
Teicoplanin—A New Use for an Old Drug in the COVID-19 Era?

Abstract
1. Introduction
2. Teicoplanin Structure
3. Teicoplanin Usage in Medicine
4. Teicoplanin Distribution in the Human Body
5. Teicoplanin Associated Side Effects
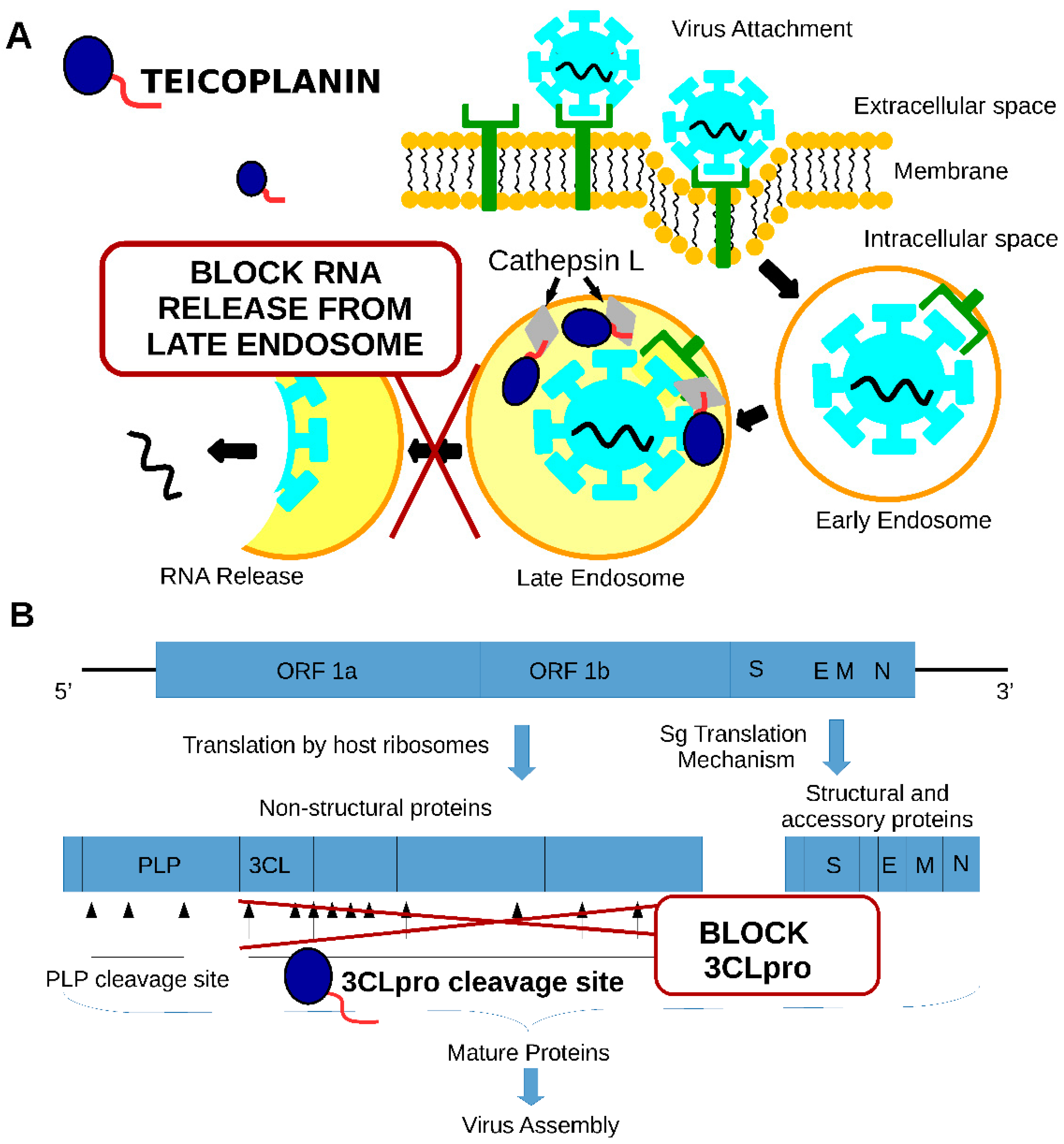
6. Anti-SARS-CoV2 Potential of Teicoplanin
7. Teicoplanin Mechanisms of Action against SARS-CoV1 and SARS-CoV2
8. Bacterial Co-Infections in COVID-19 Patients and the Potential of Teicoplanin Usage
9. Threats to Be Considered in Teicoplanin Usage
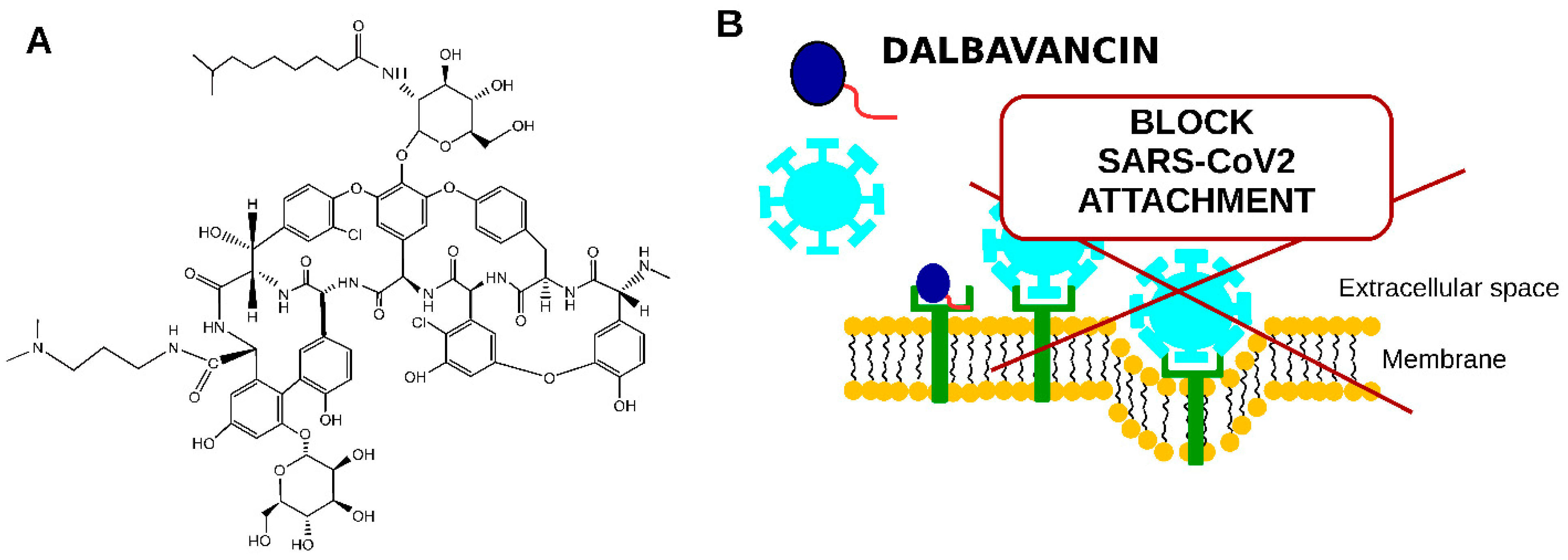
10. Novel Semisynthetic Lipoglycopeptide Antibiotics in COVID-19 Treatment
11. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Novelli, G.; Biancolella, M.; Mehrian-shai, R.; Colona, V.L.; Brito, A.F.; Grubaugh, N.D.; Vasiliou, V.; Luzzatto, L.; Reichardt, J.K. V COVID-19 one year into the pandemic: From genetics and genomics to therapy, vaccination, and policy. Hum. Genom. 2021, 15, 27. [Google Scholar] [CrossRef]
- Angeli, F.; Spanevello, A.; Reboldi, G.; Visca, D.; Verdecchia, P. SARS-CoV-2 vaccines: Lights and shadows. Eur. J. Intern. Med. 2021, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Wheatley, A.K. Neutralizing Antibody Therapeutics for COVID-19. Viruses 2021, 13, 628. [Google Scholar] [CrossRef] [PubMed]
- Kifle, Z.D.; Ayele, A.G.; Enyew, E.F. Drug Repurposing Approach, Potential Drugs, and Novel Drug Targets for COVID-19 Treatment. J. Environ. Public Health 2021, 2021, 6631721. [Google Scholar] [CrossRef] [PubMed]
- Parenti, F.; Beretta, G.; Berti, M.; Arioli, V. Teichomycins, New Antibiotics from Actinoplanes Teichomyceticus Nov. SP. I. Description of the Producer Strain, Fermentation Studies and Biological Properties. J. Antibiot. 1978, 31, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Barna, J.C.J.; Williams, D.H.; Stone, D.J.M.; Leung, T.W.C.; Doddrell, D.M. Structure Elucidation of the Teicoplanin Antibiotics. J. Am. Chem. Soc. 1984, 106, 4895–4902. [Google Scholar] [CrossRef]
- Hunt, A.H.; Molloy, R.M.; Occolowitz, J.L.; Marconi, G.G.; Debono, M. Structure of the Major Glycopeptide of the Teicoplanin Complex. J. Am. Chem. Soc. 1984, 106, 4891–4895. [Google Scholar] [CrossRef]
- Zanol, M.; Cometti, A.; Borghi, A.; Lancini, G.C. Isolation and structure determination of minor components of teicoplanin. Chromatographia 1988, 26, 234–236. [Google Scholar] [CrossRef]
