
Effects of an Atypical Antipsychotic, Zotepine, on Astroglial L-Glutamate Release through Hemichannels: Exploring the Mechanism of Mood-Stabilising Antipsychotic Actions and Antipsychotic-Induced Convulsion


Abstract
:1. Introduction
2. Results
2.1. Effects of ZTP on Astroglial l-Glutamate Release
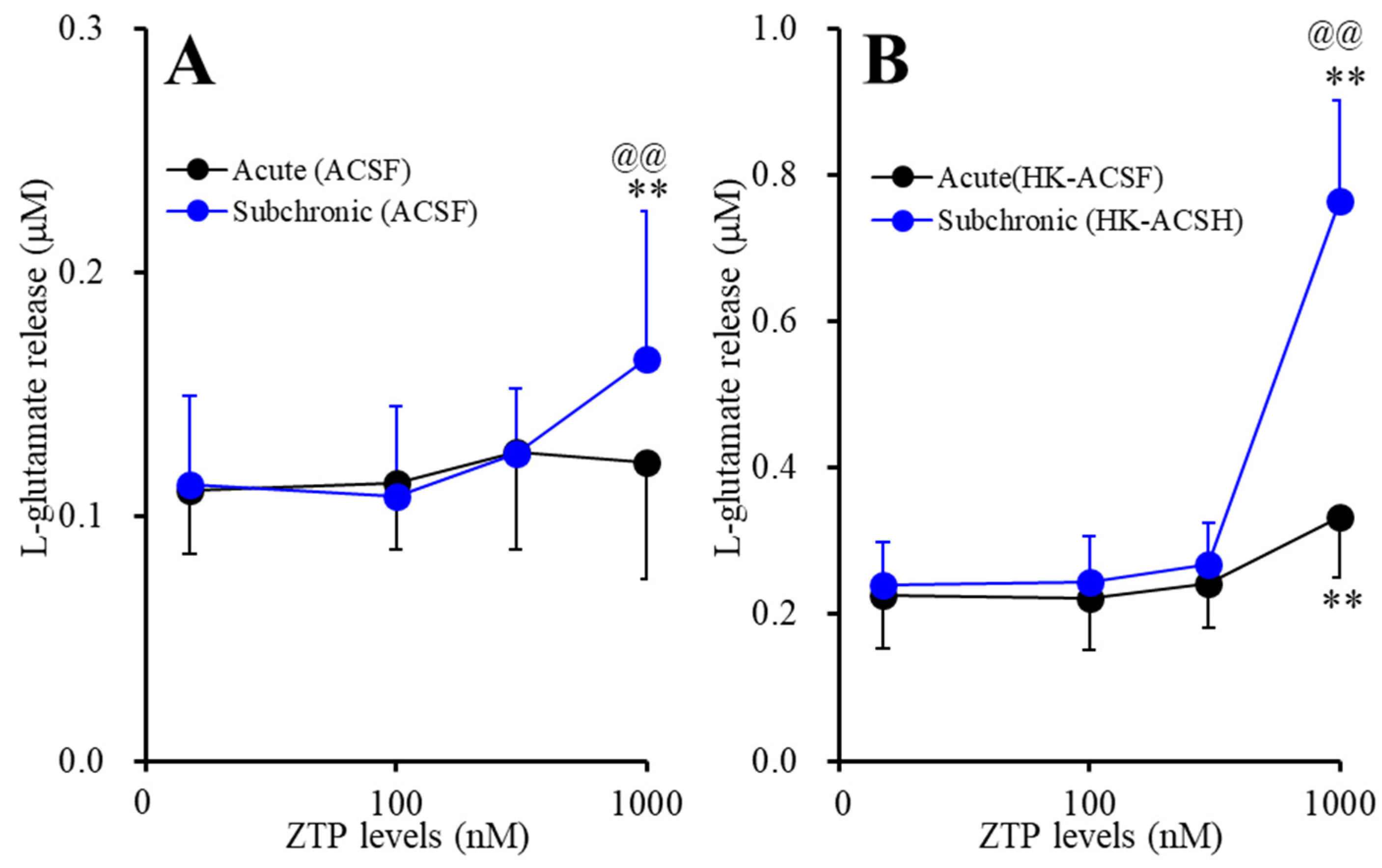
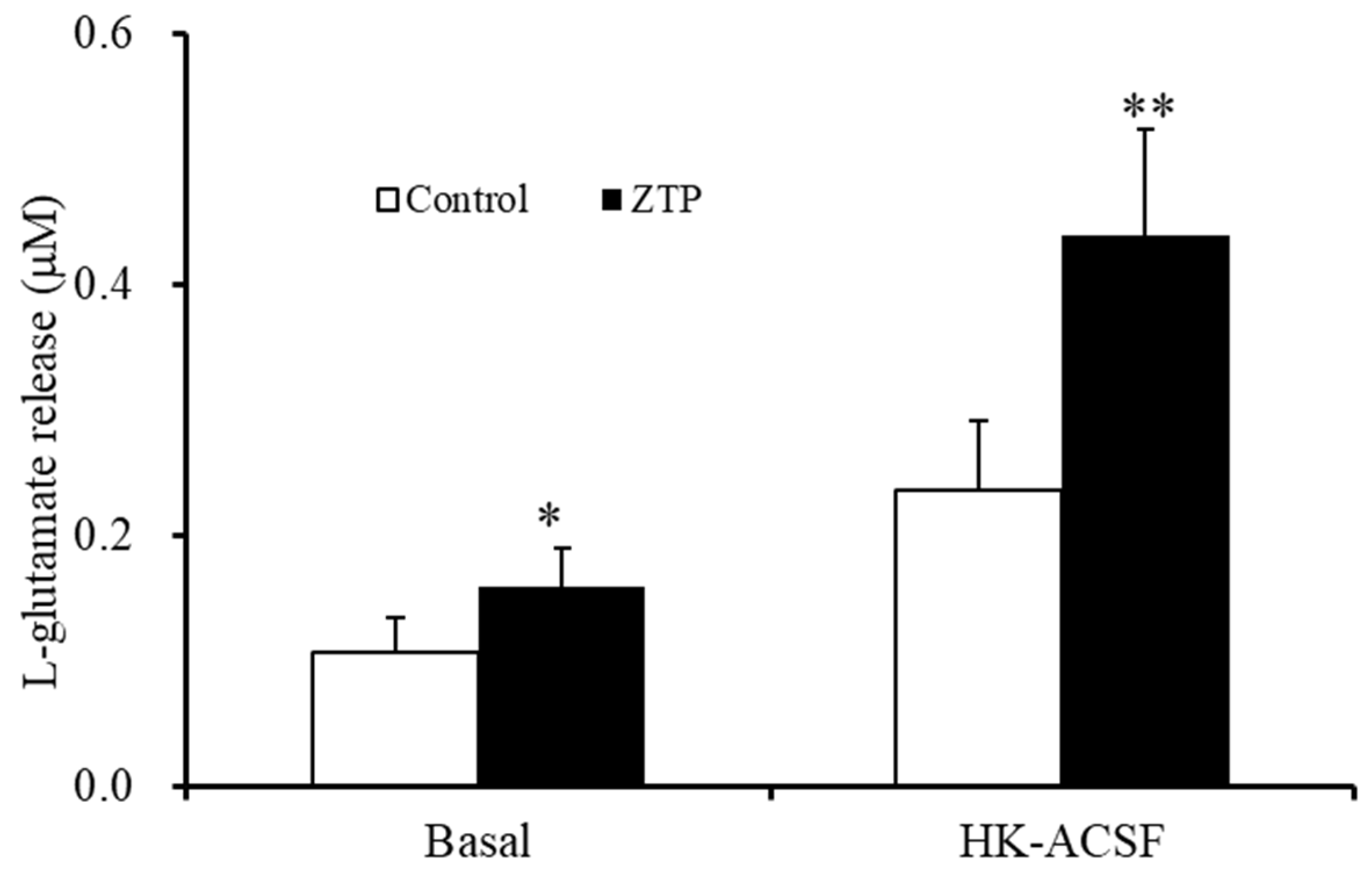
2.1.1. Concentration-Dependent Effects of Acute and Subchronic Administration of ZTP on Astroglial l-Glutamate Release
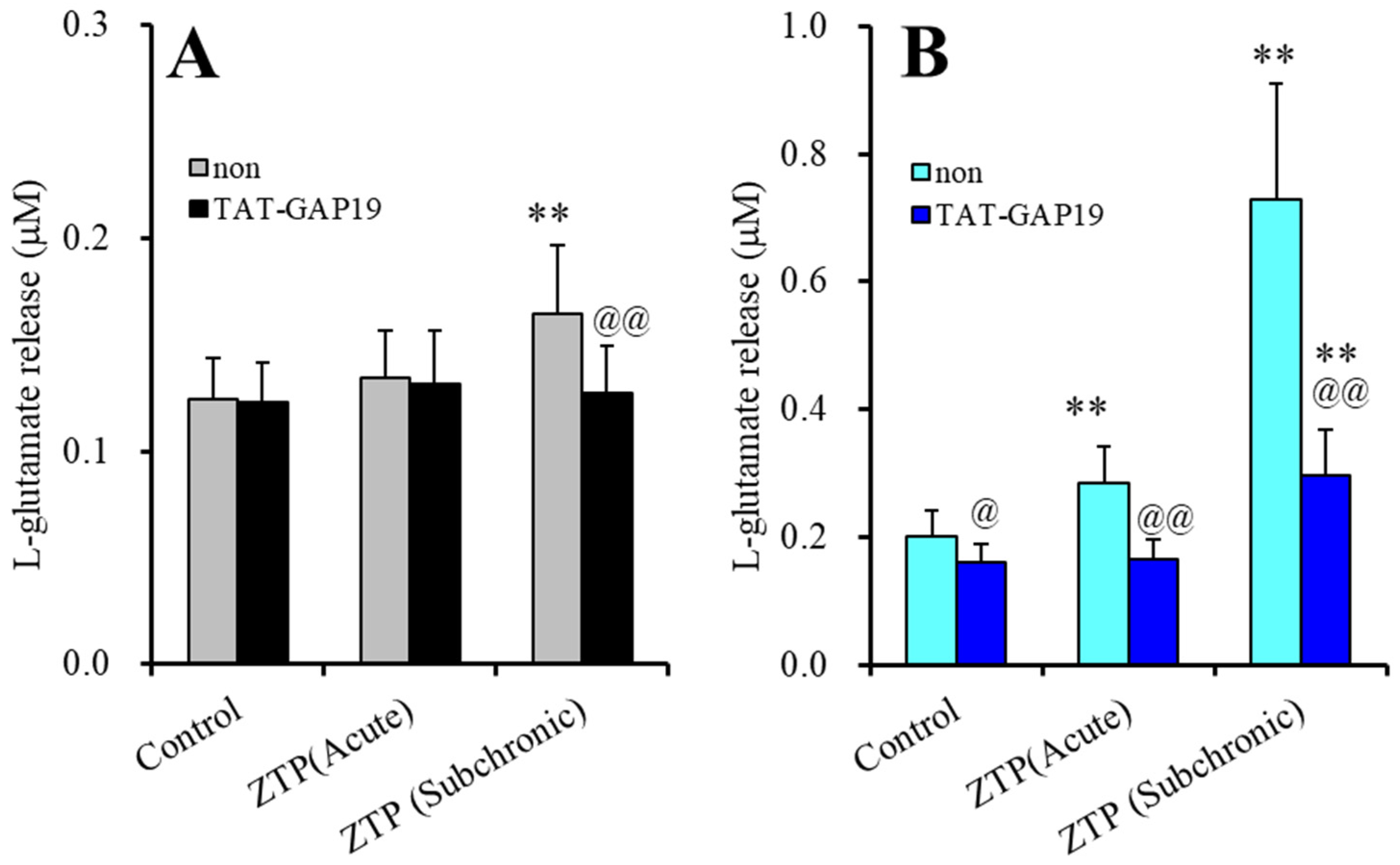
2.1.2. Interaction between Supratherapeutic Concentrations of ZTP and Astroglial Hemichannel Inhibitors on Astroglial l-Glutamate Release
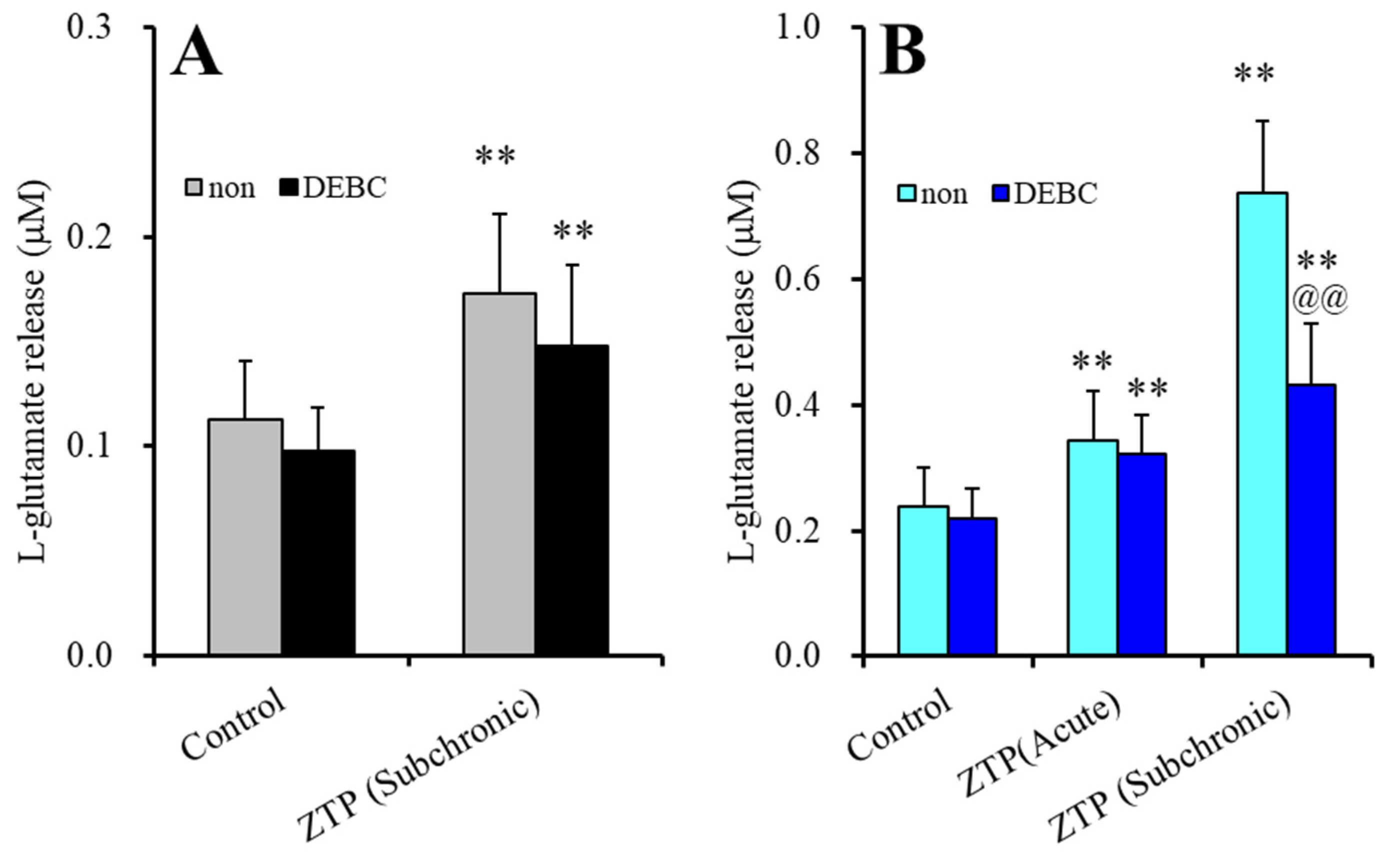
2.1.3. Interaction between Supratherapeutic Concentrations of ZTP and the Inhibitor of Protein Kinase B on Astroglial l-Glutamate Release
2.2. Effects of ZTP on the Expression of Cx43 Protein in Astrocytes
2.2.1. Concentration-Dependent Effects of Subchronic Administration of ZTP on Cx43 Expression in the Cytosol Fraction
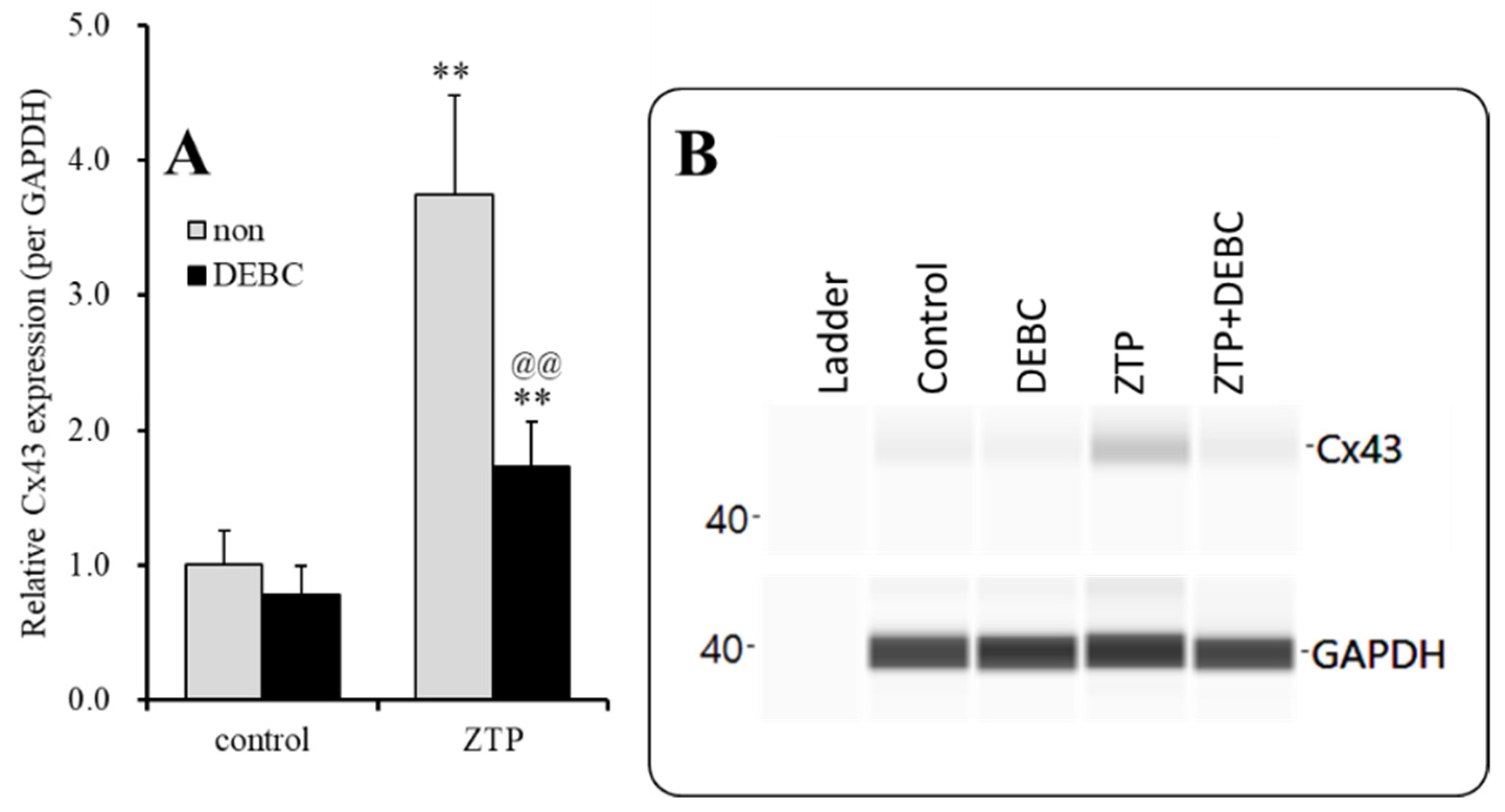
2.2.2. Interaction between Subchronic Administration of a Supratherapeutic Concentration of ZTP and Acute Administration of an Akt Inhibitor on Cx43 Protein Expression in the Plasma Membrane Fraction
2.3. Effects of Acute Administration of a Supratherapeutic Concentration of ZTP on Cx43 Expression in the Plasma Membrane Fraction and Chronic Administration of Therapeutically Relevant Concentrations of ZTP on Cx43 Expression in the Plasma Membrane Fraction and Astroglial L-Glutamate Release
2.3.1. Effects of Acute Administration of Supratherapeutic Concentration of ZTP and Chronic Administration of Therapeutic-relevant Concentration of ZTP on Cx43 Expression in the Plasma Membrane Fraction
2.3.2. Effects of Chronic Administration of Therapeutically Relevant Concentrations of ZTP on Astroglial L-glutamate Release
3. Discussion
3.1. Effects of ZTP on Astroglial L-Glutamate Release and Hemichannel Activity
3.2. A Candidate Mechanism of the Clinical Action of ZTP Associated with Astrocytes
4. Materials and Methods
4.1. Chemical Agents
4.2. Preparation of Primary Astrocyte Culture
4.3. Ultra-High-Performance Liquid Chromatography (UHPLC)
4.4. Capillary Immunoblotting Analysis
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shobo, M.; Kondo, Y.; Yamada, H.; Mihara, T.; Yamamoto, N.; Katsuoka, M.; Harada, K.; Ni, K.; Matsuoka, N. Norzotepine, a major metabolite of zotepine, exerts atypical antipsychotic-like and antidepressant-like actions through its potent inhibition of norepinephrine reuptake. J. Pharmacol. Exp. Ther. 2010, 333, 772–781. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, H.Y.; Matsubara, S.; Lee, J.C. Classification of typical and atypical antipsychotic drugs on the basis of dopamine d-1, d-2 and serotonin2 pki values. J. Pharmacol. Exp. Ther. 1989, 251, 238–246. [Google Scholar] [PubMed]
