
Pioglitazone Is a Mild Carrier-Dependent Uncoupler of Oxidative Phosphorylation and a Modulator of Mitochondrial Permeability Transition


 , and
, and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
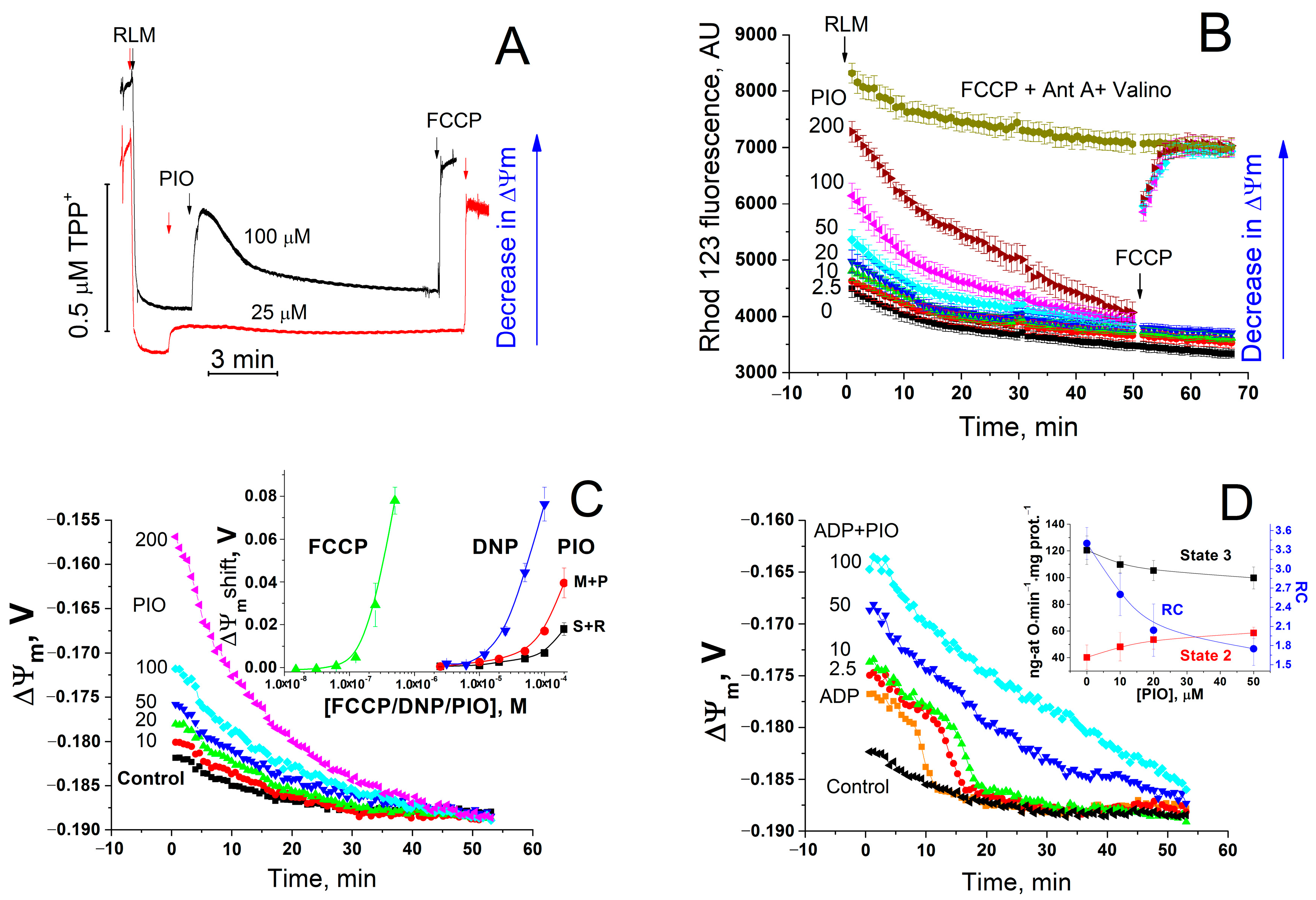
2.1. PIO Acts as a Low-Efficient Uncoupler in Isolated Mitochondria
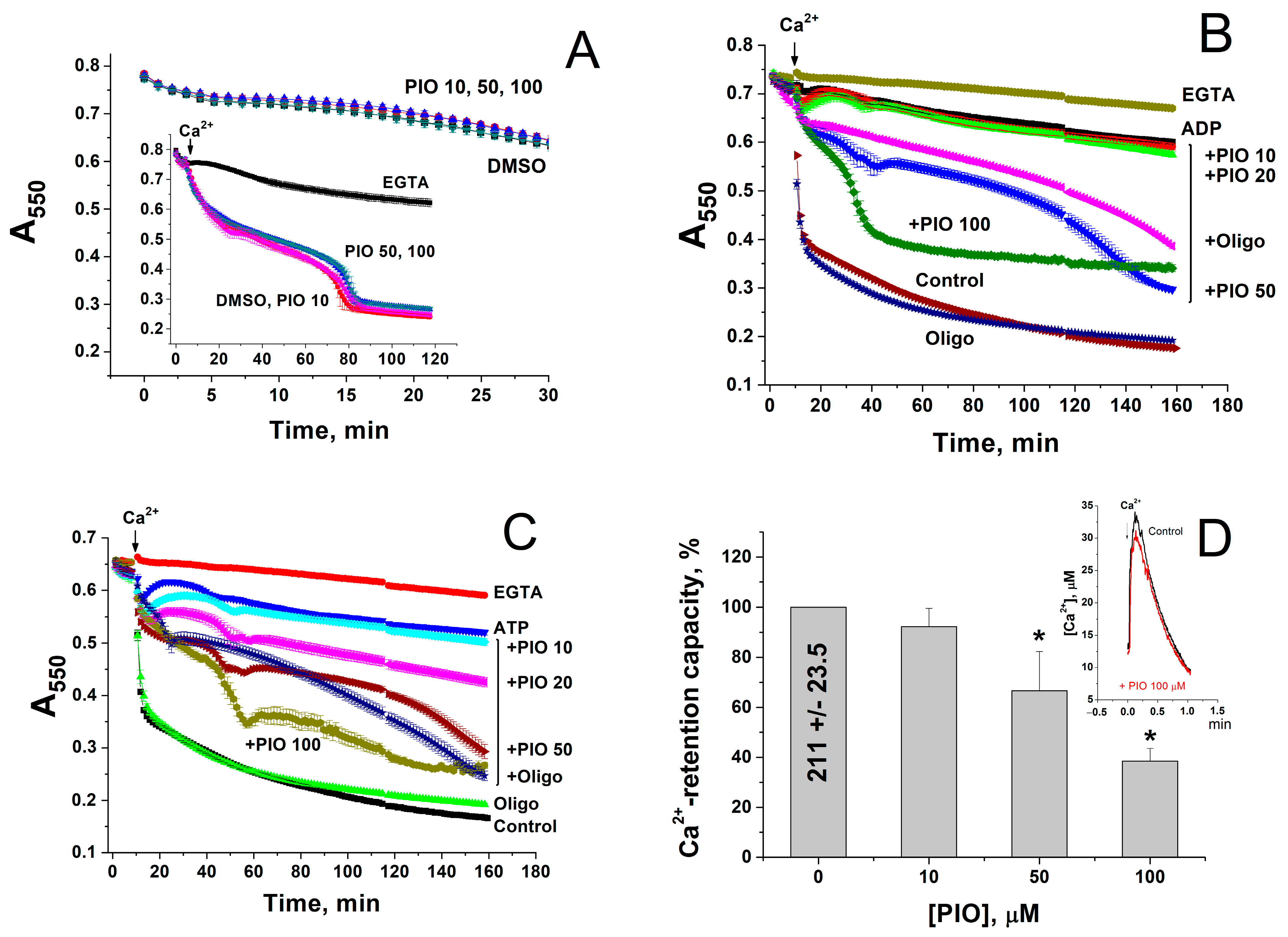
2.2. PIO Modulates the Sensitivity of the Permeability Transition Pore to Regulators
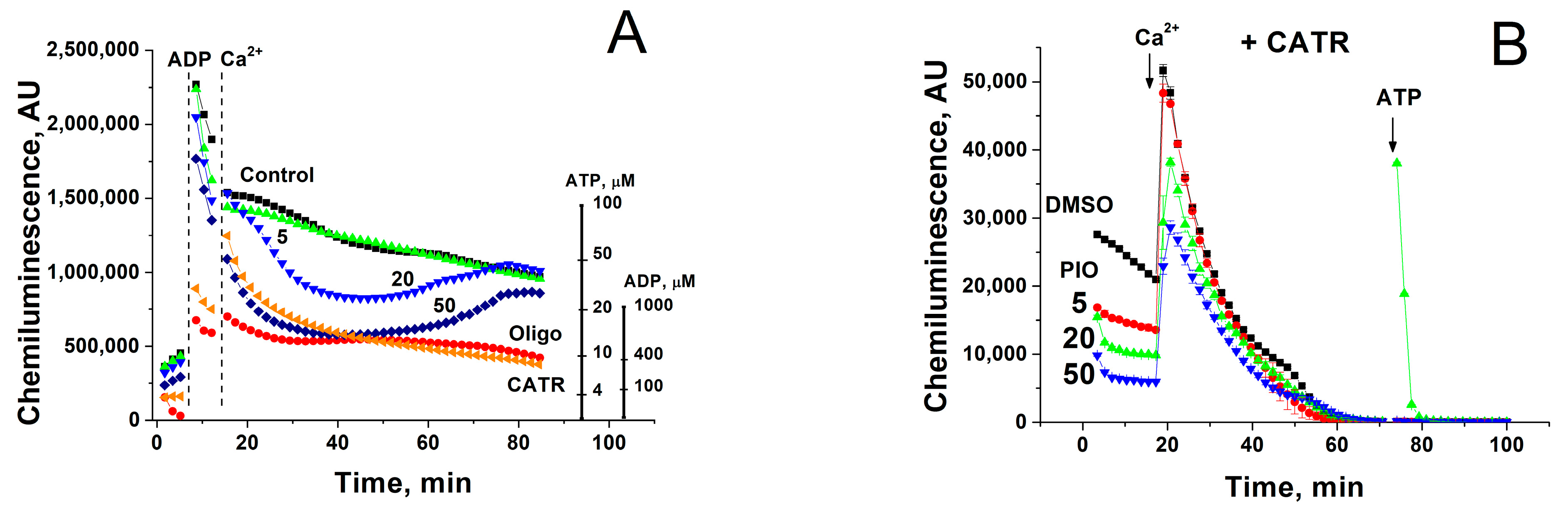
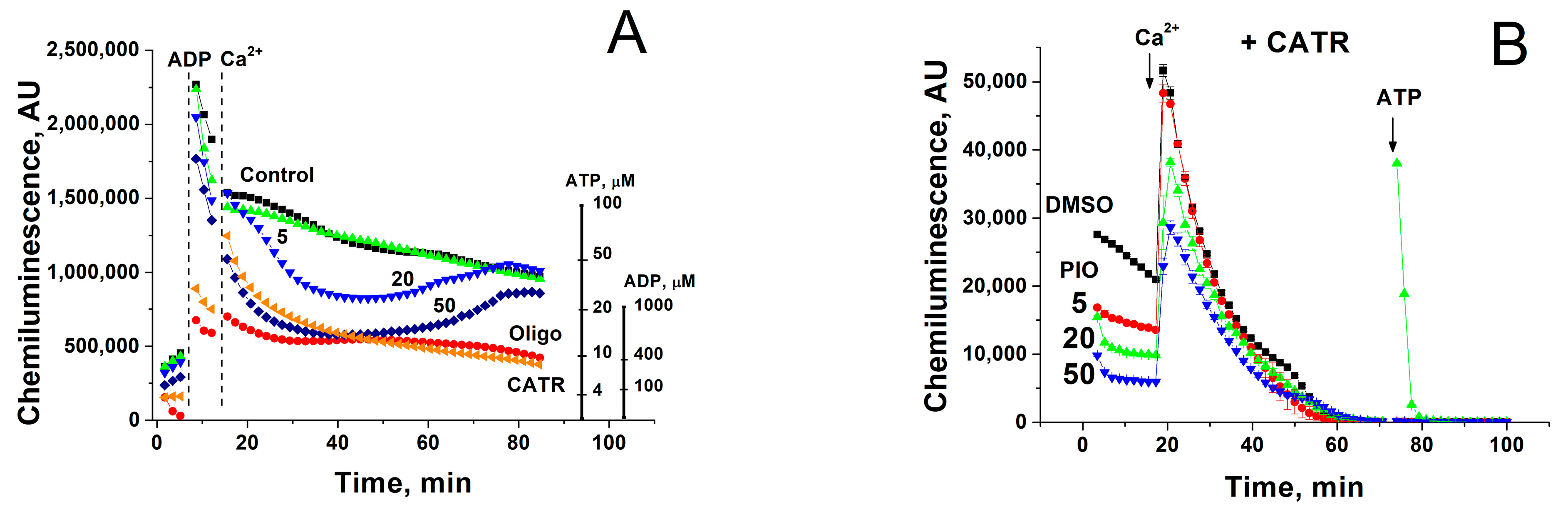
2.3. PIO Decreases the Capability of Mitochondria to Phosphorylate ADP in the Presence of Ca2+
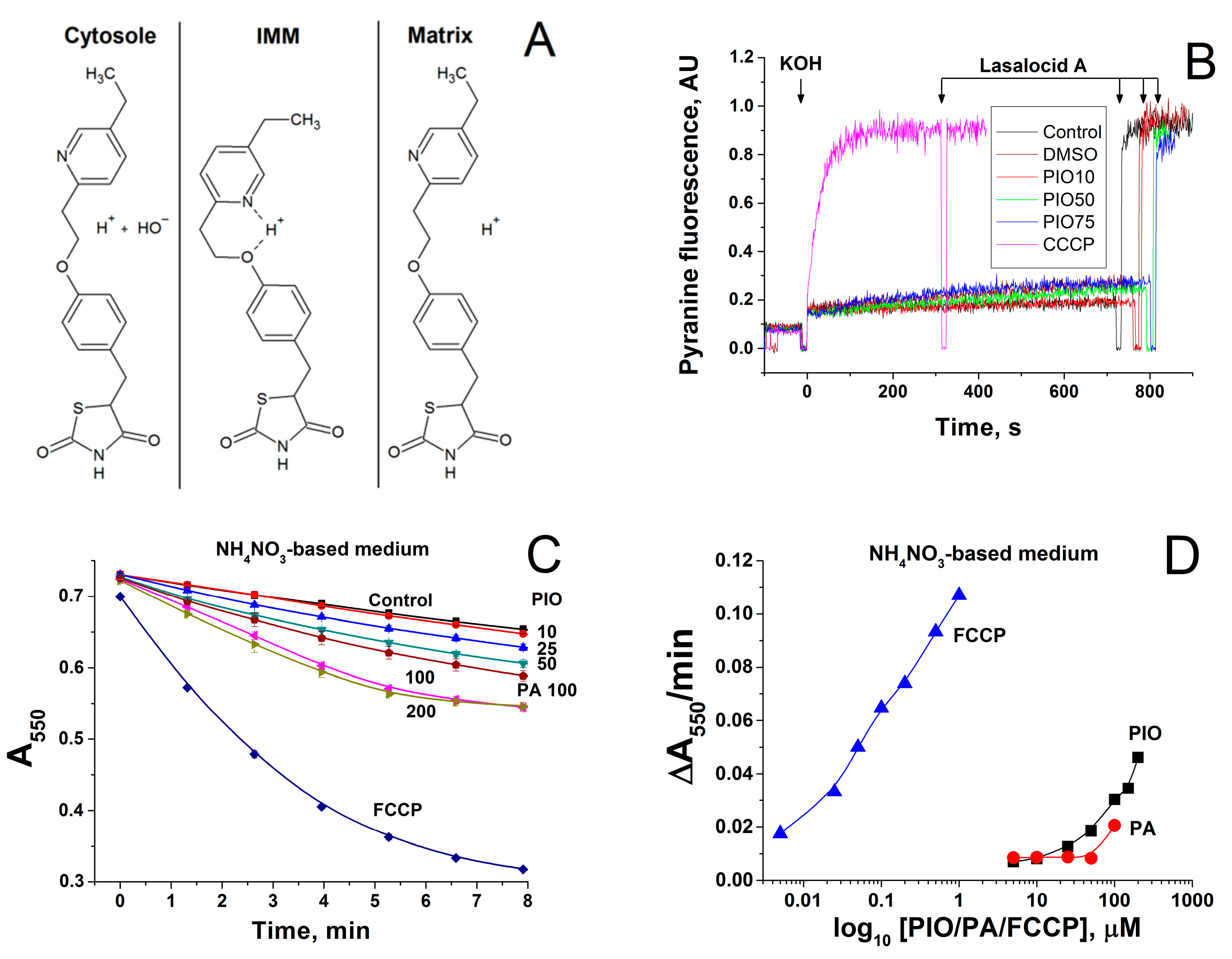
2.4. Effect of PIO on the Permeability of the Inner Mitochondrial and Lecithin Liposomal Membranes to Protons
2.5. Role of Mmitochondrial Carriers in PIO-Dependent Depolarization of Mitochondria
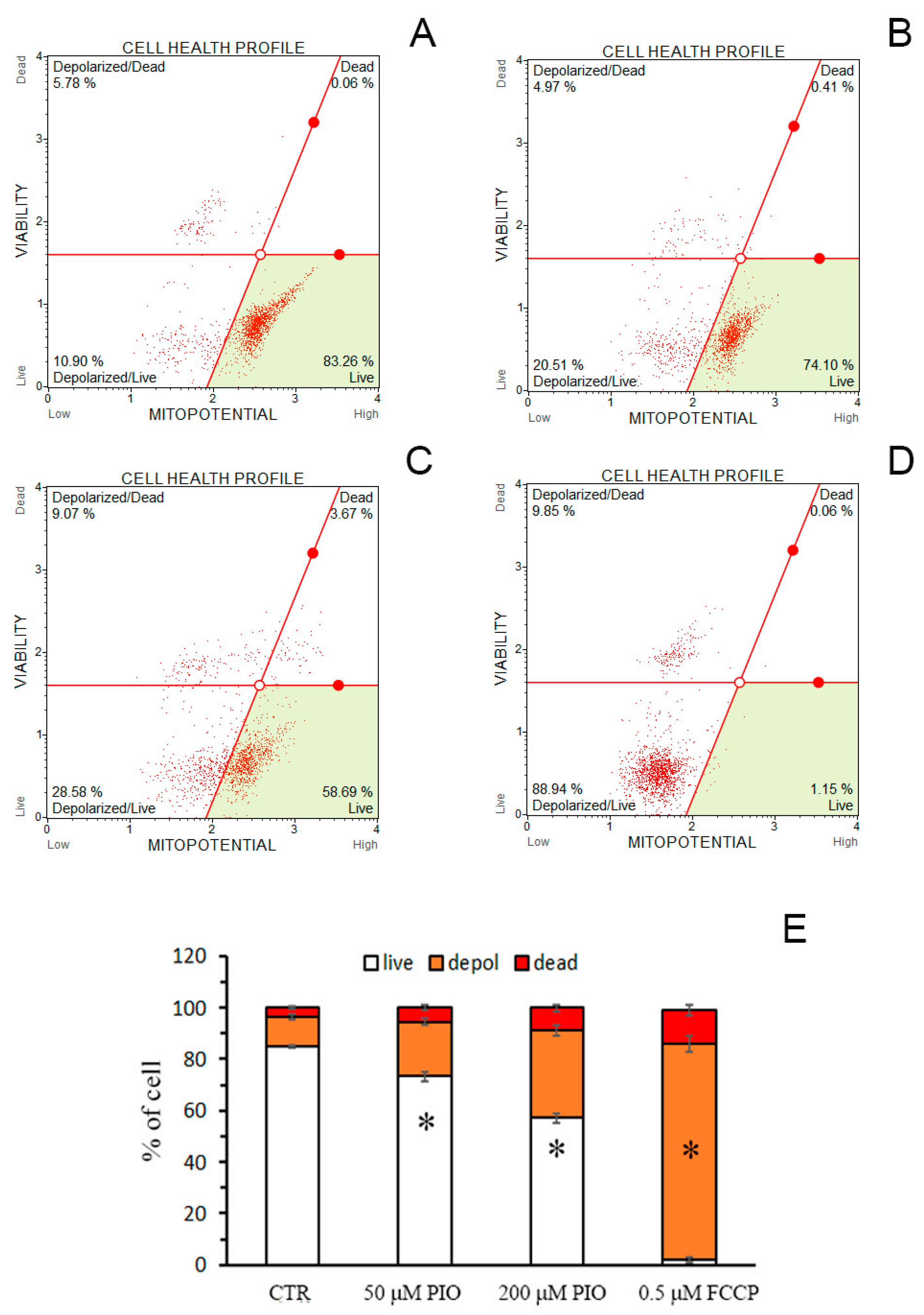
2.6. PIO Causes Mild Mitochondrial Depolarization in Intact Cells
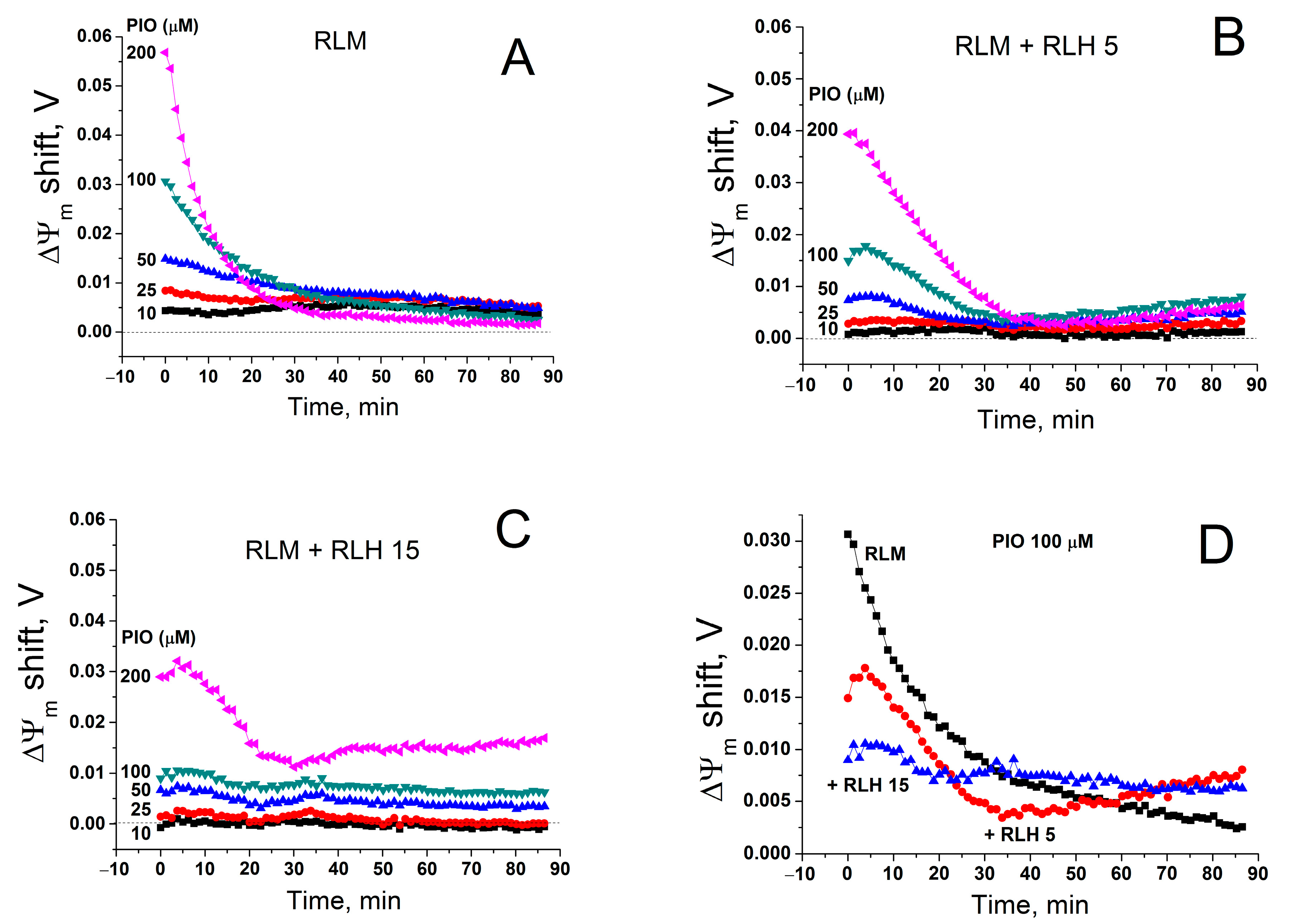
2.7. Effect of PIO on ΔΨm in Isolated Mitochondria and Mitochondria in Nuclei-Free Liver Homogenate
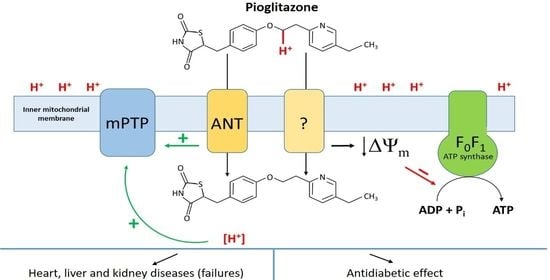
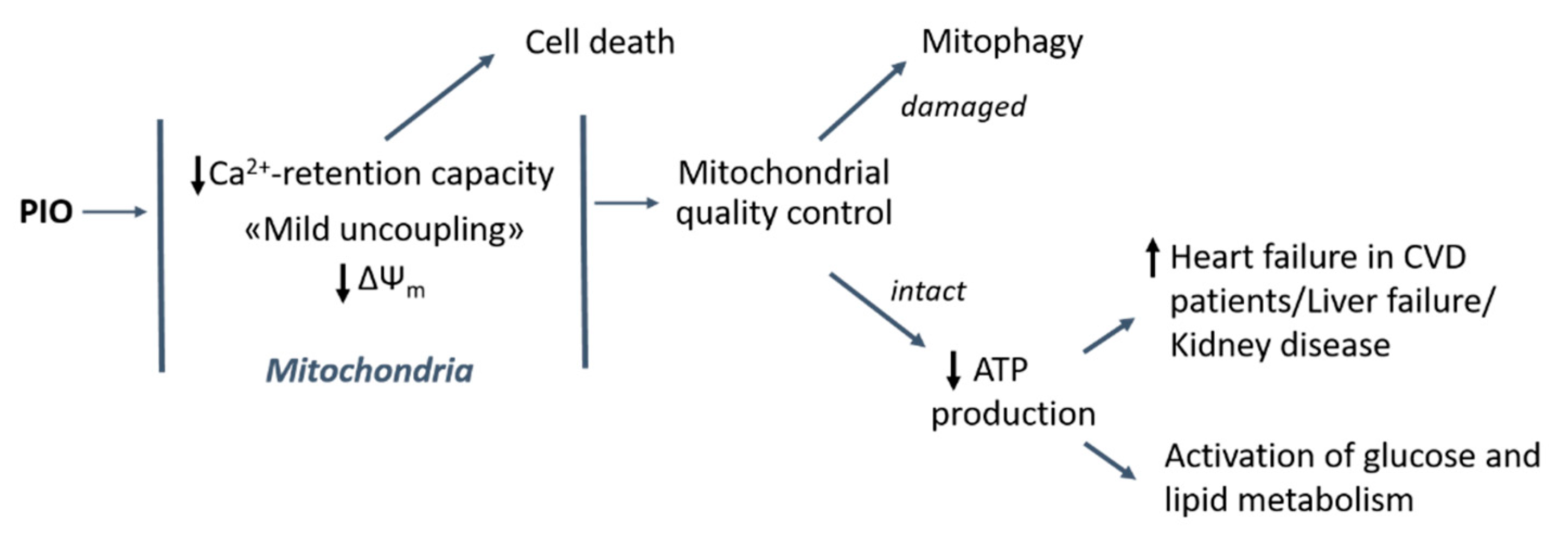
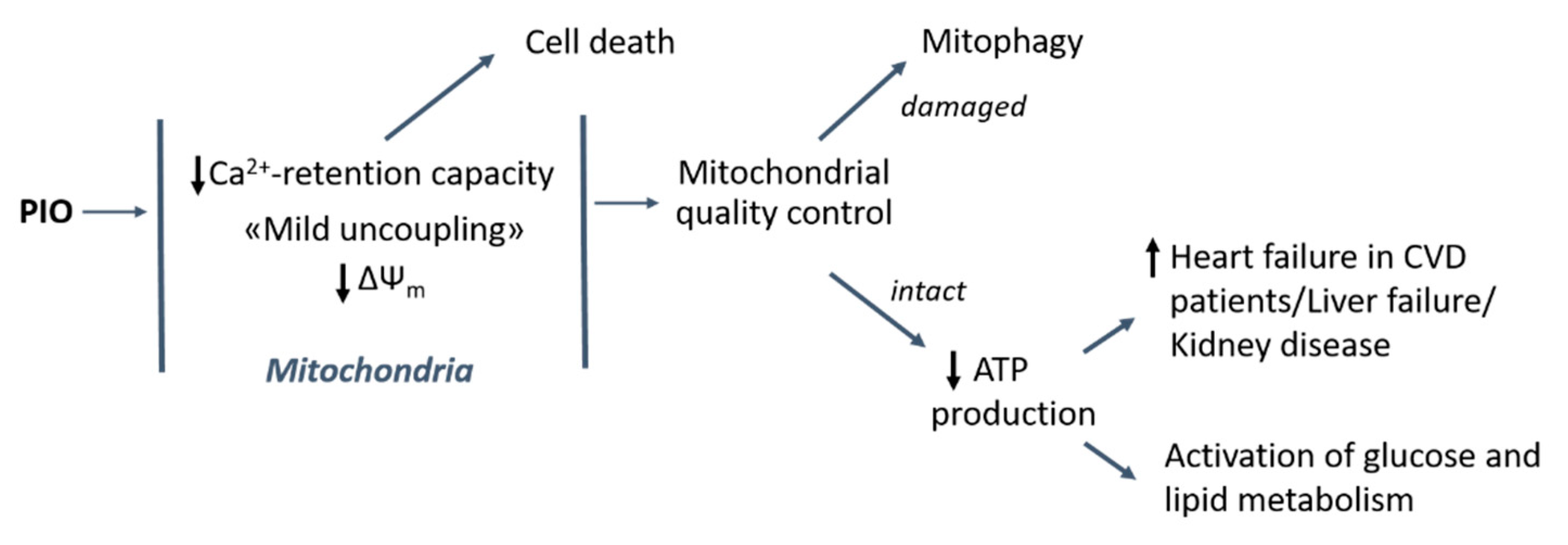
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Isolation of Rat Liver Mitochondria and Preparation of the Cytosolic Fraction of Liver Homogenate
4.3. Measurements of the Oxygen Consumption Rate
4.4. Measurements of ΔΨm in Isolated Mitochondria
