
Targeted Anti-Biofilm Therapy: Dissecting Targets in the Biofilm Life Cycle


Abstract
1. Introduction
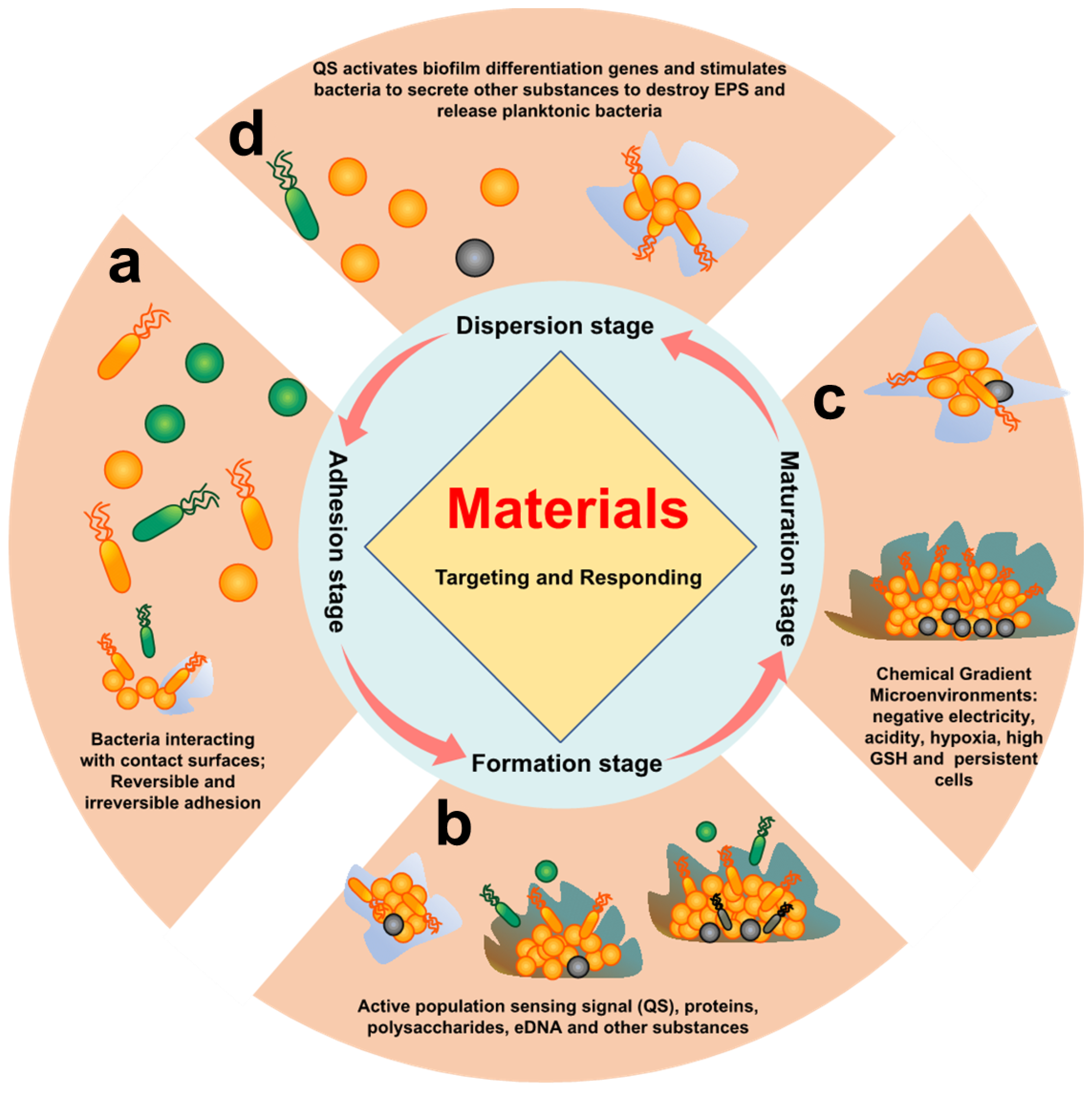
- Initial adhesion stage: The reversible adhesions are dominated by Lewis acid–base, van der Waals forces, electrostatic interactions, and hydrophilic–hydrophobic interactions [34,35]. Irreversible adhesion is triggered by the bacteria’s own adhesins and adhesion proteins [36]. Reversible and irreversible adhesion of bacteria to the surface is the main feature of this stage.
- Early biofilm formation stage: After bacteria adhere to the surface, bacteria activate their own metabolic pathways, which induces the bacteria to secrete metabolites (proteins, polysaccharides, eDNA etc.) to form EPS. At the same time, this also promotes bacteria-to-bacteria adhesion and activates quorum sensing (QS) [37]. Proteins, polysaccharides, eDNA, and QS of bacteria are the main features of this stage.
- Biofilm maturation stage: A complex spatial structure and a microenvironment with chemical gradients (acidity, hypoxia, high reduction, etc.) are gradually formed with the increase in EPS synthesized by bacteria. At the same time, some bacteria will enter a dormant and persistent state [38]. Therefore, the characteristics of this stage are mainly complex chemical gradient microenvironment, persistent cells, and dormant cells.
- Biofilm dispersion stage: Bacteria will secrete relevant secretions (enzymes, D-amino acids, surfactants, and other substances) to destroy EPS in response to nutrient deficiencies and accumulation of toxic substances, returning to a planktonic state [39]. This stage is characterized by associated secretions of bacteria and residual biofilm after dispersal.
2. Strategies of Targeting Initial Adhesion Stage
2.1. Inhibit Biofilm Formation
2.1.1. Targeted Adhesin Strategy
2.1.2. Interference Adhesion Strategy
2.1.3. Surface Anti-Adhesion Strategies
2.2. Targeting Biofilm Formation Strategy
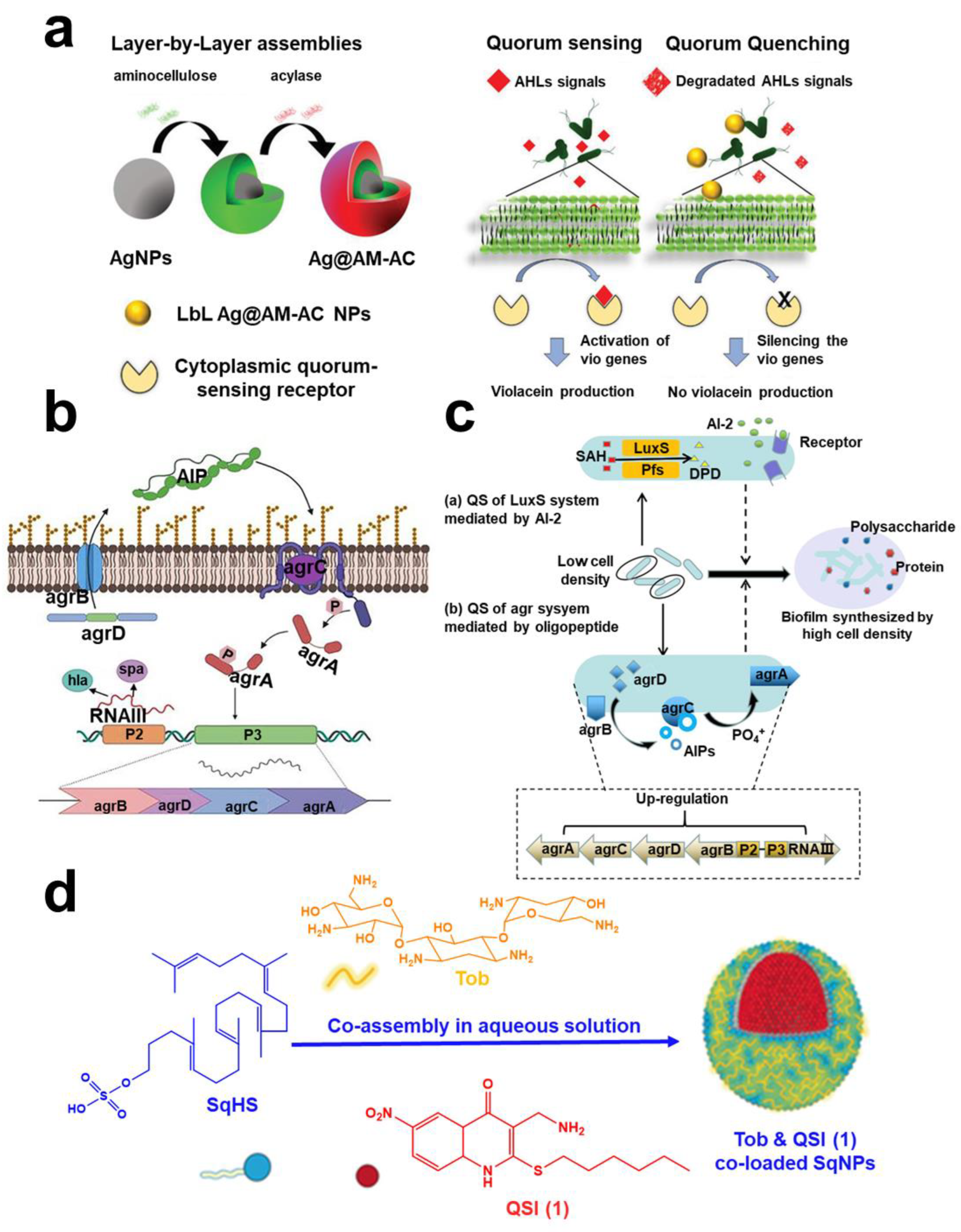
2.2.1. Quorum Quenching
2.2.2. Targeted Polysaccharide Strategy
2.2.3. Targeted eDNA Strategy
2.2.4. Targeted Protein Strategy
2.3. Targeting Strategy for Biofilm Maturation Stage
2.3.1. Targeted Persistent and Dormant Cells Strategy
2.3.2. The Intelligent Release of Microenvironment Response Strategy
Hypoxic
Low pH

Negative
Overexpression GSH

Hydrogen Peroxide
2.3.3. Other Targeting Strategies
Magnetic Targeting Technology
Phage-Targeting

Probiotic Targeting
Gene Targeting
Metabolic Targeting
2.4. Targeted Strategy for Biofilm Dispersion Stage
2.4.1. Active Dispersion Biofilm
2.4.2. Control Biofilm Dispersion

3. Conclusions and Perspectives for Future Research
- The key barrier is the in vivo biofilm model, especially for a multi-species biofilm model. In this regard, substantive research on the targeted techniques should be conducted before entering the clinic; it is extremely important to implement techniques that can accurately target the objective in multi-species biofilm.
- Further implementation of differential targeting of pathogenic bacteria and probiotics is highly beneficial and is expected to facilitate rapid development of immunotherapies.
- To clarify the metabolic pathway of targeted techniques under host pathological conditions, it is necessary to develop targeted techniques with long-term visualization or monitoring.
- Currently, targeting techniques target different phases of biofilms. Could there be a technique to observe the biofilm phase in patients to make treatment plans more effective?
- The biological effects of targeting technology among materials, cells, and bacteria are very worthy of study.
- Currently, small molecules of targeted inhibitors have the potential to replace antibiotics for treatment, but antibiotics have a chiral structure. Research on the combination of targeted inhibitors and stereochemistry may be a new generation of antibiotic research and development route.
- Targeting technology is needed to meet clinical needs. Cost-effective, simplified, and economical amplification preparation strategies need to be widely studied.
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kolpen, M.; Kragh, K.N.; Enciso, J.B.; Faurholt-Jepsen, D.; Lindegaard, B.; Egelund, G.B.; Jensen, A.V.; Ravn, P.; Mathiesen, I.H.M.; Gheorge, A.G.; et al. Bacterial Biofilms Predominate in Both Acute and Chronic Human Lung Infections. Thorax 2022, 77, 1015–1022. [Google Scholar] [CrossRef]
- Tang, Y.; Huang, Q.X.; Zheng, D.W.; Chen, Y.; Ma, L.; Huang, C.; Zhang, X.Z. Engineered Bdellovibrio bacteriovorus: A Countermeasure for Biofilm-Induced Periodontitis. Mater. Today 2022, 53, 71–83. [Google Scholar] [CrossRef]
- Silva, M.D.; Sillankorva, S. Otitis Media Pathogens—A Life Entrapped in Biofilm Communities. Crit. Rev. Microbiol. 2019, 45, 595–612. [Google Scholar] [CrossRef]
- Lerche, C.J.; Schwartz, F.; Theut, M.; Fosbøl, E.L.; Iversen, K.; Bundgaard, H.; Høiby, N.; Moser, C. Anti-Biofilm Approach in Infective Endocarditis Exposes New Treatment Strategies for Improved Outcome. Front. Cell Dev. Biol. 2021, 9, 643335. [Google Scholar] [CrossRef]
- Raheem Lateef Al-awsi, G.; Al-Hadeithi, Z.S.M.; Abdalkareem Jasim, S.; Alkhudhairy, M.K.; Ghasemian, A. Virulence Traits and Plasmid-Mediated Quinolone Resistance among Aggregatibacter actinomycetemcomitans from Iraq: Low Rate of Highly Virulent JP2 Genotype. Microb. Pathog. 2022, 164, 105438. [Google Scholar] [CrossRef]
- Xiao, Y.; Jiang, S.C.; Wang, X.; Muhammad, T.; Song, P.; Zhou, B.; Zhou, Y.; Li, Y. Mitigation of Biofouling in Agricultural Water Distribution Systems with Nanobubbles. Environ. Int. 2020, 141, 105787. [Google Scholar] [CrossRef]
- Winters, H.; Eu, H.G.; Li, S.; Alpatova, A.; Alshahri, A.H.; Nasar, N.; Ghaffour, N. Biofouling of Seawater Reverse Osmosis Membranes Caused by Dispersal of Planktonic Particulate Bacterial Aggregates (Protobiofilms) from Rotary Energy Recovery Devices. Desalination 2022, 529, 115647. [Google Scholar] [CrossRef]
- Pichardo-Romero, D.; Garcia-Arce, Z.P.; Zavala-Ramírez, A.; Castro-Muñoz, R. Current Advances in Biofouling Mitigation in Membranes for Water Treatment: An Overview. Processes 2020, 8, 137. [Google Scholar] [CrossRef]
- Ricart, M.; Guasch, H.; Barceló, D.; Brix, R.; Conceição, M.H.; Geiszinger, A.; de Alda, M.J.L.; López-Doval, J.C.; Muñoz, I.; Postigo, C.; et al. Primary and Complex Stressors in Polluted Mediterranean Rivers: Pesticide Effects on Biological Communities. J. Hydrol. 2010, 383, 52–61. [Google Scholar] [CrossRef]
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria Monocytogenes in Fresh Produce: Outbreaks, Prevalence and Contamination Levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef]
- Habash, M.; Reid, G. Microbial Biofilms: Their Development and Significance for Medical Device-Related Infections. J. Clin. Pharmacol. 1999, 39, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Schramm, A.; De Beer, D.; Gieseke, A.; Amann, R. Microenvironments and Distribution of Nitrifying Bacteria in a Membrane-Bound Biofilm. Environ. Microbiol. 2000, 2, 680–686. [Google Scholar] [CrossRef]
- Kolenbrander, P.E. Oral Microbial Communities: Biofilms, Interactions, and Genetic Systems. Annu. Rev. Virol. 2000, 54, 413–437. [Google Scholar] [CrossRef]
- Periasamy, S.; Joo, H.S.; Duong, A.C.; Bach, T.H.L.; Tan, V.Y.; Chatterjee, S.S.; Cheung, G.Y.C.; Otto, M. How Staphylococcus aureus Biofilms Develop Their Characteristic Structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1281–1286. [Google Scholar] [CrossRef]
- Sutherland, I.W. The Biofilm Matrix-An Immobilized but Dynamic Microbial Environment. Trends Microbiol. 2001, 9, 222–227. [Google Scholar] [CrossRef]
- Decho, A.W. Microbial Biofilms in Intertidal Systems: An Overview. Cont. Shelf Res. 2000, 20, 1257–1273. [Google Scholar] [CrossRef]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting Microbial Biofilms: Current and Prospective Therapeutic Strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef]
- Lewis, K. Multidrug Tolerance of Biofilms and Persister Cells. Curr. Top. Microbiol. Immunol. 2008, 322, 107–131. [Google Scholar] [CrossRef]
- Allison, K.R.; Brynildsen, M.P.; Collins, J.J. Metabolite-Enabled Eradication of Bacterial Persisters by Aminoglycosides. Nature 2011, 473, 216–220. [Google Scholar] [CrossRef]
- Keren, I.; Shah, D.; Spoering, A.; Kaldalu, N.; Lewis, K. Specialized Persister Cells and the Mechanism of Multidrug Tolerance in Escherichia coli. J. Bacteriol. 2004, 186, 8172–8180. [Google Scholar] [CrossRef]
