
PCR-Based Analytical Methods for Quantification and Quality Control of Recombinant Adeno-Associated Viral Vector Preparations


Abstract
1. Introduction
2. AAV Biology
3. AAV Production
4. Methods of AAV Quantitation
4.1. Quantitative Real-Time PCR
4.1.1. Primer Selection

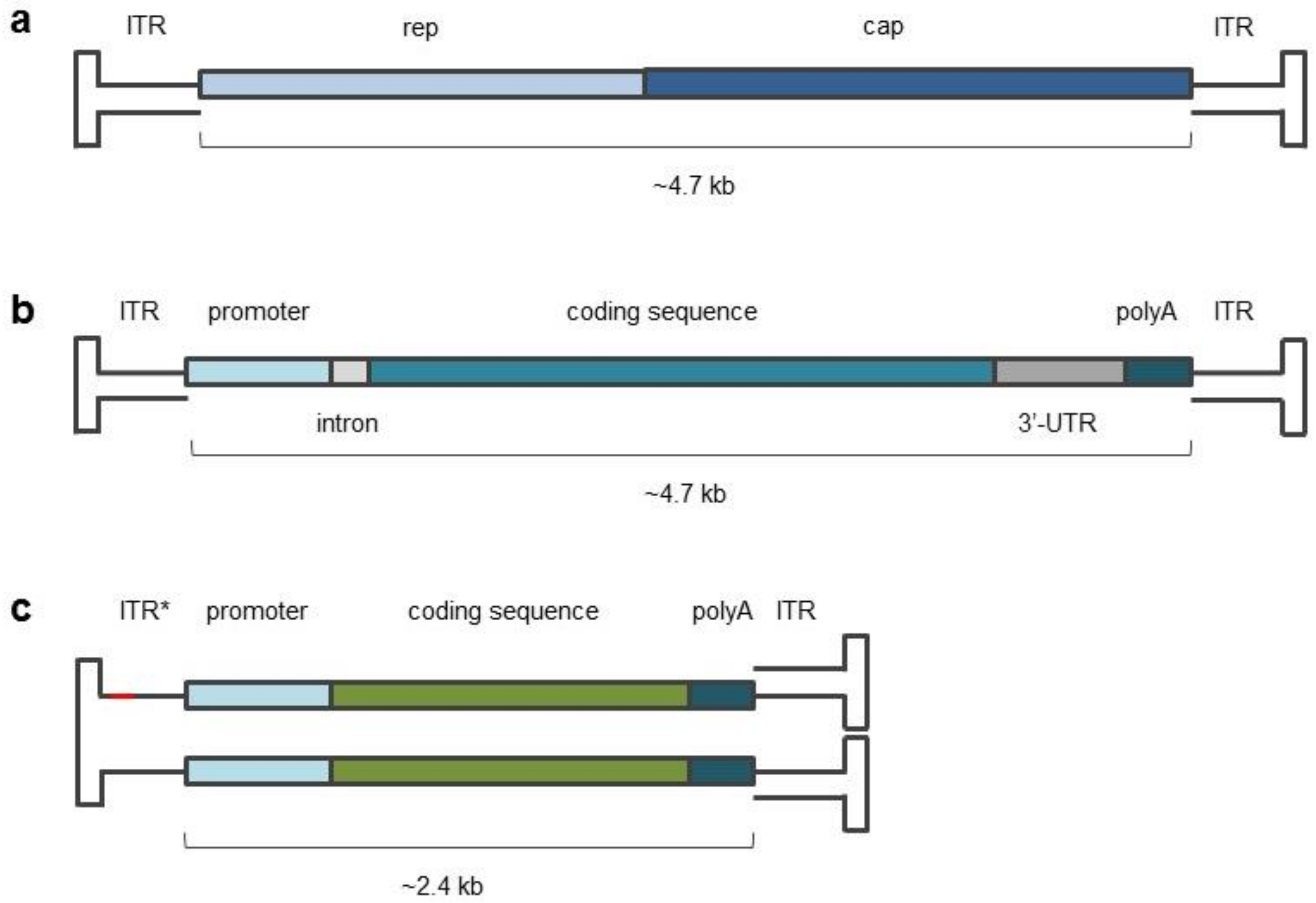{kind=link}
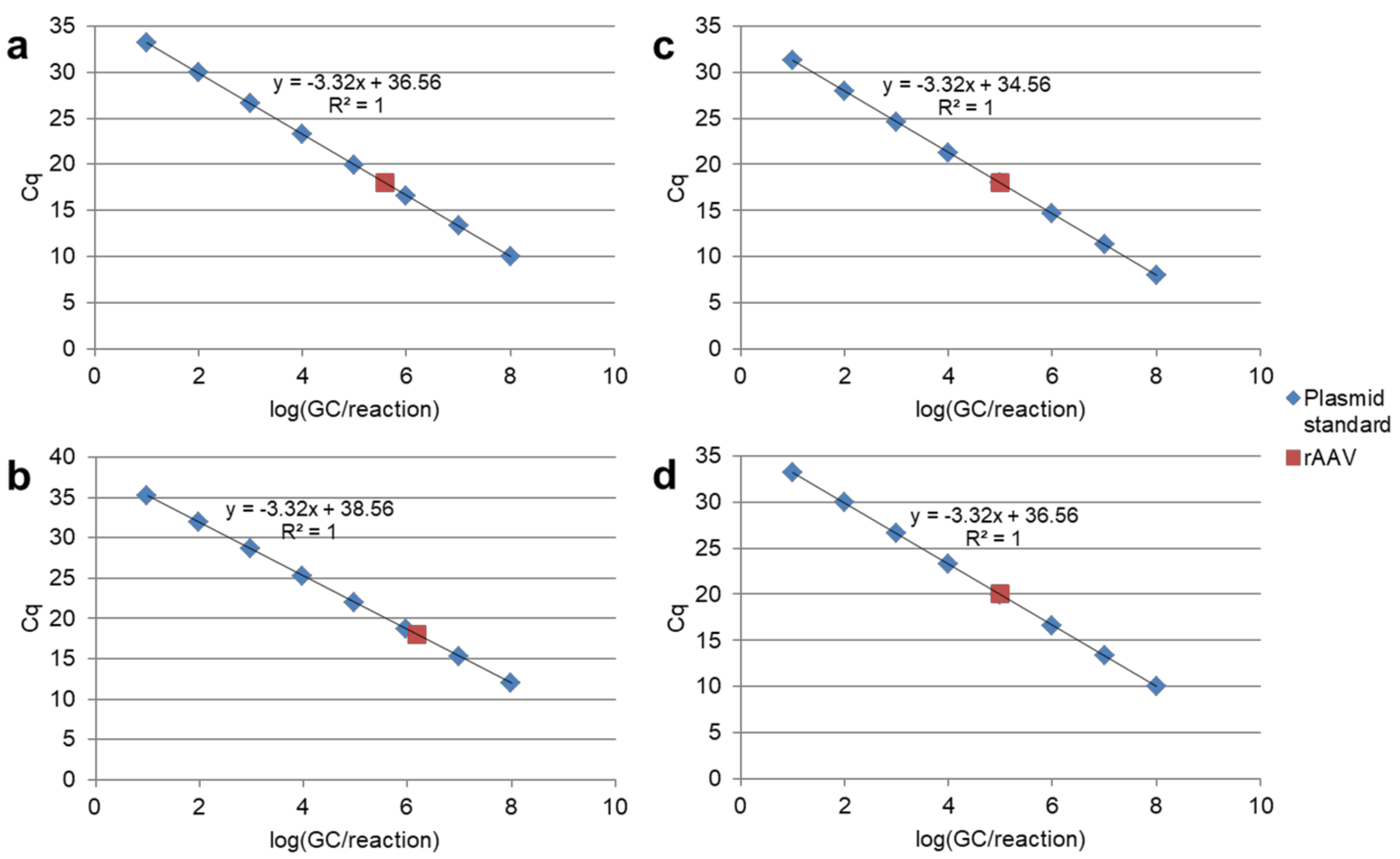{kind=link}
| Applicability | Target | Sequences | References |
|---|---|---|---|
| Universal All rAAV samples | ITR2 | primers: 5′-ggaacccctagtgatggagtt-3′ and 5′-cggcctcagtgagcga-3′; probe: R-5′-cactccctctctgcgcgctcg-3′-Q. | [44,45,46,47] |
| Semi-universal Group of rAAV samples | CMV promoter | primers: 5′-ttcctacttggcagtacatctacg-3′ and 5′-gtcaatggggtggagacttgg-3′; probe: R-5′-tgagtcaaaccgctatccacgccca-3′-Q; and other sequences. | [46,48,49] |
| CAG promoter | primers: 5′-ctgaccgcgttaatcccaca-3′ and 5′-acaagccgtgattaaaccaaga-3′. | [35] | |
| CBA promoter | primers: 5′-ccgcagccattgccttt-3′ and 5′-ccgcacagatttgggacaa-3′; probe: R-5′-atggtaatcgtgcgagagggcgc-3′-Q. | [30] | |
| TBG promoter | primers: 5′-aaactgccaattccactgctg-3′ and 5′-ccataggcaaaagcaccaaga-3′; probe: R-5′-ttggcccaatagtgagaactttttcctgc-3′-Q. | [48] | |
| GRK1 promoter | primers: 5′-tctcttaaggtagccccgg-3′ and 5′-atccgattagatcattctgccc-3′; probe: R-5′-cctcacttttcccctgagaaggaca-3′-Q. | [50] | |
| RBG intron | primers: 5′-tcaggtgcaggctgcctat-3′ and 5′-tttgtgagccagggcattg-3′; probe: R-5′-agaaggtggtggctggtgtgg-3′-Q. | [30] | |
| CMV enhancer | primers: 5′-gtcaatgggtggagtatttacgg-3′ and 5′-gcattatgcccagtacatgacct-3′; probe: R-5′-caagtgtatcatatgccaagtacgccccc-3′-Q; and other sequences. | [30,51] | |
| WPRE | primers: 5′-ttggatgctcgcctgggttg-3′ and 5′-aggaaggtccgctggatcga-3′. | [35] | |
| SV40 polyA | primers: 5′-agcaatagcatcacaaatttcacaa-3′ and 5′-ccagacatgataagatacattgatgagtt-3′; probe: R-5′-agcatttttttcactgcattctagttgtggtttgtc-3′-Q. | [45,46,47,48] | |
| BGH polyA | primers: 5′-catataaaatgaggaaattgcatcgca-3′ and 5′-tcagaacccatagagcccaccg-3′; and other sequences. | [35,48,50] | |
| RBG polyA | primers: 5′-gatttttcctcctctcctgactactc-3′ and 5′-gctgcaggtcgagggatct-3′; probe: R-5′-cagtcatagctgtccctcttctctt-3′-Q; and other sequences. | [30,48] | |
| Individual Unique rAAV samples | eGFP | primers: 5′-cacccacgtgaccacccttac-3′ and 5′-ggatgttgcagtcctccctg-3′; and other sequences. | [35,47,48,52] |
| hrGFP | primers: 5′-gatccgcagcgacatcaacc-3′ and 5′-gtacaccacctcgaagctgg-3′; probe: R-5′-gaggagatgttcgtgtaccgcgtgg-3′-Q. | [46] | |
| emGFP | primers: 5′-acggcgacgtaaacggccac-3′ and 5′-gcgaagcactgcacgccgta-3′. | [49] | |
| pU6 | primers: 5′-gggaaataggccctcttcctgccc-3′ and 5′-caccacgtgacggagcgtgac-3′. | [49] | |
| ITR5 | primers: 5′-cccccccaaacgagccag-3′ and 5′-acccccttgcttgagag-3′; probe: R-5′- cgagcgaacgcgacaggggggagagtg-3′-Q. | [47] |
4.1.2. Protocol Modifications
4.1.3. Calibration Curve
4.1.4. Reference Material
4.2. Droplet Digital PCR
4.3. Other Methods for Genome Copy Number Measurement
5. DNA Impurities
6. Complex QC Analysis of rAAV Preparations
7. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pellenz, S.; Phelps, M.; Tang, W.; Hovde, B.T.; Sinit, R.B.; Fu, W.; Li, H.; Chen, E.; Monnat, R.J. New Human Chromosomal Sites with “Safe Harbor” Potential for Targeted Transgene Insertion. Hum. Gene Ther. 2019, 30, 814–828. [Google Scholar] [CrossRef]
- Li, C.; Samulski, R.J. Engineering Adeno-Associated Virus Vectors for Gene Therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, D.A.; Shutova, M.V.; Johnston, N.R.; Smith, O.P.; Fedorin, V.V.; Kukushkin, Y.S.; van der Loo, J.C.M.; Johnstone, E.C. The Clinical Landscape for AAV Gene Therapies. Nat. Rev. Drug Discov. 2021, 20, 173–174. [Google Scholar] [CrossRef]
- Nieto, K.; Salvetti, A. AAV Vectors Vaccines Against Infectious Diseases. Front. Immunol. 2014, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Demminger, D.E.; Walz, L.; Dietert, K.; Hoffmann, H.; Planz, O.; Gruber, A.D.; von Messling, V.; Wolff, T. Adeno-Associated Virus-Vectored Influenza Vaccine Elicits Neutralizing and Fcγ Receptor-Activating Antibodies. EMBO Mol. Med. 2020, 12, e10938. [Google Scholar] [CrossRef]
- Zabaleta, N.; Dai, W.; Bhatt, U.; Hérate, C.; Maisonnasse, P.; Chichester, J.A.; Sanmiguel, J.; Estelien, R.; Michalson, K.T.; Diop, C.; et al. An AAV-Based, Room-Temperature-Stable, Single-Dose COVID-19 Vaccine Provides Durable Immunogenicity and Protection in Non-Human Primates. Cell Host Microbe 2021, 29, 1437–1453.e8. [Google Scholar] [CrossRef] [PubMed]
