
Protective Effect of Carotenoid Extract from Orange-Fleshed Sweet Potato on Gastric Ulcer in Mice by Inhibition of NO, IL-6 and PGE2 Production

 , , ,
, , ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
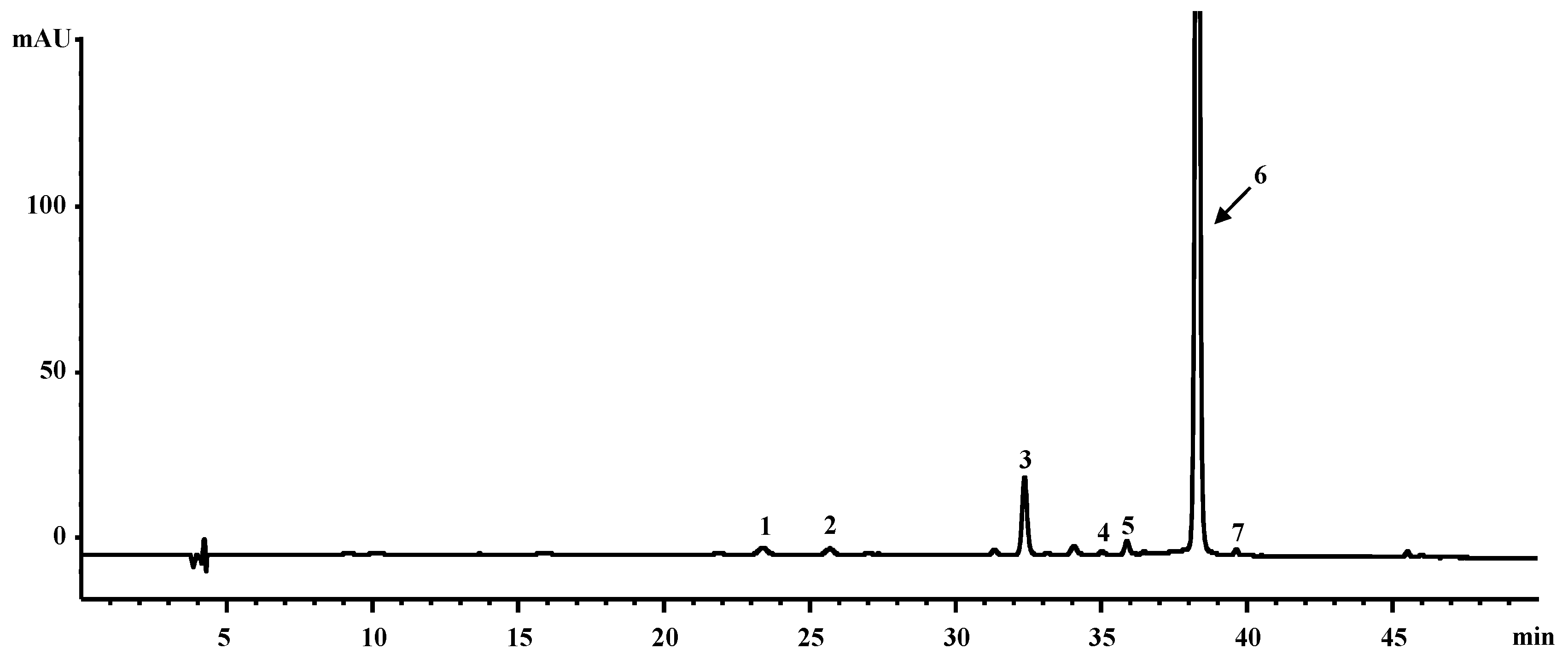
2.1. Chemical Profiles of CE from Orange-Fleshed Sweet Potato
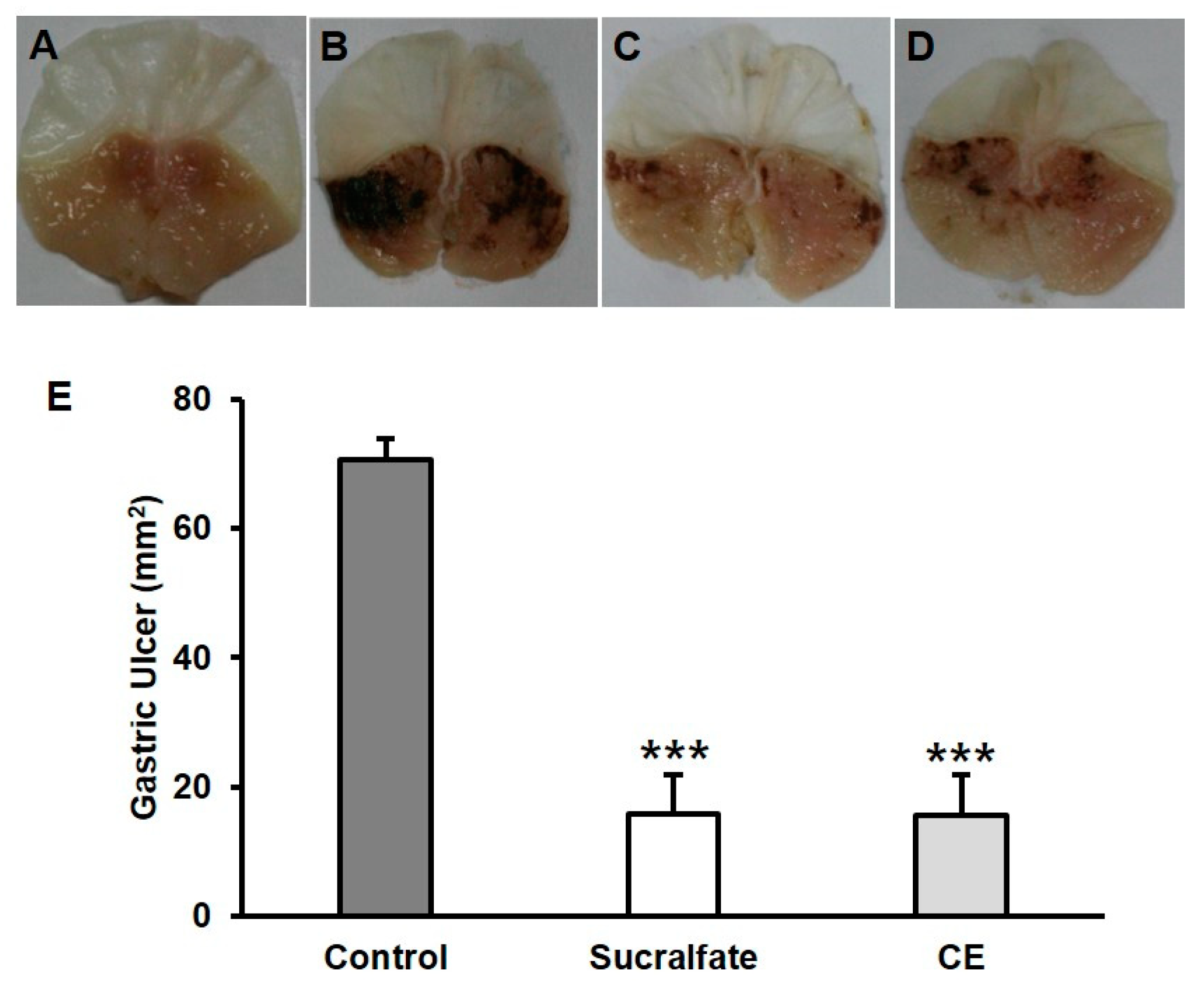
2.2. Protective Effects of Carotenoid Extract on HCl/Ethanol-Induced Gastric Ulcer Model in Mice
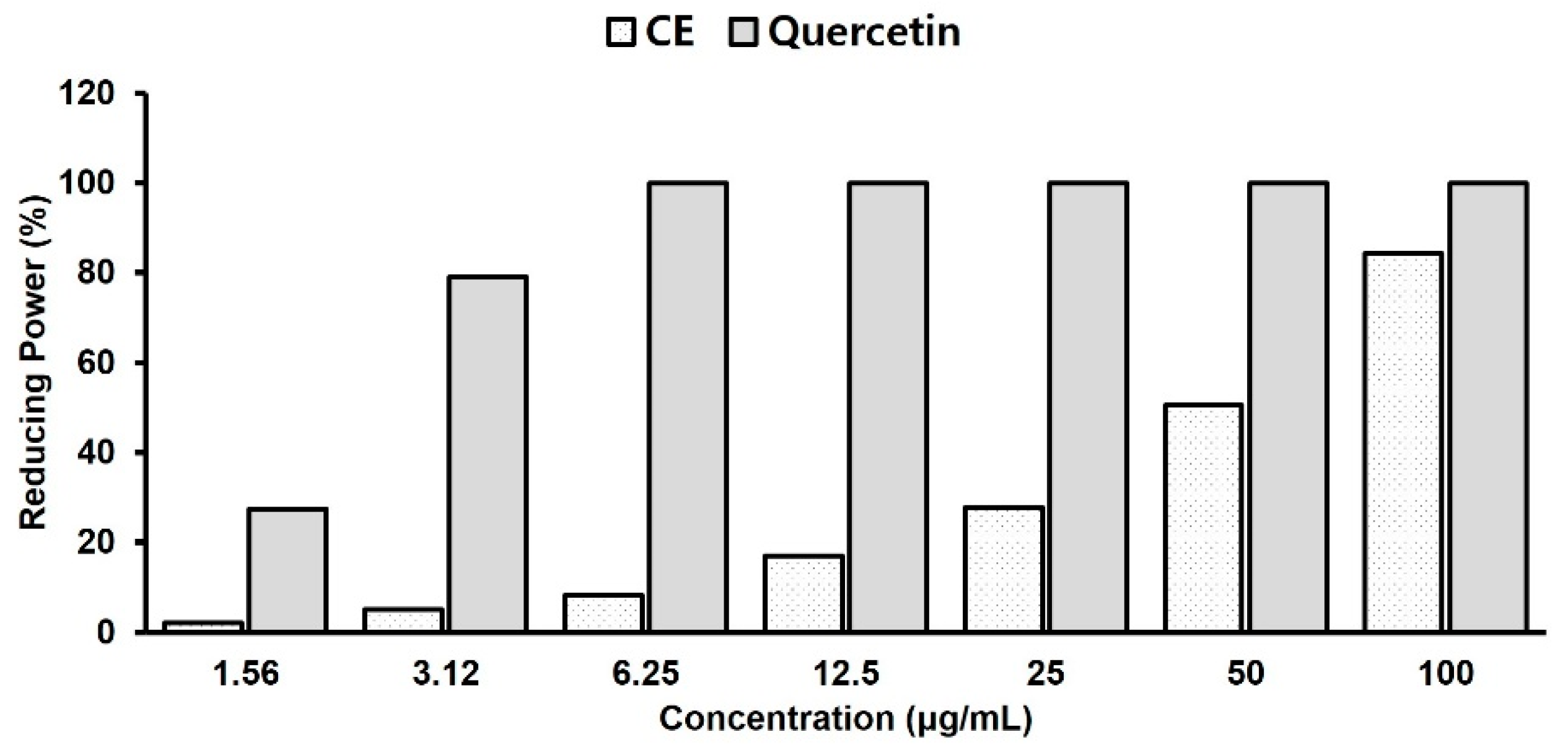
2.3. Antioxidant Activity Test with Reducing Power Assay
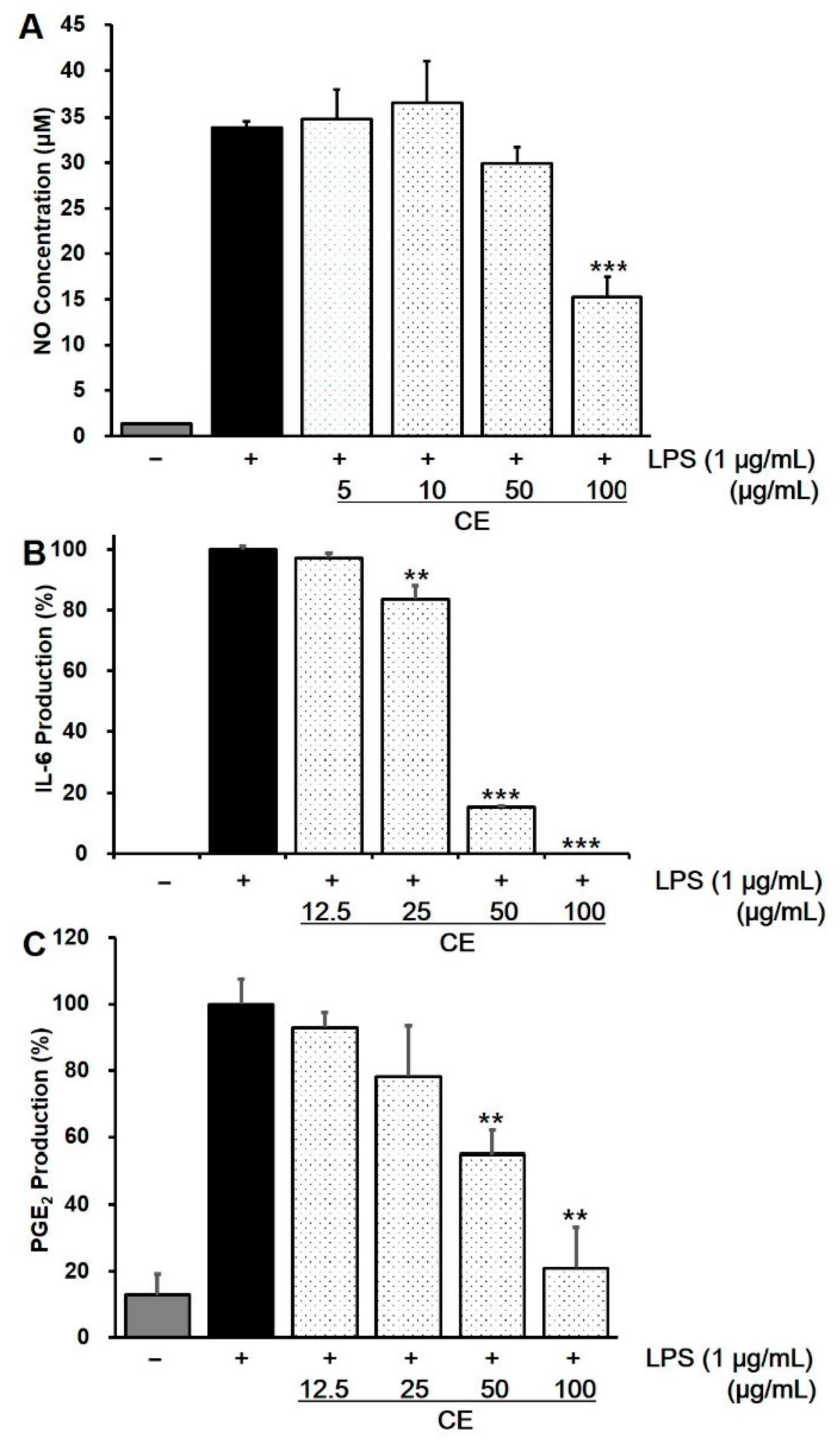
2.4. Effect of CE on Intracellular NO Production in LPS-Stimulated RAW 264.7 Cells
2.5. Effects of CE on LPS-Induced Cytokine Production in RAW 264.7 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. Sample Preparation
4.4. Liquid Chromatography Diode Array Detector Instrumental Condition
4.5. Animals
4.6. HCl/Ethanol-Induced Gastric Ulcer
4.7. Reducing Power Assay
4.8. Cell Culture
4.9. Cell Viability Assay
4.10. Intracellular Nitric Oxide Production
4.11. PGE2 Production
4.12. TNF-α, IL-1β and IL-6 Production
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, L.R. Physiology of the Gastrointestinal Tract; Elsevier Academic Press: Cambridge, UK, 2006; p. 408. [Google Scholar]
- Gugliandolo, E.; Cordaro, M.; Fusco, R.; Peritore, A.F.; Siracusa, R.; Genovese, T.; D’Amico, R.; Impellizzeri, D.; Di Paola, R.; Cuzzocrea, S.; et al. Protective effect of snail secretion filtrate against ethanol-induced gastric ulcer in mice. Sci. Rep. 2021, 11, 3638. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Hansson, L.E. Risk of stomach cancer in patients with peptic ulcer disease. World J. Surg. 2000, 24, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer. 2015, 136, 359–386. [Google Scholar] [CrossRef] [PubMed]