- Glupczynski, Y.; Lagast, H.; van der Auwera, P.; Thys, J.P.; Crokaert, F.; Yourassowsky, E.; Meunier-Carpentier, F.; Klastersky, J.; Kains, J.P.; Serruys-Schoutens, E. Clinical evaluation of teicoplanin for therapy of severe infections caused by gram-positive bacteria. Antimicrob. Agents Chemother. 1986, 29, 52–57. [Google Scholar] [CrossRef]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved glycopeptide antibacterial drugs: Mechanism of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef]
- Vimberg, V.; Gazak, R.; Szűcs, Z.; Borbás, A.; Herczegh, P.; Cavanagh, J.P.; Zieglerova, L.; Závora, J.; Adámková, V.; Balikova Novotna, G. Fluorescence assay to predict activity of the glycopeptide antibiotics. J. Antibiot. 2019, 72, 114–117. [Google Scholar] [CrossRef]
- Pea, F. Teicoplanin and therapeutic drug monitoring: An update for optimal use in different patient populations. J. Infect. Chemother. 2020, 26, 900–907. [Google Scholar] [CrossRef]
- Pea, F.; Brollo, L.; Viale, P.; Pavan, F.; Furlanut, M. Teicoplanin therapeutic drug monitoring in critically ill patients: A retrospective study emphasizing the importance of a loading dose. J. Antimicrob. Chemother. 2003, 51, 971–975. [Google Scholar] [CrossRef]
- Tascini, C.; Gemignani, G.; Doria, R.; Biancofiore, G.; Urbani, L.; Mosca, C.; Malacarne, P.; Papineschi, F.; Passaglia, C.; Dal Canto, L.; et al. Linezolid treatment for Gram-positive infections: A retrospective comparison with teicoplanin. J. Chemother. 2009, 21, 311–316. [Google Scholar] [CrossRef]
- Brink, A.J.; Richards, G.A.; Cummins, R.R.; Lambson, J. Recommendations to achieve rapid therapeutic teicoplanin plasma concentrations in adult hospitalised patients treated for sepsis. Int. J. Antimicrob. Agents 2008, 32, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Bernareggi, A.; Borgonovi, M.; Del Favero, A.; Rosina, R.; Gavanaghi, L. Teicoplanin binding in plasma following administration of increasing intravenous doses to healthy volunteers. Eur. J. Drug Metab. Pharmacokinet. 1991, Spec No 3, 256–260. [Google Scholar]
- Smithers, J.A.; Kulmala, H.K.; Thompson, G.A.; Antony, K.K.; Lewis, E.W.; Ruberg, S.J.; Kenny, M.T.; Dulworth, J.K.; Brackman, M.A. Pharmacokinetics of teicoplanin upon multiple-dose intravenous administration of 3, 12, and 30 milligrams per kilogram of body weight to healthy male volunteers. Antimicrob. Agents Chemother. 