- Fry, D.E. Antimicrobial Peptides. Surg. Infect. 2018, 19, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister Cells, Dormancy and Infectious Disease. Nat. Rev. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic Resistance of Bacterial Biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef]
- Hunter, R.C.; Beveridge, T.J. Application of a PH-Sensitive Fluoroprobe (C-SNARF-4) for PH Microenvironment Analysis in Pseudomonas aeruginosa Biofilms. Appl. Environ. Microbiol. 2005, 71, 2501–2510. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Falsetta, M.L.; Klein, M.I. The Exopolysaccharide Matrix: A Virulence Determinant of Cariogenic Biofilm. J. Dent. Res. 2013, 92, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Alhede, M.; Alhede, M.; Eickhardt-Sørensen, S.R.; Moser, C.; Kühl, M.; Jensen, P.Ø.; Høiby, N. The In Vivo Biofilm. Trends Microbiol. 2013, 21, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Hammer, B.K.; Bassler, B.L. Quorum Sensing Controls Biofilm Formation in Vibrio cholerae. Mol. Microbiol. 2003, 50, 101–104. [Google Scholar] [CrossRef]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial Extracellular Polysaccharides Involved in Biofilm Formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P. Evolving Concepts in Biofilm Infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Ciofu, O.; Molin, S.; Givskov, M.; Høiby, N. Applying Insights from Biofilm Biology to Drug Development-Can a New Approach Be Developed? Nat. Rev. Drug Discov. 2013, 12, 791–808. [Google Scholar] [CrossRef] [PubMed]
- O’Gara, J.P.; Humphreys, H. Staphylococcus epidermidis Biofilms: Importance and Implications. J. Med. Microbiol. 2001, 50, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Zachary, K.Z.; Mattew, L.B. Antimicrobial and Antifouling Strategies for Polymeric Medical Devices. ACS Macro. Lett. 2018, 7, 16–25. [Google Scholar] [CrossRef]
- Dunne, W.M. Bacterial adhesion: Seen any good biofilms lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef]
- Fu, J.; Zhang, Y.; Zhang, W.; Shu, G.; Lin, J.; Xu, F.; Tang, H.; Peng, G.; Zhao, L.; Chen, S.; et al. Strategies for Interfering With Bacterial Early Stage Biofilms. Clin. Front. Microbiol. 2021, 12, 675843. [Google Scholar] [CrossRef]
- Musa, H.M.; Aisha, L.I.; Xiao, F.; Yachong, G.; Yiyan, Y.; Xu, J.; Junzhi, Q.; Xiong, G.; Tianpei, H. Beyond Risk: Bacterial Biofilms and Their Regulating Approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef]
- Armbruster, C.; Parsek, M. New insight into the early stages of biofilm formation. Proc. Natl. Acad. Sci. USA 2018, 115, 4317–4319. [Google Scholar] [CrossRef] [PubMed]
- Conlon, B.P.; Nakayasu, E.S.; Fleck, L.E.; LaFleur, M.D.; Isabella, V.M.; Coleman, S.N.; Smith, R.D.; Adkins, J.N.; Lewis, K. Activated ClpP kills persisters and eradicates a chronic biofilm infection. Nat. Prod. Rep. 2013, 503, 365–370. [Google Scholar] [CrossRef]
- Rumbaugh, K.P.; Sauer, K. Biofilm dispersion. Nat. Rev. Microbiol. 2020, 18, 571–586. [Google Scholar] [CrossRef]
- Kindler, O.; Pulkkinen, O.; Cherstvy, A.G.; Metzler, R. Burst Statistics in an Early Biofilm Quorum Sensing Model: The Role of Spatial Colony-Growth Heterogeneity. Sci. Rep. 2019, 9, 12077. [Google Scholar] [CrossRef]
- Sharma, A.; Wood, K.B. Spatial Segregation and Cooperation in Radially Expanding Microbial Colonies under Antibiotic Stress. ISME J. 2021, 15, 3019–3033. [Google Scholar] [CrossRef] [PubMed]
- Ruhal, R.; Kataria, R. Biofilm Patterns in Gram-Positive and Gram-Negative Bacteria. Microbiol. Res. 2021, 251, 126829. [Google Scholar] [CrossRef]
- Nadell, C.D.; Drescher, K.; Foster, K.R. Spatial Structure, Cooperation and Competition in Biofilms. Nat. Rev. Microbiol. 2016, 14, 589–600. [Google Scholar] [CrossRef]
- Wu, H.; Moser, C.; Wang, H.Z.; Høiby, N.; Song, Z.J. Strategies for Combating Bacterial Biofilm Infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Doub, J.B. Bacteriophage Therapy for Clinical Biofilm Infections: Parameters That Influence Treatment Protocols and Current Treatment Approaches. Antibiotics 2020, 9, 799. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.N.; Wang, J.; Ren, K.F.; Ji, J. Functionalized Biomaterials to Combat Biofilms. Biomater. Sci. 2020, 8, 4052–4066. [Google Scholar] [CrossRef]
- Contreras-García, A.; Bucioa, E.; Brackmanc, G.; Coenyec, T.; Concheirob, A.; Alvarez-Lorenzob, C. Biofilm Inhibition and Drug-Eluting Properties of Novel DMAEMA-Modified Polyethylene and Silicone Rubber Surfaces. Biofouling 2011, 27, 123–135. [Google Scholar] [CrossRef]
- Yang, K.; Shi, J.; Wang, L.; Chen, Y.; Liang, C.; Yang, L.; Wang, L.N. Bacterial Anti-Adhesion Surface Design: Surface Patterning, Roughness and Wettability: A Review. J. Mater. Sci. Technol. 2022, 99, 82–100. [Google Scholar] [CrossRef]
- Busscher, H.J.; van der Mei, H.C.; Subbiahdoss, G.; Jutte, P.C.; van den Dungen, J.J.A.M.; Zaat, S.A.J.; Schultz, M.J.; Grainger, D.W. Biomaterial-Associated Infection: Locating the Finish Line in the Race for the Surface. Sci. Transl. Med. 2012, 4, 153rv10. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Z.; Ren, J.; Qu, X. Enzyme Mimicry for Combating Bacteria and Biofilms. Acc. Chem. Res. 2018, 51, 789–799. [Google Scholar] [CrossRef]
- Xiao, X.; Zhao, W.; Liang, J.; Sauer, K.; Libera, M. Self-Defensive Antimicrobial Biomaterial Surfaces. Colloids Surf. B 2020, 192, 110989. [Google Scholar] [CrossRef] [PubMed]
- Steenackers, H.P.; Parijs, I.; Foster, K.R.; Vanderleyden, J. Experimental Evolution in Biofilm Populations. FEMS Microbiol. Rev. 2016, 40, 373–397. [Google Scholar] [CrossRef] [PubMed]
- Thaarup, I.C.; Iversen, A.K.S.; Lichtenberg, M.; Bjarnsholt, T.; Jakobsen, T.H. Biofilm Survival Strategies in Chronic Wounds. Microorganisms 2022, 10, 775. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Ikegami, A.; Kuramitsu, H.K. Synergistic Biofilm Formation by Treponema denticola and Porphyromonas gingivalis. FEMS Microbiol. Lett. 2005, 250, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; Palmer, R.J.; Blehert, D.S.; Campagna, S.R.; Semmelhack, M.F.; Egland, P.G.; Bassler, B.L.; Kolenbrander, P.E. Autoinducer 2: A Concentration-Dependent Signal for Mutualistic Bacterial Biofilm Growth. Mol. Microbiol. 2006, 60, 1446–1456. [Google Scholar] [CrossRef]
- Kumada, M.; Motegi, M.; Nakao, R.; Yonezawa, H.; Yamamura, H.; Tagami, J.; Senpuku, H. Inhibiting Effects of Enterococcus faecium Non-Biofilm Strain on Streptococcus mutans Biofilm Formation. J. Microbiol. Immunol. Infect. 2009, 42, 188–196. [Google Scholar]
- Rao, D.; Webb, J.S.; Kjelleberg, S. Competitive Interactions in Mixed-Species Biofilms Containing the Marine Bacterium Pseudoalteromonas tunicata. Appl. Environ. Microbiol. 2005, 71, 1729–1736. [Google Scholar] [CrossRef]
- Srinivasan, R.; Santhakumari, S.; Poonguzhali, P.; Geetha, M.; Dyavaiah, M.; Xiangmin, L. Bacterial Biofilm Inhibition: A Focused Review on Recent Therapeutic Strategies for Combating the Biofilm Mediated Infections. Front. Microbiol. 2021, 12, 676458. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Uruén, C.; Chopo-Escuin, G.; Tommassen, J.; Mainar-Jaime, R.C.; Arenas, J. Antibiotics Biofilms as Promoters of Bacterial Antibiotic Resistance and Tolerance. Antibiotics 2020, 10, 3. [Google Scholar] [CrossRef]
- Jiang, Q.; Yu, Y.; Xu, R.; Zhang, Z.; Liang, C.; Sun, H.; Deng, F.; Yu, X. The Temporal Shift of Peri-Implant Microbiota during the Biofilm Formation and Maturation in a Canine Model. Microb. Pathog. 2021, 158, 105100. [Google Scholar] [CrossRef]
- Urwin, L.; Okurowska, K.; Crowther, G.; Roy, S.; Garg, P.; Karunakaran, E.; MacNeil, S.; Partridge, L.J.; Green, L.R.; Monk, P.N. Corneal Infection Models: Tools to Investigate the Role of Biofilms in Bacterial Keratitis. Cells 2020, 9, 2450. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Zhai, Z.; Gao, C. Adaptive Antibacterial Biomaterial Surfaces and Their Applications. Mater. Today Bio 2019, 2, 100017. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, J.; Peng, Q.; Liu, Y.; Lei, L.; Zhang, H. The Role of Staphylococcus aureus YycFG in Gene Regulation, Biofilm Organization and Drug Resistance. Antibiotics 2021, 10, 1555. [Google Scholar] [CrossRef] [PubMed]
- Solanki, V.; Tiwari, M.; Tiwari, V. Host-Bacteria Interaction and Adhesin Study for Development of Therapeutics. Int. J. Biol. Macromol. 2018, 112, 54–64. [Google Scholar] [CrossRef]