- Stockmeier, C.A.; DiCarlo, J.J.; Zhang, Y.; Thompson, P.; Meltzer, H.Y. Characterization of typical and atypical antipsychotic drugs based on in vivo occupancy of serotonin2 and dopamine2 receptors. J. Pharmacol. Exp. Ther. 1993, 266, 1374–1384. [Google Scholar] [PubMed]
- Prakash, A.; Lamb, H.M. Zotepine. CNS Drugs 1998, 9, 153–175. [Google Scholar] [CrossRef]
- Schotte, A.; Janssen, P.F.; Gommeren, W.; Luyten, W.H.; Van Gompel, P.; Lesage, A.S.; De Loore, K.; Leysen, J.E. Risperidone compared with new and reference antipsychotic drugs: In vitro and in vivo receptor binding. Psychopharmacology 1996, 124, 57–73. [Google Scholar] [CrossRef]
- Cooper, S.J.; Tweed, J.; Raniwalla, J.; Butler, A.; Welch, C. A placebo-controlled comparison of zotepine versus chlorpromazine in patients with acute exacerbation of schizophrenia. Acta Psychiatr. Scand. 2000, 101, 218–225. [Google Scholar] [CrossRef]
- Davis, J.M.; Chen, N.; Glick, I.D. A meta-analysis of the efficacy of second-generation antipsychotics. Arch. Gen. Psychiatry 2003, 60, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Barnas, C.; Stuppack, C.H.; Miller, C.; Haring, C.; Sperner-Unterweger, B.; Fleischhacker, W.W. Zotepine in the treatment of schizophrenic patients with prevailingly negative symptoms. A double-blind trial vs. Haloperidol. Int. Clin. Psychopharmacol. 1992, 7, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Lindenberg, A.; Gruppe, H.; Bauer, U.; Lis, S.; Krieger, S.; Gallhofer, B. Improvement of cognitive function in schizophrenic patients receiving clozapine or zotepine: Results from a double-blind study. Pharmacopsychiatry 1997, 30, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Amann, B.; Sterr, A.; Mergl, R.; Dittmann, S.; Seemuller, F.; Dobmeier, M.; Orth, M.; Schaefer, M.; Grunze, H. Zotepine loading in acute and severely manic patients: A pilot study. Bipolar Disord. 2005, 7, 471–476. [Google Scholar] [CrossRef]
- Dieterle, D.M.; Ackenheil, M.; Muller-Spahn, F.; Kapfhammer, H.P. Zotepine, a neuroleptic drug with a bipolar therapeutic profile. Pharmacopsychiatry 1987, 20, 52–57. [Google Scholar] [CrossRef]
- Scherk, H.; Pajonk, F.G.; Leucht, S. Second-generation antipsychotic agents in the treatment of acute mania: A systematic review and meta-analysis of randomized controlled trials. Arch. Gen. Psychiatry 2007, 64, 442–455. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.Y.; Jou, S.H.; Juang, Y.Y.; Chang, C.J.; Chen, J.J.; Chen, C.H.; Chiu, N.Y. A single-blind, comparative study of zotepine versus haloperidol in combination with a mood stabilizer for patients with moderate-to-severe mania. Psychiatry Clin. Neurosci. 2010, 64, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Huhn, M.; Nikolakopoulou, A.; Schneider-Thoma, J.; Krause, M.; Samara, M.; Peter, N.; Arndt, T.; Backers, L.; Rothe, P.; Cipriani, A.; et al. Comparative efficacy and tolerability of 32 oral antipsychotics for the acute treatment of adults with multi-episode schizophrenia: A systematic review and network meta-analysis. Lancet 2019, 394, 939–951. [Google Scholar] [CrossRef] [Green Version]