4.5. Measurements of ΔΨm in Isolated Thymocytes
4.6. Recording of the Permeabilization of Mitochondrial Membranes
4.7. Ca2+-Retention Capacity of Mitochondria
4.8. Measurement of ATP in Mitochondrial Suspension
4.9. Assessment of the Protonophoric Properties of PIO Using Pyranine-Loaded Vesicles
4.10. Assessment of PIO-Dependent Proton Transport in Mitochondria
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wright, M.B.; Bortolini, M.; Tadayyon, M.; Bopst, M. Minireview: Challenges and opportunities in development of PPAR agonists. Mol. Endocrinol. 2014, 28, 1756–1768. [Google Scholar] [CrossRef] [Green Version]
- Alam, F.; Islam, M.A.; Mohamed, M.; Ahmad, I.; Kamal, M.A.; Donnelly, R.; Idris, I.; Gan, S.H. Efficacy and safety of pioglitazone monotherapy in type 2 diabetes mellitus: A systematic review and meta-analysis of randomised controlled trials. Sci. Rep. 2019, 9, 5389. [Google Scholar] [CrossRef] [Green Version]
- Albert, S.G.; Wood, E.M. Meta-analysis of trials in non-alcoholic fatty liver disease with therapeutic interventions for metabolic syndrome. Diabetes Metab. Syndr. 2021, 15, 102232. [Google Scholar] [CrossRef]
- Lian, J.; Fu, J. Pioglitazone for NAFLD Patients With Prediabetes or Type 2 Diabetes Mellitus: A Meta-Analysis. Front. Endocrinol. 2021, 12, 615409. [Google Scholar] [CrossRef]