- GBD 2013 Mortality and Causes of Death Collaborators. Global, regional, and national age-sex specific all-cause and cause-specific mortality for 240 causes of death, 1990–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 385, 117–171. [Google Scholar] [CrossRef]
- Carvalho, C.A.; Fernandes, K.M.; Matta, S.L.; Silva, M.B.; Oliveira, L.L.; Fonseca, C.C. Evaluation of antiulcerogenic activity of aqueous extract of Brassica oleracea var. capitata (cabbage) on Wistar rat gastric ulceration. Arq. Gastroenterol. 2011, 48, 276–282. [Google Scholar] [CrossRef]
- Bae, J.-Y.; Lee, Y.S.; Han, S.Y.; Jeong, E.J.; Lee, M.K.; Kong, J.Y.; Lee, D.H.; Cho, K.J.; Lee, H.-S.; Ahn, M.-J. A comparison between water and ethanol extracts of Rumex acetosa for protective effects on gastric ulcers in mice. Biomol. Ther. 2012, 20, 425–430. [Google Scholar] [CrossRef]
- Panda, V.; Sonkamble, M. Anti-ulcer activity of Ipomoea batatas tubers (sweet potato). Funct. Food Health Dis. 2012, 2, 48–61. [Google Scholar] [CrossRef]
- Haniadka, R.; Saldanha, E.; Sunita, V.; Palatty, P.L.; Fayad, R.; Baliga, M.S. A review of the gastroprotective effects of ginger (Zingiber officinale Roscoe). Food Funct. 2013, 4, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Bovell-Benjamin, A.C. Sweet potato: A review of its past, present, and future role in human nutrition. Adv. Food Nutr. Res. 2007, 52, 1–59. [Google Scholar] [CrossRef]
- Neela, S.; Fanta, S.W. Review on nutritional composition of orange-fleshed sweet potato and its role in management of vitamin A deficiency. Food Sci. Nutr. 2019, 7, 1920–1945. [Google Scholar] [CrossRef] [PubMed]