- Chauhan, N.; Hatlem, D.; Orwick-Rydmark, M.; Schneider, K.; Floetenmeyer, M.; van Rossum, B.; Leo, J.C.; Linke, D. Insights into the Autotransport Process of a Trimeric Autotransporter, Yersinia Adhesin A (YadA). Mol. Microbiol. 2019, 111, 844–862. [Google Scholar] [CrossRef] [PubMed]
- Asadi, A.; Razavi, S.; Talebi, M.; Gholami, M. A Review on Anti-Adhesion Therapies of Bacterial Diseases. Infection 2019, 47, 13–23. [Google Scholar] [CrossRef]
- Monserrat-Martinez, A.; Gambin, Y.; Sierecki, E. Thinking Outside the Bug: Molecular Targets and Strategies to Overcome Antibiotic Resistance. Int. J. Mol. Sci. 2019, 20, 1255. [Google Scholar] [CrossRef]
- Filipović, U.; Dahmane, R.G.; Ghannouchi, S.; Zore, A.; Bohinc, K. Bacterial Adhesion on Orthopedic Implants. Adv. Colloid Interface Sci. 2020, 283, 102228. [Google Scholar] [CrossRef]
- Grosheva, I.; Zheng, D.; Levy, M.; Polansky, O.; Lichtenstein, A.; Golani, O.; Dori-Bachash, M.; Moresi, C.; Shapiro, H.; del Mare-Roumani, S.; et al. High-Throughput Screen Identifies Host and Microbiota Regulators of Intestinal Barrier Function. Gastroenterology 2020, 159, 1807–1823. [Google Scholar] [CrossRef]
- Paxman, J.J.; Lo, A.W.; Sullivan, M.J.; Panjikar, S.; Kuiper, M.; Whitten, A.E.; Wang, G.; Luan, C.H.; Moriel, D.G.; Tan, L.; et al. Unique Structural Features of a Bacterial Autotransporter Adhesin Suggest Mechanisms for Interaction with Host Macromolecules. Nat. Commun. 2019, 10, 1967. [Google Scholar] [CrossRef] [PubMed]
- Rao, L.; Sheng, Y.; Zhang, J.; Xu, Y.; Yu, J.; Wang, B.; Zhao, H.; Wang, X.; Guo, Y.; Wu, X.; et al. Small-Molecule Compound SYG-180-2-2 to Effectively Prevent the Biofilm Formation of Methicillin-Resistant Staphylococcus aureus. Front. Microbiol. 2022, 12, 770657. [Google Scholar] [CrossRef] [PubMed]
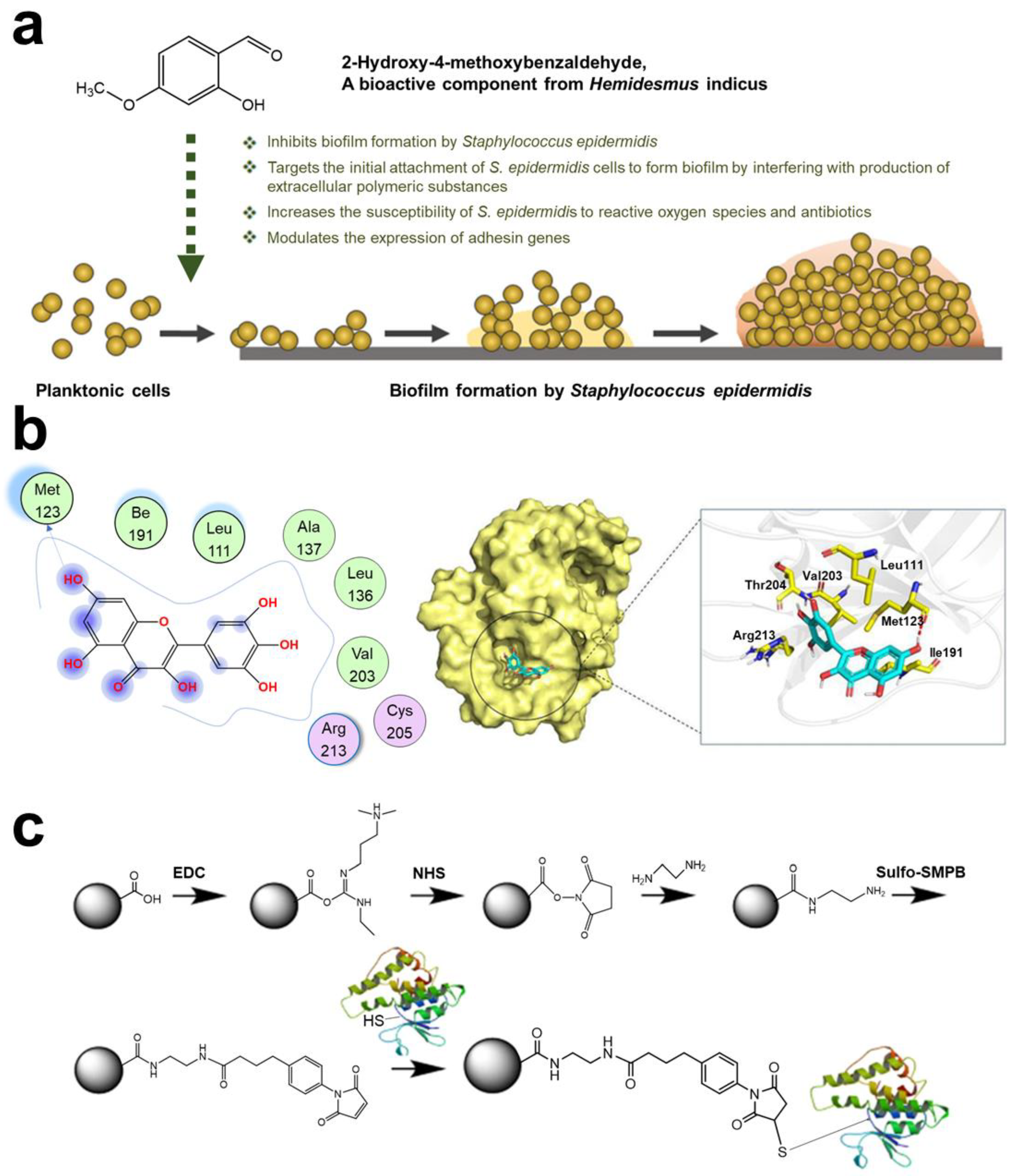
- Kannappan, A.; Durgadevi, R.; Srinivasan, R.; Lagoa, R.J.L.; Packiavathy, I.A.S.V.; Pandian, S.K.; Veera Ravi, A. 2-Hydroxy-4-Methoxybenzaldehyde from Hemidesmus indicus Is Antagonistic to Staphylococcus epidermidis Biofilm Formation. Biofouling 2020, 36, 549–563. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Lv, B.; Yang, K.; Lu, Z.; Ma, J. Discovery of Myricetin as an Inhibitor against Streptococcus mutans and an Anti-Adhesion Approach to Biofilm Formation. Int. J. Med. Microbiol. 2021, 311, 151512. [Google Scholar] [CrossRef]
- Liu, C.; Sun, D.; Liu, J.; Chen, Y.; Zhou, X.; Ru, Y.; Zhu, J.; Liu, W. CAMP and C-Di-GMP Synergistically Support Biofilm Maintenance through the Direct Interaction of Their Effectors. Nat. Commun. 2022, 13, 1493. [Google Scholar] [CrossRef]
- Awadelkareem, A.M.; Al-Shammari, E.; Elkhalifa, A.O.; Adnan, M.; Siddiqui, A.J.; Mahmood, D.; Azad, Z.R.A.A.; Patel, M.; Mehmood, K.; Danciu, C.; et al. Anti-Adhesion and Antibiofilm Activity of Eruca Sativa Miller Extract Targeting Cell Adhesion Proteins of Food-Borne Bacteria as a Potential Mechanism: Combined In Vitro-In Silico Approach. Plants 2022, 11, 610. [Google Scholar] [CrossRef]
- Huebinger, R.M.; Stones, D.H.; de Souza Santos, M.; Carlson, D.L.; Song, J.; Vaz, D.P.; Keen, E.; Wolf, S.E.; Orth, K.; Krachler, A.M. Targeting Bacterial Adherence Inhibits Multidrug-Resistant Pseudomonas aeruginosa Infection Following Burn Injury. Sci. Rep. 2016, 6, 39341. [Google Scholar] [CrossRef]
- Capeletti, L.B.; de Oliveira, J.F.A.; Loiola, L.M.D.; Galdino, F.E.; da Silva Santos, D.E.; Soares, T.A.; de Oliveira Freitas, R.; Cardoso, M.B. Gram-Negative Bacteria Targeting Mediated by Carbohydrate–Carbohydrate Interactions Induced by Surface-Modified Nanoparticles. Adv. Funct. Mater. 2019, 29, 1904216. [Google Scholar] [CrossRef]
- Sarshar, M.; Behzadi, P.; Ambrosi, C.; Zagaglia, C.; Palamara, A.T.; Scribano, D. FimH and Anti-Adhesive Therapeutics: A Disarming Strategy against Uropathogens. Antibiotics 2020, 9, 397. [Google Scholar] [CrossRef]
- Cusumano, Z.T.; Klein, R.D.; Hultgren, S.J. Innovative Solutions to Sticky Situations: Antiadhesive Strategies for Treating Bacterial Infections. Microbiol. Spectr. 2016, 4, 4.2.07. [Google Scholar] [CrossRef]
- Schembri, M.A.; Hasman, H.; Klemm, P. Expression and Purification of the Mannose Recognition Domain of the FimH Adhesin. FEMS Microbiol. Lett. 2000, 188, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Foroogh, N.; Rezvan, M.; Ahmad, K.; Mahmood, S. Structural and Functional Characterization of the FimH Adhesin of Uropathogenic Escherichia coli and Its Novel Applications. Microb. Pathog. 2021, 161, 105288. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Betz, P.; Sun, Y.; Gorb, S.N.; Lindhorst, T.K.; Krueger, A. Saccharide-Modified Nanodiamond Conjugates for the Efficient Detection and Removal of Pathogenic Bacteria. Chemistry 2012, 18, 6485–6492. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, A.; Jiménez-Barbero, J.; Casnati, A.; de Castro, C.; Darbre, T.; Fieschi, F.; Finne, J.; Funken, H.; Jaeger, K.E.; Lahmann, M.; et al. Multivalent Glycoconjugates as Anti-Pathogenic Agents. Chem. Soc. Rev. 2013, 42, 4709–4727. [Google Scholar] [CrossRef] [PubMed]
- Speziale, P.; Arciola, C.R.; Pietrocola, G. Fibronectin and Its Role in Human Infective Diseases. Cells 2019, 8, 1516. [Google Scholar] [CrossRef]
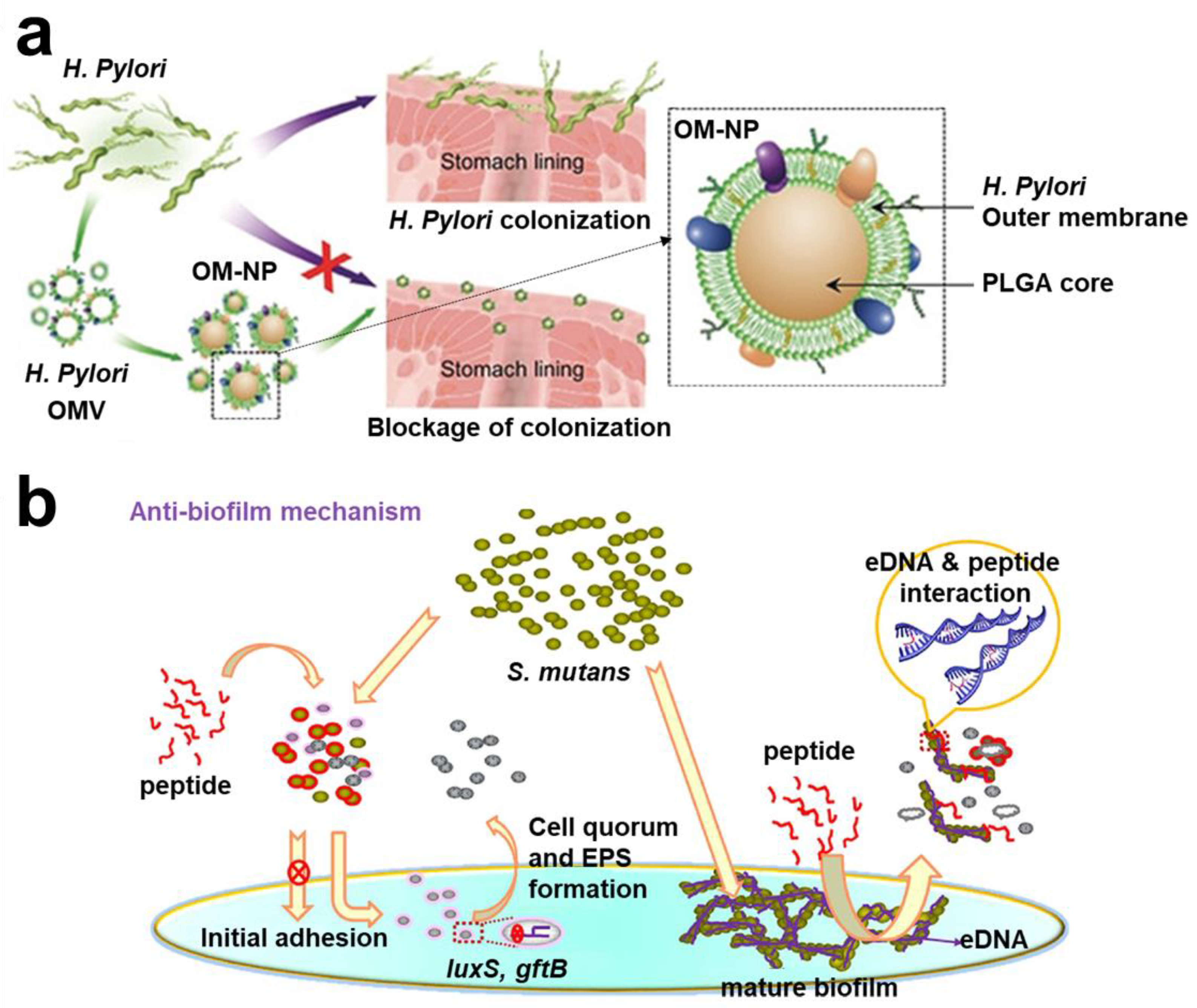
- Zhang, Y.; Chen, Y.; Lo, C.; Zhuang, J.; Angsantikul, P.; Zhang, Q.; Wei, X.; Zhou, Z.; Obonyo, M.; Fang, R.H.; et al. Inhibition of Pathogen Adhesion by Bacterial Outer Membrane-Coated Nanoparticles. Angew. Chem. Int. Ed. Engl. 2019, 58, 11404–11408. [Google Scholar] [CrossRef]
- Guo, S.; Zahiri, H.; Stevens, C.; Spaanderman, D.C.; Milroy, L.G.; Ottmann, C.; Brunsveld, L.; Voets, I.K.; Davies, P.L. Molecular Basis for Inhibition of Adhesin-Mediated Bacterial-Host Interactions through a Peptide-Binding Domain. Cell Rep. 2021, 37, 110002. [Google Scholar] [CrossRef]
- An, S.J.; Namkung, J.U.; Ha, K.W.; Jun, H.K.; Kim, H.Y.; Choi, B.K. Inhibitory Effect of D-Arabinose on Oral Bacteria Biofilm Formation on Titanium Discs. Anaerobe 2022, 75, 102533. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, C.; Chen, J.; Zhou, S.; Zhao, Y.; Xu, M.; Xu, H. Dual Mode of Anti-Biofilm Action of G3 against Streptococcus mutans. ACS Appl. Mater. Interfaces 2020, 12, 27866–27875. [Google Scholar] [CrossRef]
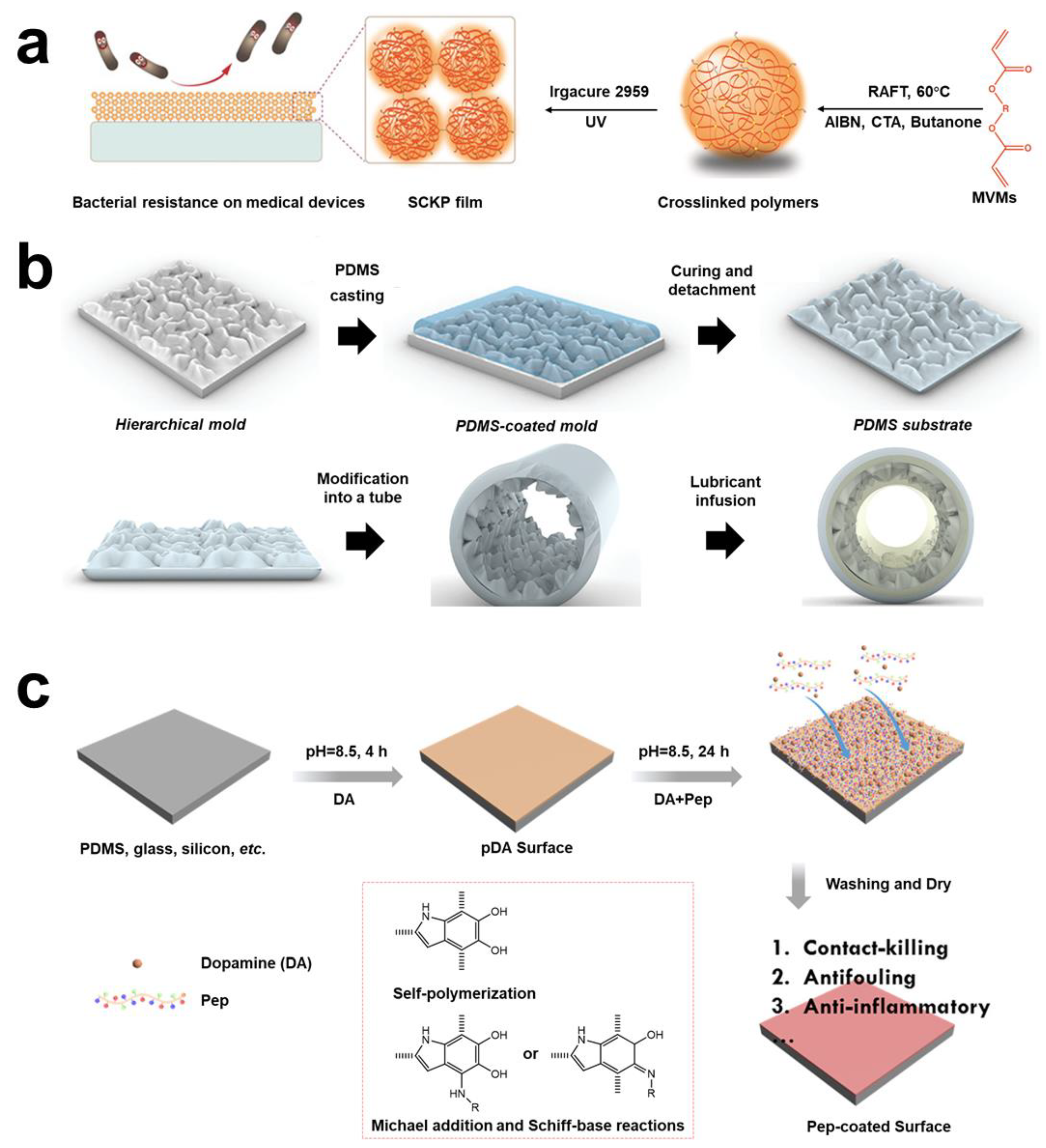
- Khoo, X.; Grinstaff, M.W. Novel Infection-Resistant Surface Coatings: A Bioengineering Approach. MRS Bull. 2011, 36, 357–366. [Google Scholar] [CrossRef]
- Xu, Q.; A, S.; Venet, M.; Gao, Y.; Zhou, D.; Wang, W.; Zeng, M.; Rotella, C.; Li, X.; Wang, X.; et al. Bacteria-Resistant Single Chain Cyclized/Knotted Polymer Coatings. Angew. Chem. Int. Ed. Engl. 2019, 58, 10616–10620. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Jin, J.; Park, S.; Kim, J.Y.; Lee, M.J.; Sun, H.; Kwon, J.S.; Lee, H.; Choi, S.H.; Hong, J. Quantitative Interpretation of Hydration Dynamics Enabled the Fabrication of a Zwitterionic Antifouling Surface. ACS Appl. Mater. Interfaces 2020, 12, 7951–7965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, J.; Yang, M.; Huang, L.; Bu, F.; Xie, Z.; Li, G.; Wang, X. Inserting Menthoxytriazine into Poly (Ethylene Terephthalate) for Inhibiting Microbial Adhesion. ACS Biomater. Sci. Eng. 2022, 8, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Tullii, G.; Donini, S.; Bossio, C.; Lodola, F.; Pasini, M.; Parisini, E.; Galeotti, F.; Antognazza, M.R. Micro-And Nanopatterned Silk Substrates for Antifouling Applications. ACS Appl. Mater. Interfaces 2020, 12, 5437–5446. [Google Scholar] [CrossRef]
- Kayes, M.I.; Galante, A.J.; Stella, N.A.; Haghanifar, S.; Shanks, R.M.Q.; Leu, P.W. Stable Lotus Leaf-Inspired Hierarchical, Fluorinated Polypropylene Surfaces for Reduced Bacterial Adhesion. React. Funct. Polym. 2018, 128, 40–46. [Google Scholar] [CrossRef]
- Khan, S.; Jarad, N.A.; Ladouceur, L.; Rachwalski, K.; Bot, V.; Shakeri, A.; Maclachlan, R.; Sakib, S.; Weitz, J.I.; Brown, E.D.; et al. Transparent and Highly Flexible Hierarchically Structured Polydimethylsiloxane Surfaces Suppress Bacterial Attachment and Thrombosis Under Static and Dynamic Conditions. Small 2022, 18, 2108112. [Google Scholar] [CrossRef]
- Liu, C.; He, Q.; Song, D.; Jackson, J.; Faria, A.F.; Jiang, X.; Li, X.; Ma, J.; Sun, Z. Electroless Deposition of Copper Nanoparticles Integrates Polydopamine Coating on Reverse Osmosis Membranes for Efficient Biofouling Mitigation. Water. Res. 2022, 217, 118375. [Google Scholar] [CrossRef]