- Mehendale, S.; van Lunzen, J.; Clumeck, N.; Rockstroh, J.; Vets, E.; Johnson, P.R.; Anklesaria, P.; Barin, B.; Boaz, M.; Kochhar, S.; et al. A Phase 1 Study to Evaluate the Safety and Immunogenicity of a Recombinant HIV Type 1 Subtype C Adeno-Associated Virus Vaccine. AIDS Res. Hum. Retrovir. 2008, 24, 873–880. [Google Scholar] [CrossRef]
- Vardas, E.; Kaleebu, P.; Bekker, L.-G.; Hoosen, A.; Chomba, E.; Johnson, P.R.; Anklesaria, P.; Birungi, J.; Barin, B.; Boaz, M.; et al. A Phase 2 Study to Evaluate the Safety and Immunogenicity of a Recombinant HIV Type 1 Vaccine Based on Adeno-Associated Virus. AIDS Res. Hum. Retrovir. 2010, 26, 933–942. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, F.; Gao, G. CRISPR-Based Therapeutic Genome Editing: Strategies and In Vivo Delivery by AAV Vectors. Cell 2020, 181, 136–150. [Google Scholar] [CrossRef]
- Amado, D.A.; Davidson, B.L. Gene Therapy for ALS: A Review. Mol. Ther. J. Am. Soc. Gene Ther. 2021, 29, 3345–3358. [Google Scholar] [CrossRef]
- Wright, J.F. Quality Control Testing, Characterization and Critical Quality Attributes of Adeno-Associated Virus Vectors Used for Human Gene Therapy. Biotechnol. J. 2021, 16, e2000022. [Google Scholar] [CrossRef]
- Wang, S.K.; Lapan, S.W.; Hong, C.M.; Krause, T.B.; Cepko, C.L. In Situ Detection of Adeno-Associated Viral Vector Genomes with SABER-FISH. Mol. Ther. Methods Clin. Dev. 2020, 19, 376. [Google Scholar] [CrossRef] [PubMed]
- Sandza, K.; Clark, A.; Koziol, E.; Akeefe, H.; Yang, F.; Holcomb, J.; Patton, K.; Hammon, K.; Mitchell, N.; Wong, W.Y.; et al. Ultra-Sensitive AAV Capsid Detection by Immunocapture-Based QPCR Following Factor VIII Gene Transfer. Gene Ther. 2021. (online ahead of print). [Google Scholar] [CrossRef]
- Balakrishnan, B.; Jayandharan, G.R. Basic Biology of Adeno-Associated Virus (AAV) Vectors Used in Gene Therapy. Curr. Gene Ther. 2014, 14, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Bulcha, J.T.; Wang, Y.; Ma, H.; Tai, P.W.L.; Gao, G. Viral Vector Platforms within the Gene Therapy Landscape. Signal Transduct. Target. Ther. 2021, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Ogden, P.J.; Kelsic, E.D.; Sinai, S.; Church, G.M. Comprehensive AAV Capsid Fitness Landscape Reveals a Viral Gene and Enables Machine-Guided Design. Science 2019, 366, 1139–1143. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, H.; Colosi, P. Effect of Genome Size on AAV Vector Packaging. Mol. Ther. J. Am. Soc. Gene Ther. 2010, 18, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Large, E.E.; Silveria, M.A.; Zane, G.M.; Weerakoon, O.; Chapman, M.S. Adeno-Associated Virus (AAV) Gene Delivery: Dissecting Molecular Interactions upon Cell Entry. Viruses 2021, 13, 1336. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, B.P.; Bailey, C.G.; Rasko, J.E.J. Journey to the Center of the Cell: Tracing the Path of AAV Transduction. Trends Mol. Med. 2021, 27, 172–184. [Google Scholar] [CrossRef]
- McCarty, D.M.; Monahan, P.E.; Samulski, R.J. Self-Complementary Recombinant Adeno-Associated Virus (ScAAV) Vectors Promote Efficient Transduction Independently of DNA Synthesis. Gene Ther. 2001, 8, 1248–1254. [Google Scholar] [CrossRef]
- McCarty, D.M.; Fu, H.; Monahan, P.E.; Toulson, C.E.; Naik, P.; Samulski, R.J. Adeno-Associated Virus Terminal Repeat (TR) Mutant Generates Self-Complementary Vectors to Overcome the Rate-Limiting Step to Transduction in Vivo. Gene Ther. 2003, 10, 2112–2118. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, W.; Zhong, L.; Han, Z.; Li, B.; Ma, W.; Weigel-Kelley, K.A.; Warrington, K.H.; Srivastava, A. Self-Complementary Recombinant Adeno-Associated Viral Vectors: Packaging Capacity and the Role of Rep Proteins in Vector Purity. Hum. Gene Ther. 2007, 18, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Meier, A.F.; Fraefel, C.; Seyffert, M. The Interplay between Adeno-Associated Virus and Its Helper Viruses. Viruses 2020, 12, 662. [Google Scholar] [CrossRef]
- Penaud-Budloo, M.; François, A.; Clément, N.; Ayuso, E. Pharmacology of Recombinant Adeno-Associated Virus Production. Mol. Ther. Methods Clin. Dev. 2018, 8, 166–180. [Google Scholar] [CrossRef]
- Galli, A.; Iaia, I.; Milella, M.S.; Cipriani, F.; Della Latta, V.; Giacca, M.; Zentilin, L.; Cervelli, T. Characterization of Viral Genome Encapsidated in Adeno-Associated Recombinant Vectors Produced in Yeast Saccharomyces Cerevisiae. Mol. Biotechnol. 2021, 63, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Dobrowsky, T.; Gianni, D.; Pieracci, J.; Suh, J. AAV Manufacturing for Clinical Use: Insights on Current Challenges from the Upstream Process Perspective. Curr. Opin. Biomed. Eng. 2021, 20, 100353. [Google Scholar] [CrossRef]