- Hermes, D.; Dudek, D.N.; Maria, M.D.; Horta, L.P.; Lima, E.N.; de Fátima, Â.; Sanches, A.C.; Modolo, L.V. In Vivo wound healing and antiulcer properties of white sweet potato (Ipomoea batatas). J. Adv. Res. 2013, 4, 411–415. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, D.J.; Lin, C.D.; Chen, H.J.; Lin, Y.H. Antioxidant and anti-proliferative activities of sweet potato (Ipomoea batatas [L.] Lam “Tainong 57’’) constituents. Bot. Bull. Acad. Sin. 2004, 45, 179–186. [Google Scholar]
- You, W.C.; Blot, W.J.; Chang, Y.S.; Ershow, A.G.; Yang, Z.T.; An, Q.; Henderson, B.; Xu, G.W.; Fraumeni, J.F., Jr.; Wang, T.G. Diet and high risk of stomach cancer in Shandong, China. Cancer Res. 1988, 48, 3518–3523. [Google Scholar] [CrossRef]
- Low, J.W.; Mwanga, R.O.M.; Andrade, M.; Carey, E.; Ball, A.-M. Tackling vitamin A deficiency with biofortified sweetpotato in sub-Saharan Africa. Glob. Food Sec. 2017, 14, 23–30. [Google Scholar] [CrossRef]
- Oki, T.; Masuda, M.; Furuta, S.; Nishiba, Y.; Terahara, N.; Suda, I. Involvement of anthocyanins and other phenolic compounds in radical-scavenging activity of purple-fleshed sweet potato cultivars. J. Food Sci. 2002, 67, 1752–1756. [Google Scholar] [CrossRef]
- Steed, L.E.; Truong, V.D. Anthocyanin content, antioxidant activity, and selected physical properties of flowable purple-fleshed sweetpotato purees. J. Food Sci. 2008, 73, S215–S221. [Google Scholar] [CrossRef] [PubMed]