- Gao, Q.; Li, X.; Yu, W.; Jia, F.; Yao, T.; Jin, Q.; Ji, J. Fabrication of Mixed-Charge Polypeptide Coating for Enhanced Hemocompatibility and Anti-Infective Effect. ACS Appl. Mater. Interfaces 2020, 12, 2999–3010. [Google Scholar] [CrossRef]
- Waters, C.M.; Bassler, B.L. Quorum Sensing: Cell-to-Cell Communication in Bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef]
- Spangler, J.R.; Dean, S.N.; Leary, D.H.; Walper, S.A. Response of Lactobacillus plantarum WCFS1 to the Gram-Negative Pathogen-Associated Quorum Sensing Molecule N-3-Oxododecanoyl Homoserine Lactone. Front. Microbiol. 2019, 10, 715. [Google Scholar] [CrossRef]
- Hense, B.A.; Schuster, M. Core Principles of Bacterial Autoinducer Systems. Microbiol. Mol. Biol. Rev. 2015, 79, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Bassler, B.L. Bacterial Quorum Sensing in Complex and Dynamically Changing Environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum Sensing Signal-Response Systems in Gram-Negative Bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C. Quorum Sensing Inhibitors: An Overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Givskov, M. Quorum-Sensing Inhibitors as Anti-Pathogenic Drugs. Int. J. Med. Microbiol. 2006, 296, 149–161. [Google Scholar] [CrossRef]
- Saipriya, K.; Swathi, C.H.; Ratnakar, K.S.; Sritharan, V. Quorum-Sensing System in Acinetobacter baumannii: A Potential Target for New Drug Development. J. Appl. Microbiol. 2020, 128, 15–27. [Google Scholar] [CrossRef]
- Christensen, Q.H.; Grove, T.L.; Booker, S.J.; Greenberg, E.P. A High-Throughput Screen for Quorum-Sensing Inhibitors That Target Acyl-Homoserine Lactone Synthases. Proc. Natl. Acad. Sci. USA 2013, 110, 13815–13820. [Google Scholar] [CrossRef]
- Piewngam, P.; Chiou, J.; Chatterjee, P.; Otto, M. Alternative Approaches to Treat Bacterial Infections: Targeting Quorum-Sensing. Expert Rev. Anti-Infect. Ther. 2020, 18, 499–510. [Google Scholar] [CrossRef]
- Tan, S.Y.Y.; Chua, S.L.; Chen, Y.; Rice, S.A.; Kjelleberg, S.; Nielsen, T.E.; Yang, L.; Givskov, M. Identification of Five Structurally Unrelated Quorum-Sensing Inhibitors of Pseudomonas aeruginosa from a Natural-Derivative Database. Antimicrob. Agents Chemother. 2013, 57, 5629–5641. [Google Scholar] [CrossRef]
- Chen, G.; Swem, L.R.; Swem, D.L.; Stauff, D.L.; O’Loughlin, C.T.; Jeffrey, P.D.; Bassler, B.L.; Hughson, F.M. A Strategy for Antagonizing Quorum Sensing. Mol. Cell 2011, 42, 199–209. [Google Scholar] [CrossRef]
- Ivanova, A.; Ivanova, K.; Tied, A.; Heinze, T.; Tzanov, T. Layer-By-Layer Coating of Aminocellulose and Quorum Quenching Acylase on Silver Nanoparticles Synergistically Eradicate Bacteria and Their Biofilms. Adv. Funct. Mater. 2020, 30, 2001284. [Google Scholar] [CrossRef]
- De Celis, M.; Serrano-Aguirre, L.; Belda, I.; Liébana-García, R.; Arroyo, M.; Marquina, D.; de la Mata, I.; Santos, A. Acylase Enzymes Disrupting Quorum Sensing Alter the Transcriptome and Phenotype of Pseudomonas aeruginosa, and the Composition of Bacterial Biofilms from Wastewater Treatment Plants. Sci Total. Environ. 2021, 799, 149401. [Google Scholar] [CrossRef]
- Kim, M.K. Staphylococcus aureus Toxins: From Their Pathogenic Roles to Anti-Virulence Therapy Using Natural Products. Biotechnol. Bioproc. E 2019, 24, 424–435. [Google Scholar] [CrossRef]
- Lu, H.; Tu, C.; Zhou, T.; Zhang, W.; Zhan, Y.; Ding, J.; Wu, X.; Yang, Z.; Cao, W.; Deng, L.; et al. A ROS-Scavenging Hydrogel Loaded with Bacterial Quorum Sensing Inhibitor Hyperbranched Poly-L-Lysine Promotes the Wound Scar-Free Healing of Infected Skin in Vivo. Chem. Eng. J. 2022, 436, 135130. [Google Scholar] [CrossRef]
- Yuan, Q.; Feng, W.; Wang, Y.; Wang, Q.; Mou, N.; Xiong, L.; Wang, X.; Xia, P.; Sun, F. Luteolin Attenuates the Pathogenesis of Staphylococcus aureus by Interfering with the Agr System. Microb. Pathog. 2022, 165, 105496. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; El-Naenaeey, E.S.Y.; Kandeel, T.M.; Hegazy, W.A.H.; Mosbah, R.A.; Nassar, M.S.; Bakhrebah, M.A.; Abdulaal, W.H.; Alhakamy, N.A.; Bendary, M.M. Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 667. [Google Scholar] [CrossRef]
- Welsh, M.A.; Blackwell, H.E. Chemical Probes of Quorum Sensing: From Compound Development to Biological Discoveries. FEMS Microbiol. Rev. 2016, 40, 774–794. [Google Scholar] [CrossRef]
- Lu, C.; Maurer, C.K.; Kirsch, B.; Steinbach, A.; Hartmann, R.W. Overcoming the Unexpected Functional Inversion of a PqsR Antagonist in Pseudomonas aeruginosa: An in Vivo Potent Antivirulence Agent Targeting Pqs Quorum Sensing. Angew. Chem. Int. Ed. Engl. 2014, 53, 1109–1112. [Google Scholar] [CrossRef]
- Wei, L.N.; Shi, C.Z.; Luo, C.X.; Hu, C.Y.; Meng, Y.H. Phloretin Inhibits Biofilm Formation by Affecting Quorum Sensing under Different Temperature. LWT 2020, 131, 109668. [Google Scholar] [CrossRef]
- Ouyang, J.; Feng, W.; Lai, X.; Chen, Y.; Zhang, X.; Rong, L.; Sun, F.; Chen, Y. Quercetin Inhibits Pseudomonas aeruginosa Biofilm Formation via the Vfr-Mediated LasIR System. Microb. Pathog. 2020, 149, 104291. [Google Scholar] [CrossRef]
- Ho, D.K.; Murgia, X.; de Rossi, C.; Christmann, R.; Hüfner de Mello Martins, A.G.; Koch, M.; Andreas, A.; Herrmann, J.; Müller, R.; Empting, M.; et al. Squalenyl Hydrogen Sulfate Nanoparticles for Simultaneous Delivery of Tobramycin and an Alkylquinolone Quorum Sensing Inhibitor Enable the Eradication of P. aeruginosa Biofilm Infections. Angew. Chem. Int. Ed. Engl. 2020, 59, 10292–10296. [Google Scholar] [CrossRef] [PubMed]
- Tomlin, H.; Piccinini, A.M. A Complex Interplay between the Extracellular Matrix and the Innate Immune Response to Microbial Pathogens. Immunology 2018, 155, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Mühlenbruch, M.; Grossart, H.P.; Eigemann, F.; Voss, M. Mini-Review: Phytoplankton-Derived Polysaccharides in the Marine Environment and Their Interactions with Heterotrophic Bacteria. Environ. Microbiol. 2018, 20, 2671–2685. [Google Scholar] [CrossRef] [PubMed]
- McKee, L.S.; la Rosa, S.L.; Westereng, B.; Eijsink, V.G.; Pope, P.B.; Larsbrink, J. Polysaccharide Degradation by the Bacteroidetes: Mechanisms and Nomenclature. Environ. Microbiol. Rep. 2021, 13, 559–581. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Wear, S.S.; Sande, C. Assembly of Bacterial Capsular Polysaccharides and Exopolysaccharides. Annu. Rev. Microbiol. 2020, 74, 521–543. [Google Scholar] [CrossRef]
- Ren, Z.; Cui, T.; Zeng, J.; Chen, L.; Zhang, W.; Xu, X.; Cheng, L.; Li, M.; Li, J.; Zhou, X.; et al. Molecule Targeting Glucosyltransferase Inhibits Streptococcus mutans Biofilm Formation and Virulence. Antimicrob. Agents. Chemother. 2015, 60, 126–135. [Google Scholar] [CrossRef]
- Falsetta, M.L.; Klein, M.I.; Lemos, J.A.; Silva, B.B.; Agidi, S.; Scott-Anne, K.K.; Koo, H. Novel Antibiofilm Chemotherapy Targets Exopolysaccharide Synthesis and Stress Tolerance in Streptococcus mutans to Modulate Virulence Expression In Vivo. Antimicrob. Agents. Chemother. 2012, 56, 6201–6211. [Google Scholar] [CrossRef]
- Laverty, G.; Gorman, S.P.; Gilmore, B.F. Biomolecular Mechanisms of Pseudomonas aeruginosa and Escherichia coli Biofilm Formation. Pathogens 2014, 3, 596–632. [Google Scholar] [CrossRef]
- De Araujo Lopes, A.C.U.; Lobo, C.I.V.; Ribeiro, S.M.; da Silva Colin, J.; Constantino, V.C.N.; Canonici, M.M.; Barbugli, P.A.; Klein, M.I. Distinct Agents Induce Streptococcus mutans Cells with Altered Biofilm Formation Capacity. Microbiol. Spectr. 2022, 10, e00650-22. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, Q.; Wang, Y.; Wu, H.; Zou, J. Molecular Mechanisms of Inhibiting Glucosyltransferases for Biofilm Formation in Streptococcus mutans. Int. J. Oral Sci. 2021, 13, 30. [Google Scholar] [CrossRef]
- Jiang, H.; Hu, Y.; Yang, M.; Liu, H.; Jiang, G. Enhanced Immune Response to a Dual-Promoter Anti-Caries DNA Vaccine Orally Delivered by Attenuated Salmonella typhimurium. Immunobiology 2017, 222, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Thorn, C.R.; Howell, P.L.; Wozniak, D.J.; Prestidge, C.A.; Thomas, N. Enhancing the Therapeutic Use of Biofilm-Dispersing Enzymes with Smart Drug Delivery Systems. Advanced Adv. Drug Deliv. Rev. 2021, 179, 113916. [Google Scholar] [CrossRef]
