
Moringa concanensis L. Alleviates DNCB-Induced Atopic Dermatitis-like Symptoms by Inhibiting NLRP3 Inflammasome-Mediated IL-1β in BALB/c Mice

 , , and
, , and
Abstract
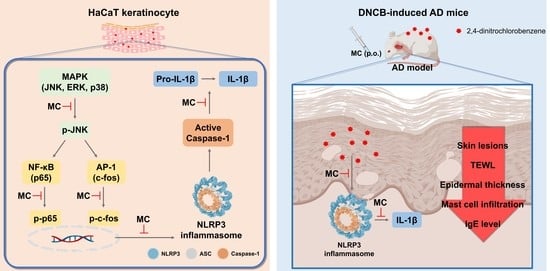
1. Introduction
2. Results
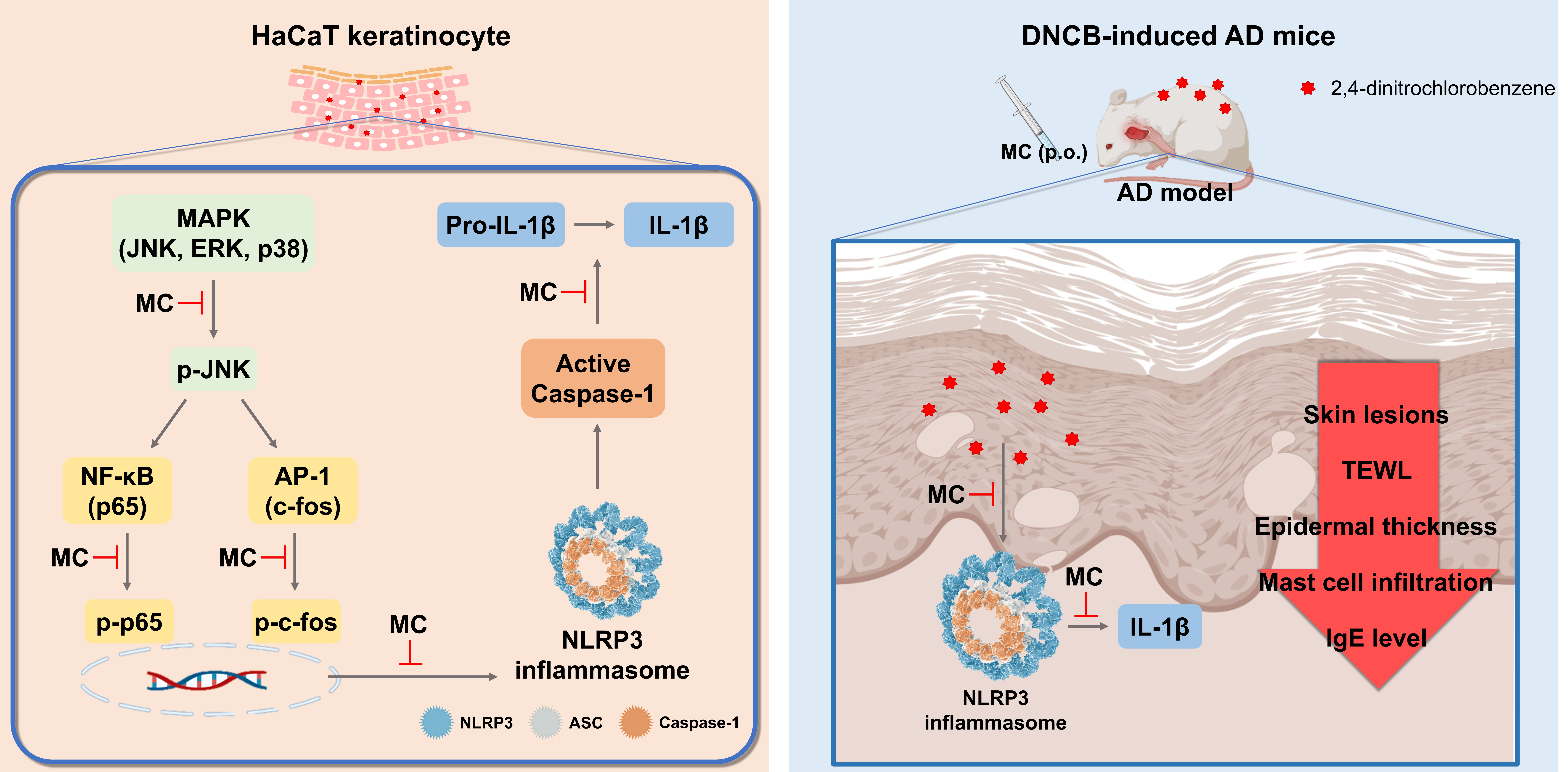
2.1. Analysis of Quadrupole Time-of-Flight (Q-TOF) Mass Spectrometry of M. concanensis
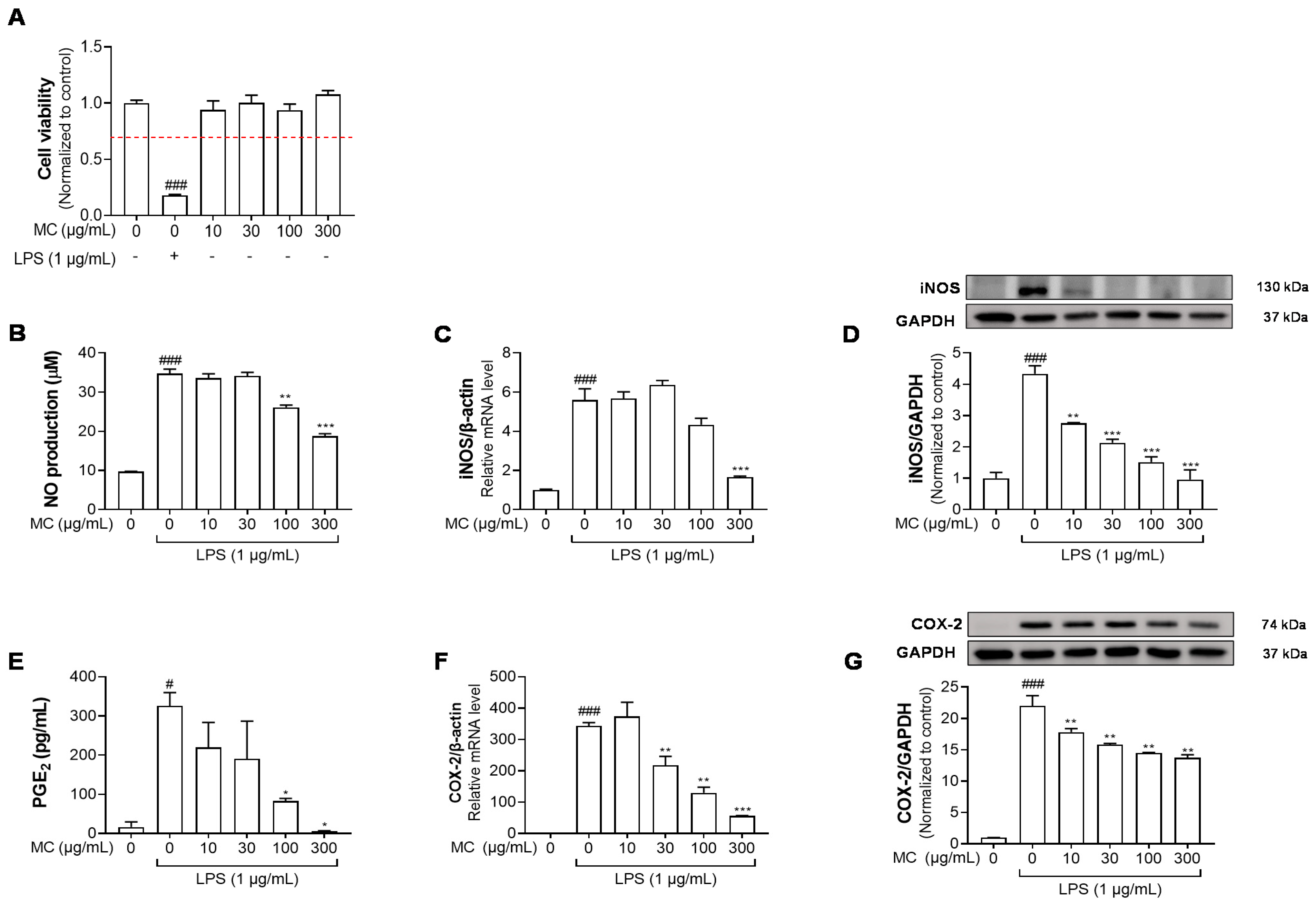
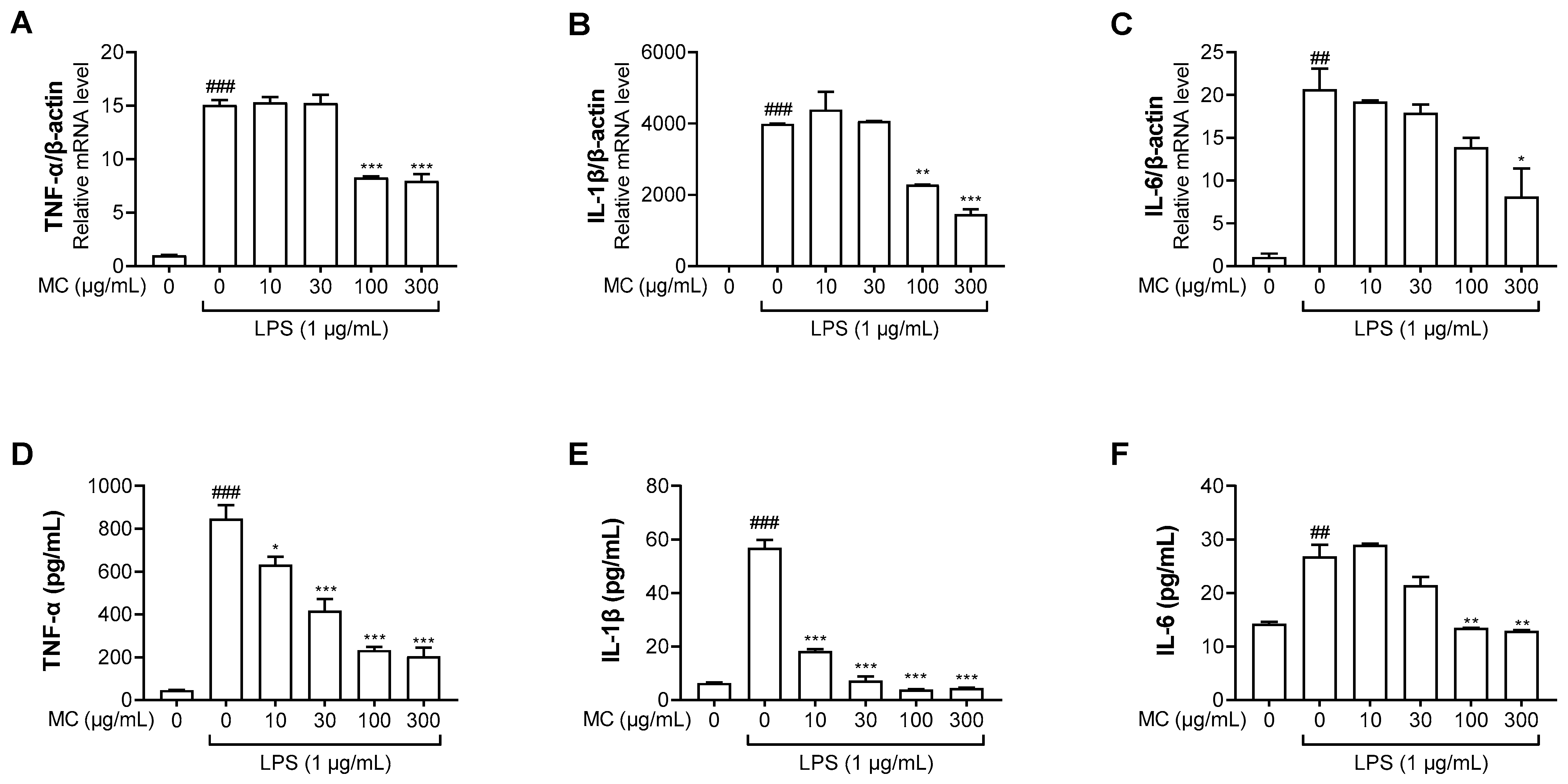
2.2. M. concanensis Inhibits the LPS-Stimulated Inflammatory Mediators in HaCaT Cells
2.3. M. concanensis Downregulated the Expression of Inflammatory Cytokines in LPS-Stimulated HaCaT Cells
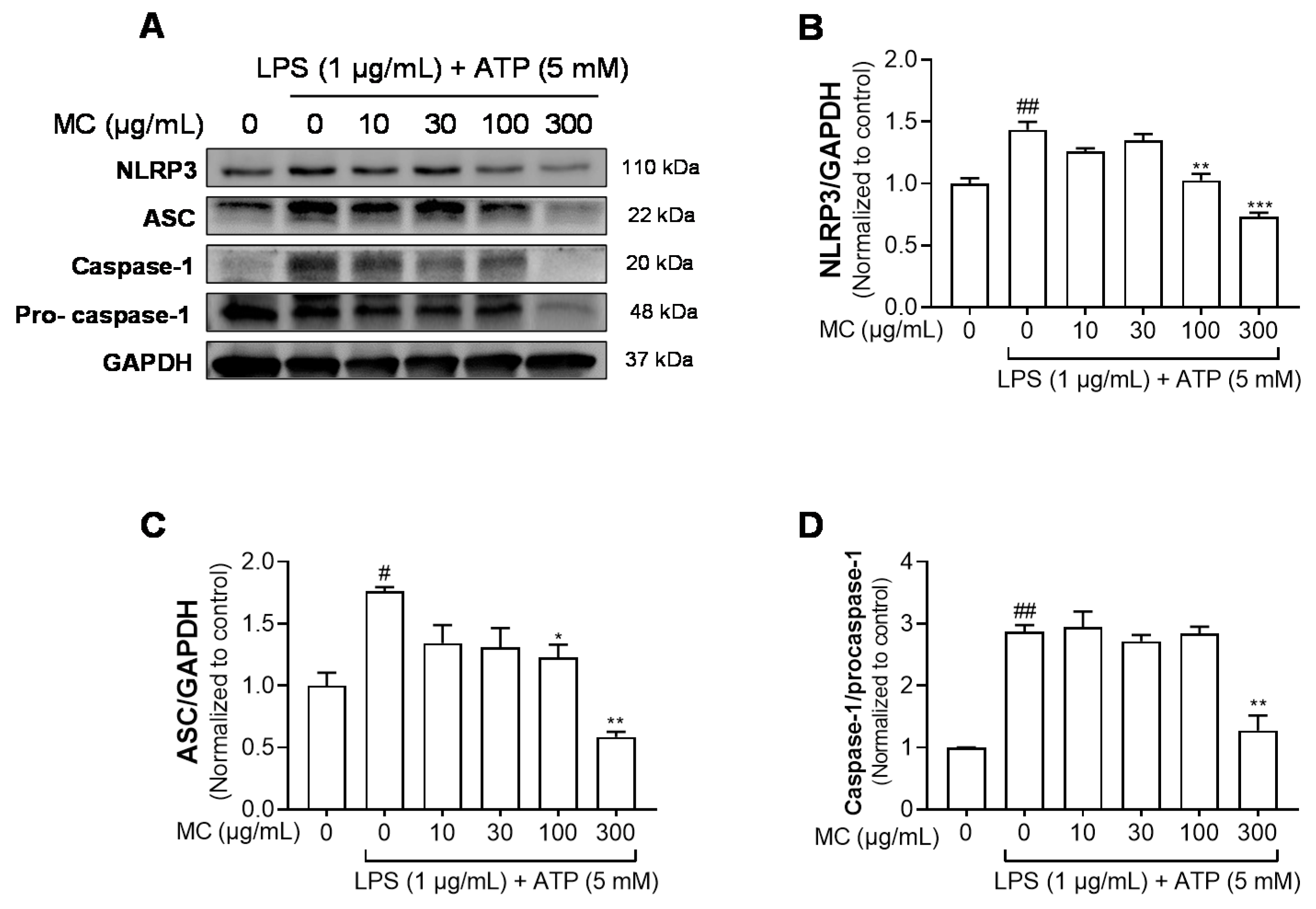
2.4. M. concanensis Reduced the Expression of IL-1β by Inhibiting the NLRP3 Inflammasome in HaCaT Cells
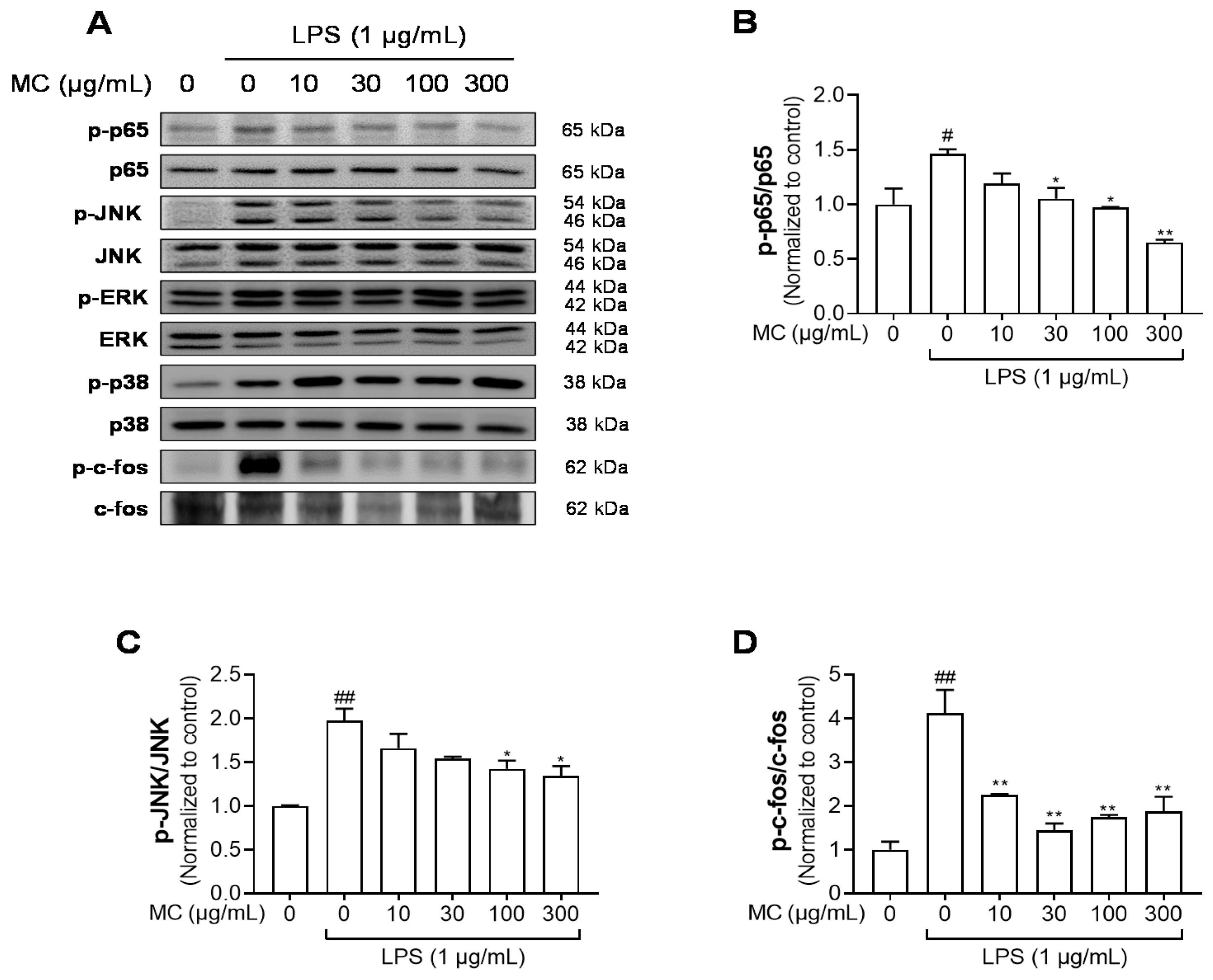
2.5. M. concanensis Inhibited the Phosphorylation of NF-κB, MAPK and AP-1 in HaCaT Cells Stimulated with LPS
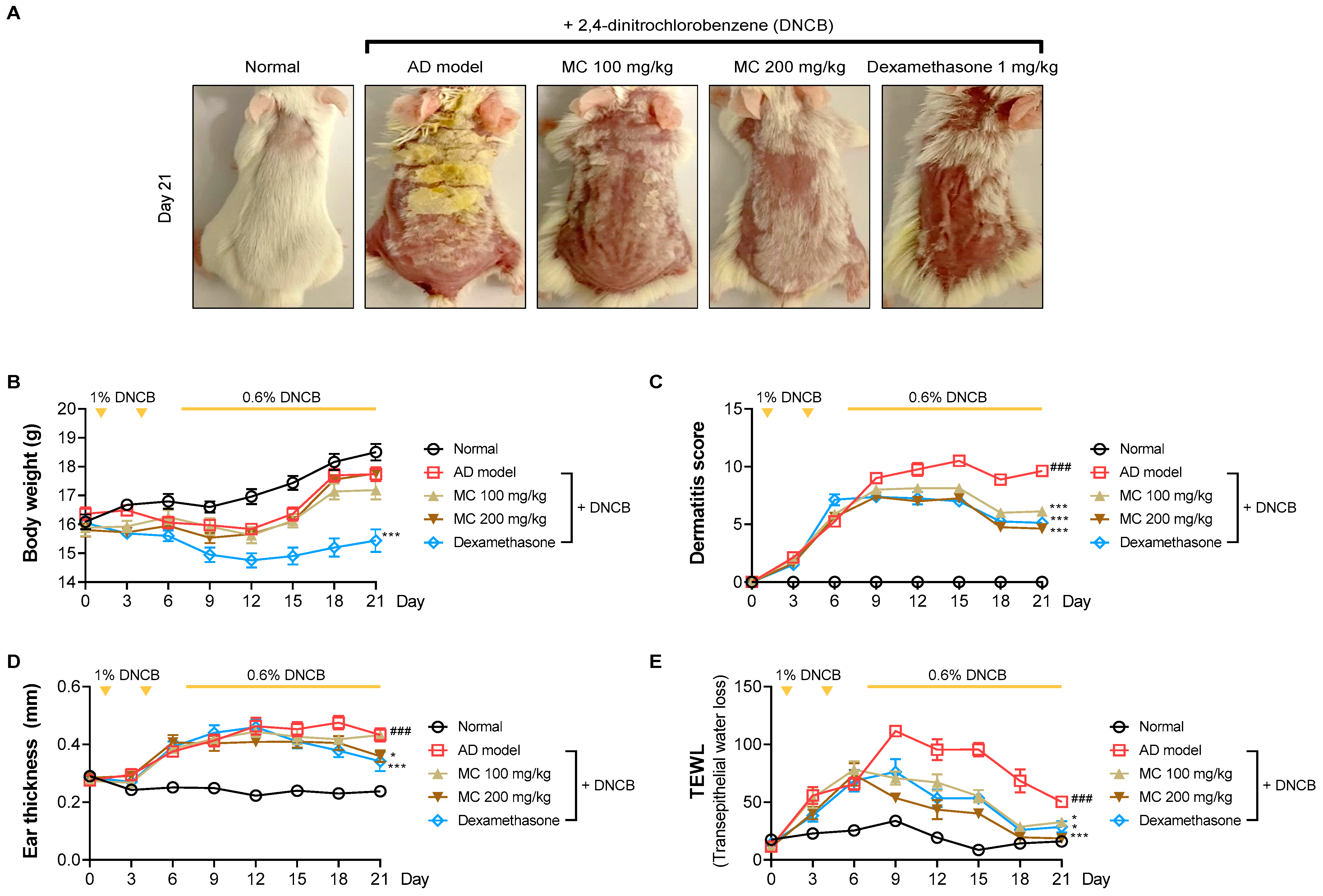
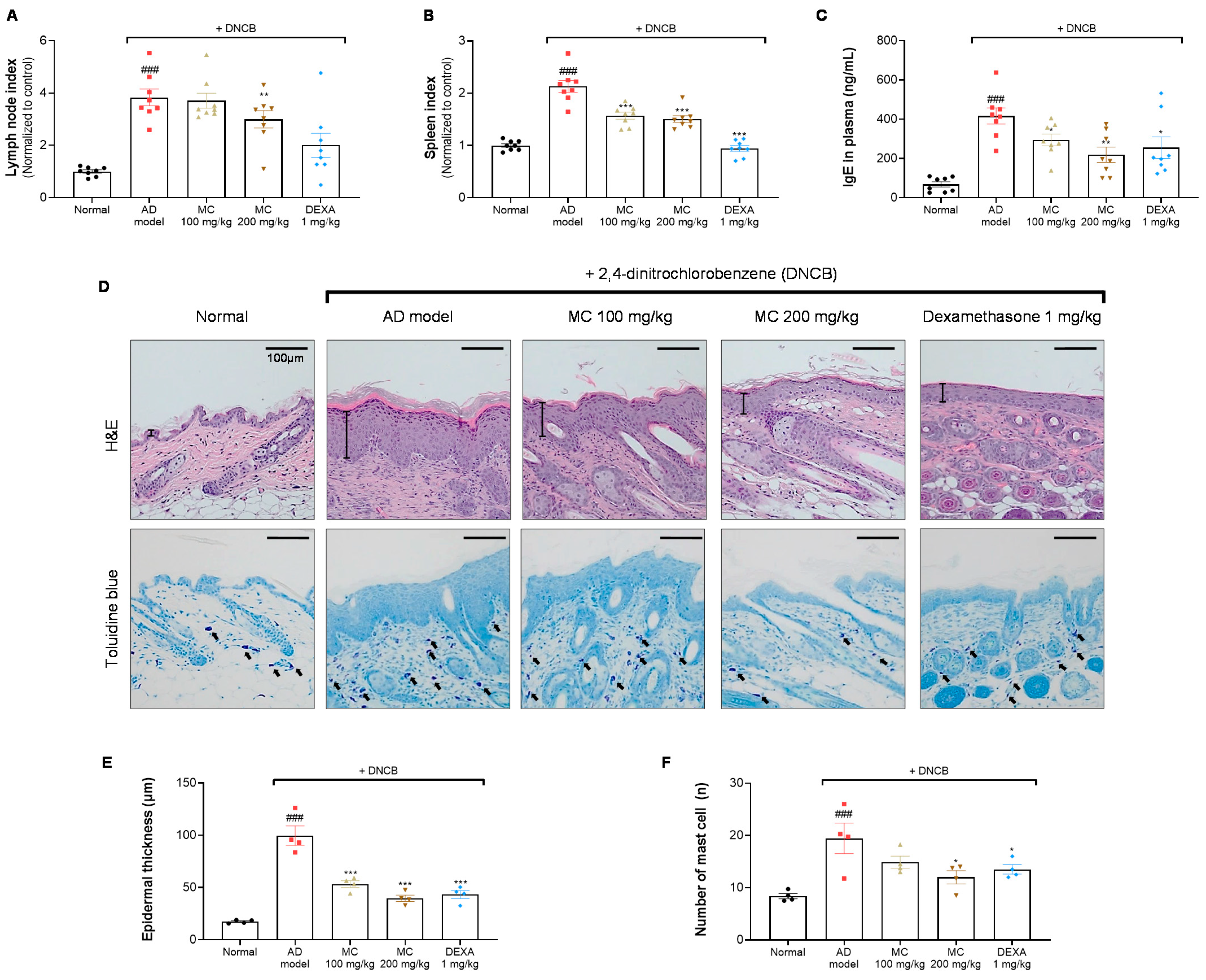
2.6. M. concanensis Improved the Clinical Symptoms in Mice with AD-like Skin Lesions Induced by DNCB
2.7. M. concanensis Ameliorated the Immunological and Histological Changes in DNCB-Challenged BALB/c Mice
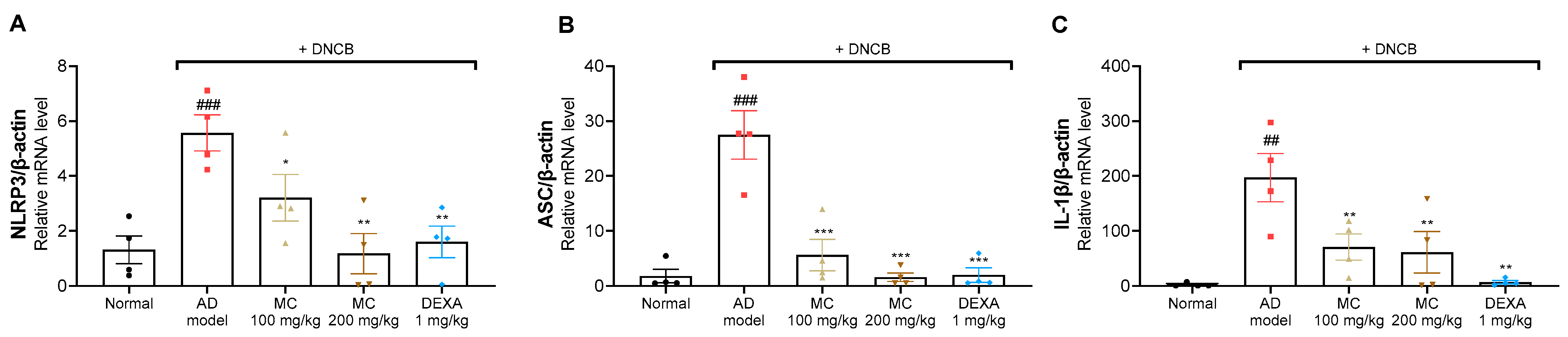
2.8. M. concanensis Inhibited the Activation of the NLRP3 Inflammasome in DNCB-Treated BALB/c Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of an Ethanolic Extract of M. concanensis
4.3. Identification of Phytochemicals in M. concanensis by UPLC-QTOF-MS/MS
4.4. Materials
4.5. Cell Culture
4.6. Cell Viability
4.7. Nitric Oxide Production
4.8. RNA Extraction and Real Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.9. PGE2, TNF-α, IL-1β, IL-6 and IgE Assays
4.10. Western Blot Analysis
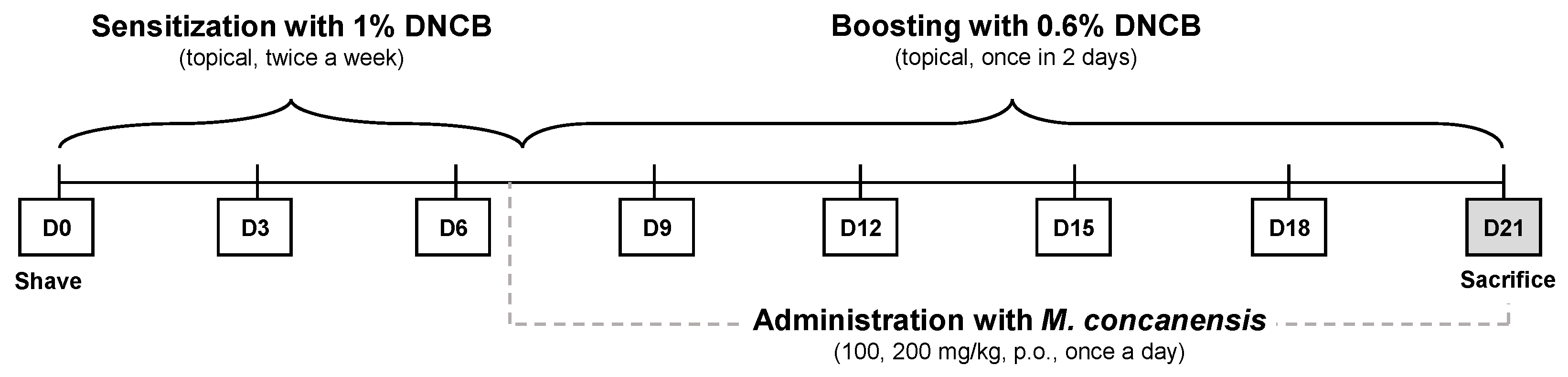
4.11. 2,4-Dinitrochlorobenzene (DNCB)-Induced Atopic Dermatitis Mice
4.12. Measurement of Clinical Symptoms and Histological Changes
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Foley, C.; Tundia, N.; Simpson, E.; Teixeira, H.D.; Litcher-Kelly, L.; Bodhani, A. Development and content validity of new patient-reported outcome questionnaires to assess the signs and symptoms and impact of atopic dermatitis: The Atopic Dermatitis Symptom Scale (ADerm-SS) and the Atopic Dermatitis Impact Scale (ADerm-IS). Curr. Med. Res. Opin. 2019, 35, 1139–1148. [Google Scholar] [CrossRef]
- Zyriax, B.C.; Augustin, M.; Abeck, F.; Mohr, N.; Kirsten, N.; Langenbruch, A. Adherence to Guideline-Oriented Preventive Measures in Patients with Atopic Dermatitis in Germany. Dermatology 2021, 238, 307–312. [Google Scholar] [CrossRef]
- Silverberg, J.I.; Barbarot, S.; Gadkari, A.; Simpson, E.L.; Weidinger, S.; Mina-Osorio, P.; Rossi, A.B.; Brignoli, L.; Saba, G.; Guillemin, I.; et al. Atopic dermatitis in the pediatric population: A cross-sectional, international epidemiologic study. Ann. Allergy Asthma Immunol. 2021, 126, 417–428.e2. [Google Scholar] [CrossRef]