- Rumachik, N.G.; Malaker, S.A.; Poweleit, N.; Maynard, L.H.; Adams, C.M.; Leib, R.D.; Cirolia, G.; Thomas, D.; Stamnes, S.; Holt, K.; et al. Methods Matter: Standard Production Platforms for Recombinant AAV Produce Chemically and Functionally Distinct Vectors. Mol. Ther. Methods Clin. Dev. 2020, 18, 98–118. [Google Scholar] [CrossRef]
- Flotte, T.R.; Trapnell, B.C.; Humphries, M.; Carey, B.; Calcedo, R.; Rouhani, F.; Campbell-Thompson, M.; Yachnis, A.T.; Sandhaus, R.A.; McElvaney, N.G.; et al. Phase 2 Clinical Trial of a Recombinant Adeno-Associated Viral Vector Expressing A1-Antitrypsin: Interim Results. Hum. Gene Ther. 2011, 22, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Lengler, J.; Coulibaly, S.; Gruber, B.; Ilk, R.; Mayrhofer, J.; Scheiflinger, F.; Hoellriegl, W.; Falkner, F.G.; Rottensteiner, H. Development of an In Vitro Biopotency Assay for an AAV8 Hemophilia B Gene Therapy Vector Suitable for Clinical Product Release. Mol. Ther. Methods Clin. Dev. 2020, 17, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Dobnik, D.; Kogovšek, P.; Jakomin, T.; Košir, N.; Tušek Žnidarič, M.; Leskovec, M.; Kaminsky, S.M.; Mostrom, J.; Lee, H.; Ravnikar, M. Accurate Quantification and Characterization of Adeno-Associated Viral Vectors. Front. Microbiol. 2019, 10, 1570. [Google Scholar] [CrossRef]
- Dorange, F.; Le Bec, C. Analytical Approaches to Characterize AAV Vector Production & Purification: Advances and Challenges. Cell Gene Ther. Insights 2018, 4, 119–129. [Google Scholar] [CrossRef]
- Li, Z.; Wu, Z.; Maekawa, T.; Cheung, W.; Webber, K.; Zhi, L.; O’Brian, K.; Howie, B.; Zhang, C.; Kondragunta, B.; et al. Analytical Technology Used in the Latest Development of Gene Therapy Candidates. Cell Gene Ther. Insights 2019, 5, 537–547. [Google Scholar] [CrossRef]
- Veldwijk, M.R.; Topaly, J.; Laufs, S.; Hengge, U.R.; Wenz, F.; Zeller, W.J.; Fruehauf, S. Development and Optimization of a Real-Time Quantitative PCR-Based Method for the Titration of AAV-2 Vector Stocks. Mol. Ther. J. Am. Soc. Gene Ther. 2002, 6, 272–278. [Google Scholar] [CrossRef]
- Lock, M.; McGorray, S.; Auricchio, A.; Ayuso, E.; Beecham, E.J.; Blouin-Tavel, V.; Bosch, F.; Bose, M.; Byrne, B.J.; Caton, T.; et al. Characterization of a Recombinant Adeno-Associated Virus Type 2 Reference Standard Material. Hum. Gene Ther. 2010, 21, 1273–1285. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cui, X.; Wang, M.; Xiao, W.; Xu, R. A Reliable and Feasible QPCR Strategy for Titrating AAV Vectors. Med. Sci. Monit. Basic Res. 2013, 19, 187–193. [Google Scholar] [CrossRef]
- Karlen, Y.; McNair, A.; Perseguers, S.; Mazza, C.; Mermod, N. Statistical Significance of Quantitative PCR. BMC Bioinform. 2007, 8, 131. [Google Scholar] [CrossRef]
- Clark, K.R.; Liu, X.; McGrath, J.P.; Johnson, P.R. Highly Purified Recombinant Adeno-Associated Virus Vectors Are Biologically Active and Free of Detectable Helper and Wild-Type Viruses. Hum. Gene Ther. 1999, 10, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Rohr, U.-P.; Wulf, M.-A.; Stahn, S.; Steidl, U.; Haas, R.; Kronenwett, R. Fast and Reliable Titration of Recombinant Adeno-Associated Virus Type-2 Using Quantitative Real-Time PCR. J. Virol. Methods 2002, 106, 81–88. [Google Scholar] [CrossRef]
- Mayginnes, J.P.; Reed, S.E.; Berg, H.G.; Staley, E.M.; Pintel, D.J.; Tullis, G.E. Quantitation of Encapsidated Recombinant Adeno-Associated Virus DNA in Crude Cell Lysates and Tissue Culture Medium by Quantitative, Real-Time PCR. J. Virol. Methods 2006, 137, 193–204. [Google Scholar] [CrossRef]
- Bainbridge, J.W.B.; Smith, A.J.; Barker, S.S.; Robbie, S.; Henderson, R.; Balaggan, K.; Viswanathan, A.; Holder, G.E.; Stockman, A.; Tyler, N.; et al. Effect of Gene Therapy on Visual Function in Leber’s Congenital Amaurosis. N. Engl. J. Med. 2008, 358, 2231–2239. [Google Scholar] [CrossRef]
- Maguire, A.M.; Simonelli, F.; Pierce, E.A.; Pugh, E.N.; Mingozzi, F.; Bennicelli, J.; Banfi, S.; Marshall, K.A.; Testa, F.; Surace, E.M.; et al. Safety and Efficacy of Gene Transfer for Leber’s Congenital Amaurosis. N. Engl. J. Med. 2008, 358, 2240–2248. [Google Scholar] [CrossRef] [PubMed]
- Bish, L.T.; Morine, K.; Sleeper, M.M.; Sanmiguel, J.; Wu, D.; Gao, G.; Wilson, J.M.; Sweeney, H.L. Adeno-Associated Virus (AAV) Serotype 9 Provides Global Cardiac Gene Transfer Superior to AAV1, AAV6, AAV7, and AAV8 in the Mouse and Rat. Hum. Gene Ther. 2008, 19, 1359–1368. [Google Scholar] [CrossRef]