- Donelli, G.; Francolini, I.; Romoli, D.; Guaglianone, E.; Piozzi, A.; Ragunath, C.; Kaplan, J.B. Synergistic Activity of Dispersin B and Cefamandole Nafate in Inhibition of Staphylococcal Biofilm Growth on Polyurethanes. Antimicrob. Agents. Chemother. 2007, 51, 2733–2740. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, Z.; Zeng, K.; Xia, Y.; Xu, W.; Wang, R.; Guo, J.; Xie, H. Functional Immobilization of a Biofilm-Releasing Glycoside Hydrolase Dispersin B on Magnetic Nanoparticles. Appl. Biochem. Biotechnol. 2022, 194, 737–747. [Google Scholar] [CrossRef] [PubMed]
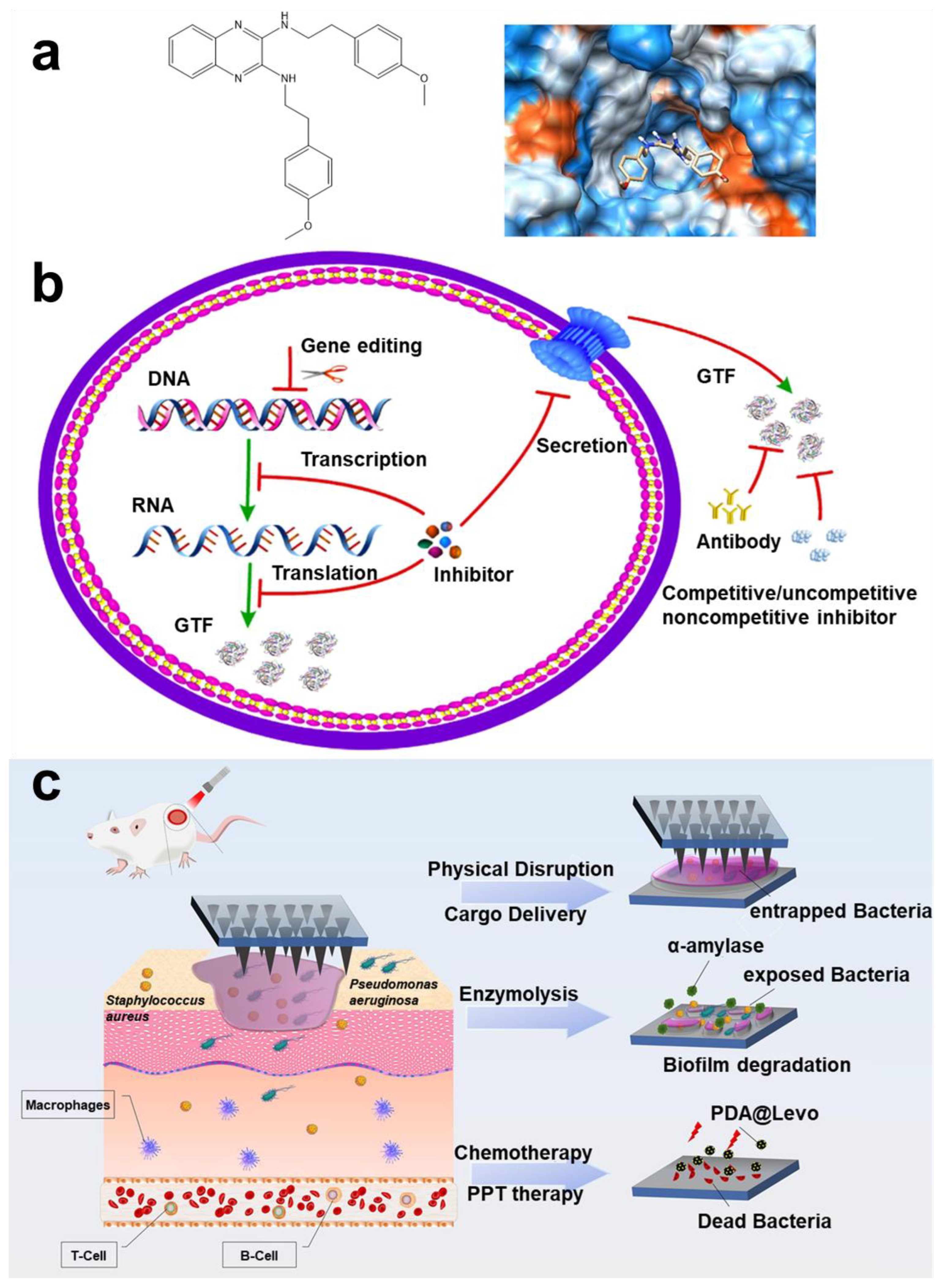
- Yu, X.; Zhao, J.; Fan, D. A Dissolving Microneedle Patch for Antibiotic/Enzymolysis/Photothermal Triple Therapy against Bacteria and Their Biofilms. Chem. Eng. J. 2022, 437, 135475. [Google Scholar] [CrossRef]
- Maunders, E.; Welch, M. Matrix Exopolysaccharides; the Sticky Side of Biofilm Formation. FEMS Microbiol. Lett. 2017, 364, fnx120. [Google Scholar] [CrossRef]
- Patel, K.K.; Tripathi, M.; Pandey, N.; Agrawal, A.K.; Gade, S.; Anjum, M.M.; Tilak, R.; Singh, S. Alginate Lyase Immobilized Chitosan Nanoparticles of Ciprofloxacin for the Improved Antimicrobial Activity against the Biofilm Associated Mucoid P. aeruginosa Infection in Cystic Fibrosis. Int. J. Pharm. 2019, 563, 30–42. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Li, X.; Lee, B.S.; Jung, S.; Lee, M.S. Enhancing the Thermo-Stability and Anti-Biofilm Activity of Alginate Lyase by Immobilization on Low Molecular Weight Chitosan Nanoparticles. Int. J. Mol. Sci. 2019, 20, 4565. [Google Scholar] [CrossRef]
- Daboor, S.M.; Rohde, J.R.; Cheng, Z. Disruption of the Extracellular Polymeric Network of Pseudomonas aeruginosa Biofilms by Alginate Lyase Enhances Pathogen Eradication by Antibiotics. J. Cyst. Fibros. 2021, 20, 264–270. [Google Scholar] [CrossRef]
- Wan, B.; Zhu, Y.; Tao, J.; Zhu, F.; Chen, J.; Li, L.; Zhao, J.; Wang, L.; Sun, S.; Yang, Y.; et al. Alginate Lyase Guided Silver Nanocomposites for Eradicating Pseudomonas aeruginosa from Lungs. ACS Appl. Mater. Interfaces 2020, 12, 9050–9061. [Google Scholar] [CrossRef]
- Szymańska, M.; Karakulska, J.; Sobolewski, P.; Kowalska, U.; Grygorcewicz, B.; Böttcher, D.; Bornscheuer, U.T.; Drozd, R. Glycoside Hydrolase (PelAh) Immobilization Prevents Pseudomonas aeruginosa Biofilm Formation on Cellulose-Based Wound Dressing. Carbohydr. Polym. 2020, 246, 116625. [Google Scholar] [CrossRef] [PubMed]
- Catlin, B.W. Extracellular deoxyribonucleic acid of bacteria and a deoxyribonuclease inhibitor. Science 1956, 124, 441–442. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S. Release Mechanisms and Molecular Interactions of Pseudomonas aeruginosa Extracellular DNA. Appl. Microbiol. Biotechnol. 2020, 104, 6549–6564. [Google Scholar] [CrossRef] [PubMed]
- Lewenza, S.; Johnson, L.; Charron-Mazenod, L.; Hong, M.; Mulcahy-O’Grady, H. Extracellular DNA Controls Expression of Pseudomonas aeruginosa Genes Involved in Nutrient Utilization, Metal Homeostasis, Acid PH Tolerance and Virulence. J. Med. Microbiol. 2020, 69, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, M.; Zheng, X.; Ma, H.; Nerenberg, R.; Chai, H. Extracellular DNA Plays a Key Role in the Structural Stability of Sulfide-Based Denitrifying Biofilms. Sci. Total Environ. 2022, 838, 155822. [Google Scholar] [CrossRef]
- Peng, N.; Cai, P.; Mortimer, M.; Wu, Y.; Gao, C.; Huang, Q. The Exopolysaccharide-EDNA Interaction Modulates 3D Architecture of Bacillus subtilis Biofilm. BMC Microbiol. 2020, 20, 115. [Google Scholar] [CrossRef]
- Tan, Y.; Ma, S.; Leonhard, M.; Moser, D.; Haselmann, G.M.; Wang, J.; Eder, D.; Schneider-Stickler, B. Enhancing Antibiofilm Activity with Functional Chitosan Nanoparticles Targeting Biofilm Cells and Biofilm Matrix. Carbohydr. Polym. 2018, 200, 35–42. [Google Scholar] [CrossRef]
- Panlilio, H.; Rice, C.v. The Role of Extracellular DNA in the Formation, Architecture, Stability, and Treatment of Bacterial Biofilms. Biotechnol. Bioeng. 2021, 118, 2129–2141. [Google Scholar] [CrossRef]
- Okshevsky, M.; Regina, V.R.; Meyer, R.L. Extracellular DNA as a Target for Biofilm Control. Curr. Opin. Biotechnol. 2015, 33, 73–80. [Google Scholar] [CrossRef]
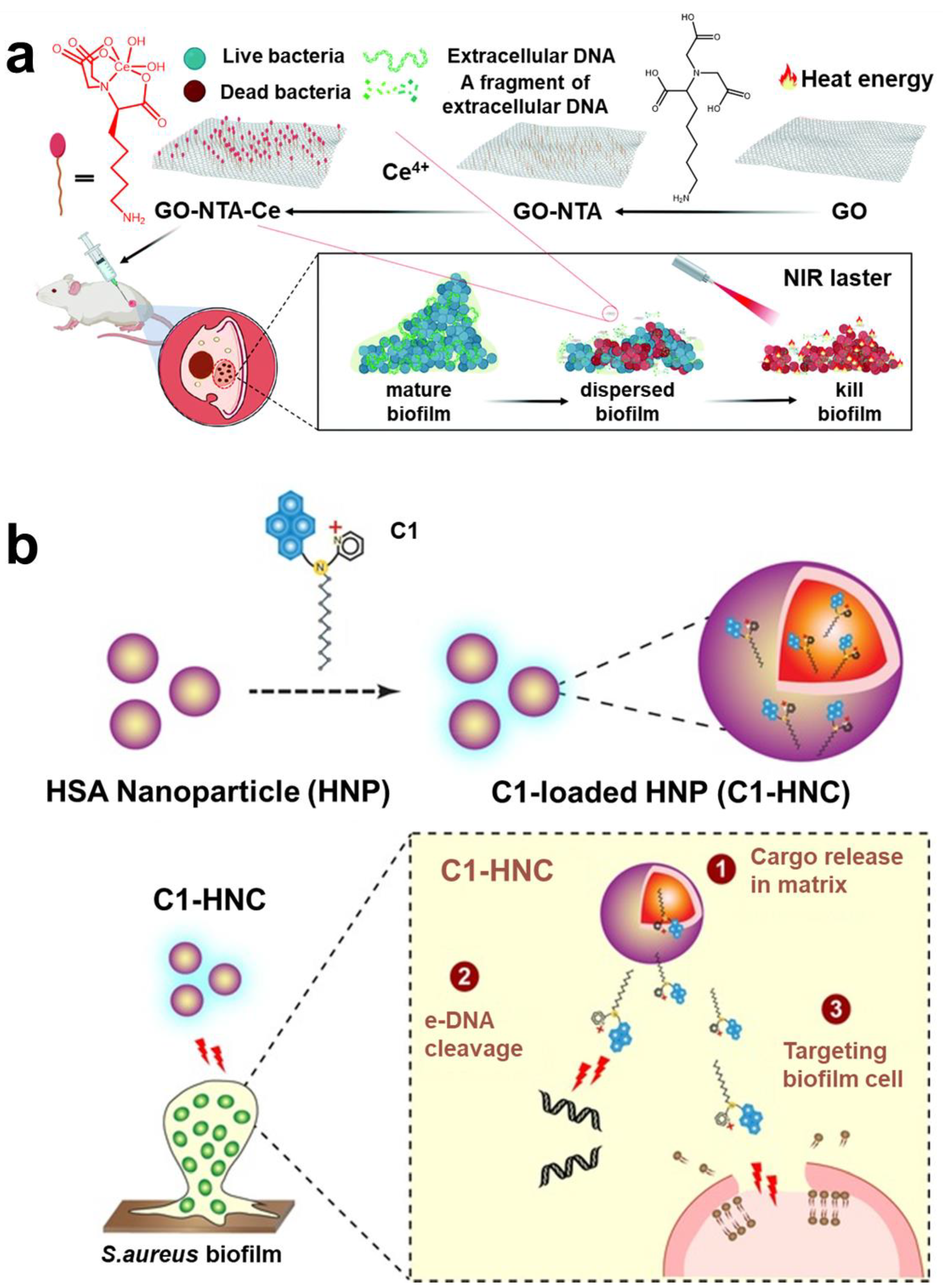
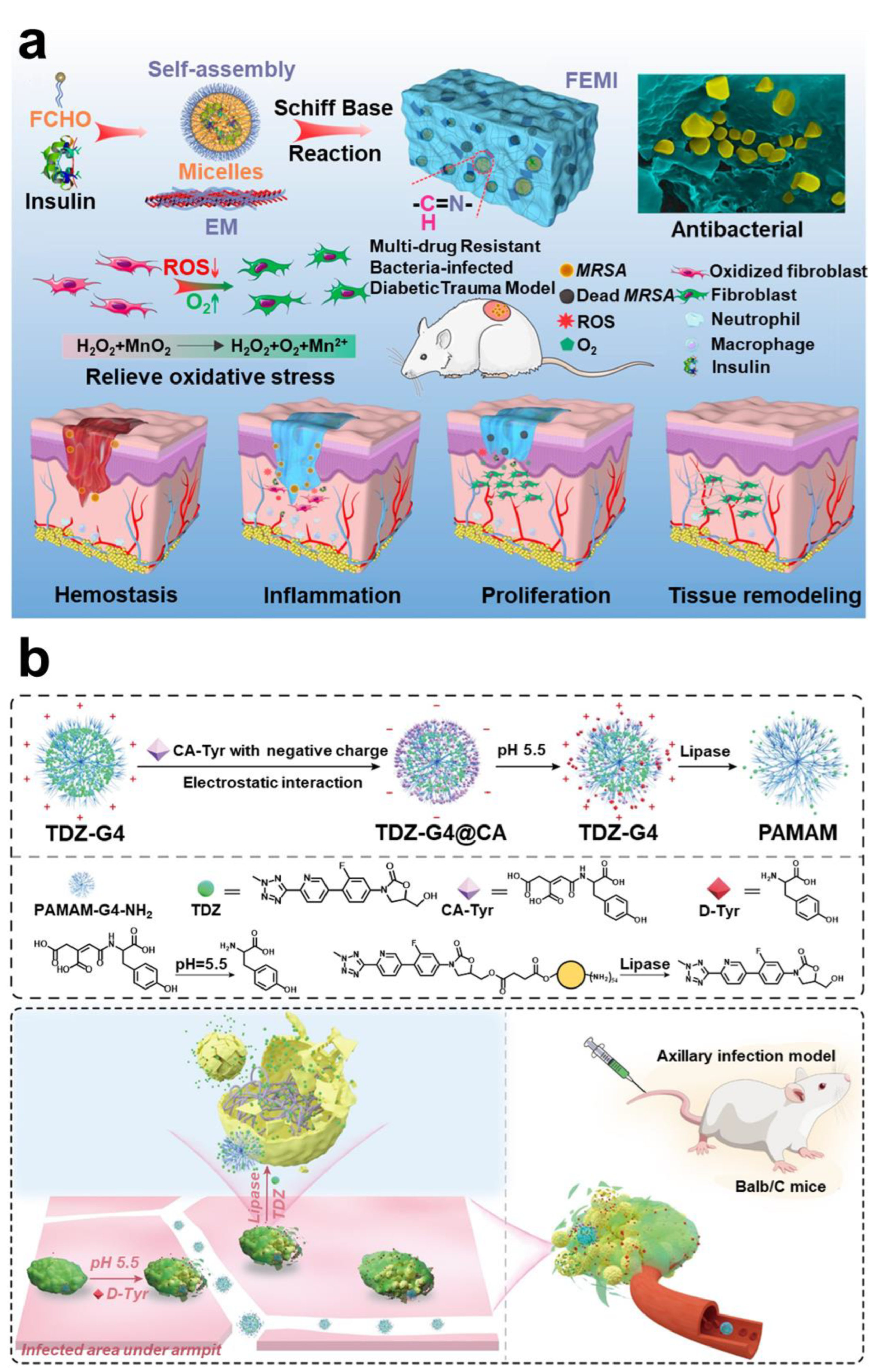
- Hu, H.; Kang, X.; Shan, Z.; Yang, X.; Bing, W.; Wu, L.; Ge, H.; Ji, H. A DNase-Mimetic Artificial Enzyme for the Eradication of Drug-Resistant Bacterial Biofilm Infections. Nanoscale 2022, 14, 2676–2685. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, F.; Ren, J.; Qu, X. A Series of MOF/Ce-Based Nanozymes with Dual Enzyme-like Activity Disrupting Biofilms and Hindering Recolonization of Bacteria. Biomaterials 2019, 208, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Gu, S.; Shi, Y.; Cui, X.; Wen, S.; Ge, J. The Effect of Emodin on Staphylococcus aureus Strains in Planktonic Form and Biofilm Formation In Vitro. Arch. Microbiol. 2017, 199, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Thiyagarajan, D.; Das, G.; Ramesh, A. Extracellular-DNA-Targeting Nanomaterial for Effective Elimination of Biofilm. ChemNanoMat 2016, 2, 879–887. [Google Scholar] [CrossRef]
- Kwon, H.Y.; Kim, J.Y.; Liu, X.; Lee, J.Y.; Yam, J.K.H.; Dahl Hultqvist, L.; Xu, W.; Rybtke, M.; Tolker-Nielsen, T.; Heo, W.; et al. Visualizing Biofilm by Targeting EDNA with Long Wavelength Probe CDr15. Biomater. Sci. 2019, 7, 3594–3598. [Google Scholar] [CrossRef]
- Erskine, E.; MacPhee, C.E.; Stanley-Wall, N.R. Functional Amyloid and Other Protein Fibers in the Biofilm Matrix. J. Mol. Biol. 2018, 430, 3642–3656. [Google Scholar] [CrossRef]
- Kavanaugh, J.S.; Flack, C.E.; Lister, J.; Ricker, E.B.; Ibberson, C.B.; Jenul, C.; Moormeier, D.E.; Delmain, E.A.; Bayles, K.W.; Horswill, A.R. Identification of Extracellular DNA-Binding Proteins in the Biofilm Matrix. mBio 2019, 10, e01137-19. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.K.; Subba Rao, T. Staphylococcus aureus Biofilm Removal by Targeting Biofilm-Associated Extracellular Proteins. Indian J. Med. Res. 2017, 146 (Suppl. 1), S1–S8. [Google Scholar] [CrossRef]
- Latasa, C.; Solano, C.; Penadés, J.R.; Lasa, I. Biofilm-Associated Proteins. Comptes Rendus Biol. 2006, 329, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Fong, J.N.C.; Yildiz, F.H. Biofilm Matrix Proteins. Microbiol. Spectr. 2015, 3, 3.2.28. [Google Scholar] [CrossRef]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A Major Protein Component of the Bacillus subtilis Biofilm Matrix. Mol. Microbiol. 2006, 59, 1229–1238. [Google Scholar] [CrossRef]
- Rocco, C.J.; Davey, M.E.; Bakaletz, L.O.; Goodman, S.D. Natural Antigenic Differences in the Functionally Equivalent Extracellular DNABII Proteins of Bacterial Biofilms Provide a Means for Targeted Biofilm Therapeutics. Mol. Oral Microbiol. 2017, 32, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Tang, N.; Aspiras, M.B.; Lau, P.C.Y.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. A Quorum-Sensing Signaling System Essential for Genetic Competence in Streptococcus mutans Is Involved in Biofilm Formation. J. Bacteriol. 2002, 184, 2699–2708. [Google Scholar] [CrossRef]
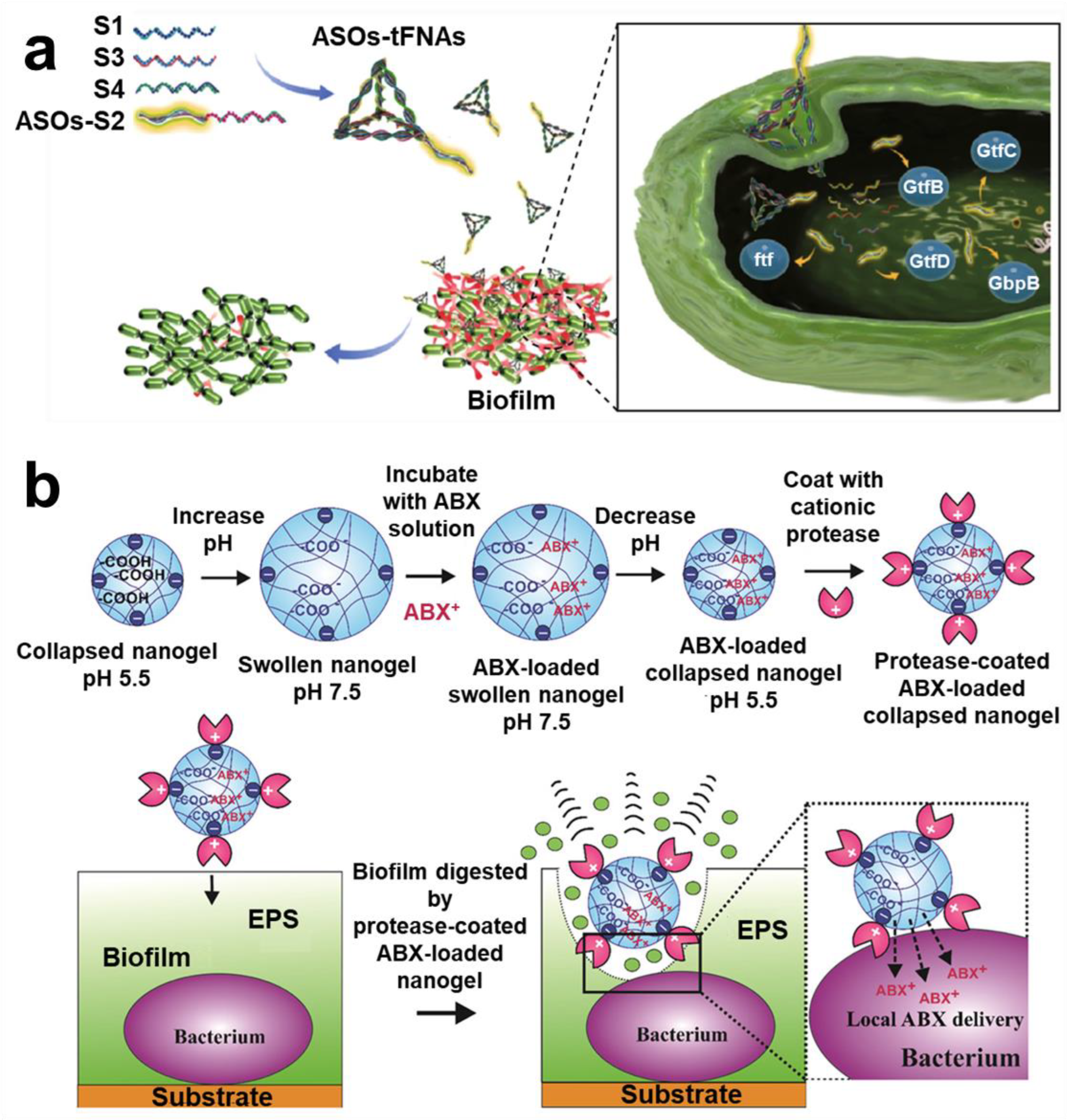
- Zhang, Y.; Xie, X.; Ma, W.; Zhan, Y.; Mao, C.; Shao, X.; Lin, Y. Multi-Targeted Antisense Oligonucleotide Delivery by a Framework Nucleic Acid for Inhibiting Biofilm Formation and Virulence. Nanomicro Lett. 2020, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Li, J.; Bai, Y.; Zhang, Y.; Sun, H.; Zhang, X. A Bacterial Infection-Microenvironment Activated Nanoplatform Based on Spiropyran-Conjugated Glycoclusters for Imaging and Eliminating of the Biofilm. Chem. Eng. J. 2020, 399, 125787. [Google Scholar] [CrossRef]
- Weldrick, P.J.; Hardman, M.J.; Paunov, V.N. Enhanced Clearing of Wound-Related Pathogenic Bacterial Biofilms Using Protease-Functionalized Antibiotic Nanocarriers. ACS Appl. Mater. Interfaces 2019, 11, 43902–43919. [Google Scholar] [CrossRef] [PubMed]
- Devlin, H.; Fulaz, S.; Hiebner, D.W.; O’gara, J.P.; Casey, E. Enzyme-Functionalized Mesoporous Silica Nanoparticles to Target Staphylococcus aureus and Disperse Biofilms. Int. J. Nanomed. 2021, 16, 1929–1942. [Google Scholar] [CrossRef]
- Chen, B.; Huang, J.; Li, H.; Zeng, Q.H.; Wang, J.J.; Liu, H.; Pan, Y.; Zhao, Y. Eradication of Planktonic Vibrio parahaemolyticus and Its Sessile Biofilm by Curcumin-Mediated Photodynamic Inactivation. Food Control 2020, 113, 107181. [Google Scholar] [CrossRef]
- Jo, J.; Price-Whelan, A.; Dietrich, L.E.P. Gradients and Consequences of Heterogeneity in Biofilms. Nat. Rev. Microbiol. 2022, 20, 593–607. [Google Scholar] [CrossRef]
