
Discovery of Novel Andrographolide Derivatives as Antiviral Inhibitors against Human Enterovirus A71

 , and
, and 
Abstract
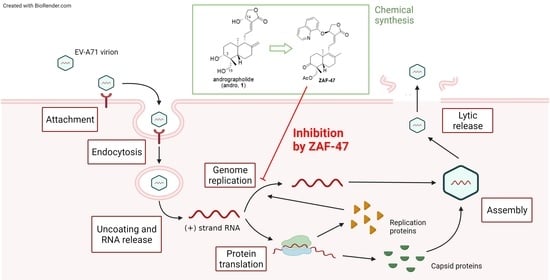
:
1. Introduction
2. Results
2.1. High Throughput Screening of Compound Library Reveals Novel EV-A71 Inhibitors
2.2. Validation of EV-A71 Hits
2.3. Synthesis of ZAF-46 and ZAF-47
2.4. ZAF-47 Targets EV-A71 Post Entry Stages
2.5. ZAF-47 Reduces Expression of EV-A71 Viral Proteins by Specifically Targeting EV-A71 RNA Replication
2.6. ZAF-47 Possibly Targets Host Factor/s Associated with EV-A71 RNA Replication
2.7. Broad-Spectrum Anti-enterovirus Activity of ZAF-47
3. Discussion
4. Materials and Methods
4.1. Synthesis of ZAF-46 and ZAF-47
4.1.1. (14β)-(8′-Quinolinoxy)-9-dehydro-17-hydro-3,19-isopropyleneoxy-andrographolide (7)
4.1.2. (14β)-(8’-Quinolinoxy)-9-dehydro-17-hydro-andrographolide (8)
4.1.3. (14β)-(8′-Quinolinoxy)-9-dehydro-17-hydro-19-acetoxy-andrographolide (ZAF-46)
4.1.4. (14β)-(8′-Quinolinoxy)-3-ketone-9-dehydro-17-hydro-19-acetoxy–andrographolide (ZAF-47)
4.2. Cells and Viruses
4.3. Compound Library
4.4. Preliminary Screen
4.5. Cell Viability Assay
4.6. Post-Treatment Assay
4.7. Viral Plaque Assays
4.8. Time-of-Addition and Time-of-Removal Assays
4.9. Pre-Treatment Assay
4.10. Co-Treatment Assay
4.11. Entry Bypass Assay
4.12. SDS-PAGE and Western Blot
4.13. Nano-Luciferase Reporter Assay
4.14. Bicistronic Luciferase Reporter Assay
4.15. Generation of Resistant Mutant
4.16. Antiviral Spectrum of ZAF-47
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Patil, R.; Jain, V. Andrographolide: A Review of Analytical Methods. J. Chromatogr. Sci. 2021, 59, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Chen, S.-R.; Chai, L.; Zhao, J.; Wang, Y.; Wang, Y. Overview of Pharmacological Activities of Andrographis Paniculata and Its Major Compound Andrographolide. Crit. Rev. Food Sci. Nutr. 2019, 59, S17–S29. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Singh, D.; Tali, J.A.; Dheer, D.; Shankar, R. Andrographolide: Chemical Modification and Its Effect on Biological Activities. Bioorganic Chem. 2020, 95, 103511. [Google Scholar] [CrossRef]
- Jiang, M.; Sheng, F.; Zhang, Z.; Ma, X.; Gao, T.; Fu, C.; Li, P. Andrographis Paniculata (Burm. f.) Nees and Its Major Constituent Andrographolide as Potential Antiviral Agents. J. Ethnopharmacol. 2021, 272, 113954. [Google Scholar] [CrossRef]
- Wintachai, P.; Kaur, P.; Lee, R.C.H.; Ramphan, S.; Kuadkitkan, A.; Wikan, N.; Ubol, S.; Roytrakul, S.; Chu, J.J.H.; Smith, D.R. Activity of Andrographolide against Chikungunya Virus Infection. Sci. Rep. 2015, 5, 14179. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, S.; Dar, L.; Kaushik, S.; Yadav, J.P. Identification and Characterization of New Potent Inhibitors of Dengue Virus NS5 Proteinase from Andrographis Paniculata Supercritical Extracts on in Animal Cell Culture and in Silico Approaches. J. Ethnopharmacol. 2021, 267, 113541. [Google Scholar] [CrossRef]
- Panraksa, P.; Ramphan, S.; Khongwichit, S.; Smith, D.R. Activity of Andrographolide against Dengue Virus. Antivir. Res. 2017, 139, 69–78. [Google Scholar] [CrossRef]