- Ayeleso, T.B.; Ramachela, K.; Mukwevho, E. Aqueous-methanol extracts of orange-fleshed sweet potato (Ipomoea batatas) ameliorate oxidative stress and modulate type 2 diabetes associated genes in insulin resistant C2C12 cells. Molecules 2018, 23, 2058. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Koo, K.A.; Park, W.S.; Kang, D.-M.; Kim, H.S.; Lee, B.Y.; Goo, Y.-M.; Kim, J.H.; Lee, M.K.; Woo, D.K.; et al. Anti-obesity activity of anthocyanin and carotenoid extracts from color-fleshed sweet potatoes. J. Food Biochem. 2020, 44, e13438. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Park, W.S.; Bae, J.-Y.; Kang, S.Y.; Yang, M.H.; Lee, S.H.; Lee, H.-S.; Kwak, S.-S.; Ahn, M.-J. Variations in the carotenoid and anthocyanin contents of Korean cultural varieties and home-processed sweet potatoes. J. Food Compos. Anal. 2015, 41, 188–193. [Google Scholar] [CrossRef]
- Murata, K.; Oyagi, A.; Takahira, D.; Tsuruma, K.; Shimazawa, M.; Ishibashi, T.; Hara, H. Protective effects of astaxanthin from Paracoccus carotinifaciens on murine gastric ulcer models. Phytother. Res. 2012, 26, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Oyaizu, M. Studies on products of the browning reaction. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Andreo, M.A.; Ballesteros, K.V.; Hiruma-Lima, C.A.; Machado da Rocha, L.R.; Souza Brito, A.R.; Vilegas, W. Effect of Mouriri pusa extracts on experimentally induced gastric lesions in rodents: Role of endogenous sulfhydryls compounds and nitric oxide in gastroprotection. J. Ethnopharmacol. 2006, 107, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar] [PubMed]
- Zhao, W.; Zhu, F.; Shen, W.; Fu, A.; Zheng, L.; Yan, Z.; Zhao, L.; Fu, G. Protective effects of DIDS against ethanol-induced gastric mucosal injury in rats. Acta Biochim. Biophys. Sin. 2009, 41, 301–308. [Google Scholar] [CrossRef]
