
Effect of Neuropeptide S Administration on Ultrasonic Vocalizations and Behaviour in Rats with Low vs. High Exploratory Activity


Abstract
1. Introduction
2. Results
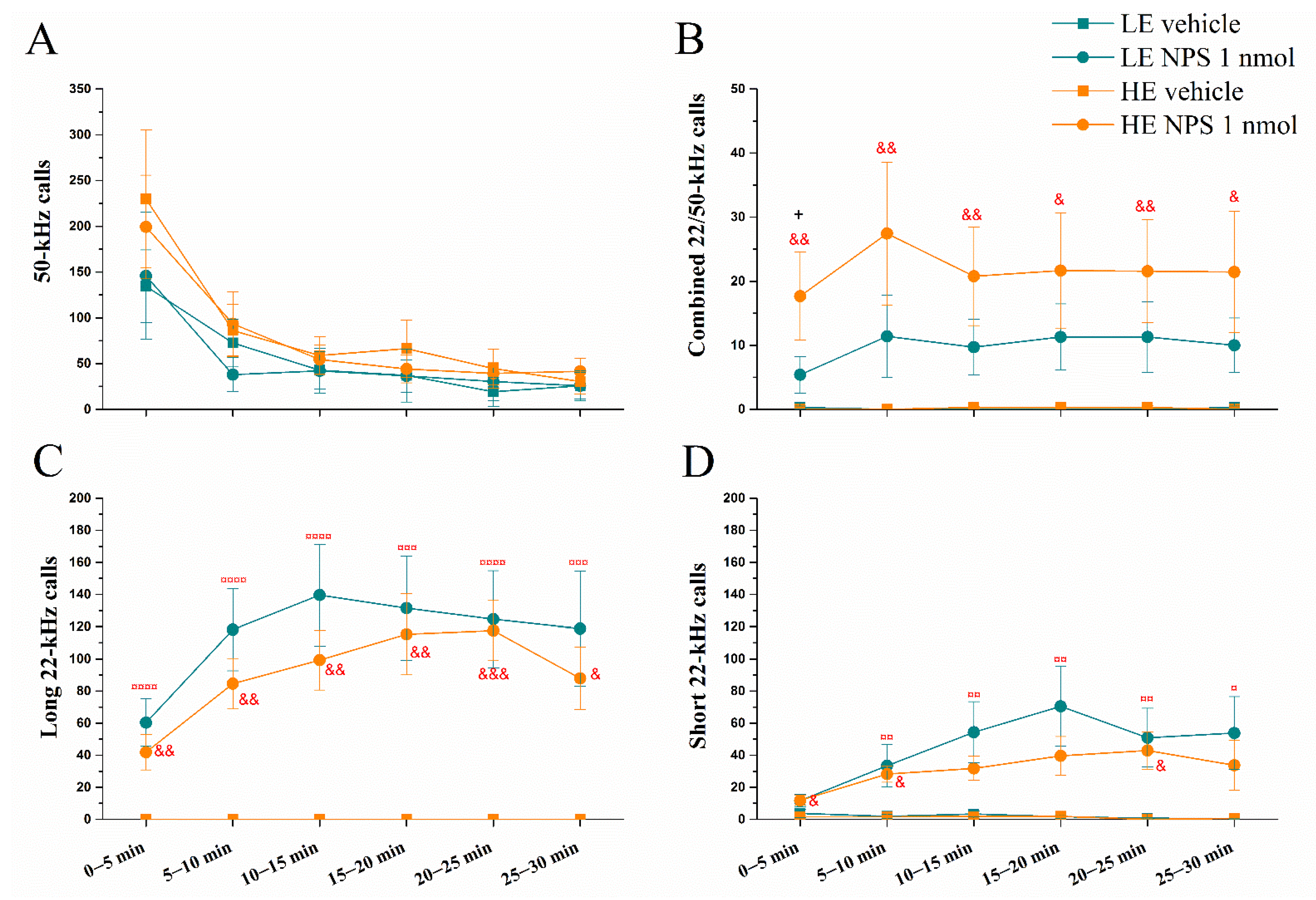
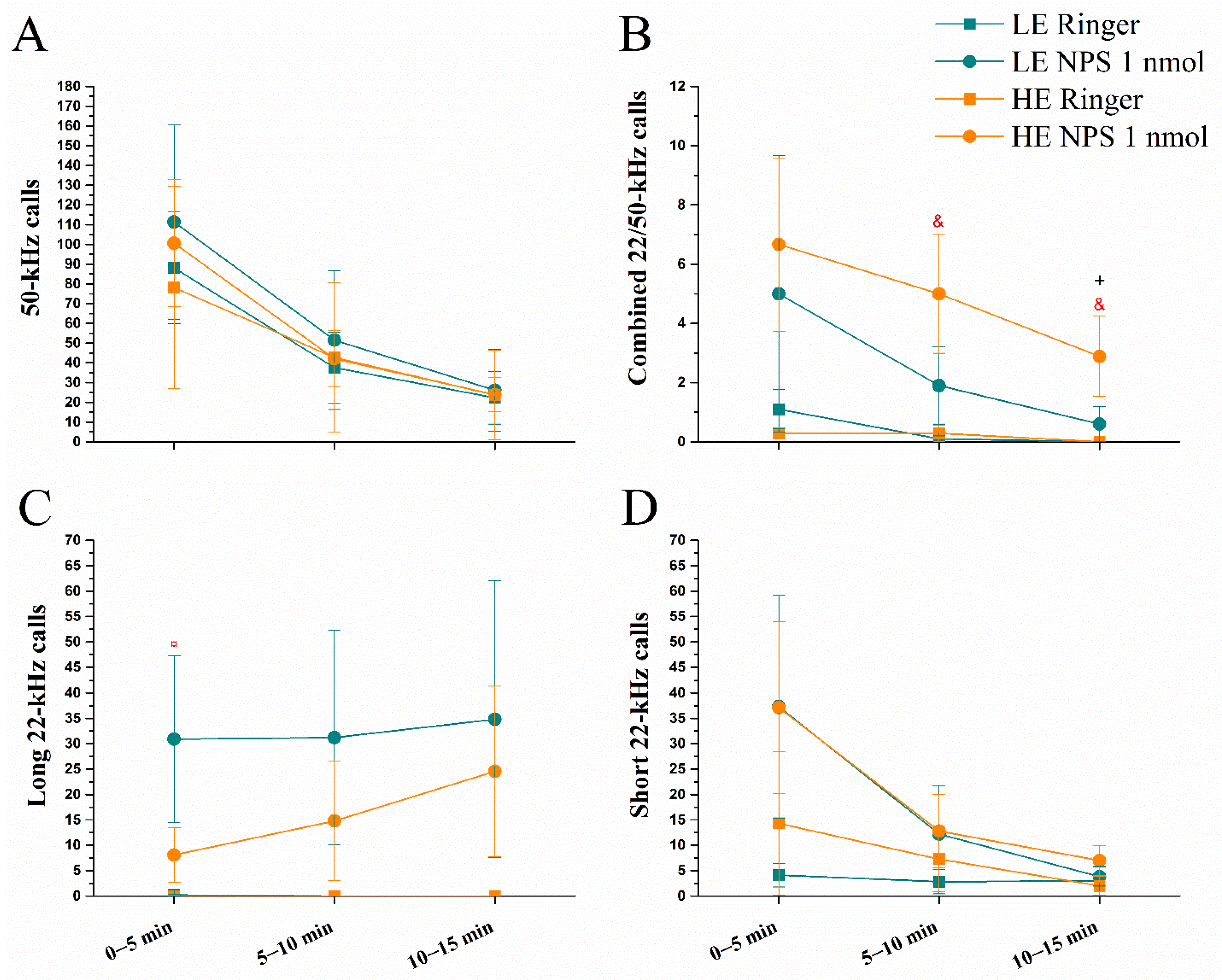
2.1. Effect of Intracerebroventricular Administration of NPS on Ultrasonic Vocalizations
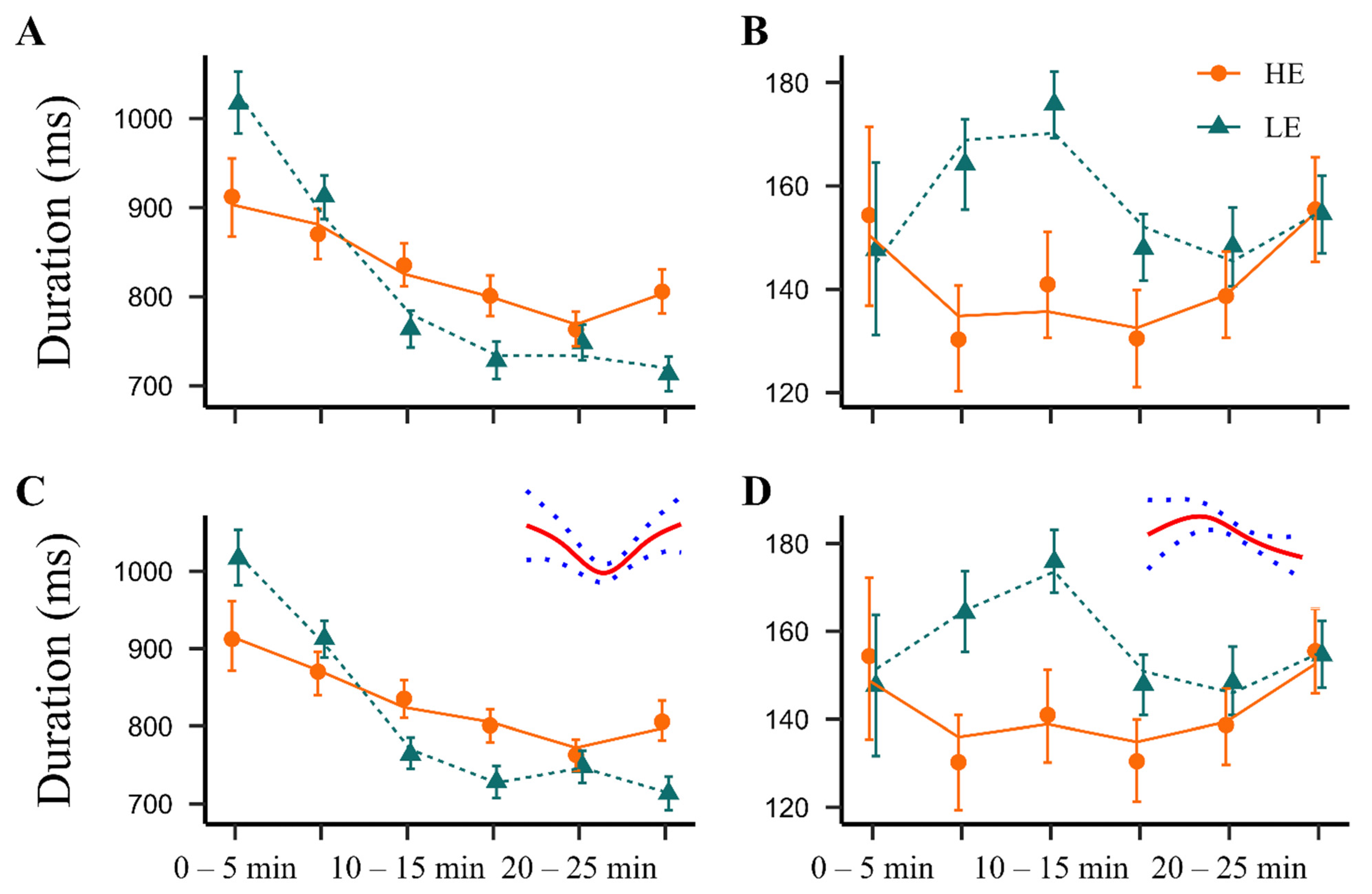
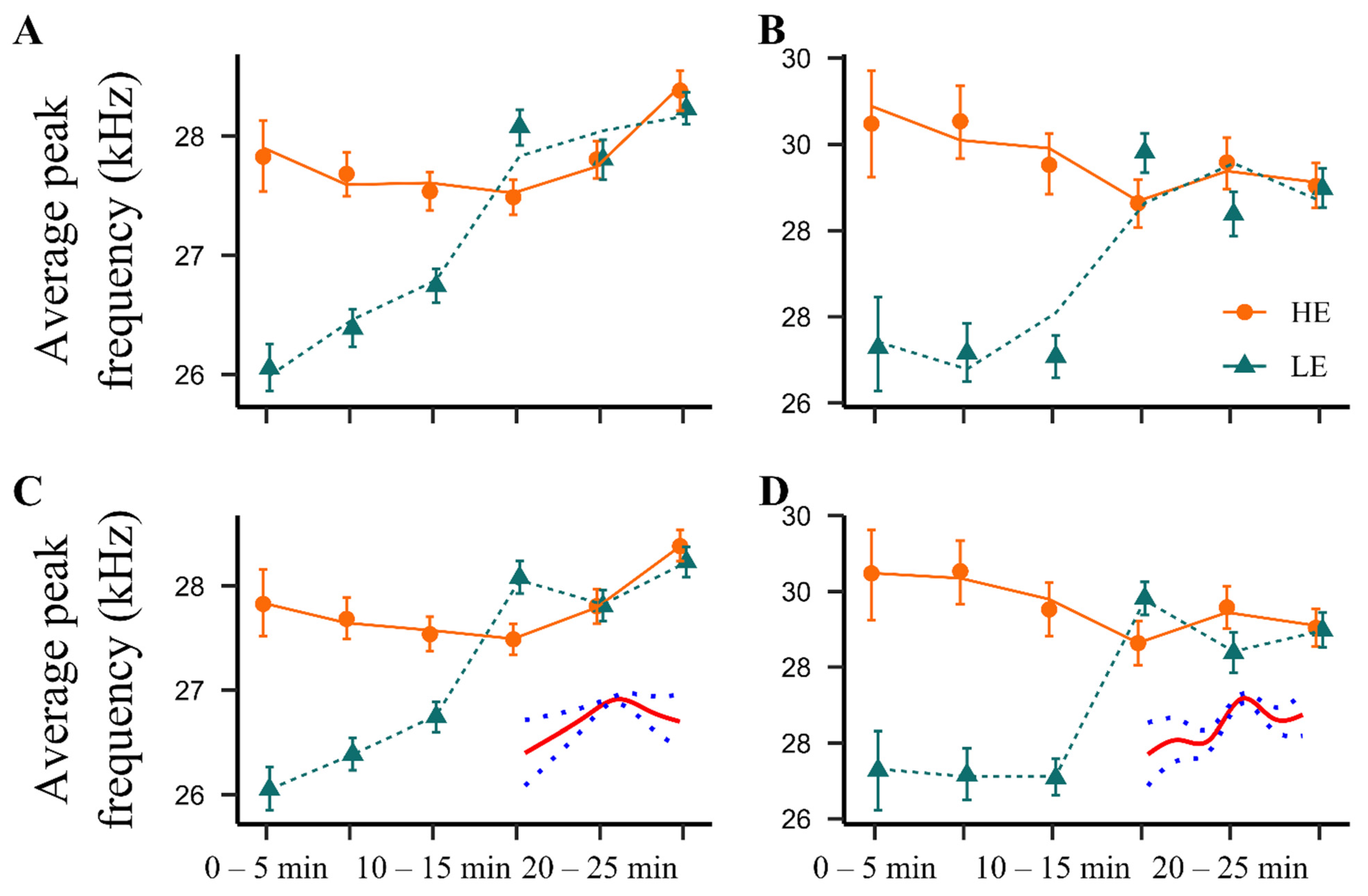
2.2. Acoustic Characteristics of the NPS-Elicited 22 kHz Calls in HE- and LE-Rats
2.2.1. Duration of Calls
2.2.2. Average Peak Frequency
2.2.3. Average Peak Amplitude
2.3. Behavioural Tests
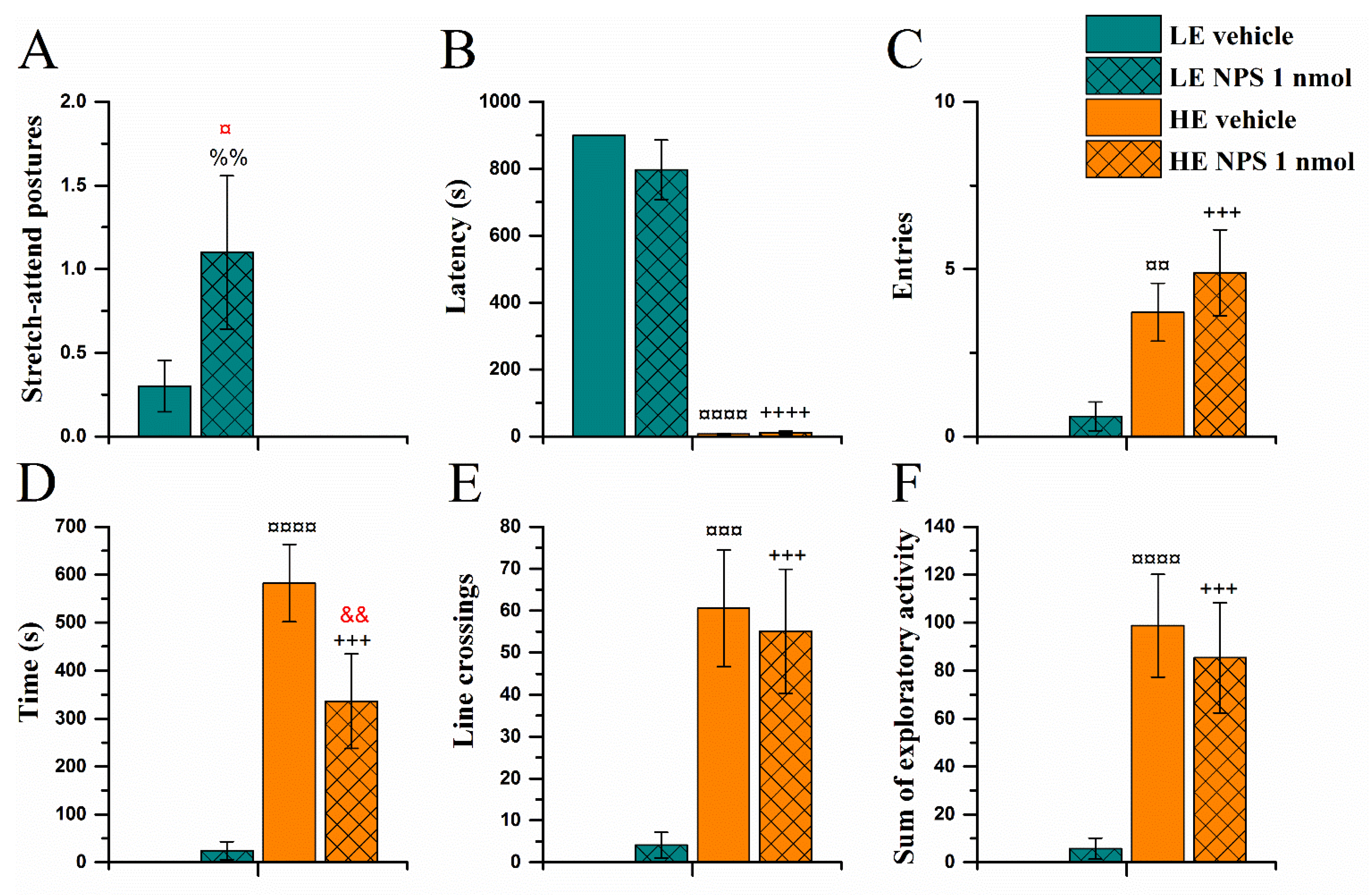
2.3.1. Exploration Box Test
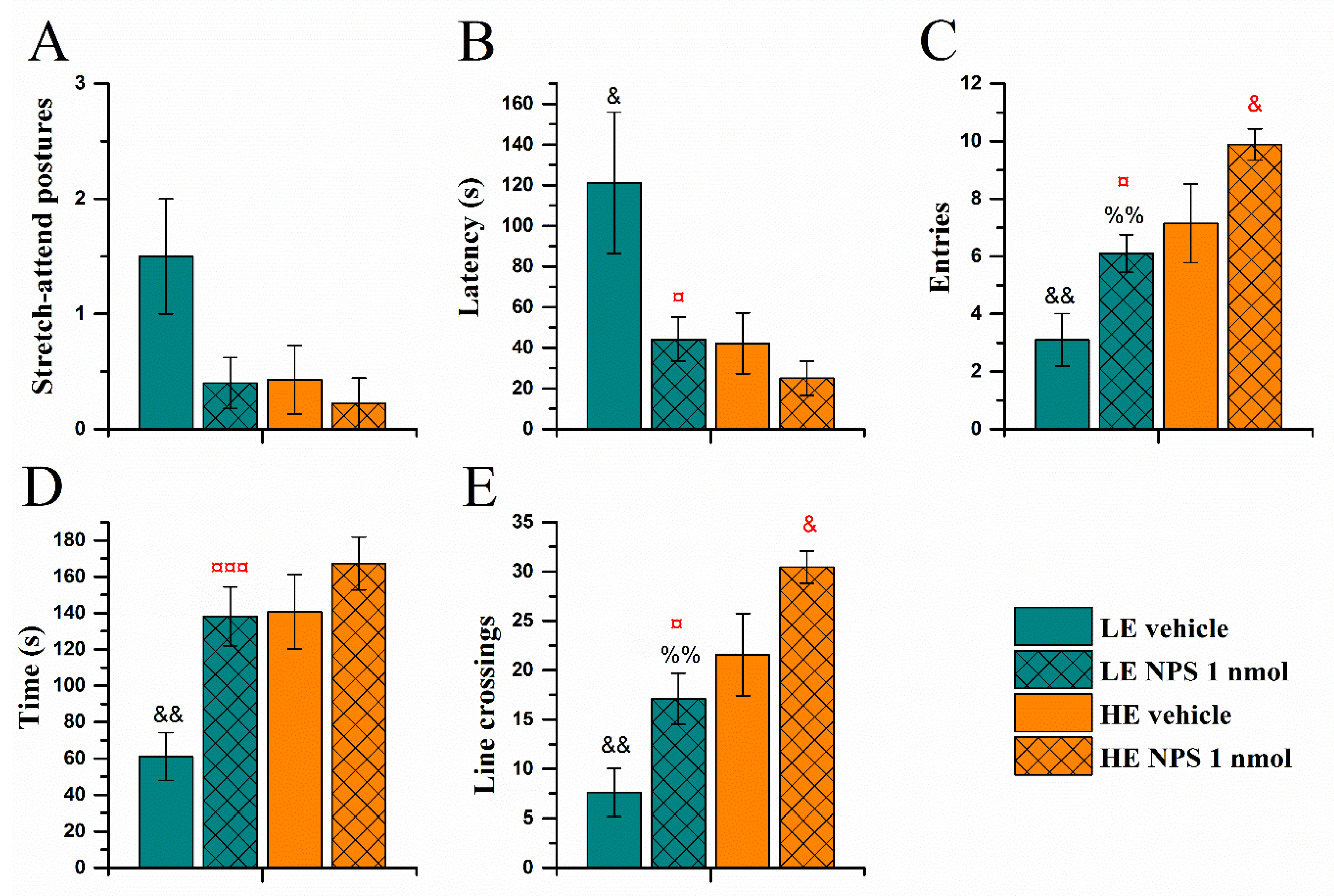
2.3.2. Elevated Zero-Maze Test
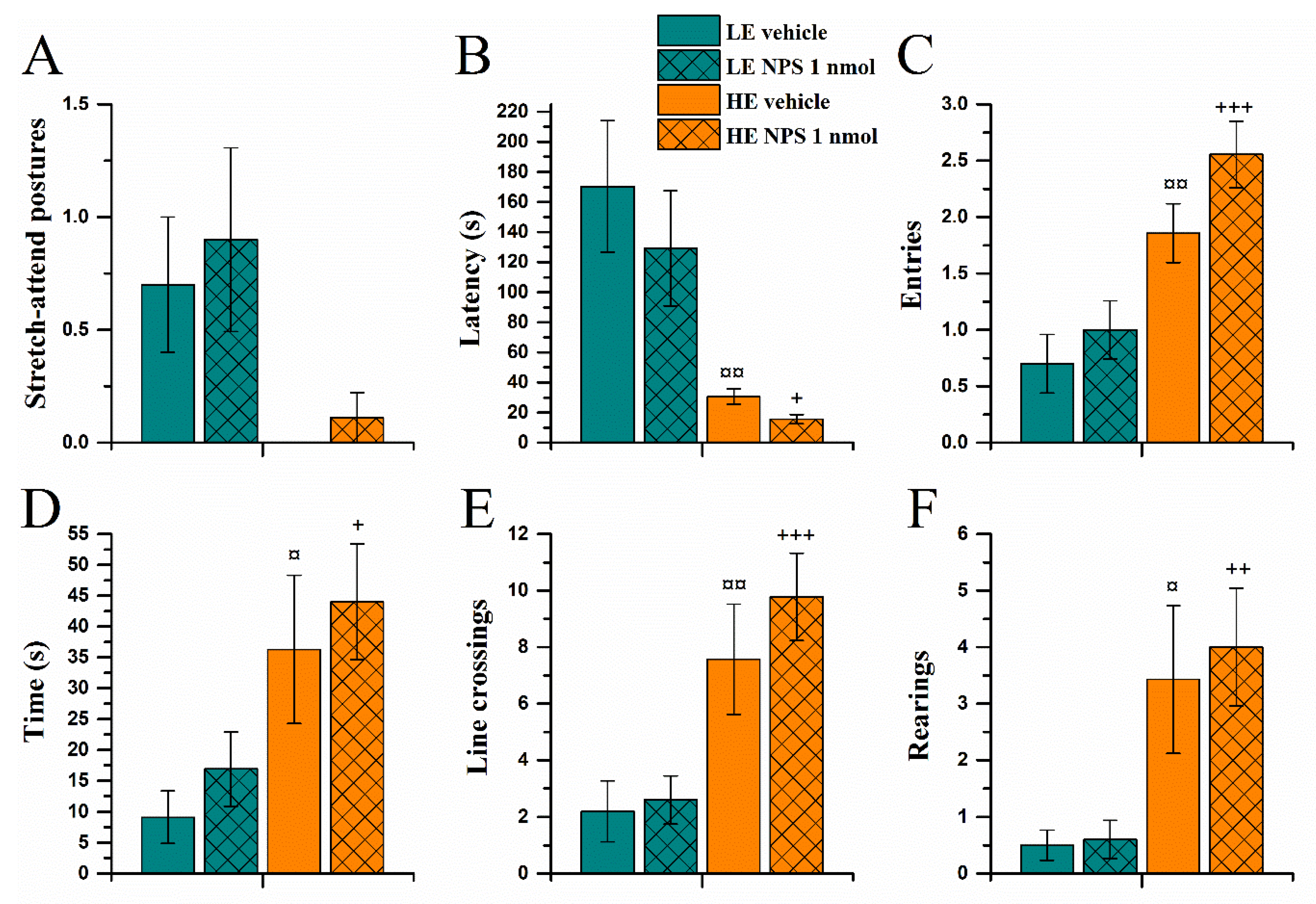
2.3.3. Light-Dark Box Test
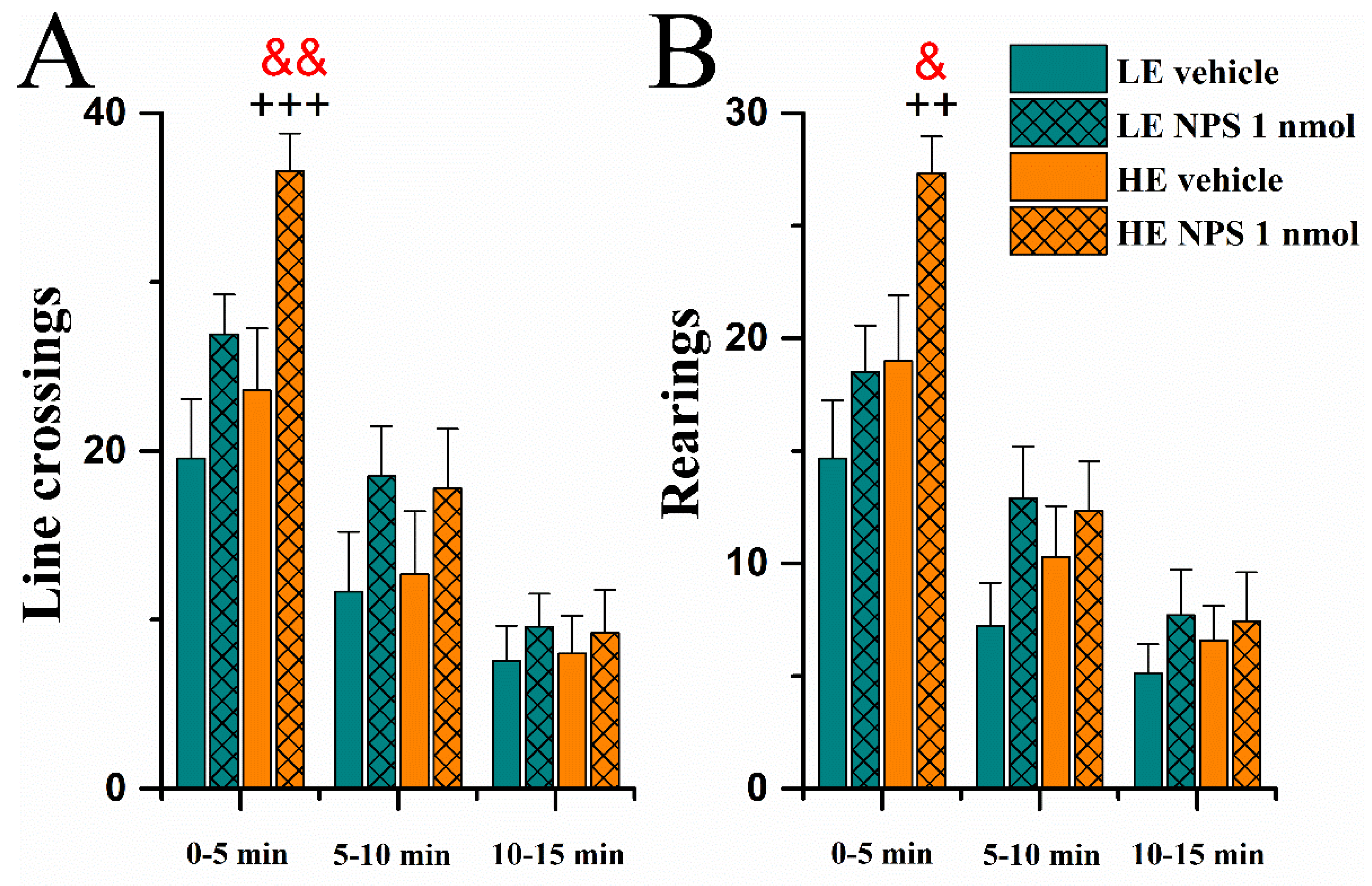
2.3.4. Novel Large Rat Housing Cage
Locomotor Activity
Number of USVs
Correlation of Locomotor Activity and USVs
3. Discussion
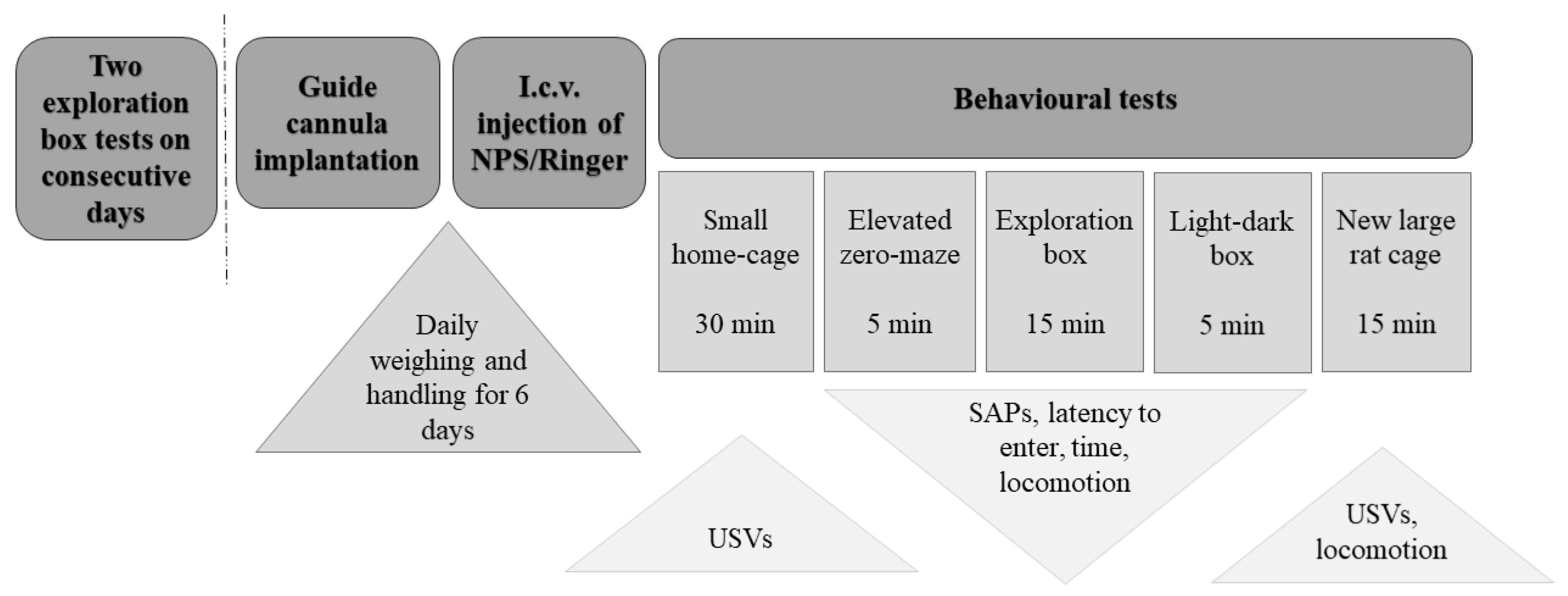
4. Materials and Methods
4.1. Animals
4.2. General Procedure
4.3. Drugs
4.4. Stereotaxic Surgery
4.5. I.c.v. Procedure
4.6. Behavioural Experiments
4.6.1. Exploration Box Test
4.6.2. Elevated Zero-Maze Test
4.6.3. Light-Dark Box Test
4.6.4. Locomotor Activity in a Novel Large Rat Housing Cage
4.7. Recording and Scoring of USVs
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, Y.-L.L.; Reinscheid, R.K.; Huitron-Resendiz, S.; Clark, S.D.; Wang, Z.; Lin, S.H.; Brucher, F.A.; Zeng, J.; Ly, N.K.; Henriksen, S.J.; et al. Neuropeptide S: A neuropeptide promoting arousal and anxiolytic-like effects. Neuron 2004, 43, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-L.; Gall, C.M.; Jackson, V.R.; Civelli, O.; Reinscheid, R.K. Distribution of neuropeptide S receptor mRNA and neurochemical characteristics of neuropeptide S-expressing neurons in the rat brain. J. Comp. Neurol. 2007, 500, 84–102. [Google Scholar] [CrossRef]
- Rizzi, A.; Vergura, R.; Marzola, G.; Ruzza, C.; Guerrini, R.; Salvadori, S.; Regoli, D.; Calo, G. Neuropeptide S is a stimulatory anxiolytic agent: A behavioural study in mice. Br. J. Pharmacol. 2008, 154, 471–479. [Google Scholar] [CrossRef]
- Pulga, A.; Ruzza, C.; Rizzi, A.; Guerrini, R.; Calo, G. Anxiolytic- and panicolytic-like effects of Neuropeptide S in the mouse elevated T-maze. Eur. J. Neurosci. 2012, 36, 3531–3537. [Google Scholar] [CrossRef]