- Alper, K.; Schwartz, K.A.; Kolts, R.L.; Khan, A. Seizure incidence in psychopharmacological clinical trials: An analysis of food and drug administration (fda) summary basis of approval reports. Biol. Psychiatry 2007, 62, 345–354. [Google Scholar] [CrossRef]
- Wu, C.S.; Wang, S.C.; Yeh, I.J.; Liu, S.K. Comparative risk of seizure with use of first- and second-generation antipsychotics in patients with schizophrenia and mood disorders. J. Clin. Psychiatry 2016, 77, e573–e579. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Suzuki, T.; Sasaki, M.; Shiraishi, H.; Koizumi, J. Convulsive seizures in schizophrenic patients induced by zotepine administration. Jpn. J. Psychiatry Neurol. 1992, 46, 161–167. [Google Scholar] [CrossRef]
- Palmgren, K.; Wighton, A.; Reynolds, C.W.; Butler, A.; Tweed, J.A.; Raniwalla, J.; Welch, C.P.; Bratty, J.R. The safety and efficacy of zotepine in the treatment of schizophrenia: Results of a one-year naturalistic clinical trial. Int. J. Psychiatry Clin. Pr. 2000, 4, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y.; Momotani, Y.; Suzuki, E.; Kaku, T. Clinical and eeg studies of zotepine, a thiepine neuroleptic, on schizophrenic patients. Pharmacopsychiatry 1987, 20, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, M.Y.; Rabey, J.M.; Orlov, E.; Korczyn, A.D. Electroencephalographic findings with low-dose clozapine treatment in psychotic parkinsonian patients. Clin. Neuropharmacol. 1996, 19, 81–86. [Google Scholar] [CrossRef]
- Freudenreich, O.; Weiner, R.D.; McEvoy, J.P. Clozapine-induced electroencephalogram changes as a function of clozapine serum levels. Biol. Psychiatry 1997, 42, 132–137. [Google Scholar] [CrossRef]
- Okada, M.; Fukuyama, K.; Shiroyama, T.; Murata, M. A working hypothesis regarding identical pathomechanisms between clinical efficacy and adverse reaction of clozapine via the activation of connexin43. Int. J. Mol. Sci. 2020, 21, 7019. [Google Scholar] [CrossRef]
- Kuo, C.J.; Yang, S.Y.; Liao, Y.T.; Chen, W.J.; Lee, W.C.; Shau, W.Y.; Chang, Y.T.; Tsai, S.Y.; Chen, C.C. Second-generation antipsychotic medications and risk of pneumonia in schizophrenia. Schizophr. Bull. 2013, 39, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackin, P. Cardiac side effects of psychiatric drugs. Hum. Psychopharmacol. 2008, 23, 3–14. [Google Scholar] [CrossRef]
- de Leon, J.; Sanz, E.J.; Noren, G.N.; De Las Cuevas, C. Pneumonia may be more frequent and have more fatal outcomes with clozapine than with other second-generation antipsychotics. World Psychiatry 2020, 19, 120–121. [Google Scholar] [CrossRef] [PubMed]
- De Leon, J.; Sanz, E.J.; De Las Cuevas, C. Data from the world health organization's pharmacovigilance database supports the prominent role of pneumonia in mortality associated with clozapine adverse drug reactions. Schizophr. Bull. 2020, 46, 1–3. [Google Scholar] [CrossRef]