- Lin, T.-P.; Chen, S.-Y.; Duh, P.-D.; Chang, L.-K.; Liu, Y.-N. Inhibition of the Epstein-Barr Virus Lytic Cycle by Andrographolide. Biol. Pharm. Bull. 2008, 31, 2018–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Dai, C.; Jiang, Z.; Li, E.; Chen, C.; Wu, X.; Chen, J.; Liu, Q.; Zhao, C.; He, J.; et al. Andrographolide as an Anti-H1N1 Drug and the Mechanism Related to Retinoic Acid-Inducible Gene-I-like Receptors Signaling Pathway. Chin. J. Integr. Med. 2014, 20, 540–545. [Google Scholar] [CrossRef]
- Chen, H.; Ma, Y.-B.; Huang, X.-Y.; Geng, C.-A.; Zhao, Y.; Wang, L.-J.; Guo, R.-H.; Liang, W.-J.; Zhang, X.-M.; Chen, J.-J. Synthesis, Structure—Activity Relationships and Biological Evaluation of Dehydroandrographolide and Andrographolide Derivatives as Novel Anti-Hepatitis B Virus Agents. Bioorganic Med. Chem. Lett. 2014, 24, 2353–2359. [Google Scholar] [CrossRef]
- Lee, J.-C.; Tseng, C.-K.; Young, K.-C.; Sun, H.-Y.; Wang, S.-W.; Chen, W.-C.; Lin, C.-K.; Wu, Y.-H. Andrographolide Exerts Anti-Hepatitis C Virus Activity by up-Regulating Haeme Oxygenase-1 via the P38 MAPK/Nrf2 Pathway in Human Hepatoma Cells. Br. J. Pharmacol. 2014, 171, 237–252. [Google Scholar] [CrossRef]
- Uttekar, M.M.; Das, T.; Pawar, R.S.; Bhandari, B.; Menon, V.; Nutan; Gupta, S.K.; Bhat, S.V. Anti-HIV Activity of Semisynthetic Derivatives of Andrographolide and Computational Study of HIV-1 Gp120 Protein Binding. Eur. J. Med. Chem. 2012, 56, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Niranjan Reddy, V.L.; Malla Reddy, S.; Ravikanth, V.; Krishnaiah, P.; Venkateshwar Goud, T.; Rao, T.P.; Siva Ram, T.; Gonnade, R.G.; Bhadbhade, M.; Venkateswarlu, Y. A New BIS-Andrographolide Ether from Andrographis Paniculata Nees and Evaluation of Anti-HIV Activity. Nat. Prod. Res. 2005, 19, 223–230. [Google Scholar] [CrossRef]
- Seubsasana, S.; Pientong, C.; Ekalaksananan, T.; Thongchai, S.; Aromdee, C. A Potential Andrographolide Analogue against the Replication of Herpes Simplex Virus Type 1 in Vero Cells. Med. Chem. 2011, 7, 237–244. [Google Scholar] [CrossRef]
- Wiart, C.; Kumar, K.; Yusof, M.Y.; Hamimah, H.; Fauzi, Z.M.; Sulaiman, M. Antiviral Properties of Ent-Labdene Diterpenes of Andrographis Paniculata Nees, Inhibitors of Herpes Simplex Virus Type 1. Phytother. Res. 2005, 19, 1069–1070. [Google Scholar] [CrossRef] [PubMed]
- Sa-ngiamsuntorn, K.; Suksatu, A.; Pewkliang, Y.; Thongsri, P.; Kanjanasirirat, P.; Manopwisedjaroen, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Pitiporn, S.; Chaopreecha, J.; et al. Anti-SARS-CoV-2 Activity of Andrographis Paniculata Extract and Its Major Component Andrographolide in Human Lung Epithelial Cells and Cytotoxicity Evaluation in Major Organ Cell Representatives. J. Nat. Prod. 2021, 84, 1261–1270. [Google Scholar] [CrossRef]
- Shi, T.-H.; Huang, Y.-L.; Chen, C.-C.; Pi, W.-C.; Hsu, Y.-L.; Lo, L.-C.; Chen, W.-Y.; Fu, S.-L.; Lin, C.-H. Andrographolide and Its Fluorescent Derivative Inhibit the Main Proteases of 2019-NCoV and SARS-CoV through Covalent Linkage. Biochem. Biophys. Res. Commun. 2020, 533, 467–473. [Google Scholar] [CrossRef]