- Ensho, T.; Nakahara, K.; Suzuki, Y.; Murakami, N. Neuropeptide S increases motor activity and thermogenesis in the rat through sympathetic activation. Neuropeptides 2017, 65, 21–27. [Google Scholar] [CrossRef]
- Vitale, G.; Filaferro, M.; Ruggieri, V.; Pennella, S.; Frigeri, C.; Rizzi, A.; Guerrini, R.; Calò, G. Anxiolytic-like effect of neuropeptide S in the rat defensive burying. Peptides 2008, 29, 2286–2291. [Google Scholar] [CrossRef]
- Leonard, S.K.; Dwyer, J.M.; Sukoff Rizzo, S.J.; Platt, B.; Logue, S.F.; Neal, S.J.; Malberg, J.E.; Beyer, C.E.; Schechter, L.E.; Rosenzweig-Lipson, S.; et al. Pharmacology of neuropeptide S in mice: Therapeutic relevance to anxiety disorders. Psychopharmacology 2008, 197, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Reinscheid, R.K.; Xu, Y.L.; Okamura, N.; Zeng, J.; Chung, S.; Pai, R.; Wang, Z.; Civelli, O. Pharmacological characterization of human and murine neuropeptide S receptor variants. J. Pharmacol. Exp. Ther. 2005, 315, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Domschke, K.; Reif, A.; Weber, H.; Richter, J.; Hohoff, C.; Ohrmann, P.; Pedersen, A.; Bauer, J.; Suslow, T.; Kugel, H.; et al. Neuropeptide S receptor gene—Converging evidence for a role in panic disorder. Mol. Psychiatry 2011, 16, 938–948. [Google Scholar] [CrossRef]
- Dannlowski, U.; Kugel, H.; Franke, F.; Stuhrmann, A.; Hohoff, C.; Zwanzger, P.; Lenzen, T.; Grotegerd, D.; Suslow, T.; Arolt, V.; et al. Neuropeptide-S (NPS) receptor genotype modulates basolateral amygdala responsiveness to aversive stimuli. Neuropsychopharmacology 2011, 36, 1879–1885. [Google Scholar] [CrossRef]
- Laas, K.; Reif, A.; Akkermann, K.; Kiive, E.; Domschke, K.; Lesch, K.P.; Veidebaum, T.; Harro, J. Interaction of the neuropeptide S receptor gene Asn107Ile variant and environment: Contribution to affective and anxiety disorders, and suicidal behaviour. Int. J. Neuropsychopharmacol. 2014, 17, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Laas, K.; Eensoo, D.; Paaver, M.; Lesch, K.P.; Reif, A.; Harro, J. Further evidence for the association of the NPSR1 gene A/T polymorphism (Asn107Ile) with impulsivity and hyperactivity. J. Psychopharmacol. 2015, 29, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, J.; Zhu, C.; Yang, L.; Ren, Y.; Ruan, J.; Fan, G.; Hu, J.; Xu, W.; Bi, X.; et al. African lungfish genome sheds light on the vertebrate water-to-land transition. Cell 2021, 184, 1362–1376.e18. [Google Scholar] [CrossRef]
