshidi MM, et al. Antiviral Effects of Artemisinin and Its Derivatives against SARS-CoV-2 Main Protease: Computational Evidences and Interactions with ACE2 Allelic Variants. Pharmaceuticals. 2022; 15(2):129. https://doi.org/10.3390/ph15020129
Chicago/Turabian StyleBadraoui, Riadh, Mongi Saoudi, Walid S. Hamadou, Salem Elkahoui, Arif J. Siddiqui, Jahoor M. Alam, Arshad Jamal, Mohd Adnan, Abdel M. E. Suliemen, Mousa M. Alreshidi, and et al. 2022. "Antiviral Effects of Artemisinin and Its Derivatives against SARS-CoV-2 Main Protease: Computational Evidences and Interactions with ACE2 Allelic Variants" Pharmaceuticals 15, no. 2: 129. https://doi.org/10.3390/ph15020129
APA StyleBadraoui, R., Saoudi, M., Hamadou, W. S., Elkahoui, S., Siddiqui, A. J., Alam, J. M., Jamal, A., Adnan, M., Suliemen, A. M. E., Alreshidi, M. M., Yadav, D. K., Naïli, H., & Ben-Nasr, H. (2022). Antiviral Effects of Artemisinin and Its Derivatives against SARS-CoV-2 Main Protease: Computational Evidences and Interactions with ACE2 Allelic Variants. Pharmaceuticals, 15(2), 129. https://doi.org/10.3390/ph15020129