- Fontes, M.S.; van Veen, T.A.; de Bakker, J.M.; van Rijen, H.V. Functional consequences of abnormal cx43 expression in the heart. Biochim. Biophys. Acta 2012, 1818, 2020–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, X.; Li, X.Z.; Fan, Z.R.; Wang, A.; Zhang, H.C.; Zhang, L.; Li, L.; Si, J.Q.; Ma, K.T. Increased expression and functionality of the gap junction in peripheral blood lymphocytes is associated with hypertension-mediated inflammation in spontaneously hypertensive rats. Cell. Mol. Biol. Lett. 2018, 23, 40. [Google Scholar] [CrossRef] [Green Version]
- Fukuyama, K.; Okada, M. Age-dependent and sleep/seizure-induced pathomechanisms of autosomal dominant sleep-related hypermotor epilepsy. Int. J. Mol. Sci. 2020, 21, 8142. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.V.; Contreras, J.E.; Bukauskas, F.F.; Saez, J.C. New roles for astrocytes: Gap junction hemichannels have something to communicate. Trends Neurosci. 2003, 26, 610–617. [Google Scholar] [CrossRef] [Green Version]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Okada, M. Can rodent models elucidate the pathomechanisms of genetic epilepsy? Br. J. Pharmacol. 2021, in press. [Google Scholar] [CrossRef]
- Okada, M.; Oka, T.; Nakamoto, M.; Fukuyama, K.; Shiroyama, T. Astroglial connexin43 as a potential target for a mood stabiliser. Int. J. Mol. Sci. 2020, 22, 339. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Rodrigues, T.M.; Martins-Marques, T.; Morel, S.; Kwak, B.R.; Girao, H. Role of connexin 43 in different forms of intercellular communication-gap junctions, extracellular vesicles and tunnelling nanotubes. J. Cell Sci. 2017, 130, 3619–3630. [Google Scholar] [CrossRef] [Green Version]
- Jeanson, T.; Pondaven, A.; Ezan, P.; Mouthon, F.; Charveriat, M.; Giaume, C. Antidepressants impact connexin 43 channel functions in astrocytes. Front. Cell. Neurosci. 2015, 9, 495. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Gangoso, E.; Yi, C.; Jeanson, T.; Kandelman, S.; Mantz, J.; Giaume, C. General anesthetics have differential inhibitory effects on gap junction channels and hemichannels in astrocytes and neurons. Glia 2016, 64, 524–536. [Google Scholar] [CrossRef]
- Walrave, L.; Vinken, M.; Leybaert, L.; Smolders, I. Astrocytic connexin43 channels as candidate targets in epilepsy treatment. Biomolecules 2020, 10, 1578. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, K.; Okada, M. Effects of atypical antipsychotics, clozapine, quetiapine and brexpiprazole on astroglial transmission associated with connexin43. Int. J. Mol. Sci. 2021, 22, 5623. [Google Scholar] [CrossRef]
- Fukuyama, K.; Okubo, R.; Murata, M.; Shiroyama, T.; Okada, M. Activation of astroglial connexin is involved in concentration-dependent double-edged sword clinical action of clozapine. Cells 2020, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Fukuyama, K.; Shiroyama, T.; Ueda, Y. Brivaracetam prevents astroglial l-glutamate release associated with hemichannel through modulation of synaptic vesicle protein. Biomed. Pharmacother. 2021, 138, 111462. [Google Scholar] [CrossRef]