- Rabinowitz, J.E.; Rolling, F.; Li, C.; Conrath, H.; Xiao, W.; Xiao, X.; Samulski, R.J. Cross-Packaging of a Single Adeno-Associated Virus (AAV) Type 2 Vector Genome into Multiple AAV Serotypes Enables Transduction with Broad Specificity. J. Virol. 2002, 76, 791–801. [Google Scholar] [CrossRef]
- Aurnhammer, C.; Haase, M.; Muether, N.; Hausl, M.; Rauschhuber, C.; Huber, I.; Nitschko, H.; Busch, U.; Sing, A.; Ehrhardt, A.; et al. Universal Real-Time PCR for the Detection and Quantification of Adeno-Associated Virus Serotype 2-Derived Inverted Terminal Repeat Sequences. Hum. Gene Ther. Methods 2012, 23, 18–28. [Google Scholar] [CrossRef]
- Furuta-Hanawa, B.; Yamaguchi, T.; Uchida, E. Two-Dimensional Droplet Digital PCR as a Tool for Titration and Integrity Evaluation of Recombinant Adeno-Associated Viral Vectors. Hum. Gene Ther. Methods 2019, 30, 127. [Google Scholar] [CrossRef] [PubMed]
- Werling, N.J.; Satkunanathan, S.; Thorpe, R.; Zhao, Y. Systematic Comparison and Validation of Quantitative Real-Time PCR Methods for the Quantitation of Adeno-Associated Viral Products. Hum. Gene Ther. Methods 2015, 26, 82–92. [Google Scholar] [CrossRef]
- D’Costa, S.; Blouin, V.; Broucque, F.; Penaud-Budloo, M.; François, A.; Perez, I.C.; Le Bec, C.; Moullier, P.; Snyder, R.O.; Ayuso, E. Practical Utilization of Recombinant AAV Vector Reference Standards: Focus on Vector Genomes Titration by Free ITR QPCR. Mol. Ther. Methods Clin. Dev. 2016, 5, 16019. [Google Scholar] [CrossRef]
- Lock, M.; Alvira, M.R.; Chen, S.-J.; Wilson, J.M. Absolute Determination of Single-Stranded and Self-Complementary Adeno-Associated Viral Vector Genome Titers by Droplet Digital PCR. Hum. Gene Ther. Methods 2014, 25, 115–125. [Google Scholar] [CrossRef]
- Wagner, A.; Röhrs, V.; Kedzierski, R.; Fechner, H.; Kurreck, J. A Novel Method for the Quantification of Adeno-Associated Virus Vectors for RNA Interference Applications Using Quantitative Polymerase Chain Reaction and Purified Genomic Adeno-Associated Virus DNA as a Standard. Hum. Gene Ther. Methods 2013, 24, 355–363. [Google Scholar] [CrossRef]
- Martinez-Fernandez de la Camara, C.; McClements, M.E.; MacLaren, R.E. Accurate Quantification of AAV Vector Genomes by Quantitative PCR. Genes 2021, 12, 601. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.R.H.; Cervera, L.; Ahmed, I.; Kondratov, O.; Zolotukhin, S.; Schrag, J.; Chahal, P.S.; Kamen, A.A. Achieving High-Yield Production of Functional AAV5 Gene Delivery Vectors via Fedbatch in an Insect Cell-One Baculovirus System. Mol. Ther. Methods Clin. Dev. 2019, 13, 279–289. [Google Scholar] [CrossRef]
- Wang, Y.; Menon, N.; Shen, S.; Feschenko, M.; Bergelson, S. A QPCR Method for AAV Genome Titer with DdPCR-Level of Accuracy and Precision. Mol. Ther. Methods Clin. Dev. 2020, 19, 341–346. [Google Scholar] [CrossRef]
- Fagone, P.; Wright, J.F.; Nathwani, A.C.; Nienhuis, A.W.; Davidoff, A.M.; Gray, J.T. Systemic Errors in Quantitative Polymerase Chain Reaction Titration of Self-Complementary Adeno-Associated Viral Vectors and Improved Alternative Methods. Hum. Gene Ther. Methods 2012, 23, 1–7. [Google Scholar] [CrossRef]
- Suoranta, T.; Laham-Karam, N.; Ylä-Herttuala, S. Optimized Protocol for Accurate Titration of Adeno-Associated Virus Vectors. Hum. Gene Ther. 2021, 32, 1270–1279. [Google Scholar] [CrossRef]
- Su, Q.; Sena-Esteves, M.; Gao, G. Titration of Recombinant Adeno-Associated Virus (RAAV) Genome Copy Number Using Real-Time Quantitative Polymerase Chain Reaction (QPCR). Cold Spring Harb. Protoc. 2020, 2020, 095646. [Google Scholar] [CrossRef]
- Rayaprolu, V.; Kruse, S.; Kant, R.; Venkatakrishnan, B.; Movahed, N.; Brooke, D.; Lins, B.; Bennett, A.; Potter, T.; McKenna, R.; et al. Comparative Analysis of Adeno-Associated Virus Capsid Stability and Dynamics. J. Virol. 2013, 87, 13150–13160. [Google Scholar] [CrossRef]
- Bennett, A.; Patel, S.; Mietzsch, M.; Jose, A.; Lins-Austin, B.; Yu, J.C.; Bothner, B.; McKenna, R.; Agbandje-McKenna, M. Thermal Stability as a Determinant of AAV Serotype Identity. Mol. Ther. Methods Clin. Dev. 2017, 6, 171–182. [Google Scholar] [CrossRef]
- Sanmiguel, J.; Gao, G.; Vandenberghe, L.H. Quantitative and Digital Droplet-Based AAV Genome Titration. Methods Mol. Biol. 2019, 1950, 51–83. [Google Scholar] [CrossRef]
- Ayuso, E.; Blouin, V.; Lock, M.; McGorray, S.; Leon, X.; Alvira, M.R.; Auricchio, A.; Bucher, S.; Chtarto, A.; Clark, K.R.; et al. Manufacturing and Characterization of a Recombinant Adeno-Associated Virus Type 8 Reference Standard Material. Hum. Gene Ther. 2014, 25, 977–987. [Google Scholar] [CrossRef]