- Wei, W.; Anderson, P.; Gadkari, A.; Blackburn, S.; Moon, R.; Piercy, J.; Shinde, S.; Gomez, J.; Ghorayeb, E. Extent and consequences of inadequate disease control among adults with a history of moderate to severe atopic dermatitis. J. Dermatol. 2018, 45, 150–157. [Google Scholar] [CrossRef]
- Eichenfield, L.F.; Tom, W.L.; Chamlin, S.L.; Feldman, S.R.; Hanifin, J.M.; Simpson, E.L.; Berger, T.G.; Bergman, J.N.; Cohen, D.E.; Cooper, K.D.; et al. Guidelines of care for the management of atopic dermatitis: Section 1. Diagnosis and assessment of atopic dermatitis. J. Am. Acad. Dermatol. 2014, 70, 338–351. [Google Scholar] [CrossRef]
- Wollenberg, A.; Kraft, S.; Oppel, T.; Bieber, T. Atopic dermatitis: Pathogenetic mechanisms. Clin. Exp. Dermatol. 2000, 25, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Eichenfield, L.F.; Hanifin, J.M.; Beck, L.A.; Lemanske, R.F., Jr.; Sampson, H.A.; Weiss, S.T.; Leung, D.Y. Atopic dermatitis and asthma: Parallels in the evolution of treatment. Pediatrics 2003, 111, 608–616. [Google Scholar] [CrossRef]
- Hussain, Z.; Sahudin, S.; Thu, H.E.; Shuid, A.N.; Bukhari, S.N.; Kumolosasi, E. Recent Advances in Pharmacotherapeutic Paradigm of Mild to Recalcitrant Atopic Dermatitis. Crit. Rev. Ther. Drug Carr. Syst. 2016, 33, 213–263. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Shi, T.; Hou, J.; Li, M. Ferulic acid alleviates atopic dermatitis-like symptoms in mice via its potent anti-inflammatory effect. Immunopharmacol. Immunotoxicol. 2020, 42, 156–164. [Google Scholar] [CrossRef]
- Rawlings, A.V.; Harding, C.R. Moisturization and skin barrier function. Dermatol. Ther. 2004, 17 (Suppl. 1), 43–48. [Google Scholar] [CrossRef]
- Zhu, T.H.; Zhu, T.R.; Tran, K.A.; Sivamani, R.K.; Shi, V.Y. Epithelial barrier dysfunctions in atopic dermatitis: A skin-gut-lung model linking microbiome alteration and immune dysregulation. Br. J. Dermatol. 2018, 179, 570–581. [Google Scholar] [CrossRef]
- Latz, E.; Xiao, T.S.; Stutz, A. Activation and regulation of the inflammasomes. Nat. Rev. Immunol. 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.H.; Ellis, L.Z.; Fujita, M. Inflammasomes as molecular mediators of inflammation and cancer: Potential role in melanoma. Cancer Lett. 2012, 314, 24–33. [Google Scholar] [CrossRef]
- Zurier, R.B.; Rossetti, R.G.; Burstein, S.H.; Bidinger, B. Suppression of human monocyte interleukin-1beta production by ajulemic acid, a nonpsychoactive cannabinoid. Biochem. Pharmacol. 2003, 65, 649–655. [Google Scholar] [CrossRef]
- Dai, X.; Sayama, K.; Tohyama, M.; Shirakata, Y.; Hanakawa, Y.; Tokumaru, S.; Yang, L.; Hirakawa, S.; Hashimoto, K. Mite allergen is a danger signal for the skin via activation of inflammasome in keratinocytes. J. Allergy Clin. Immunol. 2011, 127, 806–814.e4. [Google Scholar] [CrossRef] [PubMed]
- Anbazhakan, S.; Dhandapani, R.; Anandhakumar, P.; Balu, S. Traditional Medicinal Knowledge on Moringa concanensis Nimmo of Perambalur District, Tamilnadu. Anc. Sci. Life 2007, 26, 42–45. [Google Scholar]