- Wang, D.; Guo, H.; Chang, J.; Wang, D.; Liu, B.; Gao, P.; Wei, W. Andrographolide Prevents EV-D68 Replication by Inhibiting the Acidification of Virus-Containing Endocytic Vesicles. Front. Microbiol. 2018, 9, 2407. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.J.; Lennette, E.H.; Ho, H.H. An Apparently New Enterovirus Isolated from Patients with Disease of the Central Nervous System. J. Infect. Dis. 1974, 129, 304–309. [Google Scholar] [CrossRef]
- Kennett, M.L.; Birch, C.J.; Lewis, F.A.; Yung, A.P.; Locarnini, S.A.; Gust, I.D. Enterovirus Type 71 Infection in Melbourne. Bull. World Health Organ. 1974, 51, 609–615. [Google Scholar] [PubMed]
- Chumakov, M.; Voroshilova, M.; Shindarov, L.; Lavrova, I.; Gracheva, L.; Koroleva, G.; Vasilenko, S.; Brodvarova, I.; Nikolova, M.; Gyurova, S.; et al. Enterovirus 71 Isolated from Cases of Epidemic Poliomyelitis-like Disease in Bulgaria. Arch. Virol. 1979, 60, 329–340. [Google Scholar] [CrossRef]
- Tagaya, I.; Takayama, R.; Hagiwara, A. A Large-Scale Epidemic of Hand, Foot and Mouth Disease Associated with Enterovirus 71 Infection in Japan in 1978. Jpn. J. Med. Sci. Biol. 1981, 34, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Nagy, G.; Takátsy, S.; Kukán, E.; Mihály, I.; Dömök, I. Virological Diagnosis of Enterovirus Type 71 Infections: Experiences Gained during an Epidemic of Acute CNS Diseases in Hungary in 1978. Arch. Virol. 1982, 71, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Chua, B.H.; Lee, C.S.M.; Chem, Y.K.; Ismail, N.; Kiyu, A.; Kumarasamy, V. Genetic Diversity of Enterovirus 71 Isolated from Cases of Hand, Foot and Mouth Disease in the 1997, 2000 and 2005 Outbreaks, Peninsular Malaysia. Malays. J. Pathol. 2007, 29, 69–78. [Google Scholar] [PubMed]
- Ho, M.; Chen, E.-R.; Hsu, K.-H.; Twu, S.-J.; Chen, K.-T.; Tsai, S.-F.; Wang, J.-R.; Shih, S.-R. An Epidemic of Enterovirus 71 Infection in Taiwan. N. Engl. J. Med. 1999, 341, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-W.; Hsu, Y.-W.; Smith, D.J.; Kiang, D.; Tsai, H.-P.; Lin, K.-H.; Wang, S.-M.; Liu, C.-C.; Su, I.-J.; Wang, J.-R. Reemergence of Enterovirus 71 in 2008 in Taiwan: Dynamics of Genetic and Antigenic Evolution from 1998 to 2008. J. Clin. Microbiol. 2009, 47, 3653–3662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.-H.; Hwang, K.-P.; Ke, G.-M.; Wang, C.-F.; Ke, L.-Y.; Hsu, Y.-T.; Tung, Y.-C.; Chu, P.-Y.; Chen, B.-H.; Chen, H.-L.; et al. Evolution of EV71 Genogroup in Taiwan from 1998 to 2005: An Emerging of Subgenogroup C4 of EV71. J. Med. Virol. 2006, 78, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-Y.; Twu, S.-J.; Ho, M.-S.; Chang, L.-Y.; Lee, C.-Y. Enterovirus 71 Outbreaks, Taiwan: Occurrence and Recognition. Emerg. Infect. Dis. 2003, 9, 291–293. [Google Scholar] [CrossRef] [Green Version]
- Hosoya, M.; Kawasaki, Y.; Sato, M.; Honzumi, K.; Kato, A.; Hiroshima, T.; Ishiko, H.; Suzuki, H. Genetic Diversity of Enterovirus 71 Associated with Hand, Foot and Mouth Disease Epidemics in Japan from 1983 to 2003. Pediatr. Infect. Dis. J. 2006, 25, 691–694. [Google Scholar] [CrossRef]
- Wu, Y.; Yeo, A.; Phoon, M.C.; Tan, E.L.; Poh, C.L.; Quak, S.H.; Chow, V.T.K. The Largest Outbreak of Hand; Foot and Mouth Disease in Singapore in 2008: The Role of Enterovirus 71 and Coxsackievirus a Strains. Int. J. Infect. Dis. 2010, 14, e1076–e1081. [Google Scholar] [CrossRef] [Green Version]