- Beinhauerova, M.; Babak, V.; Bertasi, B.; Boniotti, M.B.; Kralik, P. Utilization of Digital PCR in Quantity Verification of Plasmid Standards Used in Quantitative PCR. Front. Mol. Biosci. 2020, 7, 155. [Google Scholar] [CrossRef]
- Chen, J.; Kadlubar, F.F.; Chen, J.Z. DNA Supercoiling Suppresses Real-Time PCR: A New Approach to the Quantification of Mitochondrial DNA Damage and Repair. Nucleic Acids Res. 2007, 35, 1377–1388. [Google Scholar] [CrossRef]
- Hou, Y.; Zhang, H.; Miranda, L.; Lin, S. Serious Overestimation in Quantitative PCR by Circular (Supercoiled) Plasmid Standard: Microalgal Pcna as the Model Gene. PLoS ONE 2010, 5, e9545. [Google Scholar] [CrossRef]
- Al-Zaidy, S.A.; Kolb, S.J.; Lowes, L.; Alfano, L.N.; Shell, R.; Church, K.R.; Nagendran, S.; Sproule, D.M.; Feltner, D.E.; Wells, C.; et al. AVXS-101 (Onasemnogene Abeparvovec) for SMA1: Comparative Study with a Prospective Natural History Cohort. J. Neuromuscul. Dis. 2019, 6, 307–317. [Google Scholar] [CrossRef]
- Mendell, J.R.; Shieh, P.B.; Sahenk, Z.; Lehman, K.J.; Lowes, L.P.; Reash, N.F.; Iammarino, M.; Alfano, L.N.; Woods, J.D.; Skura, C.L.; et al. A Randomized, Double-Blind, Placebo-Controlled, Genedelivery Clinical Trial of rAAVrh74.MHCK7.Microdystrophin for Duchenne Muscular Dystrophy. Available online: https://mdaconference.org/node/1140 (accessed on 24 December 2021).
- Penaud-Budloo, M.; Broucque, F.; Harrouet, K.; Bouzelha, M.; Saleun, S.; Douthe, S.; D’Costa, S.; Ogram, S.; Adjali, O.; Blouin, V.; et al. Stability of the Adeno-Associated Virus 8 Reference Standard Material. Gene Ther. 2019, 26, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Pacouret, S.; Bouzelha, M.; Shelke, R.; Andres-Mateos, E.; Xiao, R.; Maurer, A.; Mevel, M.; Turunen, H.; Barungi, T.; Penaud-Budloo, M.; et al. AAV-ID: A Rapid and Robust Assay for Batch-to-Batch Consistency Evaluation of AAV Preparations. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 1375–1386. [Google Scholar] [CrossRef]
- A Quality Control Revolution for Droplet Digital PCR. Available online: https://www.nature.com/articles/d42473-020-00123-x (accessed on 23 December 2021).
- Sidransky, D.; Tokino, T.; Hamilton, S.R.; Kinzler, K.W.; Levin, B.; Frost, P.; Vogelstein, B. Identification of Ras Oncogene Mutations in the Stool of Patients with Curable Colorectal Tumors. Science 1992, 256, 102–105. [Google Scholar] [CrossRef]
- Vogelstein, B.; Kinzler, K.W. Digital PCR. Proc. Natl. Acad. Sci. USA 1999, 96, 9236–9241. [Google Scholar] [CrossRef]
- Taylor, S.C.; Laperriere, G.; Germain, H. Droplet Digital PCR versus QPCR for Gene Expression Analysis with Low Abundant Targets: From Variable Nonsense to Publication Quality Data. Sci. Rep. 2017, 7, 2409. [Google Scholar] [CrossRef]
- Verhaegen, B.; De Reu, K.; De Zutter, L.; Verstraete, K.; Heyndrickx, M.; Van Coillie, E. Comparison of Droplet Digital PCR and QPCR for the Quantification of Shiga Toxin-Producing Escherichia Coli in Bovine Feces. Toxins 2016, 8, 157. [Google Scholar] [CrossRef]
- Xu, J.; DeVries, S.H.; Zhu, Y. Quantification of Adeno-Associated Virus with Safe Nucleic Acid Dyes. Hum. Gene Ther. 2020, 31, 1086. [Google Scholar] [CrossRef]
- Wang, Y.; Ling, C.; Song, L.; Wang, L.; Aslanidi, G.V.; Tan, M.; Ling, C.; Srivastava, A. Limitations of Encapsidation of Recombinant Self-Complementary Adeno-Associated Viral Genomes in Different Serotype Capsids and Their Quantitation. Hum. Gene Ther. Methods 2012, 23, 225. [Google Scholar] [CrossRef]
- Sommer, J.M.; Smith, P.H.; Parthasarathy, S.; Isaacs, J.; Vijay, S.; Kieran, J.; Powell, S.K.; McClelland, A.; Wright, J.F. Quantification of Adeno-Associated Virus Particles and Empty Capsids by Optical Density Measurement. Mol. Ther. J. Am. Soc. Gene Ther. 2003, 7, 122–128. [Google Scholar] [CrossRef]
- Singh, S.K. Impact of Product-Related Factors on Immunogenicity of Biotherapeutics. J. Pharm. Sci. 2011, 100, 354–387. [Google Scholar] [CrossRef]
- Wright, J.F. Product-Related Impurities in Clinical-Grade Recombinant AAV Vectors: Characterization and Risk Assessment. Biomedicines 2014, 2, 80. [Google Scholar] [CrossRef]
- Sheng-Fowler, L.; Lewis, A.M.; Peden, K. Issues Associated with Residual Cell-Substrate DNA in Viral Vaccines. Biol. J. Int. Assoc. Biol. Stand. 2009, 37, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Cellular, Tissue, and Gene Therapies Advisory Committee September 2-3, 2021 Meeting Announcement—09/02/2021–09/03/2021. Available online: https://www.fda.gov/advisory-committees/advisory-committee-calendar/cellular-tissue-and-gene-therapies-advisory-committee-september-2-3-2021-meeting-announcement (accessed on 24 December 2021).