- Burns, D.K.; Alexander, R.C.; Welsh-Bohmer, K.A.; Culp, M.; Chiang, C.; O’Neil, J.; Evans, R.M.; Harrigan, P.; Plassman, B.L.; Burke, J.R.; et al. Safety and efficacy of pioglitazone for the delay of cognitive impairment in people at risk of Alzheimer’s disease (TOMMORROW): A prognostic biomarker study and a phase 3, randomised, double-blind, placebo-controlled trial. Lancet Neurol. 2021, 20, 537–547. [Google Scholar] [CrossRef]
- Hang, X.; Zhang, Y.; Li, J.; Li, Z.; Zhang, Y.; Ye, X.; Tang, Q.; Sun, W. Comparative Efficacy and Acceptability of Anti-inflammatory Agents on Major Depressive Disorder: A Network Meta-Analysis. Front. Pharmacol. 2021, 12, 691200. [Google Scholar] [CrossRef] [PubMed]
- Della Pepa, G.; Russo, M.; Vitale, M.; Carli, F.; Vetrani, C.; Masulli, M.; Riccardi, G.; Vaccaro, O.; Gastaldelli, A.; Rivellese, A.A.; et al. Pioglitazone even at low dosage improves NAFLD in type 2 diabetes: Clinical and pathophysiological insights from a subgroup of the TOSCA. IT randomised trial. Diabetes Res. Clin. Pract. 2021, 178, 108984. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.; Zhou, Z.; Xi, X.; Huang, R.; Hu, H. Pioglitazone Ameliorates Renal Ischemia-Reperfusion Injury via Inhibition of NF-κB Activation and Inflammation in Rats. Front. Physiol. 2021, 12, 707344. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Moral, I.; Lima Ferreira, B.; de Vos, A.F.; van der Poll, T. Post-treatment with the PPAR-γ agonist pioglitazone inhibits inflammation and bacterial growth during Klebsiella pneumonia. Respir Res. 2021, 22, 230. [Google Scholar] [CrossRef]