- Chong, C.Y.; Chan, K.P.; Shah, V.A.; Ng, W.Y.M.; Lau, G.; Teo, T.E.S.; Lai, S.H.; Ling, A.E. Hand, Foot and Mouth Disease in Singapore: A Comparison of Fatal and Non-Fatal Cases. Acta Paediatr. 2003, 92, 1163–1169. [Google Scholar] [CrossRef]
- Yang, B.; Liu, F.; Liao, Q.; Wu, P.; Chang, Z.; Huang, J.; Long, L.; Luo, L.; Li, Y.; Leung, G.M.; et al. Epidemiology of Hand, Foot and Mouth Disease in China, 2008 to 2015 Prior to the Introduction of EV-A71 Vaccine. Eurosurveillance 2017, 22, 16–00824. [Google Scholar] [CrossRef] [Green Version]
- Puenpa, J.; Wanlapakorn, N.; Vongpunsawad, S.; Poovorawan, Y. The History of Enterovirus A71 Outbreaks and Molecular Epidemiology in the Asia-Pacific Region. J. Biomed. Sci. 2019, 26, 75. [Google Scholar] [CrossRef]
- Liu, M.Y.; Liu, W.; Luo, J.; Liu, Y.; Zhu, Y.; Berman, H.; Wu, J. Characterization of an Outbreak of Hand, Foot, and Mouth Disease in Nanchang, China in 2010. PLoS ONE 2011, 6, e25287. [Google Scholar] [CrossRef] [Green Version]
- Messacar, K.; Burakoff, A.; Nix, W.A.; Rogers, S.; Oberste, M.S.; Gerber, S.I.; Spence-Davizon, E.; Herlihy, R.; Dominguez, S.R. Notes from the Field: Enterovirus A71 Neurologic Disease in Children—Colorado, 2018. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1017–1018. [Google Scholar] [CrossRef] [Green Version]
- Taravilla, C.N.; Pérez-Sebastián, I.; Salido, A.G.; Serrano, C.V.; Extremera, V.C.; Rodríguez, A.D.; Marín, L.L.; Sanz, M.A.; Traba, O.M.S.; González, A.S. Enterovirus A71 Infection and Neurologic Disease, Madrid, Spain, 2016. Emerg. Infect. Dis. 2019, 25, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antona, D.; Kossorotoff, M.; Schuffenecker, I.; Mirand, A.; Leruez-Ville, M.; Bassi, C.; Aubart, M.; Moulin, F.; Lévy-Bruhl, D.; Henquell, C.; et al. Severe Paediatric Conditions Linked with EV-A71 and EV-D68, France, May to October 2016. Eurosurveillance 2016, 21, 30402. [Google Scholar] [CrossRef]
- Li, X.; Zhang, C.; Shi, Q.; Yang, T.; Zhu, Q.; Tian, Y.; Lu, C.; Zhang, Z.; Jiang, Z.; Zhou, H.; et al. Improving the Efficacy of Conventional Therapy by Adding Andrographolide Sulfonate in the Treatment of Severe Hand, Foot, and Mouth Disease: A Randomized Controlled Trial. Evid.-Based Complement. Altern. Med. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Hou, Y.; Li, Y.; He, L.; Tang, L.; Yang, T.; Zou, X.; Zhu, Q.; Yan, S.; Huang, B.; et al. Xiyanping Injection Therapy for Children with Mild Hand Foot and Mouth Disease: A Randomized Controlled Trial. J. Tradit. Chin. Med. 2017, 37, 397–403. [Google Scholar] [PubMed]
- Li, M.; Yang, X.; Guan, C.; Wen, T.; Duan, Y.; Zhang, W.; Li, X.; Wang, Y.; Zhao, Z.; Liu, S. Andrographolide Sulfonate Reduces Mortality in Enterovirus 71 Infected Mice by Modulating Immunity. Int. Immunopharmacol. 2018, 55, 142–150. [Google Scholar] [CrossRef]