- Reinscheid, R.K.; Mafessoni, F.; Lüttjohann, A.; Jüngling, K.; Pape, H.C.; Schulz, S. Neandertal introgression and accumulation of hypomorphic mutations in the neuropeptide S (NPS) system promote attenuated functionality. Peptides 2021, 138, 170506. [Google Scholar] [CrossRef] [PubMed]
- Ebner, K.; Rjabokon, A.; Pape, H.C.; Singewald, N. Increased in vivo release of neuropeptide S in the amygdala of freely moving rats after local depolarisation and emotional stress. Amino Acids 2011, 41, 991–996. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Smith, K.L.; Patterson, M.; Dhillo, W.S.; Patel, S.R.; Semjonous, N.M.; Gardiner, J.V.; Ghatei, M.A.; Bloom, S.R. Neuropeptide S stimulates the hypothalamo-pituitary-adrenal axis and inhibits food intake. Endocrinology 2006, 147, 3510–3518. [Google Scholar] [CrossRef]
- Jüngling, K.; Liu, X.; Lesting, J.; Coulon, P.; Sosulina, L.; Reinscheid, R.K.; Pape, H.C. Activation of neuropeptide S-expressing neurons in the locus coeruleus by corticotropin-releasing factor. J. Physiol. 2012, 590, 3701–3717. [Google Scholar] [CrossRef] [PubMed]
- Pañeda, C.; Huitron-Resendiz, S.; Frago, L.M.; Chowen, J.A.; Picetti, R.; De Lecea, L.; Roberts, A.J. Neuropeptide S reinstates cocaine-seeking behavior and increases locomotor activity through corticotropin-releasing factor receptor 1 in mice. J. Neurosci. 2009, 29, 4155–4161. [Google Scholar] [CrossRef][Green Version]
- Mochizuki, T.; Kim, J.; Sasaki, K. Microinjection of neuropeptide S into the rat ventral tegmental area induces hyperactivity and increases extracellular levels of dopamine metabolites in the nucleus accumbens shell. Peptides 2010, 31, 926–931. [Google Scholar] [CrossRef]
- Si, W.; Aluisio, L.; Okamura, N.; Clark, S.D.; Fraser, I.; Sutton, S.W.; Bonaventure, P.; Reinscheid, R.K. Neuropeptide S stimulates dopaminergic neurotransmission in the medial prefrontal cortex. J. Neurochem. 2010, 115, 475–482. [Google Scholar] [CrossRef]
- Harro, J. Animals, anxiety, and anxiety disorders: How to measure anxiety in rodents and why. Behav. Brain Res. 2018, 352, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J. Affective Neuroscience: The Foundations of Human and Animal Emotions, 1st ed.; Oxford University Press: New York, NY, USA, 1998; ISBN 0195096738. [Google Scholar]
- Brudzynski, S.M. Ethotransmission: Communication of emotional states through ultrasonic vocalization in rats. Curr. Opin. Neurobiol. 2013, 23, 310–317. [Google Scholar] [CrossRef]
- Simola, N.; Granon, S. Ultrasonic vocalizations as a tool in studying emotional states in rodent models of social behavior and brain disease. Neuropharmacology 2019, 159, 107420. [Google Scholar] [CrossRef]