- Hassan, F.E.; Sakr, H.I.; Mohie, P.M.; Suliman, H.S.; Mohamed, A.S.; Attia, M.H.; Eid, D.M. Pioglitazone improves skeletal muscle functions in reserpine-induced fibromyalgia rat model. Ann. Med. 2021, 53, 1032–1040. [Google Scholar] [CrossRef]
- Yeligar, S.M.; Mehta, A.J.; Harris, F.L.; Brown, L.A.S.; Hart, C.M. Pioglitazone Reverses Alcohol-Induced Alveolar Macrophage Phagocytic Dysfunction. J. Immunol. 2021, 207, 483–492. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Alblihed, M.; Guerreiro, S.G.; Cruz-Martins, N.; Batiha, G.E. COVID-19 in Relation to Hyperglycemia and Diabetes Mellitus. Front. Cardiovasc. Med. 2021, 8, 644095. [Google Scholar] [CrossRef] [PubMed]
- Abdelhafez, A.T.; Gomaa, A.M.S.; Ahmed, A.M.; Sayed, M.M.; Ahmed, M.A. Pioglitazone and/or irbesartan ameliorate COPD-induced endothelial dysfunction in side stream cigarette smoke-exposed mice model. Life Sci. 2021, 280, 119706. [Google Scholar] [CrossRef]
- Gouveia-Eufrasio, L.; Ribeiro, N.Q.; Alves Santos, J.R.; Carvalho da Costa, M.; Peres Emídio, E.C.; Cota de Freitas, G.J.; Fonseca do Carmo, P.H.; Miranda, B.A.; Dornelas de Oliveira, J.C.M.; Vitorino da Silva, L.M.; et al. Randomized, phase 1/2, double-blind pioglitazone repositioning trial combined with antifungals for the treatment of cryptococcal meningitis—PIO study. Contemp. Clin. Trials Commun. 2021, 22, 100745. [Google Scholar] [CrossRef] [PubMed]