- Chen, H.; Nie, Q.; Xie, M.; Yao, H.; Zhang, K.; Yin, J.; Nie, S. Protective effects of β-glucan isolated from highland barley on ethanol-induced gastric damage in rats and its benefits to mice gut conditions. Food Res. Int. 2019, 122, 157–166. [Google Scholar] [CrossRef]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Gao, H.; Chen, S.; Wang, Q.; Li, X.; Du, L.J.; Li, J.; Luo, Y.Y.; Li, J.X.; Zhao, L.C.; et al. Procyanidin A1 alleviates inflammatory response induced by LPS through NF-κB, MAPK, and Nrf2/HO-1 pathways in RAW 264.7 cells. Sci. Rep. 2019, 9, 15087. [Google Scholar] [CrossRef]
- Ronchetti, D.; Impagnatiello, F.; Guzzetta, M.; Gasparini, L.; Borgatti, M.; Gambari, R.; Ongini, E. Modulation of iNOS expression by a nitric oxide-releasing derivative of the natural antioxidant ferulic acid in activated RAW 264.7 macrophages. Eur. J. Pharmacol. 2006, 532, 162–169. [Google Scholar] [CrossRef]
- Yang, Y.; Li, R.; Hui, J.; Li, L.; Zheng, X. β-Carotene attenuates LPS-induced rat intestinal inflammation via modulating autophagy and regulating the JAK2/STAT3 and JNK/p38 MAPK signaling pathways. J. Food Biochem. 2021, 45, e13544. [Google Scholar] [CrossRef]
- Dorrington, M.G.; Fraser, I.D.C. NF-κB signaling in macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Feghali, C.A.; Wright, T.M. Cytokines in acute and chronic inflammation. Front. Biosci. 1997, 2, 12–26. [Google Scholar] [CrossRef]
- Tsuge, K.; Inazumi, T.; Shimamoto, A.; Sugimoto, Y. Molecular mechanisms underlying prostaglandin E2-exacerbated inflammation and immune diseases. Int. Immunol. 2019, 31, 597–606. [Google Scholar] [CrossRef] [PubMed]
- El-Kholy, A.A.; Elkablawy, M.A.; El-Agamy, D.S. Lutein mitigates cyclophosphamide induced lung and liver injury via NF-κB/MAPK dependent mechanism. Biomed. Pharmacother. 2017, 92, 519–527. [Google Scholar] [CrossRef]