- Burgdorf, J.; Panksepp, J.; Moskal, J.R. Frequency-modulated 50 kHz ultrasonic vocalizations: A tool for uncovering the molecular substrates of positive affect. Neurosci. Biobehav. Rev. 2011, 35, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- Kroes, R.A.; Burgdorf, J.; Otto, N.J.; Panksepp, J.; Moskal, J.R. Social defeat, a paradigm of depression in rats that elicits 22-kHz vocalizations, preferentially activates the cholinergic signaling pathway in the periaqueductal gray. Behav. Brain Res. 2007, 182, 290–300. [Google Scholar] [CrossRef]
- Burgdorf, J.; Knutson, B.; Panksepp, J.; Shippenberg, T.S. Evaluation of rat ultrasonic vocalizations as predictors of the conditioned aversive effects of drugs. Psychopharmacology 2001, 155, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, R.J.; Weiss, S. Twenty-two kHz alarm cries to presentation of a predator, by laboratory rats living in visible burrow systems. Physiol. Behav. 1991, 50, 1991. [Google Scholar] [CrossRef]
- Sánchez, C. Stress-induced vocalisation in adult animals. A valid model of anxiety? Eur. J. Pharmacol. 2003, 463, 133–143. [Google Scholar] [CrossRef]
- Burgdorf, J.; Kroes, R.A.; Moskal, J.R.; Pfaus, J.G.; Brudzynski, S.M.; Panksepp, J. Ultrasonic vocalizations of rats (Rattus norvegicus) during mating, play, and aggression: Behavioral concomitants, relationship to reward, and self-administration of playback. J. Comp. Psychol. 2008, 122, 357–367. [Google Scholar] [CrossRef]
- Panksepp, J.; Burgdorf, J. 50-kHz chirping (laughter?) in response to conditioned and unconditioned tickle-induced reward in rats: Effects of social housing and genetic variables. Behav. Brain Res. 2000, 115, 25–38. [Google Scholar] [CrossRef]
- Burgdorf, J.; Wood, P.L.; Kroes, R.A.; Moskal, J.R.; Panksepp, J. Neurobiology of 50-kHz ultrasonic vocalizations in rats: Electrode mapping, lesion, and pharmacology studies. Behav. Brain Res. 2007, 182, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, S.M.; Bihari, F.; Ociepa, D.; Fu, X.W. Analysis of 22 kHz ultrasonic vocalization in laboratory rats: Long and short calls. Physiol. Behav. 1993, 54, 215–221. [Google Scholar] [CrossRef]
- Brudzynski, S.M. Pharmacology of ultrasonic vocalizations in adult rats: Significance, call classification and neural substrate. Curr. Neuropharmacol. 2015, 13, 180–192. [Google Scholar] [CrossRef]
- Brudzynski, S.M.; Iku, A.; Harness neé Savoy, A. Activity of cholinergic neurons in the laterodorsal tegmental nucleus during emission of 22kHz vocalization in rats. Behav. Brain Res. 2011, 225, 276–283. [Google Scholar] [CrossRef]