- Jung, S.H.; Ryu, C.M.; Kim, J.S. Bacterial Persistence: Fundamentals and Clinical Importance. J. Microbiol. 2019, 57, 829–835. [Google Scholar] [CrossRef]
- Soares, A.; Alexandre, K.; Etienne, M. Tolerance and Persistence of Pseudomonas aeruginosa in Biofilms Exposed to Antibiotics: Molecular Mechanisms, Antibiotic Strategies and Therapeutic Perspectives. Front. Microbiol. 2020, 11, 2057. [Google Scholar] [CrossRef]
- Alexander, C.; Guru, A.; Pradhan, P.; Mallick, S.; Mahanandia, N.C.; Subudhi, B.B.; Beuria, T.K. MazEF-rifampicin interaction suggests a mechanism for rifampicin induced inhibition of persisters. BMC Mol. Cell Biol. 2020, 21, 73. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J.F.; Rôças, I.N. Clinical Implications and Microbiology of Bacterial Persistence after Treatment Procedures. J. Endod. 2008, 34, 1291–1301.e3. [Google Scholar] [CrossRef] [PubMed]
- Cardona, P.J.; Ruiz-Manzano, J. On the Nature of Mycobacterium tuberculosis-Latent Bacilli. Eur. Respir. J. 2004, 24, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Prabowo, S.A.; Gröschel, M.I.; Schmidt, E.D.L.; Skrahina, A.; Mihaescu, T.; Hastürk, S.; Mitrofanov, R.; Pimkina, E.; Visontai, I.; de Jong, B.; et al. Targeting Multidrug-Resistant Tuberculosis (MDR-TB) by Therapeutic Vaccines. Med. Microbiol. Immunol. 2013, 202, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Koul, A.; Dendouga, N.; Vergauwen, K.; Molenberghs, B.; Vranckx, L.; Willebrords, R.; Ristic, Z.; Lill, H.; Dorange, I.; Guillemont, J.; et al. Diarylquinolines Target Subunit c of Mycobacterial ATP Synthase. Nat. Chem. Biol. 2007, 3, 323–324. [Google Scholar] [CrossRef]
- Balemans, W.; Vranckx, L.; Lounis, N.; Pop, O.; Guillemont, J.; Vergauwen, K.; Mol, S.; Gilissen, R.; Motte, M.; Lançois, D.; et al. Novel Antibiotics Targeting Respiratory ATP Synthesis in Gram-Positive Pathogenic Bacteria. Antimicrob. Agents Chemother. 2012, 56, 4131–4139. [Google Scholar] [CrossRef]
- Garrison, A.T.; Abouelhassan, Y.; Norwood, V.M.; Kallifidas, D.; Bai, F.; Nguyen, M.T.; Rolfe, M.; Burch, G.M.; Jin, S.; Luesch, H.; et al. Structure-Activity Relationships of a Diverse Class of Halogenated Phenazines That Targets Persistent, Antibiotic-Tolerant Bacterial Biofilms and Mycobacterium tuberculosis. J. Med. Chem. 2016, 59, 3808–3825. [Google Scholar] [CrossRef]
- Dutta, N.K.; Klinkenberg, L.G.; Vazquez, M.J.; Segura-Carro, D.; Colmenarejo, G.; Ramon, F.; Rodriguez-Miquel, B.; Mata-Cantero, L.; de Francisco, E.P.; Chuang, Y.M.; et al. Inhibiting the Stringent Response Blocks Mycobacterium tuberculosis Entry into Quiescence and Reduces Persistence. Sci. Adv. 2019, 5, eaav2104. [Google Scholar] [CrossRef]
- Tkachenko, A.G.; Kashevarova, N.M.; Sidorov, R.Y.; Nesterova, L.Y.; Akhova, A.v.; Tsyganov, I.v.; Vaganov, V.Y.; Shipilovskikh, S.A.; Rubtsov, A.E.; Malkov, A.v. A Synthetic Diterpene Analogue Inhibits Mycobacterial Persistence and Biofilm Formation by Targeting (p)PpGpp Synthetases. Cell Chem. Biol. 2021, 28, 1420–1432.e9. [Google Scholar] [CrossRef]
- Kaur, P.; Potluri, V.; Ahuja, V.K.; Naveenkumar, C.N.; Krishnamurthy, R.V.; Gangadharaiah, S.T.; Shivarudraiah, P.; Eswaran, S.; Nirmal, C.R.; Mahizhaveni, B.; et al. A Multi-Targeting Pre-Clinical Candidate against Drug-Resistant Tuberculosis. Tuberculosis 2021, 129, 102104. [Google Scholar] [CrossRef]
- Roy, S.; Bahar, A.A.; Gu, H.; Nangia, S.; Sauer, K.; Ren, D. Persister Control by Leveraging Dormancy Associated Reduction of Antibiotic Efflux. PLoS Pathog. 2021, 17, e1010144. [Google Scholar] [CrossRef] [PubMed]
- Kitzenberg, D.A.; Lee, J.S.; Mills, K.B.; Kim, J.-S.; Liu, L.; Vázquez-Torres, A.; Colgan, S.P.; Kao, D.J. Adenosine Awakens Metabolism to Enhance Growth-Independent Killing of Tolerant and Persister Bacteria across Multiple Classes of Antibiotics. mBio 2022, 13, e00480-22. [Google Scholar] [CrossRef] [PubMed]
- Nabawy, A.; Makabenta, J.M.; Schmidt-Malan, S.; Park, J.; Li, C.H.; Huang, R.; Fedeli, S.; Chattopadhyay, A.N.; Patel, R.; Rotello, V.M. Dual Antimicrobial-Loaded Biodegradable Nanoemulsions for Synergistic Treatment of Wound Biofilms. J. Control. Release 2022, 347, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; An, Q.; Zhang, Y.; Sun, K.; Bai, L.; Luo, Y. Recent Advances in Clp Protease Modulation to Address Virulence, Resistance and Persistence of MRSA Infection. Drug Discov. Today 2021, 26, 2190–2197. [Google Scholar] [CrossRef]
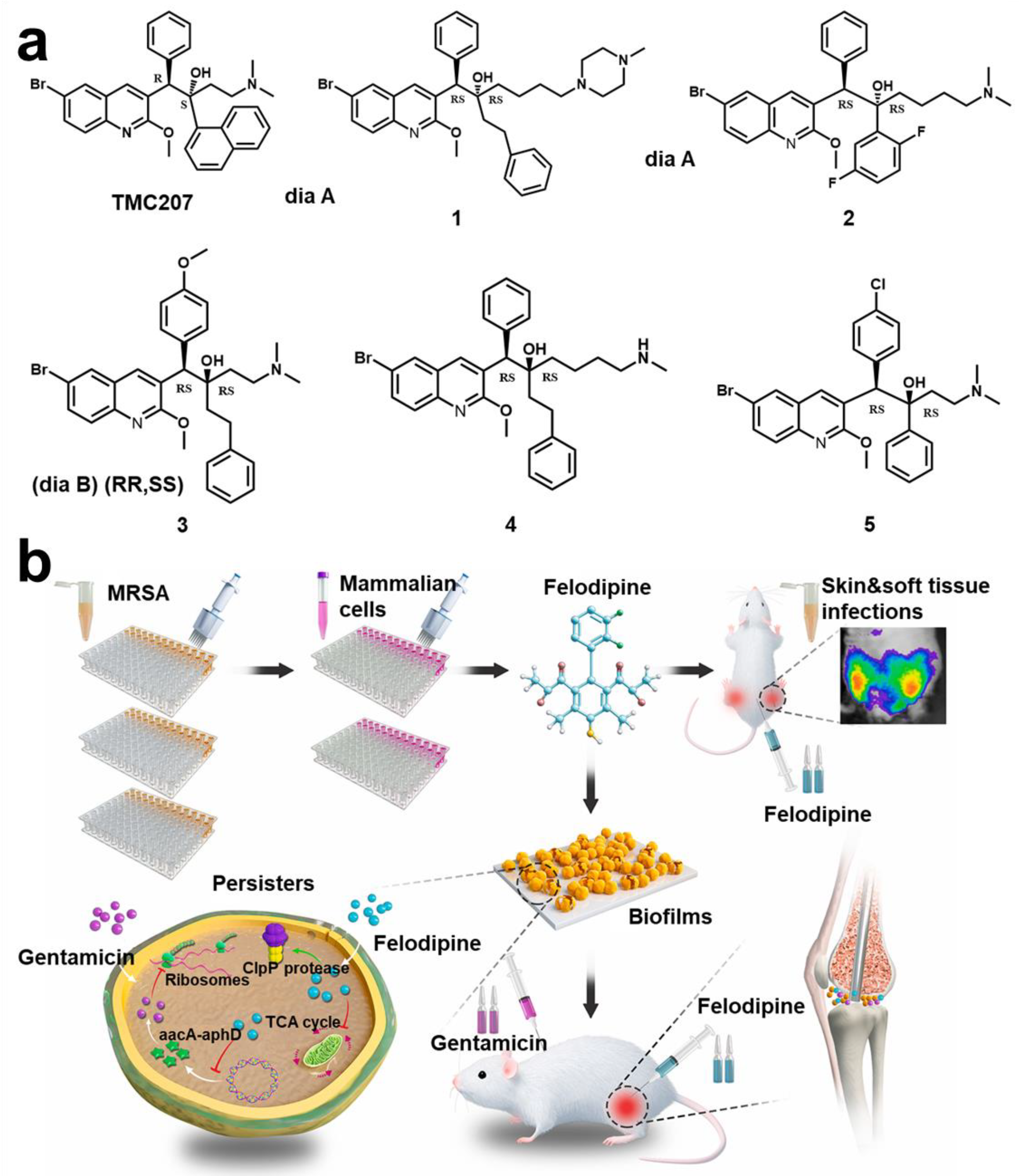
- Zhang, S.; Qu, X.; Jiao, J.; Tang, H.; Wang, M.; Wang, Y.; Yang, H.; Yuan, W.; Yue, B. Felodipine Enhances Aminoglycosides Efficacy against Implant Infections Caused by Methicillin-Resistant Staphylococcus aureus, Persisters and Biofilms. Bioact. Mater. 2022, 14, 272–289. [Google Scholar] [CrossRef]
- Walters, M.C.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Stewart, P.S. Contributions of Antibiotic Penetration, Oxygen Limitation, and Low Metabolic Activity to Tolerance of Pseudomonas aeruginosa Biofilms to Ciprofloxacin and Tobramycin. Antimicrob. Agents Chemother. 2003, 47, 317–323. [Google Scholar] [CrossRef]
- Marteyn, B.; West, N.P.; Browning, D.F.; Cole, J.A.; Shaw, J.G.; Palm, F.; Mounier, J.; Prévost, M.C.; Sansonetti, P.; Tang, C.M. Modulation of Shigella Virulence in Response to Available Oxygen in Vivo. Nature 2010, 465, 355–358. [Google Scholar] [CrossRef]
- Bai, Y.; Hu, Y.; Gao, Y.; Wei, X.; Li, J.; Zhang, Y.; Wu, Z.; Zhang, X. Oxygen Self-Supplying Nanotherapeutic for Mitigation of Tissue Hypoxia and Enhanced Photodynamic Therapy of Bacterial Keratitis. ACS Appl. Mater. Interfaces 2021, 13, 33790–33801. [Google Scholar] [CrossRef]
- Zou, L.; Hu, D.; Wang, F.; Jin, Q.; Ji, J. The Relief of Hypoxic Microenvironment Using an O2 Self-Sufficient Fluorinated Nanoplatform for Enhanced Photodynamic Eradication of Bacterial Biofilms. Nano Res. 2022, 15, 1636–1644. [Google Scholar] [CrossRef]
- Hu, D.; Zou, L.; Yu, W.; Jia, F.; Han, H.; Yao, K.; Jin, Q.; Ji, J. Relief of Biofilm Hypoxia Using an Oxygen Nanocarrier: A New Paradigm for Enhanced Antibiotic Therapy. Adv. Sci. 2020, 7, 2000398. [Google Scholar] [CrossRef]
- Deng, Q.; Sun, P.; Zhang, L.; Liu, Z.; Wang, H.; Ren, J.; Qu, X. Porphyrin MOF Dots–Based, Function-Adaptive Nanoplatform for Enhanced Penetration and Photodynamic Eradication of Bacterial Biofilms. Adv. Funct. Mater. 2019, 29, 1903018. [Google Scholar] [CrossRef]
- Xiu, W.; Gan, S.; Wen, Q.; Qiu, Q.; Dai, S.; Dong, H.; Li, Q.; Yuwen, L.; Weng, L.; Teng, Z.; et al. Biofilm Microenvironment-Responsive Nanotheranostics for Dual-Mode Imaging and Hypoxia-Relief-Enhanced Photodynamic Therapy of Bacterial Infections. Research 2020, 2020, 9426453. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Hua, Y.; Zhang, Y.; Lv, M.; Wang, H.; Pi, Y.; Xie, J.; Wang, C.; Yong, Y. A Biofilm Microenvironment-Activated Single-Atom Iron Nanozyme with NIR-Controllable Nanocatalytic Activities for Synergetic Bacteria-Infected Wound Therapy. Adv. Healthc. Mater. 2021, 10, 2101374. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zheng, H.; Zhou, L.; Cheng, F.; Liu, Z.; Zhang, H.; Wang, L.; Zhang, Q. Nanoenzyme-Reinforced Injectable Hydrogel for Healing Diabetic Wounds Infected with Multidrug Resistant Bacteria. Nano Lett. 2020, 20, 5149–5158. [Google Scholar] [CrossRef]
- Zhang, S.; Chai, Q.; Man, Z.; Tang, C.; Li, Z.; Zhang, J.; Xu, H.; Xu, X.; Chen, C.; Liu, Y.; et al. Bioinspired Nano-Painting on Orthopedic Implants Orchestrates Periprosthetic Anti-Infection and Osseointegration in a Rat Model of Arthroplasty. Chem. Eng. J. 2022, 435, 134848. [Google Scholar] [CrossRef]
- Scalise, A.; Bianchi, A.; Tartaglione, C.; Bolletta, E.; Pierangeli, M.; Torresetti, M.; Marazzi, M.; di Benedetto, G. Microenvironment and microbiology of skin wounds: The role of bacterial biofilms and related factors. Semin. Vasc. Surg. 2015, 28, 151–159. [Google Scholar] [CrossRef]
- Wilton, M.; Charron-Mazenod, L.; Moore, R.; Lewenza, S. Extracellular DNA Acidifies Biofilms and Induces Aminoglycoside Resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2015, 60, 544–553. [Google Scholar] [CrossRef]
- Chen, H.; Yang, J.; Sun, L.; Zhang, H.; Guo, Y.; Qu, J.; Jiang, W.; Chen, W.; Ji, J.; Yang, Y.W.; et al. Synergistic Chemotherapy and Photodynamic Therapy of Endophthalmitis Mediated by Zeolitic Imidazolate Framework-Based Drug Delivery Systems. Small 2019, 15, 1903880. [Google Scholar] [CrossRef]