- Balakrishnan, B.B.; Krishnasamy, K.; Choi, K.C. Moringa concanensis Nimmo ameliorates hyperglycemia in 3T3-L1 adipocytes by upregulating PPAR-gamma, C/EBP-alpha via Akt signaling pathway and STZ-induced diabetic rats. Biomed. Pharmacother. 2018, 103, 719–728. [Google Scholar] [CrossRef]
- Maitreya, C.S.a.B. Seasonal analysis of phytochemicals in Moringa concanensis Nimmo ex Dalz. and Gibson from south saurasthara zone, Junagadh-Gujarat, India. Int. Assoc. Biol. Comput. Dig. 2021, 6, 116–125. [Google Scholar]
- Hur, S.S.; Choi, S.W.; Lee, D.R.; Park, J.H.; Chung, T.H. Advanced Effect of Moringa oleifera Bioconversion by Rhizopus oligosporus on the Treatment of Atopic Dermatitis: Preliminary Study. Evid. Based Complement. Altern. Med. 2018, 2018, 7827565. [Google Scholar] [CrossRef]
- Choi, E.J.; Debnath, T.; Tang, Y.; Ryu, Y.B.; Moon, S.H.; Kim, E.K. Topical application of Moringa oleifera leaf extract ameliorates experimentally induced atopic dermatitis by the regulation of Th1/Th2/Th17 balance. Biomed. Pharmacother. 2016, 84, 870–877. [Google Scholar] [CrossRef]
- Zeng, K.; Thompson, K.E.; Yates, C.R.; Miller, D.D. Synthesis and biological evaluation of quinic acid derivatives as anti-inflammatory agents. Bioorg. Med. Chem. Lett. 2009, 19, 5458–5460. [Google Scholar] [CrossRef]
- Pragasam, S.J.; Venkatesan, V.; Rasool, M. Immunomodulatory and anti-inflammatory effect of p-coumaric acid, a common dietary polyphenol on experimental inflammation in rats. Inflammation 2013, 36, 169–176. [Google Scholar] [CrossRef]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Sreedhar, R.; Giridharan, V.V.; Watanabe, K. Molecular targets of quercetin with anti-inflammatory properties in atopic dermatitis. Drug Discov. Today 2016, 21, 632–639. [Google Scholar] [CrossRef]
- Chanjitwiriya, K.; Roytrakul, S.; Kunthalert, D. Quercetin negatively regulates IL-1beta production in Pseudomonas aeruginosa-infected human macrophages through the inhibition of MAPK/NLRP3 inflammasome pathways. PLoS ONE 2020, 15, e0237752. [Google Scholar] [CrossRef]
- Xue, Y.; Du, M.; Zhu, M.J. Quercetin suppresses NLRP3 inflammasome activation in epithelial cells triggered by Escherichia coli O157:H7. Free Radic. Biol. Med. 2017, 108, 760–769. [Google Scholar] [CrossRef]
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of atopic dermatitis: Clinical implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef]
- Ahn, S.; Siddiqi, M.H.; Aceituno, V.C.; Simu, S.Y.; Zhang, J.; Jimenez Perez, Z.E.; Kim, Y.J.; Yang, D.C. Ginsenoside Rg5:Rk1 attenuates TNF-α/IFN-γ-induced production of thymus- and activation-regulated chemokine (TARC/CCL17) and LPS-induced NO production via downregulation of NF-κB/p38 MAPK/STAT1 signaling in human keratinocytes and macrophages. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 287–295. [Google Scholar] [CrossRef]
- Lim, J.S.; Kim, J.Y.; Lee, S.; Choi, J.K.; Kim, E.N.; Choi, Y.A.; Jang, Y.H.; Jeong, G.S.; Kim, S.H. Bakuchicin attenuates atopic skin inflammation. Biomed. Pharmacother. 2020, 129, 110466. [Google Scholar] [CrossRef]