- Min, N.; Leong, P.T.; Lee, R.C.H.; Khuan, J.S.E.; Chu, J.J.H. A Flavonoid Compound Library Screen Revealed Potent Antiviral Activity of Plant-Derived Flavonoids on Human Enterovirus A71 Replication. Antivir. Res. 2018, 150, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-H.; Chung, T.D.Y.; Oldenburg, K.R. A Simple Statistical Parameter for Use in Evaluation and Validation of High Throughput Screening Assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Dai, K.; Tan, J.K.; Qian, W.; Lee, R.C.H.; Chu, J.J.H.; Zhou, G.-C. Discovery of 14S-(2′-Chloro-4′-Nitrophenoxy)-8R/S, 17-Epoxy Andrographolide as EV-A71 Infection Inhibitor. Biochem. Pharmacol. 2021, 194, 114820. [Google Scholar] [CrossRef]
- Li, F.; Lee, E.M.; Sun, X.; Wang, D.; Tang, H.; Zhou, G.-C. Design, Synthesis and Discovery of Andrographolide Derivatives against Zika Virus Infection. Eur. J. Med. Chem. 2020, 187, 111925. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Law, W.-K.; Wang, D.; Nie, X.; Sheng, D.; Song, G.; Guo, K.; Wei, P.; Ouyang, P.; Wong, C.-W.; et al. Synthesis and Discovery of Andrographolide Derivatives as Non-Steroidal Farnesoid X Receptor (FXR) Antagonists. RSC Adv. 2014, 4, 13533–13545. [Google Scholar] [CrossRef]
- Dey, A.; Chen, R.; Li, F.; Maitra, S.; Hernandez, J.-F.; Zhou, G.-C.; Vincent, B. Synthesis and Characterization of Andrographolide Derivatives as Regulators of ΒAPP Processing in Human Cells. Molecules 2021, 26, 7660. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; He, Y.Q.; Yi, L.N.; Zan, H.; Kung, H.F.; He, M.L. Viral Kinetics of Enterovirus 71 in Human Habdomyosarcoma Cells. World J. Gastroenterol. 2011, 17, 4135–4142. [Google Scholar] [CrossRef]
- Wu, K.X.; Chu, J.J.-H. Antiviral Screen Identifies EV71 Inhibitors and Reveals Camptothecin-Target, DNA Topoisomerase 1 as a Novel EV71 Host Factor. Antivir. Res. 2017, 143, 122–133. [Google Scholar] [CrossRef]
- Yuan, J.; Shen, L.; Wu, J.; Zou, X.; Gu, J.; Chen, J.; Mao, L. Enterovirus A71 Proteins: Structure and Function. Front. Microbiol. 2018, 9, 286. [Google Scholar] [CrossRef] [Green Version]
- Murray, K.E.; Nibert, M.L. Guanidine Hydrochloride Inhibits Mammalian Orthoreovirus Growth by Reversibly Blocking the Synthesis of Double-Stranded RNA. J. Virol. 2007, 81, 4572–4584. [Google Scholar] [CrossRef] [Green Version]
- Schneider-Poetsch, T.; Ju, J.; Eyler, D.E.; Dang, Y.; Bhat, S.; Merrick, W.C.; Green, R.; Shen, B.; Liu, J.O. Inhibition of Eukaryotic Translation Elongation by Cycloheximide and Lactimidomycin. Nat. Chem. Biol. 2010, 6, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Gunaseelan, S.; Wong, K.Z.; Min, N.; Sun, J.; Ismail, N.K.B.M.; Tan, Y.J.; Lee, R.C.H.; Chu, J.J.H. Prunin Suppresses Viral IRES Activity and Is a Potential Candidate for Treating Enterovirus A71 Infection. Sci. Transl. Med. 2019, 11, eaar5759. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Qiu, M.; Chen, D.; Zheng, N.; Jin, Y.; Wu, Z. Apigenin Inhibits Enterovirus 71 Replication through Suppressing Viral IRES Activity and Modulating Cellular JNK Pathway. Antivir. Res. 2014, 109, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Swinney, D.C. Phenotypic vs. Target-Based Drug Discovery for First-in-Class Medicines. Clin. Pharmacol. Ther. 2013, 93, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Loureiro Damasceno, J.P.; Silva da Rosa, H.; Silva de Araújo, L.; Jacometti Cardoso Furtado, N.A. Andrographis Paniculata Formulations: Impact on Diterpene Lactone Oral Bioavailability. Eur. J. Drug Metab. Pharmacokinet. 