- Wang, D.Y.; Yang, G.; van der Mei, H.C.; Ren, Y.; Busscher, H.J.; Shi, L. Liposomes with Water as a PH-Responsive Functionality for Targeting of Acidic Tumor and Infection Sites. Angew. Chem. Int. Ed. Engl. 2021, 60, 17714–17719. [Google Scholar] [CrossRef]
- Jiang, X.; Li, W.; Chen, X.; Wang, C.; Guo, R.; Hong, W. On-Demand Multifunctional Electrostatic Complexation for Synergistic Eradication of MRSA Biofilms. ACS Appl. Mater. Interfaces 2022, 14, 10200–10211. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, J.; Chai, M.; Li, X.; Deng, Y.; Jin, Q.; Ji, J. Size and Charge Adaptive Clustered Nanoparticles Targeting the Biofilm Microenvironment for Chronic Lung Infection Management. ACS Nano 2020, 14, 5686–5699. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, F.; Hu, X.; Lu, J.; Sun, X.; Gao, J.; Ling, D. Responsive Assembly of Silver Nanoclusters with a Biofilm Locally Amplified Bactericidal Effect to Enhance Treatments against Multi-Drug-Resistant Bacterial Infections. ACS Cent. Sci. 2019, 5, 1366–1376. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cheng, J.; Cai, X.; Han, J.; Chen, X.; You, L.; Xiong, C.; Wang, S. PH-Switchable Antimicrobial Supramolecular Hydrogels for Synergistically Eliminating Biofilm and Promoting Wound Healing. ACS Appl. Mater. Interfaces 2022, 14, 18120–18132. [Google Scholar] [CrossRef]
- Fulaz, S.; Devlin, H.; Vitale, S.; Quinn, L.; O’gara, J.P.; Casey, E. Tailoring Nanoparticle-Biofilm Interactions to Increase the Efficacy of Antimicrobial Agents against Staphylococcus aureus. Int. J. Nanomed. 2020, 15, 4779–4791. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Qiao, Z.; Yan, D.; Yang, M.; Yang, L.; Wan, X.; Chen, H.; Luo, J.; Xiao, H. Ciprofloxacin Conjugated Gold Nanorods with PH Induced Surface Charge Transformable Activities to Combat Drug Resistant Bacteria and Their Biofilms. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 128, 112292. [Google Scholar] [CrossRef] [PubMed]
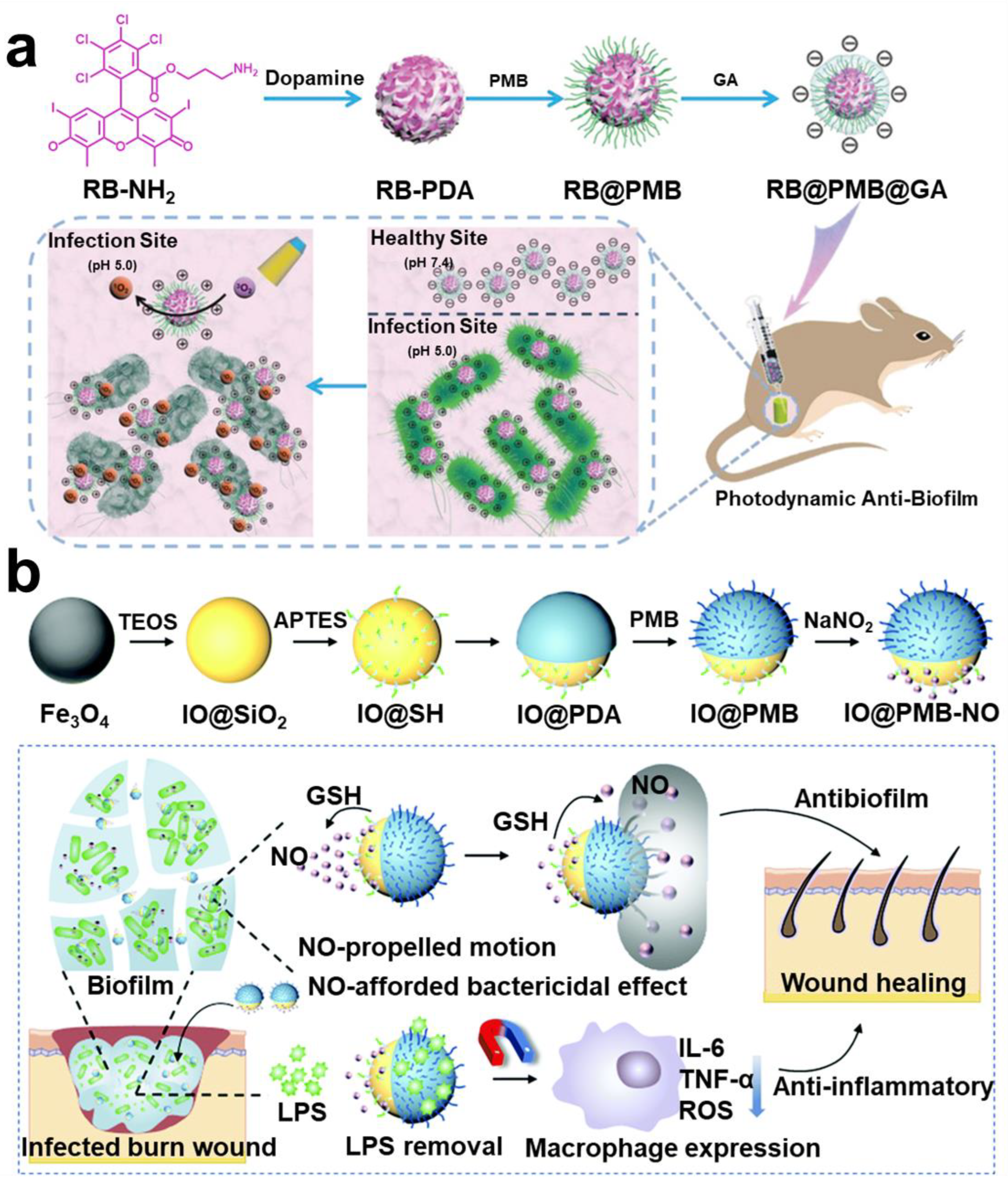
- Wu, S.; Xu, C.; Zhu, Y.; Zheng, L.; Zhang, L.; Hu, Y.; Yu, B.; Wang, Y.; Xu, F.J. Biofilm-Sensitive Photodynamic Nanoparticles for Enhanced Penetration and Antibacterial Efficiency. Adv. Funct. Mater. 2021, 31, 2103591. [Google Scholar] [CrossRef]
- Tian, S.; Su, L.; Liu, Y.; Cao, J.; Yang, G.; Ren, Y.; Huang, F.; Liu, J.; An, Y.; van der Mei, H.C.; et al. Self-Targeting, Zwitterionic Micellar Dispersants Enhance Antibiotic Killing of Infectious Biofilms—An Intravital Imaging Study in Mice. Sci. Adv. 2020, 6, eabb1112. [Google Scholar] [CrossRef]
- Shatalin, K.; Nuthanakanti, A.; Kaushik, A.; Shishov, D.; Peselis, A.; Shamovsky, I.; Pani, B.; Lechpammer, M.; Vasilyev, N.; Shatalina, E.; et al. Inhibitors of Bacterial H2S Biogenesis Targeting Antibiotic Resistance and Tolerance. Science 2021, 372, 1169–1175. [Google Scholar] [CrossRef]
- Zeng, J.; Wang, Y.; Sun, Z.; Chang, H.; Cao, M.; Zhao, J.; Lin, K.; Xie, Y. A Novel Biocompatible PDA/IR820/DAP Coating for Antibiotic/Photodynamic/Photothermal Triple Therapy to Inhibit and Eliminate Staphylococcus aureus Biofilm. Chem. Eng. J. 2020, 394, 125017. [Google Scholar] [CrossRef]
- Peng, J.; Xie, S.; Huang, K.; Ran, P.; Wei, J.; Zhang, Z.; Li, X. Nitric Oxide-Propelled Nanomotors for Bacterial Biofilm Elimination and Endotoxin Removal to Treat Infected Burn Wounds. J. Mater. Chem. B 2022, 10, 4189–4202. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Y.; Shah, S.; Kim, D.; Simon-Soro, A.; Ito, T.; Hajfathalian, M.; Li, Y.; Hsu, J.C.; Nieves, L.M.; et al. Precision Targeting of Bacterial Pathogen via Bi-Functional Nanozyme Activated by Biofilm Microenvironment. Biomaterials 2021, 268, 120581. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, X.; Ma, S.; Guo, Q.; Zhang, W.; Cheng, L.; Ding, L.; Xu, Z.; Jiang, J.; Gao, L. Oral Biofilm Elimination by Combining Iron-Based Nanozymes and Hydrogen Peroxide-Producing Bacteria. Biomater. Sci. 2020, 8, 2447–2458. [Google Scholar] [CrossRef]
- Xu, M.; Hu, Y.; Xiao, Y.; Zhang, Y.; Sun, K.; Wu, T.; Lv, N.; Wang, W.; Ding, W.; Li, F.; et al. Near-Infrared-Controlled Nanoplatform Exploiting Photothermal Promotion of Peroxidase-like and OXD-like Activities for Potent Antibacterial and Anti-Biofilm Therapies. ACS Appl. Mater. Interfaces 2020, 12, 50260–50274. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Z.; Hoop, M.; Mushtaq, F.; Siringil, E.; Hu, C.; Nelson, B.J.; Pané, S. Recent Developments in Magnetically Driven Micro- and Nanorobots. Appl. Mater. Today 2017, 9, 37–48. [Google Scholar] [CrossRef]
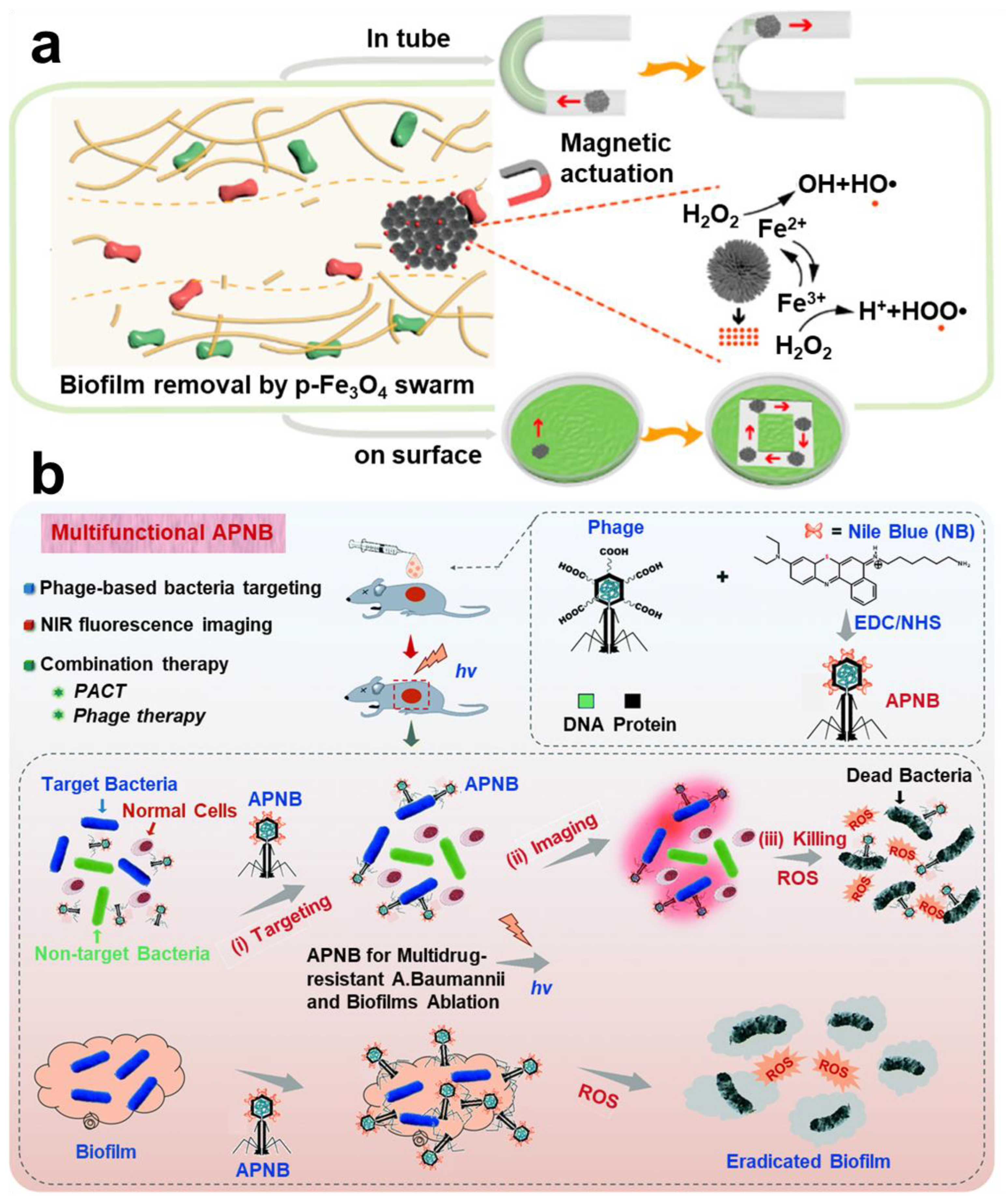
- Dong, Y.; Wang, L.; Yuan, K.; Ji, F.; Gao, J.; Zhang, Z.; Du, X.; Tian, Y.; Wang, Q.; Zhang, L. Magnetic Microswarm Composed of Porous Nanocatalysts for Targeted Elimination of Biofilm Occlusion. ACS Nano 2021, 15, 5056–5067. [Google Scholar] [CrossRef]
- Yang, G.; Wang, D.Y.; Liu, Y.; Huang, F.; Tian, S.; Ren, Y.; Liu, J.; An, Y.; van der Mei, H.C.; Busscher, H.J.; et al. In-Biofilm Generation of Nitric Oxide Using a Magnetically-Targetable Cascade-Reaction Container for Eradication of Infectious Biofilms. Bioact. Mater. 2022, 14, 321–334. [Google Scholar] [CrossRef]
- Yuan, K.; Jurado-Sánchez, B.; Escarpa, A. Dual-Propelled Lanbiotic Based Janus Micromotors for Selective Inactivation of Bacterial Biofilms. Angew. Chem. Int. Ed. Engl. 2021, 60, 4915–4924. [Google Scholar] [CrossRef]
- Javaudin, F.; Latour, C.; Debarbieux, L.; Lamy-Besnier, Q. Intestinal Bacteriophage Therapy: Looking for Optimal Efficacy. Clin. Microbiol. Rev. 2021, 34, e0013621. [Google Scholar] [CrossRef]
- Yan, W.; Banerjee, P.; Xu, M.; Mukhopadhyay, S.; Ip, M.; Carrigy, N.B.; Lechuga-Ballesteros, D.; To, K.K.W.; Leung, S.S.Y. Formulation Strategies for Bacteriophages to Target Intracellular Bacterial Pathogens. Adv. Drug Deliv. Rev. 2021, 176, 113864. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, K.; Liu, Y.; Huang, C.; Wu, M. Dynamic Impact of Virome on Colitis and Colorectal Cancer: Immunity, Inflammation, Prevention and Treatment. Semin. Cancer Biol. 2021, in press. [Google Scholar] [CrossRef]
- He, X.; Yang, Y.; Guo, Y.; Lu, S.; Du, Y.; Li, J.J.; Zhang, X.; Leung, N.L.C.; Zhao, Z.; Niu, G.; et al. Phage-Guided Targeting, Discriminative Imaging, and Synergistic Killing of Bacteria by AIE Bioconjugates. J. Am. Chem. Soc. 2020, 142, 3959–3969. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, S.B.; Mehta, M.; Sood, S.; Sharma, J. Isolation of a Novel Phage and Targeting Biofilms of Drug-Resistant Oral Enterococci. J. Glob. Infect. Dis. 2020, 12, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, L.; Brosh, Y.; Gelman, D.; Coppenhagen-Glazer, S.; Beyth, S.; Poradosu-Cohen, R.; Que, Y.A.; Beyth, N.; Hazan, R. Targeting Enterococcus faecalis Biofilms with Phage Therapy. Appl. Environ. Microbiol. 2015, 81, 2696–2705. [Google Scholar] [CrossRef] [PubMed]
- Ran, B.; Yuan, Y.; Xia, W.; Li, M.; Yao, Q.; Wang, Z.; Wang, L.; Li, X.; Xu, Y.; Peng, X. A Photo-Sensitizable Phage for Multidrug-Resistant Acinetobacter baumannii therapy and Biofilm Ablation. Chem. Sci. 2021, 12, 1054–1061. [Google Scholar] [CrossRef]
- Little, J.S.; Dedrick, R.M.; Freeman, K.G.; Cristinziano, M.; Smith, B.E.; Benson, C.A.; Jhaveri, T.A.; Baden, L.R.; Solomon, D.A.; Hatfull, G.F. Bacteriophage Treatment of Disseminated Cutaneous Mycobacterium chelonae Infection. Nat. Commun. 2022, 13, 2313. [Google Scholar] [CrossRef]
- Chapman, C.M.C.; Gibson, G.R.; Rowland, I. Effects of Single- and Multi-Strain Probiotics on Biofilm Formation and In Vitro Adhesion to Bladder Cells by Urinary Tract Pathogens. Anaerobe 2014, 27, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Bidossi, A.; de Grandi, R.; Toscano, M.; Bottagisio, M.; de Vecchi, E.; Gelardi, M.; Drago, L. Probiotics Streptococcus salivarius 24SMB and Streptococcus oralis 89a Interfere with Biofilm Formation of Pathogens of the Upper Respiratory Tract. BMC Infect. Dis. 2018, 18, 653. [Google Scholar] [CrossRef]
- Meroni, G.; Panelli, S.; Zuccotti, G.; Bandi, C.; Drago, L.; Pistone, D. Probiotics as Therapeutic Tools against Pathogenic Biofilms: Have We Found the Perfect Weapon? Microbiol. Res. 2021, 12, 916–937. [Google Scholar] [CrossRef]
- Santos, R.A.; Oliva-Teles, A.; Pousão-Ferreira, P.; Jerusik, R.; Saavedra, M.J.; Enes, P.; Serra, C.R. Isolation and Characterization of Fish-Gut Bacillus spp. as Source of Natural Antimicrobial Compounds to Fight Aquaculture Bacterial Diseases. Mar. Biotechnol. 2021, 23, 276–293. [Google Scholar] [CrossRef]