- Zhou, B.R.; Zhang, J.A.; Zhang, Q.; Permatasari, F.; Xu, Y.; Wu, D.; Yin, Z.Q.; Luo, D. Palmitic acid induces production of proinflammatory cytokines interleukin-6, interleukin-1beta, and tumor necrosis factor-alpha via a NF-κB-dependent mechanism in HaCaT keratinocytes. Mediat. Inflamm. 2013, 2013, 530429. [Google Scholar] [CrossRef]
- Zito, G.; Buscetta, M.; Cimino, M.; Dino, P.; Bucchieri, F.; Cipollina, C. Cellular Models and Assays to Study NLRP3 Inflammasome Biology. Int. J. Mol. Sci. 2020, 21, 4294. [Google Scholar] [CrossRef]
- Strowig, T.; Henao-Mejia, J.; Elinav, E.; Flavell, R. Inflammasomes in health and disease. Nature 2012, 481, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Yang, Y.; Wang, Q.; Li, M.; Tian, C.; Liu, Y.; Aung, L.H.H.; Li, P.F.; Yu, T.; Chu, X.M. NLRP3 inflammasome in endothelial dysfunction. Cell Death Dis. 2020, 11, 776. [Google Scholar] [CrossRef] [PubMed]
- Budai, M.M.; Varga, A.; Milesz, S.; Tozser, J.; Benko, S. Aloe vera downregulates LPS-induced inflammatory cytokine production and expression of NLRP3 inflammasome in human macrophages. Mol. Immunol. 2013, 56, 471–479. [Google Scholar] [CrossRef]
- Subedi, L.; Lee, J.H.; Yumnam, S.; Ji, E.; Kim, S.Y. Anti-Inflammatory Effect of Sulforaphane on LPS-Activated Microglia Potentially through JNK/AP-1/NF-κB Inhibition and Nrf2/HO-1 Activation. Cells 2019, 8, 194. [Google Scholar] [CrossRef]
- Zitnik, S.E.; Ruschendorf, F.; Muller, S.; Sengler, C.; Lee, Y.A.; Griffioen, R.W.; Meglio, P.; Wahn, U.; Witt, H.; Nickel, R. IL13 variants are associated with total serum IgE and early sensitization to food allergens in children with atopic dermatitis. Pediatr. Allergy Immunol. 2009, 20, 551–555. [Google Scholar] [CrossRef]
- Zheng, J.; Yao, L.; Zhou, Y.; Gu, X.; Wang, C.; Bao, K.; Sun, Y.; Hong, M. A novel function of NLRP3 independent of inflammasome as a key transcription factor of IL-33 in epithelial cells of atopic dermatitis. Cell Death Dis. 2021, 12, 871. [Google Scholar] [CrossRef]
- Garmhausen, D.; Hagemann, T.; Bieber, T.; Dimitriou, I.; Fimmers, R.; Diepgen, T.; Novak, N. Characterization of different courses of atopic dermatitis in adolescent and adult patients. Allergy 2013, 68, 498–506. [Google Scholar] [CrossRef]
- Illi, S.; von Mutius, E.; Lau, S.; Nickel, R.; Gruber, C.; Niggemann, B.; Wahn, U.; The Multicenter Allergy Study Group. The natural course of atopic dermatitis from birth to age 7 years and the association with asthma. J. Allergy Clin. Immunol. 2004, 113, 925–931. [Google Scholar] [CrossRef]
- Skabytska, Y.; Kaesler, S.; Volz, T.; Biedermann, T. How the innate immune system trains immunity: Lessons from studying atopic dermatitis and cutaneous bacteria. J. Dtsch. Dermatol. Ges. 2016, 14, 153–156. [Google Scholar] [CrossRef][Green Version]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Hajar, T.; Leshem, Y.A.; Hanifin, J.M.; Nedorost, S.T.; Lio, P.A.; Paller, A.S.; Block, J.; Simpson, E.L. A systematic review of topical corticosteroid withdrawal (“steroid addiction”) in patients with atopic dermatitis and other dermatoses. J. Am. Acad. Dermatol. 2015, 72, 541–549.e2. [Google Scholar] [CrossRef] [PubMed]