1992, 36, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Takesue, Y.; Nakajima, K.; Ichiki, K.; Doita, A.; Wada, Y.; Tsuchida, T.; Takahashi, Y.; Ishihara, M.; Ikeuchi, H.; et al. Enhanced loading regimen of teicoplanin is necessary to achieve therapeutic pharmacokinetics levels for the improvement of clinical outcomes in patients with renal dysfunction. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1501–1509. [Google Scholar] [CrossRef]
- Rybak, M.J.; Lerner, S.A.; Levine, D.P.; Albrecht, L.M.; McNeil, P.L.; Thompson, G.A.; Kenny, M.T.; Yuh, L. Teicoplanin pharmacokinetics in intravenous drug abusers being treated for bacterial endocarditis. Antimicrob. Agents Chemother. 1991, 35, 696–700. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bergeron, M.G.; Saginur, R.; Desaulniers, D.; Trottier, S.; Goldstein, W.; Foucault, P.; Lessard, C. Concentrations of Teicoplanin in serum and atrial appendages of patients undergoing cardiac surgery. Antimicrob. Agents Chemother. 1990, 34, 1699–1702. [Google Scholar] [CrossRef] [PubMed]
- Mimoz, O.; Rolland, D.; Adoun, M.; Marchand, S.; Breilh, D.; Brumpt, I.; Debaene, B.; Couet, W. Steady-state trough serum and epithelial lining fluid concentrations of teicoplanin 12 mg/kg per day in patients with ventilator-associated pneumonia. Intensive Care Med. 2006, 32, 775–779. [Google Scholar] [CrossRef]
- Yanagihara, K.; Kaneko, Y.; Sawai, T.; Miyazaki, Y.; Tsukamoto, K.; Hirakata, Y.; Tomono, K.; Kadota, J.I.; Tashiro, T.; Murata, I.; et al. Efficacy of linezolid against methicillin-resistant or vancomycin-insensitive Staphylococcus aureus in a model of hematogenous pulmonary infection. Antimicrob. Agents Chemother. 2002, 46, 3288–3291. [Google Scholar] [CrossRef]
- Luna, C.M.; Bruno, D.A.; García-Morato, J.; Mann, K.C.; Patrón, J.R.; Sagardía, J.; Absi, R.; Bottino, M.G.; Marchetti, D.; Famiglietti, A.; et al. Effect of linezolid compared with glycopeptides in methicillin-resistant Staphylococcus aureus severe pneumonia in piglets. Chest 2009, 135, 1564–1571. [Google Scholar] [CrossRef]
- Naruse, N.; Oka, M.; Konishi, M.; Oki, T. New antiviral antibiotics, kistamicins a and b ii. structure determination. J. Antibiot. 1993, 46, 1812–1818. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Naruse, N.; Tenmyo, O.; Kobaru, S.; Hatori, M.; Tomita, K.; Hamagishi, Y.; Oki, T. New antiviral antibiotics, kistamicins a and b i. taxonomy, production, isolation, physico-chemical properties and biological activities. J. Antibiot. 