- Fukuyama, K.; Ueda, Y.; Okada, M. Effects of carbamazepine, lacosamide and zonisamide on gliotransmitter release associated with activated astroglial hemichannels. Pharmaceuticals 2020, 13, 117. [Google Scholar] [CrossRef] [PubMed]
- Shiroyama, T.; Fukuyama, K.; Okada, M. Distinct effects of escitalopram and vortioxetine on astroglial l-glutamate release associated with connexin43. Int. J. Mol. Sci. 2021, 22, 10013. [Google Scholar] [CrossRef]
- Quesseveur, G.; Gardier, A.M.; Guiard, B.P. The monoaminergic tripartite synapse: A putative target for currently available antidepressant drugs. Curr. Drug Targets 2013, 14, 1277–1294. [Google Scholar] [CrossRef] [PubMed]
- Kikuoka, R.; Miyazaki, I.; Kubota, N.; Maeda, M.; Kagawa, D.; Moriyama, M.; Sato, A.; Murakami, S.; Kitamura, Y.; Sendo, T.; et al. Mirtazapine exerts astrocyte-mediated dopaminergic neuroprotection. Sci. Rep. 2020, 10, 20698. [Google Scholar] [CrossRef] [PubMed]
- Stroth, N.; Svenningsson, P. S100b interacts with the serotonin 5-ht7 receptor to regulate a depressive-like behavior. Eur. Neuropsychopharmacol. 2015, 25, 2372–2380. [Google Scholar] [CrossRef]
- Okubo, R.; Hasegawa, T.; Fukuyama, K.; Shiroyama, T.; Okada, M. Current limitations and candidate potential of 5-ht7 receptor antagonism in psychiatric pharmacotherapy. Front. Psychiatry 2021, 12, 623684. [Google Scholar] [CrossRef] [PubMed]
- Schoretsanitis, G.; Paulzen, M.; Unterecker, S.; Schwarz, M.; Conca, A.; Zernig, G.; Grunder, G.; Haen, E.; Baumann, P.; Bergemann, N.; et al. Tdm in psychiatry and neurology: A comprehensive summary of the consensus guidelines for therapeutic drug monitoring in neuropsychopharmacology, update 2017; a tool for clinicians. World Biol. Psychiatry 2018, 19, 162–174. [Google Scholar] [CrossRef]
- Hiemke, C.; Bergemann, N.; Clement, H.W.; Conca, A.; Deckert, J.; Domschke, K.; Eckermann, G.; Egberts, K.; Gerlach, M.; Greiner, C.; et al. Consensus guidelines for therapeutic drug monitoring in neuropsychopharmacology: Update 2017. Pharmacopsychiatry 2018, 51, 9–62. [Google Scholar]
- Yamamura, S.; Ohoyama, K.; Hamaguchi, T.; Nakagawa, M.; Suzuki, D.; Matsumoto, T.; Motomura, E.; Tanii, H.; Shiroyama, T.; Okada, M. Effects of zotepine on extracellular levels of monoamine, gaba and glutamate in rat prefrontal cortex. Br. J. Pharmacol. 2009, 157, 656–665. [Google Scholar] [CrossRef]
- Yamamura, S.; Ohoyama, K.; Hamaguchi, T.; Kashimoto, K.; Nakagawa, M.; Kanehara, S.; Suzuki, D.; Matsumoto, T.; Motomura, E.; Shiroyama, T.; et al. Effects of quetiapine on monoamine, gaba, and glutamate release in rat prefrontal cortex. Psychopharmacology 2009, 206, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Fukuyama, K.; Shiroyama, T.; Ueda, Y. Lurasidone inhibits nmda antagonist-induced functional abnormality of thalamocortical glutamatergic transmission via 5-ht7 receptor blockade. Br. J. Pharmacol. 2019, 176, 4002–4018. [Google Scholar] [CrossRef]