- Gutfreund, K.; Bienias, W.; Szewczyk, A.; Kaszuba, A. Topical calcineurin inhibitors in dermatology. Part I: Properties, method and effectiveness of drug use. Postep. Dermatol. Alergol. 2013, 30, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Eichenfield, L.F.; Tom, W.L.; Berger, T.G.; Krol, A.; Paller, A.S.; Schwarzenberger, K.; Bergman, J.N.; Chamlin, S.L.; Cohen, D.E.; Cooper, K.D.; et al. Guidelines of care for the management of atopic dermatitis: Section 2. Management and treatment of atopic dermatitis with topical therapies. J. Am. Acad. Dermatol. 2014, 71, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef]
- Begon, E.; Michel, L.; Flageul, B.; Beaudoin, I.; Jean-Louis, F.; Bachelez, H.; Dubertret, L.; Musette, P. Expression, subcellular localization and cytokinic modulation of Toll-like receptors (TLRs) in normal human keratinocytes: TLR2 up-regulation in psoriatic skin. Eur. J. Dermatol. 2007, 17, 497–506. [Google Scholar] [CrossRef]
- Lee, S.H.; Kwon, N.S.; Baek, K.J.; Yun, H.Y.; Kim, D.S. LGI3 is secreted and binds to ADAM22 via TRIF-dependent NF-κB pathway in response to LPS in human keratinocytes. Cytokine 2020, 126, 154872. [Google Scholar] [CrossRef]
- Tang, L.; Zhou, F. Inflammasomes in Common Immune-Related Skin Diseases. Front. Immunol. 2020, 11, 882. [Google Scholar] [CrossRef]
- Abd Rani, N.Z.; Husain, K.; Kumolosasi, E. Moringa Genus: A Review of Phytochemistry and Pharmacology. Front. Pharmacol. 2018, 9, 108. [Google Scholar] [CrossRef]
- Arulselvan, P.; Tan, W.S.; Gothai, S.; Muniandy, K.; Fakurazi, S.; Esa, N.M.; Alarfaj, A.A.; Kumar, S.S. Anti-Inflammatory Potential of Ethyl Acetate Fraction of Moringa oleifera in Downregulating the NF-κB Signaling Pathway in Lipopolysaccharide-Stimulated Macrophages. Molecules 2016, 21, 1452. [Google Scholar] [CrossRef]
- Tamrat, Y.; Nedi, T.; Assefa, S.; Teklehaymanot, T.; Shibeshi, W. Anti-inflammatory and analgesic activities of solvent fractions of the leaves of Moringa stenopetala Bak. (Moringaceae) in mice models. BMC Complement. Altern. Med. 2017, 17, 473. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.S.; Arulselvan, P.; Karthivashan, G.; Fakurazi, S. Moringa oleifera Flower Extract Suppresses the Activation of Inflammatory Mediators in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages via NF-κB Pathway. Mediat. Inflamm. 2015, 2015, 720171. [Google Scholar] [CrossRef] [PubMed]
- Chitra, M.J.a.M. Evaluation of anti-inflammatory, analgesic and antipyretic activity of Moringa concanensis Nimmo. J. Chem. Pharm. Res. 2011, 3, 802–806. [Google Scholar]
- Douglas, T.; Champagne, C.; Morizot, A.; Lapointe, J.M.; Saleh, M. The Inflammatory Caspases-1 and -11 Mediate the Pathogenesis of Dermatitis in Sharpin-Deficient Mice. J. Immunol. 2015, 195, 2365–2373. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, K.; Yamate, Y.; Yokoyama, S. Ultraviolet B eye irradiation aggravates atopic dermatitis via adrenocorticotropic hormone and NLRP3 inflammasome in NC/Nga mice. Photodermatol. Photoimmunol. Photomed. 2018, 34, 200–210. [Google Scholar] [CrossRef]