1993, 46, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Pannecouque, C.; De Clercq, E.; Pavlov, A.Y.; Printsevskaya, S.S.; Miroshnikova, O.V.; Reznikova, M.I.; Preobrazhenskaya, M.N. Antiretroviral activity of semisynthetic derivatives of glycopeptide antibiotics. J. Med. Chem. 2003, 46, 2755–2764. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cui, R.; Li, G.; Gao, Q.; Yuan, S.; Altmeyer, R.; Zou, G. Teicoplanin inhibits Ebola pseudovirus infection in cell culture. Antivir. Res. 2016, 125, 1–7. [Google Scholar] [CrossRef]
- Zhou, N.; Pan, T.; Zhang, J.; Li, Q.; Zhang, X.; Bai, C.; Huang, F.; Peng, T.; Zhang, J.; Liu, C.; et al. Glycopeptide antibiotics potently inhibit cathepsin l in the late endosome/lysosome and block the entry of ebola virus, middle east respiratory syndrome coronavirus (MERS-CoV), and severe acute respiratory syndrome coronavirus (SARS-CoV). J. Biol. Chem. 2016, 291, 9218–9232. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, X.; Yu, F.; Liu, J.; Zou, F.; Pan, T.; Zhang, H. Teicoplanin potently blocks the cell entry of 2019-nCoV. BioRxiv 2020. [Google Scholar] [CrossRef]
- Gomes, C.P.; Fernandes, D.E.; Casimiro, F.; da Mata, G.F.; Passos, M.T.; Varela, P.; Mastroianni-Kirsztajn, G.; Pesquero, J.B. Cathepsin L in COVID-19: From Pharmacological Evidences to Genetics. Front. Cell. Infect. Microbiol. 2020, 10, 589505. [Google Scholar] [CrossRef]
- Tripathi, P.K.; Upadhyay, S.; Singh, M.; Raghavendhar, S.; Bhardwaj, M.; Sharma, P.; Patel, A.K. Screening and evaluation of approved drugs as inhibitors of main protease of SARS-CoV-2. Int. J. Biol. Macromol. 2020, 164, 2622–2631. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.S.; Hsueh, P.R. Old and re-purposed drugs for the treatment of COVID-19. Expert Rev. Anti-Infect. Ther. 2020, 18, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.A.; Devaux, C.; Colson, P.; Raoult, D.; Rolain, J.M. Teicoplanin: An alternative drug for the treatment of COVID-19? Int. J. Antimicrob. Agents 2020, 55, 105944. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, G.; Alessandri, F.; d’Ettorre, G.; Borrazzo, C.; Spagnolello, O.; Oliva, A.; Ruberto, F.; Mastroianni, C.M.; Pugliese, F.; Venditti, M. Is teicoplanin a complementary treatment option for COVID-19? The question remains. Int. J. Antimicrob. Agents 2020, 56, 106029. [Google Scholar] [CrossRef]