- Pratt, J.A.; Winchester, C.; Egerton, A.; Cochran, S.M.; Morris, B.J. Modelling prefrontal cortex deficits in schizophrenia: Implications for treatment. Br. J. Pharmacol. 2008, 153, S465–S470. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, H.; von Bardeleben, U.; Holsboer, F.; Benkert, O. Zotepine versus perazine in patients with paranoid schizophrenia: A double-blind controlled trial of its effectiveness. Fortschr. Neurol. -Psychiatr. 1991, 59, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Sotnikova, T.D.; Marion, S.; Lefkowitz, R.J.; Gainetdinov, R.R.; Caron, M.G. An akt/beta-arrestin 2/pp2a signaling complex mediates dopaminergic neurotransmission and behavior. Cell 2005, 122, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu, J.M.; Sotnikova, T.D.; Yao, W.D.; Kockeritz, L.; Woodgett, J.R.; Gainetdinov, R.R.; Caron, M.G. Lithium antagonizes dopamine-dependent behaviors mediated by an akt/glycogen synthase kinase 3 signaling cascade. Proc. Natl. Acad. Sci. USA 2004, 101, 5099–5104. [Google Scholar] [CrossRef] [Green Version]
- Johnson-Farley, N.N.; Kertesy, S.B.; Dubyak, G.R.; Cowen, D.S. Enhanced activation of akt and extracellular-regulated kinase pathways by simultaneous occupancy of gq-coupled 5-ht2a receptors and gs-coupled 5-ht7a receptors in pc12 cells. J. Neurochem. 2005, 92, 72–82. [Google Scholar] [CrossRef]
- Daniele, S.; Zappelli, E.; Martini, C. Trazodone regulates neurotrophic/growth factors, mitogen-activated protein kinases and lactate release in human primary astrocytes. J. Neuroinflammation 2015, 12, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxishima, M.; Shiga, T.; Shutoh, F.; Hamada, S.; Maeshima, T.; Okado, N. Serotonin 2a receptor-like immunoreactivity is detected in astrocytes but not in oligodendrocytes of rat spinal cord. Brain Res. 2001, 889, 270–273. [Google Scholar] [CrossRef]
- Harris, K.L.; Mason, S.L.; Vallin, B.; Barker, R.A. Reduced expression of dopamine d2 receptors on astrocytes in r6/1 hd mice and hd post-mortem tissue. Neurosci. Lett. 2021, 136289, in press. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.S.; Sato, W.; Ataka, K.; Yagisawa, K.; Omori, Y.; Kanbayashi, T.; Shimizu, T. Clozapine-induced seizures, electroencephalography abnormalities, and clinical responses in japanese patients with schizophrenia. Neuropsychiatr. Dis. Treat. 2014, 10, 1973–1978. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.M.; Park, S.H. Seizure associated with clozapine: Incidence, etiology, and management. CNS Drugs 2015, 29, 101–111. [Google Scholar] [CrossRef]
- Ali, M.; Das, S. Zotepine-induced convulsion at a low dose in a case of paranoid schizophrenia. Psychiatry Clin. Neurosci. 2018, 72, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khairkar, P.; Gupta, N.; Varma, S.K. Zotepine-induced convulsive seizures in a chronic case of treatment resistant paranoid schizophrenia. Indian J. Pharm. 2013, 45, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, K.; Fukuzawa, M.; Ruri, O.; Okada, M. Upregulated connexin 43 induced by loss-of-functional s284l-mutant alpha4 subunit of nicotinic ach receptor contributes to pathomechanisms of autosomal dominant sleep-related hypermotor epilepsy. Pharmaceuticals 2020, 13, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuyama, K.; Fukuzawa, M.; Okada, M. Upregulated and hyperactivated thalamic connexin 43 plays important roles in pathomechanisms of cognitive impairment and seizure of autosomal dominant sleep-related hypermotor epilepsy with s284l-mutant α4 subunit of nicotinic ach receptor. Pharmaceuticals 2020, 13, 99. [Google Scholar] [CrossRef]