- Rovani, B.T.; de Freitas, R.B.; Augusti, P.R.; Araldi, I.C.; Somacal, S.; Quatrin, A.; Emanuelli, T.; da Rocha, M.P.; Bauermann Lde, F. Prooxidant activity of norbixin in model of acute gastric ulcer induced by ethanol in rats. Hum. Exp. Toxicol. 2016, 35, 737–746. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, E.H.; Hahm, K.B. Oxidative stress in inflammation-based gastrointestinal tract diseases: Challenges and opportunities. J. Gastroenterol. Hepatol. 2012, 27, 1004–1010. [Google Scholar] [CrossRef]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid radical chemistry and antioxidant/pro-oxidant properties. Arch. Biochem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Román, J.; García-Gil, S.; Rodríguez-Luna, A.; Motilva, V.; Talero, E. Anti-inflammatory and anticancer effects of microalgal carotenoids. Mar. Drugs 2021, 19, 531. [Google Scholar] [CrossRef]
- Sindhu, E.R.; Kuttan, R. Carotenoid lutein protects rats from gastric ulcer induced by ethanol. J. Basic Clin. Physiol Pharmacol. 2012, 23, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Goulart da Silva, G.; de Oliveira Braga, L.E.; Souza de Oliveira, E.C.; Valério Tinti, S.; de Carvalho, J.E.; Goldoni Lazarini, J.; Rosalen, P.L.; Dionísio, A.P.; Tasca Gois Ruiz, A.L. Cashew apple byproduct: Gastroprotective effects of standardized extract. J. Ethnopharmacol. 2021, 269, 113744. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, D.; Saleh, S.; Al-Shorbagy, M. Beta-carotene protects against indomethacin-induced gastric ulceration: Involvement of antioxidant mechanisms. Az. J. Pharm. Sci. 2007, 36, 265–275. [Google Scholar]