- Weil, Z.M.; Karelina, K.; Whitehead, B.; Velazquez-Cruz, R.; Oliverio, R.; Pinti, M.; Nwafor, D.C.; Nicholson, S.; Fitzgerald, J.A.; Hollander, J.; et al. Mild traumatic brain injury increases vulnerability to cerebral ischemia in mice. Exp. Neurol. 2021, 342, 113765. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Tripathy, D.; Schwenke, D.C.; Banerji, M.; Bray, G.A.; Buchanan, T.A.; Clement, S.C.; Henry, R.R.; Hodis, H.N.; Kitabchi, A.E.; et al. Pioglitazone for diabetes prevention in impaired glucose tolerance. N. Engl. J. Med. 2011, 364, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- de Jong, M.; van der Worp, H.B.; van der Graaf, Y.; Visseren, F.L.J.; Westerink, J. Pioglitazone and the secondary prevention of cardiovascular disease. A meta-analysis of randomized-controlled trials. Cardiovasc. Diabetol. 2017, 16, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polyzos, S.A.; Kechagias, S.; Tsochatzis, E.A. Review article: Non-alcoholic fatty liver disease and cardiovascular diseases: Associations and treatment considerations. Aliment. Pharmacol. Ther. 2021, 54, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Nesto, R.W.; Bell, D.; Bonow, R.O.; Fonseca, V.; Grundy, S.M.; Horton, E.S.; Le Winter, M.; Porte, D.; Semenkovich, C.F.; Smith, S.; et al. Thiazolidinedione use, fluid retention, and congestive heart failure: A consensus statement from the American Heart Association and American Diabetes Association. Diabetes Care 2004, 27, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.Y.; Hsiao, P.J.; Yang, Y.H.; Lin, K.D.; Shin, S.J. The association of pioglitazone and urinary tract disease in type 2 diabetic Taiwanese: Bladder cancer and chronic kidney disease. PLoS ONE 2014, 9, e85479. [Google Scholar] [CrossRef]
- McEvoy, G.K. AHFS Drug Information, 59th ed.; American Society of Health-System Pharmacists: Bethesda, MD, USA, 2017; p. 3299. [Google Scholar]
- Mehtälä, J.; Khanfir, H.; Bennett, D.; Ye, Y.; Korhonen, P.; Hoti, F. Pioglitazone use and risk of bladder cancer: A systematic literature review and meta-analysis of observational studies. Diabetol. Int. 2019, 10, 24–36. [Google Scholar] [CrossRef]
- Pérez-Ortiz, J.M.; Tranque, P.; Burgos, M.; Vaquero, C.F.; Llopis, J. Glitazones induce astroglioma cell death by releasing reactive oxygen species from mitochondria: Modulation of cytotoxicity by nitric oxide. Mol. Pharmacol. 2007, 72, 407–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassef, M.A.E.; Tork, O.M.; Rashed, L.A.; Ibrahim, W.; Morsi, H.; Rabie, D.M.M. Mitochondrial dysfunction in diabetic cardiomyopathy: Effect of mesenchymal stem cell with PPAR-γ agonist or exendin-4. Exp. Clin. Endocrinol. Diabetes 2018, 126, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Kiran, A.V.V.V.R.; Kumari, G.K.; Krishnamurthy, P.T. Preliminary evaluation of anticancer efficacy of pioglitazone combined with celecoxib for the treatment of non-small cell lung cancer. Investig. New Drugs 2021. [Google Scholar] [CrossRef] [PubMed]