- Xiao, Y.; Xu, W.; Su, W. NLRP3 inflammasome: A likely target for the treatment of allergic diseases. Clin. Exp. Allergy 2018, 48, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Bhattacharyya, S.; Pal, T.K. Antioxidant Activities of Moringa concanensis Flowers (fresh and dried) Grown in West Bengal. Int. J. Res. Chem. Environ. 2014, 4, 64–70. [Google Scholar]
- Balakrishnan, B.B.; Krishnasamy, K. Evaluation of Free Radical Screening and Antioxidant Potential of Moringa concanensis Nimmo-a Medicinal Plant Used in Indian Traditional Medication System. Int. J. Pharm. Pharm. Sci. 2018, 10, 91–97. [Google Scholar] [CrossRef]
- Comalada, M.; Camuesco, D.; Sierra, S.; Ballester, I.; Xaus, J.; Gálvez, J.; Zarzuelo, A. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through down-regulation of the NF-κB pathway. Eur. J. Immunol. 2005, 35, 584–592. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Lee, S.Y.; Moon, E.; Kim, S.Y.; Lee, K.R. Quinic acid derivatives from Pimpinella brachycarpa exert anti-neuroinflammatory activity in lipopolysaccharide-induced microglia. Bioorg. Med. Chem. Lett. 2013, 23, 2140–2144. [Google Scholar] [CrossRef] [PubMed]
- Domiciano, T.P.; Wakita, D.; Jones, H.D.; Crother, T.R.; Verri, W.A., Jr.; Arditi, M.; Shimada, K. Quercetin Inhibits Inflammasome Activation by Interfering with ASC Oligomerization and Prevents Interleukin-1 Mediated Mouse Vasculitis. Sci. Rep. 2017, 7, 41539. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xu, X.; Li, Y.; Kou, J.; Huang, F.; Liu, B.; Liu, K. Quercetin, luteolin and epigallocatechin gallate alleviate TXNIP and NLRP3-mediated inflammation and apoptosis with regulation of AMPK in endothelial cells. Eur. J. Pharmacol. 2014, 745, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Park, S.; Kim, H.; Choi, G.J.; Kim, S.H. Application of UPLC-QTOF-MS Based Untargeted Metabolomics in Identification of Metabolites Induced in Pathogen-Infected Rice. Plants 2021, 10, 213. [Google Scholar] [CrossRef]
- Ko, W.K.; Lee, S.H.; Kim, S.J.; Jo, M.J.; Kumar, H.; Han, I.B.; Sohn, S. Anti-inflammatory effects of ursodeoxycholic acid by lipopolysaccharide-stimulated inflammatory responses in RAW 264.7 macrophages. PLoS ONE 2017, 12, e0180673. [Google Scholar] [CrossRef]
- Yang, Y.I.; Woo, J.H.; Seo, Y.J.; Lee, K.T.; Lim, Y.; Choi, J.H. Protective Effect of Brown Alga Phlorotannins against Hyper-inflammatory Responses in Lipopolysaccharide-Induced Sepsis Models. J. Agric. Food Chem. 2016, 64, 570–578. [Google Scholar] [CrossRef]
- Oranje, A.P.; Glazenburg, E.J.; Wolkerstorfer, A.; de Waard-van der Spek, F.B. Practical issues on interpretation of scoring atopic dermatitis: The SCORAD index, objective SCORAD and the three-item severity score. Br. J. Dermatol. 2007, 157, 645–648. [Google Scholar] [CrossRef]