- Fukuyama, K.; Fukuzawa, M.; Shiroyama, T.; Okada, M. Pathogenesis and pathophysiology of autosomal dominant sleep-related hypermotor epilepsy with s284l-mutant alpha4 subunit of nicotinic ach receptor. Br. J. Pharmacol. 2020, 177, 2143–2162. [Google Scholar] [CrossRef]
- Ernst, C.; Nagy, C.; Kim, S.; Yang, J.P.; Deng, X.; Hellstrom, I.C.; Choi, K.H.; Gershenfeld, H.; Meaney, M.J.; Turecki, G. Dysfunction of astrocyte connexins 30 and 43 in dorsal lateral prefrontal cortex of suicide completers. Biol. Psychiatry 2011, 70, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Bernard, R.; Kerman, I.A.; Thompson, R.C.; Jones, E.G.; Bunney, W.E.; Barchas, J.D.; Schatzberg, A.F.; Myers, R.M.; Akil, H.; Watson, S.J. Altered expression of glutamate signaling, growth factor, and glia genes in the locus coeruleus of patients with major depression. Mol. Psychiatry 2011, 16, 634–646. [Google Scholar] [CrossRef] [Green Version]
- Miguel-Hidalgo, J.J.; Wilson, B.A.; Hussain, S.; Meshram, A.; Rajkowska, G.; Stockmeier, C.A. Reduced connexin 43 immunolabeling in the orbitofrontal cortex in alcohol dependence and depression. J. Psychiatr. Res. 2014, 55, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, C.; Torres-Platas, S.G.; Mechawar, N.; Turecki, G. Repression of astrocytic connexins in cortical and subcortical brain regions and prefrontal enrichment of h3k9me3 in depression and suicide. Int. J. Neuropsychopharmacol. 2017, 20, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Nagy, C.; Suderman, M.; Yang, J.; Szyf, M.; Mechawar, N.; Ernst, C.; Turecki, G. Astrocytic abnormalities and global DNA methylation patterns in depression and suicide. Mol. Psychiatry 2015, 20, 320–328. [Google Scholar] [CrossRef]
- Rajkowska, G.; Selemon, L.D.; Goldman-Rakic, P.S. Neuronal and glial somal size in the prefrontal cortex: A postmortem morphometric study of schizophrenia and huntington disease. Arch. Gen. Psychiatry 1998, 55, 215–224. [Google Scholar] [CrossRef]
- Selemon, L.D.; Rajkowska, G.; Goldman-Rakic, P.S. Elevated neuronal density in prefrontal area 46 in brains from schizophrenic patients: Application of a three-dimensional, stereologic counting method. J. Comp. Neurol. 1998, 392, 402–412. [Google Scholar] [CrossRef]