- Ceccarelli, G.; Alessandri, F.; Oliva, A.; Borrazzo, C.; Dell’Isola, S.; Ialungo, A.M.; Rastrelli, E.; Pelli, M.; Raponi, G.; Turriziani, O.; et al. The role of teicoplanin in the treatment of SARS-CoV-2 infection: A retrospective study in critically ill COVID-19 patients (Tei-COVID study). J. Med. Virol. 2021, 93, 4319–4325. [Google Scholar] [CrossRef]
- Westblade, L.F.; Simon, M.S.; Satlin, M.J. Bacterial Co-Infections in Coronavirus Disease 2019. Trends Microbiol. 2021, 29, 930–941. [Google Scholar] [CrossRef]
- Hughes, S.; Troise, O.; Donaldson, H.; Mughal, N.; Moore, L.S.P. Bacterial and fungal coinfection among hospitalized patients with COVID-19: A retrospective cohort study in a UK secondary-care setting. Clin. Microbiol. Infect. 2020, 26, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- Kreitmann, L.; Monard, C.; Dauwalder, O.; Simon, M.; Argaud, L. Early bacterial co-infection in ARDS related to COVID-19. Intensive Care Med. 2020, 46, 1787–1789. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.; Chen, S.; Ko, W.; Hsueh, P. Increased antimicrobial resistance during the COVID-19 pandemic. Int. J. Antimicrob. Agents 2021, 57, 106324. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Hansford, K.A.; Blaskovich, M.A.T.; Halai, R.; Cooper, M.A. Glycopeptide antibiotics: Back to the future. J. Antibiot. 2014, 67, 631–644. [Google Scholar] [CrossRef]
- Blaskovich, M.A.T.; Hansford, K.A.; Butler, M.S.; Jia, Z.; Mark, A.E.; Cooper, M.A. Developments in Glycopeptide Antibiotics. ACS Infect. Dis. 2018, 4, 715–735. [Google Scholar] [CrossRef] [PubMed]
- Vimberg, V.; Cavanagh, J.P.; Benada, O.; Kofroňová, O.; Hjerde, E.; Zieglerová, L.; Balíková Novotná, G. Teicoplanin resistance in Staphylococcus haemolyticus is associated with mutations in histidine kinases VraS and WalK. Diagn. Microbiol. Infect. Dis. 2018, 90, 233–240. [Google Scholar] [CrossRef]
- Lee, J.Y.H.; Monk, I.R.; Gonçalves da Silva, A.; Seemann, T.; Chua, K.Y.L.; Kearns, A.; Hill, R.; Woodford, N.; Bartels, M.D.; Strommenger, B.; et al. Global spread of three multidrug-resistant lineages of Staphylococcus epidermidis. Nat. Microbiol. 2018, 3, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Vimberg, V.; Zieglerova, L.; Závora, J.; Šemberová, L.; Prásilová, J.; Adámková, V.; Novotna, G.B. Draft genome sequences of three clinical isolates of teicoplanin-resistant Staphylococcus epidermidis from patients without prior exposure to glycopeptide antibiotics. J. Glob. Antimicrob. Resist. 2019, 16, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Malabarba, A.; Goldstein, B.P. Origin, structure, and activity in vitro and in vivo of dalbavancin. J. Antimicrob. Chemother. 2005, 55, ii15–ii20. [Google Scholar] [CrossRef] [PubMed]