- Burgdorf, J.; Knutson, B.; Panksepp, J.; Ikemoto, S. Nucleus accumbens amphetamine microinjections unconditionally elicit 50-kHz ultrasonic vocalizations in rats. Behav. Neurosci. 2001, 115, 940–944. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, S.M. Pharmacological and behavioral characteristics of 22 kHz alarm calls in rats. Neurosci. Biobehav. Rev. 2001, 25, 611–617. [Google Scholar] [CrossRef]
- Ahrens, A.M.; Ma, S.T.; Maier, E.Y.; Duvauchelle, C.L.; Schallert, T. Repeated intravenous amphetamine exposure: Rapid and persistent sensitization of 50-kHz ultrasonic trill calls in rats. Behav. Brain Res. 2009, 197, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Simola, N.; Fenu, S.; Costa, G.; Pinna, A.; Plumitallo, A.; Morelli, M. Pharmacological characterization of 50-kHz ultrasonic vocalizations in rats: Comparison of the effects of different psychoactive drugs and relevance in drug-induced reward. Neuropharmacology 2012, 63, 224–234. [Google Scholar] [CrossRef]
- Harro, J. Inter-individual differences in neurobiology as vulnerability factors for affective disorders: Implications for psychopharmacology. Pharmacol. Ther. 2010, 125, 402–422. [Google Scholar] [CrossRef]
- Gould, T.D.; Gottesman, I.I. Psychiatric endophenotypes and the development of valid animal models. Genes Brain Behav. 2006, 5, 113–119. [Google Scholar] [CrossRef]
- Montgomery, K.C. The relation between fear induced by novel stimulation and exploratory drive. J. Comp. Physiol. Psychol. 1955, 48, 254–260. [Google Scholar] [CrossRef]
- Otter, M.H.; Matto, V.; Sõukand, R.; Skrebuhhova, T.; Allikmets, L.; Harro, J. Characterization of rat exploratory behavior using the exploration box test. Methods Find. Exp. Clin. Pharmacol. 1997, 19, 683–691. [Google Scholar]
- Mällo, T.; Alttoa, A.; Kõiv, K.; Tõnissaar, M.; Eller, M.; Harro, J. Rats with persistently low or high exploratory activity: Behaviour in tests of anxiety and depression, and extracellular levels of dopamine. Behav. Brain Res. 2007, 177, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Alttoa, A.; Kõiv, K.; Hinsley, T.A.; Brass, A.; Harro, J. Differential gene expression in a rat model of depression based on persistent differences in exploratory activity. Eur. Neuropsychopharmacol. 2010, 20, 288–300. [Google Scholar] [CrossRef]