- Ibeiro, F.C.; Iglesias, M.C.; Barros, P.P.; Santos, S.S.F.; Jorge, A.O.C.; Leão, M.V.P. Lactobacillus rhamnosus Interferes with Candida albicans Adherence and Biofilm Formation: A Potential Alternative Treatment of Candidiasis. Austin J. Pharmacol. Ther. 2021, 9, 1133. [Google Scholar] [CrossRef]
- Shokouhfard, M.; Kermanshahi, R.K.; Feizabadi, M.M.; Teimourian, S.; Safari, F. Lactobacillus spp. Derived Biosurfactants Effect on Expression of Genes Involved in Proteus Mirabilis Biofilm Formation. Infect. Genet. Evol. 2022, 100, 105264. [Google Scholar] [CrossRef]
- Frohlich, K.M.; Weintraub, S.F.; Bell, J.T.; Todd, G.C.; Väre, V.Y.P.; Schneider, R.; Kloos, Z.A.; Tabe, E.S.; Cantara, W.A.; Stark, C.J.; et al. Discovery of Small-Molecule Antibiotics against a Unique TRNA-Mediated Regulation of Transcription in Gram-Positive Bacteria. ChemMedChem 2019, 14, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Seyler, T.M.; Moore, C.; Kim, H.; Ramachandran, S.; Agris, P.F. A New Promising Anti-Infective Agent Inhibits Biofilm Growth by Targeting Simultaneously a Conserved RNA Function That Controls Multiple Genes. Antibiotics 2021, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M.; Colavin, A.; Shi, H.; Czarny, T.L.; Larson, M.H.; Wong, S.; Hawkins, J.S.; Lu, C.H.S.; Koo, B.M.; Marta, E.; et al. A Comprehensive, CRISPR-Based Functional Analysis of Essential Genes in Bacteria. Cell 2016, 165, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Afonina, I.; Ong, J.; Chua, J.; Lu, T.; Kline, K.A. Multiplex Crispri System Enables the Study of Stage-Specific Biofilm Genetic Requirements in Enterococcus faecalis. mBio 2020, 11, e01101-20. [Google Scholar] [CrossRef]
- Asaad, A.M.; Soma, S.A.; Ajlan, E.; Awad, S.M. Epidemiology of Biofilm Producing Acinetobacter baumannii Nosocomial Isolates from a Tertiary Care Hospital in Egypt: A Cross-Sectional Study. Infect. Drug Resist. 2021, 14, 709–717. [Google Scholar] [CrossRef]
- Subramanian, D.; Natarajan, J. Integrated Meta-Analysis and Machine Learning Approach Identifies Acyl-CoA Thioesterase with Other Novel Genes Responsible for Biofilm Development in Staphylococcus aureus. Infect. Genet. Evol. 2021, 88, 104702. [Google Scholar] [CrossRef]
- Abdulhaq, N.; Nawaz, Z.; Zahoor, M.A.; Siddique, A.B. Association of Biofilm Formation with Multi Drug Resistance in Clinical Isolates of Pseudomonas aeruginosa. EXCLI J. 2020, 19, 201–208. [Google Scholar] [CrossRef]
- Wu, S.; Liu, Y.; Lei, L.; Zhang, H. Antisense YycG Modulates the Susceptibility of Staphylococcus aureus to Hydrogen Peroxide via the SarA. BMC Microbiol. 2021, 21, 160. [Google Scholar] [CrossRef]
- Wille, J.; Coenye, T. Biofilm Dispersion: The Key to Biofilm Eradication or Opening Pandora’s Box? Biofilm 2020, 2, 100027. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. Escaping the Biofilm in More than One Way: Desorption, Detachment or Dispersion. Curr. Opin. Microbiol. 2016, 30, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Fux, C.A.; Wilson, S.; Stoodley, P. Detachment Characteristics and Oxacillin Resistance of Staphylococcus aureus Biofilm Emboli in an In Vitro Catheter Infection Model. J. Bacteriol. 2004, 186, 4486–4491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Li, X.; Yu, C.; Wang, Y. Promising Therapeutic Strategies Against Microbial Biofilm Challenges. Front. Cell. Infect. Microbiol. 2020, 10, 359. [Google Scholar] [CrossRef] [PubMed]
- Kolodkin-Gal, I.; Romero, D.; Cao, S.; Clardy, J.; Kolter, R.; Losick, R. D-Amino Acids Trigger Biofilm Disassembly. Science 2010, 328, 624–627. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.M.; Monteiro, C.; Costa Lima, S.A.; Casal, S.; van Dijck, P.; Martins, M.C.L.; Nunes, C.; Reis, S. N-Acetyl-l-Cysteine-Loaded Nanosystems as a Promising Therapeutic Approach Toward the Eradication of Pseudomonas aeruginosa Biofilms. ACS Appl. Mater. Interfaces 2021, 13, 42329–42343. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.Y.K.; Li, M.; Chow, M.Y.T.; Ke, W.R.; Tai, W.; Chan, H.K. A Dual Action of D-Amino Acids on Anti-Biofilm Activity and Moisture-Protection of Inhalable Ciprofloxacin Powders. Eur. J. Pharm. Biopharm. 2022, 173, 132–140. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, H.; Feng, J.; Zhou, Y.; Wang, B. Synergistic Chemotherapy, Physiotherapy and Photothermal Therapy against Bacterial and Biofilms Infections through Construction of Chiral Glutamic Acid Functionalized Gold Nanobipyramids. Chem. Eng. J. 2020, 393, 124778. [Google Scholar] [CrossRef]
- Chen, M.; Wei, J.; Xie, S.; Tao, X.; Zhang, Z.; Ran, P.; Li, X. Bacterial Biofilm Destruction by Size/Surface Charge-Adaptive Micelles. Nanoscale 2019, 11, 1410–1422. [Google Scholar] [CrossRef]
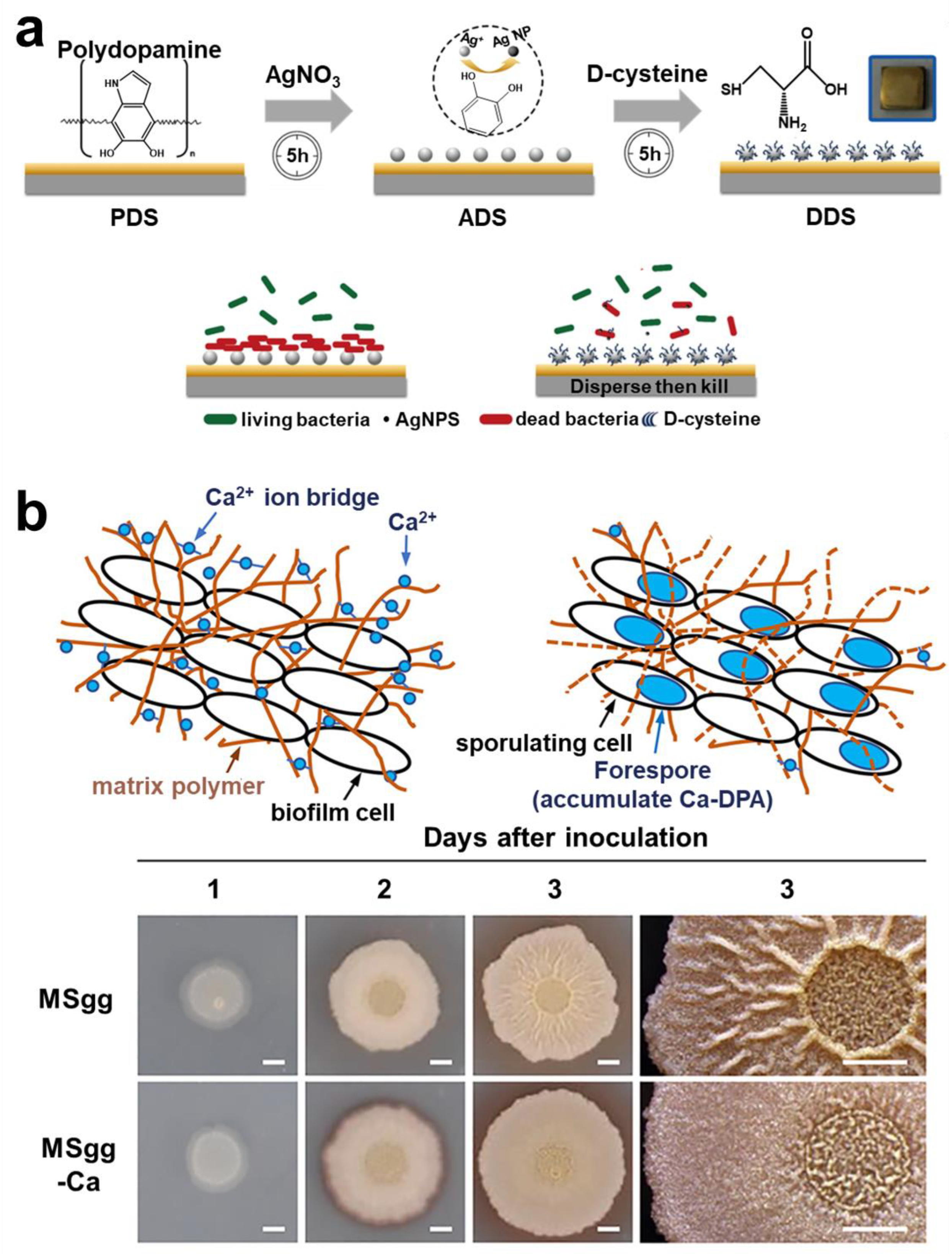
- Huang, L.; Lou, Y.; Zhang, D.; Ma, L.; Qian, H.; Hu, Y.; Ju, P.; Xu, D.; Li, X. D-Cysteine Functionalised Silver Nanoparticles Surface with a “Disperse-Then-Kill” Antibacterial Synergy. Chem. Eng. J. 2020, 381, 122662. [Google Scholar] [CrossRef]
- Liu, X.; Li, Z.; Fan, Y.; Lekbach, Y.; Song, Y.; Xu, D.; Zhang, Z.; Ding, L.; Wang, F. A Mixture of D-Amino Acids Enhances the Biocidal Efficacy of CMIT/MIT Against Corrosive Vibrio harveyi Biofilm. Front. Microbiol. 2020, 11, 557435. [Google Scholar] [CrossRef]
- Louis, M.; Clamens, T.; Tahrioui, A.; Desriac, F.; Rodrigues, S.; Rosay, T.; Harmer, N.; Diaz, S.; Barreau, M.; Racine, P.J.; et al. Pseudomonas aeruginosa Biofilm Dispersion by the Human Atrial Natriuretic Peptide. Adv. Sci. 2022, 9, 2103262. [Google Scholar] [CrossRef] [PubMed]
- Bharti, S.; Maurya, R.K.; Venugopal, U.; Singh, R.; Akhtar, M.S.; Krishnan, M.Y. Rv1717 Is a Cell Wall-Associated β-Galactosidase of Mycobacterium tuberculosis That Is Involved in Biofilm Dispersion. Front. Microbiol. 2021, 11, 611122. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.; Kobayashi, K. Calcium Prevents Biofilm Dispersion in Bacillus subtilis. J. Bacteriol. 2021, 203, e00114-21. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biofilm Types | Characteristic | Target | Pros | Cons |
|---|---|---|---|---|
| Initial adhesion stage | Reversible and Irreversible adhesion. | Adhesin and Adhesion protein. | Prevention preferable to treatment. | Stability of surface coatings. |
| Will not cause drug resistance. | Not necessarily kill bacteria. | |||
| Access not required after implantation. | Potential substrate utilization by host. | |||
| Early formation stage | Active intercellular Communication and progressive formation of EPS. | QS; Polysaccharide Intracellular Adhesin (PIA); eDNA; Polysaccharides and Proteins. | Molecular medicine. | Potential degradation by nucleases, proteins, or enzyme. |
| Controlled locally. | Highly localized. | |||
| It will affect metabolism and will not produce drug resistance. | Composition variability. | |||
| Maturation stage | Mature EPS and Gradient chemical microenvironment and Changes in bacterial metabolism. | Hypoxic; Low pH; Negative; Overexpression GSH; H2O2; Persistent and dormant bacteria. | Disrupt pathogenic microenvironment. | Difficult to simulate in vivo models. |
| Readily functionalized. | Incomplete eradication. | |||
| Active on dormant cells. | Interaction with host. | |||
| Dispersion stage | Accumulation of biofilm residues and associated secretions. | Enzymes, D-amino acids; surfactants and others. | Readily combined with antimicrobials. | Low spatiotemporal controllability. |
| Avoid cell dormancy. | Residues to be resolved. | |||
| High universality. | Release of pathogens may result in recolonization and acute infection. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bu, F.; Liu, M.; Xie, Z.; Chen, X.; Li, G.; Wang, X. Targeted Anti-Biofilm Therapy: Dissecting Targets in the Biofilm Life Cycle. Pharmaceuticals 2022, 15, 1253. https://doi.org/10.3390/ph15101253
Bu F, Liu M, Xie Z, Chen X, Li G, Wang X. Targeted Anti-Biofilm Therapy: Dissecting Targets in the Biofilm Life Cycle. Pharmaceuticals. 2022; 15(10):1253. https://doi.org/10.3390/ph15101253
Chicago/Turabian StyleBu, Fanqiang, Mengnan Liu, Zixu Xie, Xinyu Chen, Guofeng Li, and Xing Wang. 2022. "Targeted Anti-Biofilm Therapy: Dissecting Targets in the Biofilm Life Cycle" Pharmaceuticals 15, no. 10: 1253. https://doi.org/10.3390/ph15101253
APA StyleBu, F., Liu, M., Xie, Z., Chen, X., Li, G., & Wang, X. (2022). Targeted Anti-Biofilm Therapy: Dissecting Targets in the Biofilm Life Cycle. Pharmaceuticals, 15(10), 1253. https://doi.org/10.3390/ph15101253