- Sun, J.-D.; Liu, Y.; Yuan, Y.-H.; Li, J.; Chen, N.-H. Gap junction dysfunction in the prefrontal cortex induces depressive-like behaviors in rats. Neuropsychopharmacology 2012, 37, 1305–1320. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, S.H.; Folsom, T.D.; Reutiman, T.J.; Pandian, T.; Braun, N.N.; Haug, K. Chronic psychotropic drug treatment causes differential expression of connexin 43 and gfap in frontal cortex of rats. Schizophr. Res. 2008, 104, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi, H.; Khaksarian, M.; Joghataei, M.T.; Hassanzadeh, G.; Soleimani, M.; Eftekhari, S.; Soleimani, M.; Mousavizadeh, K.; Hadjighassem, M.R. Fluoxetin upregulates connexin 43 expression in astrocyte. Basic Clin. Neurosci. 2014, 5, 74–79. [Google Scholar] [PubMed]
- Morioka, N.; Suekama, K.; Zhang, F.F.; Kajitani, N.; Hisaoka-Nakashima, K.; Takebayashi, M.; Nakata, Y. Amitriptyline up-regulates connexin43-gap junction in rat cultured cortical astrocytes via activation of the p38 and c-fos/ap-1 signalling pathway. Br. J. Pharmacol. 2014, 171, 2854–2867. [Google Scholar] [CrossRef] [Green Version]
- Curtis, M.J.; Alexander, S.; Cirino, G.; Docherty, J.R.; George, C.H.; Giembycz, M.A.; Hoyer, D.; Insel, P.A.; Izzo, A.A.; Ji, Y.; et al. Experimental design and analysis and their reporting ii: Updated and simplified guidance for authors and peer reviewers. Br. J. Pharmacol. 2018, 175, 987–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Therapeutic-Relevant | Supratherapeutic | Therapeutic-Relevant | Supratherapeutic | ||
|---|---|---|---|---|---|
| Release | Basal | (ACSF) | Activated | (HK-ACSF) | |
| Acute | Non | Increase GAP19(-) | Non | Increase GAP19(+) DEBC(-) | Figure 1, Figure 2 and Figure 3 |
| Subchronic | Non | Increase, GAP19(+) DEBC (-) | Non | Increase GAP19(+) DEBC(+) | Figure 1, Figure 2 and Figure 3 |
| Chronic | Increase | Increase | Figure 8 | ||
| Cx43 Expression | Cytosol | Plasma membrane | |||
| Acute | Non | Figure 7 | |||
| Subchronic | Non | Increase | Non | Increase DEBC (+) | Figure 4, Figure 5 and Figure 6 |
| Chronic | Increase | Figure 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukuyama, K.; Okada, M. Effects of an Atypical Antipsychotic, Zotepine, on Astroglial L-Glutamate Release through Hemichannels: Exploring the Mechanism of Mood-Stabilising Antipsychotic Actions and Antipsychotic-Induced Convulsion. Pharmaceuticals 2021, 14, 1116. https://doi.org/10.3390/ph14111116
Fukuyama K, Okada M. Effects of an Atypical Antipsychotic, Zotepine, on Astroglial L-Glutamate Release through Hemichannels: Exploring the Mechanism of Mood-Stabilising Antipsychotic Actions and Antipsychotic-Induced Convulsion. Pharmaceuticals. 2021; 14(11):1116. https://doi.org/10.3390/ph14111116
Chicago/Turabian StyleFukuyama, Kouji, and Motohiro Okada. 2021. "Effects of an Atypical Antipsychotic, Zotepine, on Astroglial L-Glutamate Release through Hemichannels: Exploring the Mechanism of Mood-Stabilising Antipsychotic Actions and Antipsychotic-Induced Convulsion" Pharmaceuticals 14, no. 11: 1116. https://doi.org/10.3390/ph14111116
APA StyleFukuyama, K., & Okada, M. (2021). Effects of an Atypical Antipsychotic, Zotepine, on Astroglial L-Glutamate Release through Hemichannels: Exploring the Mechanism of Mood-Stabilising Antipsychotic Actions and Antipsychotic-Induced Convulsion. Pharmaceuticals, 14(11), 1116. https://doi.org/10.3390/ph14111116