- Alttoa, A.; Kõiv, K.; Eller, M.; Uustare, A.; Rinken, A.; Harro, J. Effects of low dose N-(2-chloroethyl)-N-ethyl-2-bromobenzylamine administration on exploratory and amphetamine-induced behavior and dopamine D2 receptor function in rats with high or low exploratory activity. Neuroscience 2005, 132, 979–990. [Google Scholar] [CrossRef]
- Alttoa, A.; Seeman, P.; Kõiv, K.; Eller, M.; Harro, J. Rats with persistently high exploratory activity have both higher extracellular dopamine levels and higher proportion of D2 High receptors in the striatum. Synapse 2009, 63, 443–446. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, A.; Kõiv, K.; Raudkivi, K.; Harro, J. Antidepressants differentially affect striatal amphetamine-stimulated dopamine and serotonin release in rats with high and low novelty-oriented behaviour. Pharmacol. Res. 2016, 113, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, S.M. Principles of rat communication: Quantitative parameters of ultrasonic calls in rats. Behav. Genet. 2005, 35, 85–92. [Google Scholar] [CrossRef]
- Basken, J.N.; Connor, N.P.; Ciucci, M.R. Effect of aging on ultrasonic vocalizations and laryngeal sensorimotor neurons in rats. Exp. Brain Res. 2012, 219, 351–361. [Google Scholar] [CrossRef]
- Zhu, H.; Mingler, M.K.; McBride, M.L.; Murphy, A.J.; Valenzuela, D.M.; Yancopoulos, G.D.; Williams, M.T.; Vorhees, C.V.; Rothenberg, M.E. Abnormal response to stress and impaired NPS-induced hyperlocomotion, anxiolytic effect and corticosterone increase in mice lacking NPSR1. Psychoneuroendocrinology 2010, 35, 1119–1132. [Google Scholar] [CrossRef]
- Simola, N.; Paci, E.; Serra, M.; Costa, G.; Morelli, M. Modulation of Rat 50-kHz Ultrasonic Vocalizations by Glucocorticoid Signaling: Possible Relevance to Reward and Motivation. Int. J. Neuropsychopharmacol. 2018, 21, 73–83. [Google Scholar] [CrossRef]
- Ise, S.; Nagano, N.; Okuda, S.; Ohta, H. Corticotropin-releasing factor modulates maternal separation-induced ultrasonic vocalization in rat pups via activation of CRF1 receptor. Brain Res. 2008, 1234, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; McCloskey, N.; Phillips, J.; Simmons, S.J.; Kirby, L.G. CRF-5-HT interactions in the dorsal raphe nucleus and motivation for stress-induced opioid reinstatement. Psychopharmacology 2020, 238, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Swiergiel, A.H.; Zhou, Y.; Dunn, A.J. Effects of chronic footshock, restraint and corticotropin-releasing factor on freezing, ultrasonic vocalization and forced swim behavior in rats. Behav. Brain Res. 2007, 183, 178–187. [Google Scholar] [CrossRef]