- Kumari, G.K.; Kiran, A.V.V.V.R.; Krishnamurthy, P.T. Preliminary evaluation on the beneficial effects of pioglitazone in the treatment of endometrial cancer. Med. Oncol. 2021, 38, 71. [Google Scholar] [CrossRef]
- Nesti, L.; Tricò, D.; Mengozzi, A.; Natali, A. Rethinking pioglitazone as a cardioprotective agent: A new perspective on an overlooked drug. Cardiovasc. Diabetol. 2021, 18, 109. [Google Scholar] [CrossRef] [PubMed]
- Butterick, T.A.; Stone, L.H.; Duffy, C.; Holley, C.; Cabrera, J.A.; Crampton, M.; Ward, H.B.; Kelly, R.F.; McFalls, E.O. Pioglitazone increases PGC1-α signaling within chronically ischemic myocardium. Basic Res. Cardiol. 2016, 111, 37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, X.; Meng, L.; Gong, M.; Li, J.; Shi, W.; Qiu, J.; Yang, Y.; Zhao, J.; Suo, Y.; et al. Pioglitazone Inhibits Diabetes-Induced Atrial Mitochondrial Oxidative Stress and Improves Mitochondrial Biogenesis, Dynamics, and Function Through the PPAR-γ/PGC-1α Signaling Pathway. Front. Pharmacol. 2021, 12, 658362. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.Y.; Perez-Polo, J.R.; Birnbaum, Y. Oral glyburide, but not glimepiride, blocks the infarct-size limiting effects of pioglitazone. Cardiovasc. Drugs Ther. 2008, 22, 429–436. [Google Scholar] [CrossRef]
- Wynne, A.M.; Mocanu, M.M.; Yellon, D.M. Pioglitazone mimics preconditioning in the isolated perfused rat heart: A role for the prosurvival kinases PI3K and P42/44MAPK. J. Cardiovasc. Pharmacol. 2005, 46, 817–822. [Google Scholar] [CrossRef]