- Yushchuk, O.; Andreo-Vidal, A.; Marcone, G.L.; Bibb, M.; Marinelli, F.; Binda, E. New Molecular Tools for Regulation and Improvement of A40926 Glycopeptide Antibiotic Production in Nonomuraea gerenzanensis ATCC 39727. Front. Microbiol. 2020, 11, 8. [Google Scholar] [CrossRef]
- Smith, J.R.; Roberts, K.D.; Rybak, M.J. Dalbavancin: A Novel Lipoglycopeptide Antibiotic with Extended Activity against Gram-Positive Infections. Infect. Dis. Ther. 2015, 4, 245–258. [Google Scholar] [CrossRef]
- Economou, N.J.; Nahoum, V.; Weeks, S.D.; Grasty, K.C.; Zentner, I.J.; Townsend, T.M.; Bhuiya, M.W.; Cocklin, S.; Loll, P.J. A carrier protein strategy yields the structure of dalbavancin. J. Am. Chem. Soc. 2012, 134, 4637–4645. [Google Scholar] [CrossRef]
- Wang, G.; Yang, M.L.; Duan, Z.L.; Liu, F.L.; Jin, L.; Long, C.B.; Zhang, M.; Tang, X.P.; Xu, L.; Li, Y.C.; et al. Dalbavancin binds ACE2 to block its interaction with SARS-CoV-2 spike protein and is effective in inhibiting SARS-CoV-2 infection in animal models. Cell Res. 2021, 31, 17–24. [Google Scholar] [CrossRef]
- Rappo, U.; Dunne, M.W.; Puttagunta, S.; Baldassarre, J.S.; Su, S.; Desai-Krieger, D.; Inoue, M. Epithelial lining fluid and plasma concentrations of dalbavancin in healthy adults after a single 1,500-milligram infusion. Antimicrob. Agents Chemother. 2019, 63, e01024-19. [Google Scholar] [CrossRef]
- Hoffmann, M.; Jin, Y.; Pöhlmann, S. Dalbavancin: Novel candidate for COVID-19 treatment. Cell Res. 2021, 31, 243–244. [Google Scholar] [CrossRef] [PubMed]
- Szűcs, Z.; Kelemen, V.; Le Thai, S.; Csávás, M.; Rőth, E.; Batta, G.; Stevaert, A.; Vanderlinden, E.; Naesens, L.; Herczegh, P.; et al. Structure-activity relationship studies of lipophilic teicoplanin pseudoaglycon derivatives as new anti-influenza virus agents. Eur. J. Med. Chem. 2018, 157, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Bereczki, I.; Csávás, M.; Szűcs, Z.; Rőth, E.; Batta, G.; Ostorházi, E.; Naesens, L.; Borbás, A.; Herczegh, P. Synthesis of Antiviral Perfluoroalkyl Derivatives of Teicoplanin and Vancomycin. ChemMedChem 2020, 15, 1661–1671. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infections in | Cured (%) | Improved (%) | Failed (%) |