- Brudzynski, S.M.; Ociepa, D.; Bihari, F. Comparison between cholinergically and naturally induced ultrasonic vocalization in the rat. J. Psychiatry Neurosci. 1991, 16, 221–226. [Google Scholar]
- Brudzynski, S.M. Ultrasonic vocalization induced by intracerebral carbachol in rats: Localization and a dose-response study. Behav. Brain Res. 1994, 63, 133–143. [Google Scholar] [CrossRef]
- Reinscheid, R.K. Neuropeptide S: Anatomy, pharmacology, genetics and physiological functions. Results Probl. Cell Differ. 2008, 46, 145–158. [Google Scholar] [CrossRef]
- Cao, J.; De Lecea, L.; Ikemoto, S. Intraventricular administration of neuropeptide S has reward-like effects. Eur. J. Pharmacol. 2011, 658, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Matrov, D.; Kõiv, K.; Kanarik, M.; Peet, K.; Raudkivi, K.; Harro, J. Middle-range exploratory activity in adult rats suggests higher resilience to chronic social defeat. Acta Neuropsychiatr. 2016, 28, 125–140. [Google Scholar] [CrossRef]
- Wegener, G.; Finger, B.C.; Elfving, B.; Keller, K.; Liebenberg, N.; Fischer, C.W.; Singewald, N.; Slattery, D.A.; Neumann, I.D.; Mathé, A.A. Neuropeptide S alters anxiety, but not depression-like behaviour in Flinders Sensitive Line rats: A genetic animal model of depression. Int. J. Neuropsychopharmacol. 2012, 15, 375–387. [Google Scholar] [CrossRef]
- Slattery, D.A.; Naik, R.R.; Grund, T.; Yen, Y.C.; Sartori, S.B.; Füchsl, A.; Finger, B.C.; Elfving, B.; Nordemann, U.; Guerrini, R.; et al. Selective breeding for high anxiety introduces a synonymous SNP that increases Neuropeptide S receptor activity. J. Neurosci. 2015, 35, 4599–4613. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: London, UK, 2007; ISBN 9780080475158. [Google Scholar]
- Shepherd, J.K.; Grewal, S.S.; Fletcher, A.; Bill, D.J.; Dourish, C.T. Behavioural and pharmacological characterisation of the elevated “zero-maze” as an animal model of anxiety. Psychopharmacology 1994, 116, 56–64. [Google Scholar] [CrossRef]
- Matto, V.; Harro, J.; Allikmets, L. The effects of cholecystokinin A and B receptor antagonists on exploratory behaviour in the elevated zero-maze in rat. Neuropharmacology 1997, 36, 389–396. [Google Scholar] [CrossRef]
- Henniger, M.S.H.; Ohl, F.; Hölter, S.M.; Weißenbacher, P.; Toschi, N.; Lörscher, P.; Wigger, A.; Spanagel, R.; Landgraf, R. Unconditioned anxiety and social behaviour in two rat lines selectively bred for high and low anxiety-related behaviour. Behav. Brain Res. 2000, 111, 153–163. [Google Scholar] [CrossRef]