- Logger, J.G.M.; Driessen, R.J.B.; de Jong, E.; van Erp, P.E.J. Value of GPSkin for the measurement of skin barrier impairment and for monitoring of rosacea treatment in daily practice. Skin Res. Technol. 2021, 27, 15–23. [Google Scholar] [CrossRef]
- Yang, H.; Jung, E.M.; Ahn, C.; Lee, G.S.; Lee, S.Y.; Kim, S.H.; Choi, I.G.; Park, M.J.; Lee, S.S.; Choi, D.H.; et al. Elemol from Chamaecyparis obtusa ameliorates 2,4-dinitrochlorobenzene-induced atopic dermatitis. Int. J. Mol. Med. 2015, 36, 463–472. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) | Tentative Identification | Formula | m/z [M-H]− | Mass Error (ppm) | Response | Fragmentation (m/z) |
|---|---|---|---|---|---|---|
| 0.46 | Maltose | C12H22O11 | 341.1090 | −1.3 | 1,038,656 | 179.0561 |
| 0.47 | Quinic acid | C7H12O6 | 191.0559 | −1.3 | 1,983,085 | 85.0301 |
| 0.50 | Coumaroylquinic acid | C16H18O8 | 337.0918 | −3.3 | 111,547 | 191.0561 |
| 0.62 | Malic acid | C4H6O5 | 133.0137 | −3.9 | 27,811 | 89.0246, 114.9995 |
| 0.89 | 4-O-Caffeoylquinic acid | C16H18O9 | 353.0874 | −1.2 | 180,161 | 135.0452, 173.0445, 191.0562 |
| 0.98 | Esculin | C15H16O9 | 339.0715 | −2 | 2322 | 177.0191 |
| 1.03 | tryptophan | C11H12N2O2 | 203.0823 | −1.5 | 31,577 | 116.0510, 142.0665 |
| 1.13 | Coumaric acid | C9H8O3 | 163.0402 | 1.1 | 44,900 | 119.0504 |
| 1.15 | Coumaroylquinic acid | C16H18O8 | 337.0926 | −0.9 | 505,720 | 163.0402, 173.0454, 191.0562 |
| 1.24 | Hydroxybenzoic acid | C7H6O3 | 137.0244 | −0.2 | 14,703 | 93.0351 |
| 1.25 | Coumaric acid | C9H8O3 | 163.0403 | 1.1 | 7702 | 93.0351, 119.0505 |
| 1.30 | 4-Feruloylquinic acid | C17H20O9 | 367.1030 | −1.3 | 25,889 | 134.0376, 193.0509 |
| 1.44 | Apigenin 6,8 C-dihexose | C27H30O15 | 593.1513 | 0.3 | 626,091 | 353.0671, 473.1010 |
| 1.49 | Coumaroylquinic acid | C16H18O8 | 337.0926 | −0.8 | 131,771 | 173.0455, 191.0562 |
| 1.55 | Coumaroylquinic acid | C16H18O8 | 337.0926 | −0.8 | 110,283 | 163.0402, 173.0454 |
| 1.73 | Orientin | C21H20O11 | 447.0934 | 0.2 | 2271 | 327.052 |
| 1.78 | isopentyl β-primeveroside | C16H30O10 | 381.1766 | 0.6 | 1,128,403 | 249.135 |
| 2.09 | Quercetin hydroxymethylglutaroyl glycoside | C27H28O16 | 607.1307 | 0.4 | 51,494 | 300.0287 |
| 2.15 | Vitexin | C21H20O10 | 431.0983 | −0.3 | 135,245 | 283.0617, 311.0566, 341.0674 |
| 2.31 | Kaempferol-3-O-β-D-glucopyranoside | C21H20O11 | 447.0936 | 0.8 | 3530 | 285.041 |
| 2.45 | Nicotiflorin | C27H30O15 | 593.1520 | 1.3 | 576,122 | 285.0409 |
| 2.54 | Isorhamnetin 3-O-rutinoside | C28H32O16 | 623.1620 | 0.4 | 209,986 | 300.0279, 315.0514 |
| 2.74 | Isorhamnetin 3-glucoside | C22H22O12 | 477.1038 | −0.2 | 7248 | 299.0205, 314.0435 |
| 2.94 | Azelaic acid | C9H16O4 | 187.0975 | −0.5 | 78,016 | 125.0972 |
| 3.07 | Unknown | C23H34O13 | 517.1930 | 0.7 | 447,712 | - |
| 3.71 | Quercetin | C15H10O7 | 301.0354 | 0.2 | 6398 | 151.0032 |
| 4.15 | Unknown | C21H36O10 | 447.2234 | 0.2 | 493,377 | - |
| 4.88 | Kaempferide | C16H12O6 | 299.0557 | −1.5 | 3863 | 284.0326 |
| 8.63 | (E,E)-9-Oxooctadeca-10,12-dienoic acid | C18H30O3 | 293.2123 | 0.2 | 367,072 | - |