|---|---|---|---|
| Soft Tissue | 76 | 15 | 9 |
| Bone/Joint | 50 | 19 | 31 |
| Septicemia | 65 | 11 | 24 |
| Lung | 68 | 11 | 21 |
| Lower Respiratory Tract | 73 | 8 | 19 |
| Upper Respiratory Tract | 73 | 20 | 7 |
| Endocarditis | 70 | 5 | 25 |
| Urinary Tract | 76 | 3 | 21 |
| Total | 67 | 17 | 16 |
| Human | Dosage | Concentration | |
| Urine | 2 or 3 mg/kg (single dose) | 22.4 µg/mL | |
| 440 mg (single dose) | 25–61 µg/mL | ||
| Bone | 400–800 mg (single dose) | 1.3–12.7 µg/mL | |
| 10 mg/kg | 0.55–25.91 µg/mL | ||
| CFS | 400 mg (single dose) | 1.3 µg/mL (peak on Day 2) | |
| 10–15 mg/kg | 2.1–7.2 µg/mL | ||
| Epithelial Lining Fluid | 12 mg/kg | 4.9 (2.0–11.8) µg/mL | |
| Heart | 6–12 mg/kg | 70.6–139.8 µg/g | |
| Dialysate | 6 mg/kg (single dose) | 0.69–1.63 µg/mL | |
| Skin | 400–800 mg (single dose) | 1–8.2 µg/mL |
| Side Effects | Frequency (%) |
|---|---|
| Nephrotoxicity | 5.98 |
| Pain in injection site | 4.93 |
| Hearing problem | 4.76 |
| Drug fever | 3.15 |
| Rash | 1.68 |
| Year | Virus | Cell Line | IC50 (Luciferase) |
|---|---|---|---|
| 2016 | HIV-luc/SARS-CoV-S pseudotyped viruses | HEK293T | 0.39 µM |
| 2019 | 2019-nCoV-Spike-pseudoviruses | A549 cells | 1.66 μM |
| Year | Virus | Cell Line | EC50 |
| 2021 | SARS-CoV-2 | Vero E6 cell | 15.7 µM |
| Gram-Positive Bacteria | Gram-Negative Bacteria |
|---|---|
| CoNS | P. aeruginosa |
| S. aureus | K. pneumoniae |
| S. pneumoniae | H. influenzae |
| E. faecium | E. coli |
| E. faecalis | S. maltophilia |
| μg/mL | ||
|---|---|---|
| Time (Hours) | Plasma (SD) | ELF (SD) |
| 4 | 279 (32) | 1.9 (1.0) |
| 8 | 222 (27) | 3.1 (1.9) |
| 12 | 194 (24) | 3.6 (2.1) |
| 24 | 169 (20) | 2.7 (0.5) |
| 72 | 120 (14) | 7.3 (8.2) |
| 120 | 94 (11) | 11.9 (20.1) |
| 168 | 79 (9) | 2.0 (0.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vimberg, V. Teicoplanin—A New Use for an Old Drug in the COVID-19 Era? Pharmaceuticals 2021, 14, 1227. https://doi.org/10.3390/ph14121227
Vimberg V. Teicoplanin—A New Use for an Old Drug in the COVID-19 Era? Pharmaceuticals. 2021; 14(12):1227. https://doi.org/10.3390/ph14121227
Chicago/Turabian StyleVimberg, Vladimir. 2021. "Teicoplanin—A New Use for an Old Drug in the COVID-19 Era?" Pharmaceuticals 14, no. 12: 1227. https://doi.org/10.3390/ph14121227
APA StyleVimberg, V. (2021). Teicoplanin—A New Use for an Old Drug in the COVID-19 Era? Pharmaceuticals, 14(12), 1227. https://doi.org/10.3390/ph14121227