- Kõiv, K.; Tiitsaar, K.; Laugus, K.; Harro, J. Extracellular Dopamine Levels in Nucleus Accumbens after Chronic Stress in Rats with Persistently High vs. Low 50-kHz Ultrasonic Vocalization Response. Brain Sci. 2021, 11, 470. [Google Scholar] [CrossRef] [PubMed]
- Mirman, D. Growth Curve Analysis and Visualization Using R, 1st ed.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2014; ISBN 9781466584327. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2017; ISBN 9781498728348. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measure | LE | HE |
|---|---|---|
| USVs | ||
| Home-cage | ↑ short and long 22 kHz USVs | ↑ short and long 22 kHz USVs |
| ↑ 22/50 kHz USVs | ↑ 22/50 kHz USVs | |
| --- 50 kHz USVs | --- 50 kHz USVs | |
| ↓ average peak frequency in long 22 kHz USVs * | ||
| ↑ average peak amplitude in long 22 kHz USVs * | ||
| Novel large rat cage | ↑ long 22 kHz USVs | ↑ long 22 kHz USVs |
| ↑ 22/50 kHz USVs | ↑ 22/50 kHz USVs | |
| --- 50 kHz, short 22 kHz USVs | --- 50 kHz, short 22 kHz USVs | |
| Behavioural tests | ||
| Exploration box | ↑ stretch-attend postures | ↓ time in the open area while no change in exploratory activity |
| Elevated 0-maze | ↓ latency to enter | |
| ↑ time in the open quadrants | ||
| ↑ entries | ↑ entries | |
| ↑ line crossings | ↑ line crossings | |
| Light-dark box | --- | --- |
| Novel large rat cage | --- | ↑ rearings and line crossings during first 5 min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kõiv, K.; Matrov, D.; Uusen, T.; Harro, J. Effect of Neuropeptide S Administration on Ultrasonic Vocalizations and Behaviour in Rats with Low vs. High Exploratory Activity. Pharmaceuticals 2021, 14, 524. https://doi.org/10.3390/ph14060524
Kõiv K, Matrov D, Uusen T, Harro J. Effect of Neuropeptide S Administration on Ultrasonic Vocalizations and Behaviour in Rats with Low vs. High Exploratory Activity. Pharmaceuticals. 2021; 14(6):524. https://doi.org/10.3390/ph14060524
Chicago/Turabian StyleKõiv, Kadri, Denis Matrov, Trine Uusen, and Jaanus Harro. 2021. "Effect of Neuropeptide S Administration on Ultrasonic Vocalizations and Behaviour in Rats with Low vs. High Exploratory Activity" Pharmaceuticals 14, no. 6: 524. https://doi.org/10.3390/ph14060524
APA StyleKõiv, K., Matrov, D., Uusen, T., & Harro, J. (2021). Effect of Neuropeptide S Administration on Ultrasonic Vocalizations and Behaviour in Rats with Low vs. High Exploratory Activity. Pharmaceuticals, 14(6), 524. https://doi.org/10.3390/ph14060524