- Vandermeulen, G.; Marie, C.; Scherman, D.; Préat, V. New Generation of Plasmid Backbones Devoid of Antibiotic Resistance Marker for Gene Therapy Trials. Mol. Ther. 2011, 19, 1942. [Google Scholar] [CrossRef]
- Kojabad, A.A.; Farzanehpour, M.; Galeh, H.E.G.; Dorostkar, R.; Jafarpour, A.; Bolandian, M.; Nodooshan, M.M. Droplet Digital PCR of Viral DNA/RNA, Current Progress, Challenges, and Future Perspectives. J. Med. Virol. 2021, 93, 4182–4197. [Google Scholar] [CrossRef]
- Soudy, A.F. Prophylactic Control Of Mycoplasma Contamination In Starting Biological Materials Used In Viral Vaccine Production. J. Appl. Vet. Sci. 2019, 4, 30–34. [Google Scholar] [CrossRef]
- Angart, P.; Kohnhorst, C.; Chiang, M.-J.; Arden, N.S. Considerations for Risk and Control of Mycoplasma in Bioprocessing. Curr. Opin. Chem. Eng. 2018, 22, 161–166. [Google Scholar] [CrossRef]
- Center for Biologics Evaluation and Research (CBER). Available online: https://www.fda.gov/about-fda/fda-organization/center-biologics-evaluation-and-research-cber (accessed on 24 December 2021).
- Tustian, A.D.; Bak, H. Assessment of Quality Attributes for Adeno-Associated Viral Vectors. Biotechnol. Bioeng. 2021, 118, 4186–4203. [Google Scholar] [CrossRef]
- Rabinowitz, J.; Chan, Y.K.; Samulski, R.J. Adeno-Associated Virus (AAV) Versus Immune Response. Viruses 2019, 11, 102. [Google Scholar] [CrossRef]
- Guerin, K.; Rego, M.; Bourges, D.; Ersing, I.; Haery, L.; DeMaio, K.H.; Sanders, E.; Tasissa, M.; Kostman, M.; Tillgren, M.; et al. A Novel Next-Generation Sequencing and Analysis Platform to Assess the Identity of Recombinant Adeno-Associated Viral Preparations from Viral DNA Extracts. Hum. Gene Ther. 2020, 31, 664. [Google Scholar] [CrossRef]
- André, M.; Reghin, S.; Boussard, E.; Lempereur, L.; Maisonneuve, S. Universal Real-Time PCR Assay for Quantitation and Size Evaluation of Residual Cell DNA in Human Viral Vaccines. Biol. J. Int. Assoc. Biol. Stand. 2016, 44, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cooper, R.; Kiladjian, A.; Bergelson, S.; Feschenko, M. A Digestion-Free Method for Quantification of Residual Host Cell DNA in RAAV Gene Therapy Products. Mol. Ther. Methods Clin. Dev. 2019, 13, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wu, M.; Menesale, E.; Lu, T.; Magliola, A.; Bergelson, S. Development and Qualification of a High Sensitivity, High Throughput Q-PCR Assay for Quantitation of Residual Host Cell DNA in Purification Process Intermediate and Drug Substance Samples. J. Pharm. Biomed. Anal. 2014, 100, 145–149. [Google Scholar] [CrossRef]
- Allay, J.A.; Sleep, S.; Long, S.; Tillman, D.M.; Clark, R.; Carney, G.; Fagone, P.; McIntosh, J.H.; Nienhuis, A.W.; Davidoff, A.M.; et al. Good Manufacturing Practice Production of Self-Complementary Serotype 8 Adeno-Associated Viral Vector for a Hemophilia B Clinical Trial. Hum. Gene Ther. 2011, 22, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Hauck, B.; Murphy, S.L.; Smith, P.H.; Qu, G.; Liu, X.; Zelenaia, O.; Mingozzi, F.; Sommer, J.M.; High, K.A.; Wright, J.F. Undetectable Transcription of Cap in a Clinical AAV Vector: Implications for Preformed Capsid in Immune Responses. Mol. Ther. J. Am. Soc. Gene Ther. 2008, 17, 144. [Google Scholar] [CrossRef] [PubMed]
- Sastry, L.; Xu, Y.; Cooper, R.; Pollok, K.; Cornetta, K. Evaluation of Plasmid DNA Removal from Lentiviral Vectors by Benzonase Treatment. Hum. Gene Ther. 2004, 15, 221–226. [Google Scholar] [CrossRef]
- Garvey, C.E.; McGowin, C.L.; Foster, T.P. Development and Evaluation of SYBR Green-I Based Quantitative PCR Assays for Herpes Simplex Virus Type 1 Whole Transcriptome Analysis. J. Virol. Methods 2014, 201, 101–111. [Google Scholar] [CrossRef]
- Ye, G.-J.; Scotti, M.M.; Liu, J.; Wang, L.; Knop, D.R.; Veres, G. Clearance and Characterization of Residual HSV DNA in Recombinant Adeno-Associated Virus Produced by an HSV Complementation System. Gene Ther. 2011, 18, 135–144. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Halbert, C.L.; Metzger, M.J.; Lam, S.-L.; Miller, A.D. Capsid-Expressing DNA in AAV Vectors and Its Elimination by Use of an Oversize Capsid Gene for Vector Production. Gene Ther. 2011, 18, 411–417. [Google Scholar] [CrossRef][Green Version]
- Bardelli, M.; Zárate-Pérez, F.; Agúndez, L.; Linden, R.M.; Escalante, C.R.; Henckaerts, E. Identification of a Functionally Relevant Adeno-Associated Virus Rep68 Oligomeric Interface. J. Virol. 2016, 90, 6612. [Google Scholar] [CrossRef]
- Funakoshi, K.; Bagheri, M.; Zhou, M.; Suzuki, R.; Abe, H.; Akashi, H. Highly Sensitive and Specific Alu-Based Quantification of Human Cells among Rodent Cells. Sci. Rep. 2017, 7, 13202. [Google Scholar] [CrossRef] [PubMed]
- Nicklas, J.A.; Buel, E. Simultaneous Determination of Total Human and Male DNA Using a Duplex Real-Time PCR Assay. J. Forensic Sci. 2006, 51, 1005–1015. [Google Scholar] [CrossRef]
- Preston Campbell, J.; Mulcrone, P.; Masood, S.K.; Karolak, M.; Merkel, A.; Hebron, K.; Zijlstra, A.; Sterling, J.; Elefteriou, F. TRIzol and Alu QPCR-Based Quantification of Metastatic Seeding within the Skeleton. Sci. Rep. 2015, 5, 12635. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.A.; Hedges, D.J.; Perodeau, B.P.; Landry, K.E.; Stoilova, N.; Laborde, M.E.; Shewale, J.; Sinha, S.K.; Batzer, M.A. Multiplex Polymerase Chain Reaction for Simultaneous Quantitation of Human Nuclear, Mitochondrial, and Male Y-Chromosome DNA: Application in Human Identification. Anal. Biochem. 2005, 337, 89–97. [Google Scholar] [CrossRef]