| 8.57 | Unknown | C30H54 N2O19 | 745.3250 | 0.3 | 30,373 | - |
| 8.74 | (E,E)-9-Oxooctadeca-10,12-dienoic acid | C18H30O3 | 293.2121 | −0.5 | 436,590 | - |
| 9.45 | Coronaric acid | C18H32O3 | 295.2280 | 0.6 | 119,446 | - |
| 10.00 | Ricinoleic acid | C18H34O3 | 297.2434 | −0.5 | 9993 | - |
| 10.27 | Unknown | C34H40 N2O5 | 555.285 | −2.6 | 1,277,907 | - |
| 11.54 | Ursolic Acid | C30H48O3 | 455.3528 | −0.5 | 144,205 | - |
| 11.83 | Linolenic acid | C18H30O2 | 277.2174 | 0.2 | 108,147 | - |
| 11.96 | Pentadecanal | C15H30O | [M + COOH]− 271.2277 | −0.6 | 60,182 | - |
| 13.09 | n-Heptadecanal | C17H34O | [M + COOH]− 299.2591 | −0.3 | 73,029 | - |
| 13.70 | Oleic acid | C18H34O2 | 281.2486 | 0.1 | 45,678 | - |
| Target Gene | Primer Sequence | |
|---|---|---|
| iNOS | F | 5′-CAT GCT ACT GGA GGT GGG TG-3′ |
| R | 5′-CAT TGA TCT CCG TGA CAG CC-3′ | |
| COX-2 | F | 5′-TGC TGT ACA AGC AGT GGC AA-3′ |
| R | 5′-GCA GCC ATT TCC TTC TCT CC-3′ | |
| TNF-α | F | 5′-AGC ACA GAA AGC ATG ATC CG-3′ |
| R | 5′-CTG ATG AGA GGG AGG CCA TT-3′ | |
| IL-1β | F | 5′-ACCT GCT GGT GTG TGA CGT T-3′ |
| R | 5′-TCG TTG CTT GGT TCT CCT TG-3′ | |
| IL-6 | F | 5′-GAG GAT ACC ACT CCC AAC AGA CC-3′ |
| R | 5′-AAG TGC ATC ATC GTT GTT CAT ACA-3′ | |
| NLRP3 | F | 5′-GCGTGTTGTCAGGATCTCGCATTGG-3′ |
| R | 5′-GTGTCTCCAAGGGCATTGCTTCGTAG-3′ | |
| ASC | F | 5′-ACAGAAGTGGACGGAGTGCT-3′ |
| R | 5′-CTCCAGGTCCATCACCAAGT-3′ | |
| β-actin | F | 5′-ATC ACT ATT GGC AAC GAG CG-3′ |
| R | 5′-TCA GCA ATG CCT GGG TAC AT-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.-M.; Kim, S.-Y.; Mony, T.J.; Bae, H.J.; Choi, S.-H.; Choi, Y.-Y.; An, J.-Y.; Kim, H.-J.; Cho, Y.E.; Sowndhararajan, K.; et al. Moringa concanensis L. Alleviates DNCB-Induced Atopic Dermatitis-like Symptoms by Inhibiting NLRP3 Inflammasome-Mediated IL-1β in BALB/c Mice. Pharmaceuticals 2022, 15, 1217. https://doi.org/10.3390/ph15101217
Kim K-M, Kim S-Y, Mony TJ, Bae HJ, Choi S-H, Choi Y-Y, An J-Y, Kim H-J, Cho YE, Sowndhararajan K, et al. Moringa concanensis L. Alleviates DNCB-Induced Atopic Dermatitis-like Symptoms by Inhibiting NLRP3 Inflammasome-Mediated IL-1β in BALB/c Mice. Pharmaceuticals. 2022; 15(10):1217. https://doi.org/10.3390/ph15101217
Chicago/Turabian StyleKim, Kyeong-Min, So-Yeon Kim, Tamanna Jahan Mony, Ho Jung Bae, Seung-Hyuk Choi, Yu-Yeong Choi, Ju-Yeon An, Hyun-Jeong Kim, Ye Eun Cho, Kandhasamy Sowndhararajan, and et al. 2022. "Moringa concanensis L. Alleviates DNCB-Induced Atopic Dermatitis-like Symptoms by Inhibiting NLRP3 Inflammasome-Mediated IL-1β in BALB/c Mice" Pharmaceuticals 15, no. 10: 1217. https://doi.org/10.3390/ph15101217
APA StyleKim, K.-M., Kim, S.-Y., Mony, T. J., Bae, H. J., Choi, S.-H., Choi, Y.-Y., An, J.-Y., Kim, H.-J., Cho, Y. E., Sowndhararajan, K., & Park, S. J. (2022). Moringa concanensis L. Alleviates DNCB-Induced Atopic Dermatitis-like Symptoms by Inhibiting NLRP3 Inflammasome-Mediated IL-1β in BALB/c Mice. Pharmaceuticals, 15(10), 1217. https://doi.org/10.3390/ph15101217