2022, 47, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wei, H.; Wang, J.; Wang, W.; Gao, J. Synthesis of Andrographolide Analogues and Their Neuroprotection and Neurite Outgrowth-Promoting Activities. Bioorganic Med. Chem. 2019, 27, 2209–2219. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Xu, C.; Ji, L.; Wang, Z. A Novel Semi-Synthetic Andrographolide Analogue A5 Inhibits Tumor Angiogenesis via Blocking the VEGFR2-P38/ERK1/2 Signal Pathway. Biosci. Trends 2013, 7, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.-W.; Fu, H.-Z.; Luo, Y.-H.; Wei, X.-Y. Potential Anti-Angiogenic Sulfates of Andrographolide. J. Asian Nat. Prod. Res. 2013, 15, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.; Xu, W.; Liang, L.; Li, J.; Ding, X.; Chen, X.; Hu, J.; Lv, A.; Li, X. Clinical Efficacy of Andrographolide Sulfonate in the Treatment of Severe Hand, Foot, and Mouth Disease (HFMD) Is Dependent upon Inhibition of Neutrophil Activation. Phytother. Res. 2015, 29, 1161–1167. [Google Scholar] [CrossRef]
- Delvecchio, R.; Higa, L.; Pezzuto, P.; Valadão, A.; Garcez, P.; Monteiro, F.; Loiola, E.; Dias, A.; Silva, F.; Aliota, M.; et al. Chloroquine, an Endocytosis Blocking Agent, Inhibits Zika Virus Infection in Different Cell Models. Viruses 2016, 8, 322. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Liang, B.; Aarthy, M.; Singh, S.K.; Garg, N.; Mysorekar, I.U.; Giri, R. Hydroxychloroquine Inhibits Zika Virus NS2B-NS3 Protease. ACS Omega 2018, 3, 18132–18141. [Google Scholar] [CrossRef] [Green Version]
- Roldan, E.Q.; Biasiotto, G.; Magro, P.; Zanella, I. The Possible Mechanisms of Action of 4-Aminoquinolines (Chloroquine/Hydroxychloroquine) against SARS-Cov-2 Infection (COVID-19): A Role for Iron Homeostasis? Pharmacol. Res. 2020, 158, 104904. [Google Scholar] [CrossRef]
- Biot, C.; Daher, W.; Chavain, N.; Fandeur, T.; Khalife, J.; Dive, D.; De Clercq, E. Design and Synthesis of Hydroxyferroquine Derivatives with Antimalarial and Antiviral Activities. J. Med. Chem. 2006, 49, 2845–2849. [Google Scholar] [CrossRef] [Green Version]
- Vishvakarma, V.K.; Nand, B.; Kumar, V.; Kumari, K.; Bahadur, I.; Singh, P. Xanthene Based Hybrid Analogues to Inhibit Protease of Novel Corona Virus: Molecular Docking and ADMET Studies. Comput. Toxicol. 2020, 16, 100140. [Google Scholar] [CrossRef]
- Li, F.; Li, X.-M.; Sheng, D.; Chen, S.-R.; Nie, X.; Liu, Z.; Wang, D.; Zhao, Q.; Wang, Y.; Wang, Y.; et al. Discovery and Preliminary SAR of 14-Aryloxy-Andrographolide Derivatives as Antibacterial Agents with Immunosuppressant Activity. RSC Adv. 2018, 8, 9440–9456. [Google Scholar] [CrossRef] [Green Version]
- Richter, S.; Parolin, C.; Palumbo, M.; Palu, G. Antiviral Properties of Quinolone-Based Drugs. Curr. Drug Target-Infect. Disord. 2004, 4, 111–116. [Google Scholar] [CrossRef]