- McBride, C.; Gaupp, D.; Phinney, D.G. Quantifying Levels of Transplanted Murine and Human Mesenchymal Stem Cells in Vivo by Real-Time PCR. Cytotherapy 2003, 5, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Puntel, M.; Curtin, J.F.; Zirger, J.M.; Muhammad, A.K.M.; Xiong, W.; Liu, C.; Hu, J.; Kroeger, K.M.; Czer, P.; Sciascia, S.; et al. Quantification of High-Capacity Helper-Dependent Adenoviral Vector Genomes in Vitro and in Vivo, Using Quantitative TaqMan Real-Time Polymerase Chain Reaction. Hum. Gene Ther. 2006, 17, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Jiang, L.; Lei, Q.; Yang, J.; Gao, X.; Wang, W.; Zhang, Y.; Kong, T.; Chen, Q.; Li, G. Development and Validation of Quantitative Real-Time PCR for the Detection of Residual CHO Host Cell DNA and Optimization of Sample Pretreatment Method in Biopharmaceutical Products. Biol. Proced. Online 2019, 21, 1–7. [Google Scholar] [CrossRef]
- Schnödt, M.; Schmeer, M.; Kracher, B.; Krüsemann, C.; Espinosa, L.E.; Grünert, A.; Fuchsluger, T.; Rischmüller, A.; Schleef, M.; Büning, H. DNA Minicircle Technology Improves Purity of Adeno-Associated Viral Vector Preparations. Mol. Ther. Nucleic Acids 2016, 5, e355. [Google Scholar] [CrossRef]
- Lecomte, E.; Tournaire, B.; Cogné, B.; Dupont, J.-B.; Lindenbaum, P.; Martin-Fontaine, M.; Broucque, F.; Robin, C.; Hebben, M.; Merten, O.-W.; et al. Advanced Characterization of DNA Molecules in RAAV Vector Preparations by Single-Stranded Virus Next-Generation Sequencing. Mol. Ther. Nucleic Acids 2015, 4, e260. [Google Scholar] [CrossRef]
- Tai, P.W.L.; Xie, J.; Fong, K.; Seetin, M.; Heiner, C.; Su, Q.; Weiand, M.; Wilmot, D.; Zapp, M.L.; Gao, G. Adeno-Associated Virus Genome Population Sequencing Achieves Full Vector Genome Resolution and Reveals Human-Vector Chimeras. Mol. Ther. Methods Clin. Dev. 2018, 9, 130–141. [Google Scholar] [CrossRef]
- Chadeuf, G.; Ciron, C.; Moullier, P.; Salvetti, A. Evidence for Encapsidation of Prokaryotic Sequences during Recombinant Adeno-Associated Virus Production and Their in Vivo Persistence after Vector Delivery. Mol. Ther. J. Am. Soc. Gene Ther. 2005, 12, 744–753. [Google Scholar] [CrossRef]
- Penaud-Budloo, M.; Lecomte, E.; Guy-Duché, A.; Saleun, S.; Roulet, A.; Lopez-Roques, C.; Tournaire, B.; Cogné, B.; Léger, A.; Blouin, V.; et al. Accurate Identification and Quantification of DNA Species by Next-Generation Sequencing in Adeno-Associated Viral Vectors Produced in Insect Cells. Hum. Gene Ther. Methods 2017, 28, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Mao, Q.; Tai, P.W.L.; He, R.; Ai, J.; Su, Q.; Zhu, Y.; Ma, H.; Li, J.; Gong, S.; et al. Short DNA Hairpins Compromise Recombinant Adeno-Associated Virus Genome Homogeneity. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 1363–1374. [Google Scholar] [CrossRef]
- Tran, N.T.; Heiner, C.; Weber, K.; Weiand, M.; Wilmot, D.; Xie, J.; Wang, D.; Brown, A.; Manokaran, S.; Su, Q.; et al. AAV-Genome Population Sequencing of Vectors Packaging CRISPR Components Reveals Design-Influenced Heterogeneity. Mol. Ther. Methods Clin. Dev. 2020, 18, 639–651. [Google Scholar] [CrossRef]
- Tanaka, T.; Hanaoka, H.; Sakurai, S. Optimization of the Quality by Design Approach for Gene Therapy Products: A Case Study for Adeno-Associated Viral Vectors. Eur. J. Pharm. Biopharm. Off. J. Arbeitsgemeinschaft Pharm. Verfahrenstechnik EV 2020, 155, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Mallela, K.M.G.; Deorkar, N.; Brophy, G. Manufacturing Challenges and Rational Formulation Development for AAV Viral Vectors. J. Pharm. Sci. 2021, 110, 2609–2624. [Google Scholar] [CrossRef]
- Wilmott, P.; Lisowski, L.; Alexander, I.E.; Logan, G.J. A User’s Guide to the Inverted Terminal Repeats of Adeno-Associated Virus. Hum. Gene Ther. Methods 2019, 30, 206–213. [Google Scholar] [CrossRef]
- Savy, A.; Dickx, Y.; Nauwynck, L.; Bonnin, D.; Merten, O.-W.; Galibert, L. Impact of Inverted Terminal Repeat Integrity on RAAV8 Production Using the Baculovirus/Sf9 Cells System. Hum. Gene Ther. Methods 2017, 28, 277. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.; Sorrentino, M.; Woodcock, D.; O’Riordan, C.R.; Dhawan, V.; Verhagen, M.F.; Davies, C. Size Exclusion Chromatography with Dual Wavelength Detection as a Sensitive and Accurate Method for Determining the Empty and Full Capsids of Recombinant Adeno-Associated Viral Vectors. Hum. Gene Ther. 2021. (online ahead of print). [Google Scholar] [CrossRef]
- Fu, X.; Chen, W.-C.; Argento, C.; Clarner, P.; Bhatt, V.; Dickerson, R.; Bou-Assaf, G.; Bakhshayeshi, M.; Lu, X.; Bergelson, S.; et al. Analytical Strategies for Quantification of Adeno-Associated Virus Empty Capsids to Support Process Development. Hum. Gene Ther. Methods 2019, 30, 144–152. [Google Scholar] [CrossRef]
- Joshi, P.R.H.; Bernier, A.; Chahal, P.S.; Kamen, A. Development and Validation of an Anion Exchange High-Performance Liquid Chromatography Method for Analysis of Empty Capsids and Capsids Encapsidating Genetic Material in a Purified Preparation of Recombinant Adeno-Associated Virus Serotype 5. Hum. Gene Ther. 2021, 32, 1390–1402. [Google Scholar] [CrossRef] [PubMed]
- Karawdeniya, B.I.; Bandara, Y.M.N.D.Y.; Khan, A.I.; Chen, W.T.; Vu, H.-A.; Morshed, A.; Suh, J.; Dutta, P.; Kim, M.J. Adeno-Associated Virus Characterization for Cargo Discrimination through Nanopore Responsiveness. Nanoscale 2020, 12, 23721–23731. [Google Scholar] [CrossRef]