- Ogawa, K.; Oyagi, A.; Tanaka, J.; Kobayashi, S.; Hara, H. The protective effect and action mechanism of Vaccinium myrtillus L. on gastric ulcer in mice. Phytother. Res. 2011, 25, 1160–1165. [Google Scholar] [CrossRef] [PubMed]
- Malyshev, I.Y.; Shnyra, A. Controlled modulation of inflammatory, stress and apoptotic responses in macrophages. Curr. Drug Targets Immune Endocr. Metabol. Disord. 2003, 3, 1–22. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, J.-Y.; Park, W.-S.; Kim, H.-J.; Kim, H.-S.; Kang, K.-K.; Kwak, S.-S.; Ahn, M.-J. Protective Effect of Carotenoid Extract from Orange-Fleshed Sweet Potato on Gastric Ulcer in Mice by Inhibition of NO, IL-6 and PGE2 Production. Pharmaceuticals 2021, 14, 1320. https://doi.org/10.3390/ph14121320
Bae J-Y, Park W-S, Kim H-J, Kim H-S, Kang K-K, Kwak S-S, Ahn M-J. Protective Effect of Carotenoid Extract from Orange-Fleshed Sweet Potato on Gastric Ulcer in Mice by Inhibition of NO, IL-6 and PGE2 Production. Pharmaceuticals. 2021; 14(12):1320. https://doi.org/10.3390/ph14121320
Chicago/Turabian StyleBae, Ji-Yeong, Woo-Sung Park, Hye-Jin Kim, Ho-Soo Kim, Kwon-Kyoo Kang, Sang-Soo Kwak, and Mi-Jeong Ahn. 2021. "Protective Effect of Carotenoid Extract from Orange-Fleshed Sweet Potato on Gastric Ulcer in Mice by Inhibition of NO, IL-6 and PGE2 Production" Pharmaceuticals 14, no. 12: 1320. https://doi.org/10.3390/ph14121320
APA StyleBae, J.-Y., Park, W.-S., Kim, H.-J., Kim, H.-S., Kang, K.-K., Kwak, S.-S., & Ahn, M.-J. (2021). Protective Effect of Carotenoid Extract from Orange-Fleshed Sweet Potato on Gastric Ulcer in Mice by Inhibition of NO, IL-6 and PGE2 Production. Pharmaceuticals, 14(12), 1320. https://doi.org/10.3390/ph14121320