- Tran, Q.T.N.; Tan, W.S.D.; Wong, W.S.F.; Chai, C.L.L. Polypharmacology of Andrographolide: Beyond One Molecule One Target. Nat. Prod. Rep. 2021, 38, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Duann, Y.-F.; Yen, T.-L.; Chen, Y.-Y.; Jayakumar, T.; Ong, E.-T.; Sheu, J.-R. Andrographolide, a Novel NF-κB Inhibitor, Inhibits Vascular Smooth Muscle Cell Proliferation and Cerebral Endothelial Cell Inflammation. Acta Cardiol. Sin. 2014, 30, 308–315. [Google Scholar] [PubMed]
- Jin, Y.; Zhang, R.; Wu, W.; Duan, G. Antiviral and Inflammatory Cellular Signaling Associated with Enterovirus 71 Infection. Viruses 2018, 10, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikanth, L.; Sarma, P.V.G.K. Andrographolide Binds to Spike Glycoprotein and RNA-Dependent RNA Polymerase (NSP12) of SARS-CoV-2 by in Silico Approach: A Probable Molecule in the Development of Anti-Coronaviral Drug. J. Genet. Eng. Biotechnol. 2021, 19, 101. [Google Scholar] [CrossRef]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang, I.H.; Friman, O.; Guertin, D.A.; Chang, J.H.; Lindquist, R.A.; Moffat, J.; et al. CellProfiler: Image Analysis Software for Identifying and Quantifying Cell Phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Nuclei Count | Inhibition Rate (%) |
|---|---|---|
| 0.1% DMSO | 998 | |
| AGF-57 | 845 | 85.02 |
| AGL-16 | 991 | 86.63 |
| AGP-8e | 1076 | 51.10 |
| ZAF-46 | 1037 | 96.04 |
| AGP-7 | 813 | 90.13 |
| AGP-72 | 914 | 79.59 |
| ZAF-47 | 955 | 77.14 |
| AGP-49 | 945 | 77.30 |
| AGP-254 | 873 | 90.52 |
| AGP-196 | 795 | 66.49 |
| AGP-261 | 999 | 88.91 |
| AGP-177 | 668 | 88.24 |
| AGP-253 [43] | 902 | 98.91 |
| AGP-263 | 1006 | 96.52 |
| AGP-240a | 780 | 84.83 |
| AGP-240b | 819 | 86.69 |
| AGP-218 | 816 | 94.62 |
| AGP-208 | 775 | 97.77 |
| AGP-236a | 885 | 53.13 |
| Compound | ZAF-46 | ZAF-47 |
|---|---|---|
| CC50 (µM) | 29.91 | 29.57 |
| IC50 (µM) | 4.44 | 2.06 |
| Selectivity Index (SI) | 6.73 | 14.35 |
| Enterovirus | Species | Drug Treatment Duration (h) | IC50 (µM) |
|---|---|---|---|
| CV-A6 | Enterovirus A | 96 | 3.16 |
| CV-A16 | Enterovirus A | 16 | 1.38 |
| Echo7 | Enterovirus B | 12 | 6.70 |
| CV-B5 | Enterovirus B | 12 | 2.46 |
| CV-A24 | Enterovirus C | 12 | 1.09 |
| EV-D68 | Enterovirus D | 12 | 5.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, J.K.; Chen, R.; Lee, R.C.H.; Li, F.; Dai, K.; Zhou, G.-C.; Chu, J.J.H. Discovery of Novel Andrographolide Derivatives as Antiviral Inhibitors against Human Enterovirus A71. Pharmaceuticals 2022, 15, 115. https://doi.org/10.3390/ph15020115
Tan JK, Chen R, Lee RCH, Li F, Dai K, Zhou G-C, Chu JJH. Discovery of Novel Andrographolide Derivatives as Antiviral Inhibitors against Human Enterovirus A71. Pharmaceuticals. 2022; 15(2):115. https://doi.org/10.3390/ph15020115
Chicago/Turabian StyleTan, Jie Kai, Ran Chen, Regina Ching Hua Lee, Feng Li, Kun Dai, Guo-Chun Zhou, and Justin Jang Hann Chu. 2022. "Discovery of Novel Andrographolide Derivatives as Antiviral Inhibitors against Human Enterovirus A71" Pharmaceuticals 15, no. 2: 115. https://doi.org/10.3390/ph15020115
APA StyleTan, J. K., Chen, R., Lee, R. C. H., Li, F., Dai, K., Zhou, G.-C., & Chu, J. J. H. (2022). Discovery of Novel Andrographolide Derivatives as Antiviral Inhibitors against Human Enterovirus A71. Pharmaceuticals, 15(2), 115. https://doi.org/10.3390/ph15020115