- Strasser, L.; Morgan, T.E.; Guapo, F.; Füssl, F.; Forsey, D.; Anderson, I.; Bones, J. A Native Mass Spectrometry-Based Assay for Rapid Assessment of the Empty:Full Capsid Ratio in Adeno-Associated Virus Gene Therapy Products. Anal. Chem. 2021, 93, 12817–12821. [Google Scholar] [CrossRef]
- Barnes, L.F.; Draper, B.E.; Chen, Y.-T.; Powers, T.W.; Jarrold, M.F. Quantitative Analysis of Genome Packaging in Recombinant AAV Vectors by Charge Detection Mass Spectrometry. Mol. Ther. Methods Clin. Dev. 2021, 23, 87. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.; Fernandes, D.; Hussain, M.T.; Kaszuba, M.; Stenson, J.; Markova, N. Characterization of Recombinant Adeno-Associated Viruses (RAAVs) for Gene Therapy Using Orthogonal Techniques. Pharmaceutics 2021, 13, 586. [Google Scholar] [CrossRef]
- Goodwin, M.S.; Croft, C.L.; Futch, H.S.; Ryu, D.; Ceballos-Diaz, C.; Liu, X.; Paterno, G.; Mejia, C.; Deng, D.; Menezes, K.; et al. Utilizing Minimally Purified Secreted RAAV for Rapid and Cost-Effective Manipulation of Gene Expression in the CNS. Mol. Neurodegener. 2020, 15, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Ibraheim, R.; Tai, P.W.L.; Gao, G. A Scalable and Accurate Method for Quantifying Vector Genomes of Recombinant Adeno-Associated Viruses in Crude Lysate. Hum. Gene Ther. Methods 2017, 28, 139–147. [Google Scholar] [CrossRef]
- Green, E.A.; Lee, K.H. Analytical Methods to Characterize Recombinant Adeno-Associated Virus Vectors and the Benefit of Standardization and Reference Materials. Curr. Opin. Biotechnol. 2021, 71, 65–76. [Google Scholar] [CrossRef]
- Moullier, P.; Snyder, R.O. International Efforts for Recombinant Adeno-Associated Viral Vector Reference Standards. Mol. Ther. J. Am. Soc. Gene Ther. 2008, 16, 1185–1188. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, E.; Saleun, S.; Bolteau, M.; Guy-Duché, A.; Adjali, O.; Blouin, V.; Penaud-Budloo, M.; Ayuso, E. The SSV-Seq 2.0 PCR-Free Method Improves the Sequencing of Adeno-Associated Viral Vector Genomes Containing GC-Rich Regions and Homopolymers. Biotechnol. J. 2021, 16, e2000016. [Google Scholar] [CrossRef] [PubMed]


| DNA Impurity | Target | Sequences | References |
|---|---|---|---|
| Plasmid | KanR | primers: 5′-gggcgcccggttctttttgtc-3′ and 5′-gccagtcccttcccgcttcagtg-3′. | [90] |
| AmpR | primers: 5′-cgcgccacatagcagaactt-3′ and 5′-cgccccgaagaacgttt-3′; probe: R-5′-aaaagtgctcatcattg-3′-Q; and other sequences. | [91,92] | |
| Adenovirus E2A | primers: 5′-ttgctgaaacccaccatttg-3′ and 5′-tcgtggacagcgaggaaga-3′; probe: R-5′-cgccacatcttctct-3′-Q. | [91] | |
| Adenovirus E4 | primers: 5′-tcggcgcactccgtaca-3′ and 5′-cgcgggtctctgtctcaaaa-3′; probe: R-5′-tagggatcgcctacctc-3′-Q. | [91] | |
| HSV UL23 | primers: 5′-tcgatgtgtctgtcctccg-3′ and 5′-atcccatcgccgccctc-3′. | [93] | |
| HSV UL24 | primers: 5′-gccgcgagaacgcgcag-3′ and 5′-cctcgaataccgagcgacc-3′. | [93] | |
| HSV UL26.5 | primers: 5′-catgtccttccacccagac-3′ and 5′-cccatcatctgagagacgaa-3′; probe: R-5′-cagcacacgtggacgttgacac-3′-Q. | [94] | |
| HSV UL29 | primers: 5′-ccgcctatggttaccttgtc-3′ and 5′-ccctcctgtatctggtcgtt-3′; probe: R-5′-agcctcccaggtgcagaaaggt-3′-Q. | [94] | |
| HSV UL33 | primers: 5′- cgaactttacgggacacaatc-3′ and 5′-cgtagtcgggaagacaacct-3′; probe: R-5′-tagacgcgcgctacgtctcg-3′-Q. | [94] | |
| HSV UL35 | primers: 5′-acgcaaacaacacgtttacc-3′ and 5′-tcgaaggttctcgaacgac-3′; probe: R-5′-cggcgcacctattcaccgttt-3′-Q. | [94] | |
| Ori | primers: 5′-gcgcgtaatctgctgcttg-3′ and 5′-ctacggctacactagaagaacagta-3′; probe: R-5′-cgctctgctgaagccagttaccttcgg-3′-Q. | [45] | |
| cap8 | primers: 5′-tcagccaaggtgggcctaatacaa-3′ and 5′-ttgctgctgcaagttatctgccac-3′; and other sequences. | [90,95] | |
| rep2 | primers: 5′-cggggttttacgagattgtg-3′ and 5′-cgccatttctggtctttgtg-3′. | [95] | |
| cap2 | primers: 5′-ttctcagatgctgcgtaccggaaa-3′ and 5′-tctgccattgaggtggtacttggt-3′. | [96] | |
| Host cell DNA | human Alu | primers: 5′-gaggcgggcggatca-3′ and 5′-cccggctaatttttgtatttttagtag-3′; probe: R-5′-cagcctggccaacatggtgaaacc-3′-Q; and other sequences. | [88,89,97,98,99,100,101] |
| Adenovirus E1A | primers: 5′-gggtgaggagtttgtgttagattatg-3′ and 5′-tcctccggtgataatgacaaga-3′; probe: R-5′-agcaccccgggcacggttg-3′-Q. | [102] | |
| chinese hamster Alu | primers: 5′-agagatggctcgaggttaag-3′ and 5′-tctgcacaccagaagagg-3′; probe: R-5′-agcaccaactgctcttccagagg-3′-Q. | [103] | |
| Syrian hamster 5S rRNA | primers: 5′-cgcagcagcaggctct-3′ and 5′-accctgcttagcttccgaga-3′; probe: R-5′-ccgccgtcgtctacggccatacc-3′-Q. | [94] | |
| rcAAV | ITR-Rep recombinants | primers: 5′-actccatcactaggggttct-3′ and 5′-gctggggaccttaatcacaa-3′. | [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shmidt, A.A.; Egorova, T.V. PCR-Based Analytical Methods for Quantification and Quality Control of Recombinant Adeno-Associated Viral Vector Preparations. Pharmaceuticals 2022, 15, 23. https://doi.org/10.3390/ph15010023
Shmidt AA, Egorova TV. PCR-Based Analytical Methods for Quantification and Quality Control of Recombinant Adeno-Associated Viral Vector Preparations. Pharmaceuticals. 2022; 15(1):23. https://doi.org/10.3390/ph15010023
Chicago/Turabian StyleShmidt, Anna A., and Tatiana V. Egorova. 2022. "PCR-Based Analytical Methods for Quantification and Quality Control of Recombinant Adeno-Associated Viral Vector Preparations" Pharmaceuticals 15, no. 1: 23. https://doi.org/10.3390/ph15010023
APA StyleShmidt, A. A., & Egorova, T. V. (2022). PCR-Based Analytical Methods for Quantification and Quality Control of Recombinant Adeno-Associated Viral Vector Preparations. Pharmaceuticals, 15(1), 23. https://doi.org/10.3390/ph15010023