- Ye, Y.; Lin, Y.; Atar, S.; Huang, M.H.; Perez-Polo, J.R.; Uretsky, B.F.; Birnbaum, Y. Myocardial protection by pioglita6zone, atorvastatin, and their combination: Mechanisms and possible interactions. Am. J. Physiol. 2006, 291, 1158–1169. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lang, M.J.; Mao, X.B.; Tian, L.; Feng, Y.B. Antiapoptosis and mitochondrial effect of pioglitazone preconditioning in the ischemic/reperfused heart of rat. Cardiovasc. Drugs Ther. 2008, 22, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Jarrar, Y.B.; Jarrar, Q.; Abaalkhail, S.J.; Kalloush, H.M.; Naser, W.; Zihlif, M.; Shhab, M.A.; Madani, A.E.; Jamous, Y.; Lee, S.J. Molecular toxicological alterations in the mouse hearts induced by sub-chronic thiazolidinedione drugs administration. Fundam Clin Pharmacol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.F.; Whitaker-Menezes, D.; Howell, A.; Sotgia, F.; Lisanti, M.P. Mitochondrial biogenesis in epithelial cancer cells promotes breast cancer tumor growth and confers autophagy resistance. Cell Cycle 2012, 11, 4174–4180. [Google Scholar] [CrossRef] [Green Version]
- Rezaiean Mehrabadi, A.; Jamshidzadeh, A.; Rashedinia, M.; Niknahad, H. Study of the effects of ATP suppliers and thiol reductants on toxicity of pioglitazone in isolated rat liver mitochondria. Iran. J. Pharm. Res. 2015, 14, 825–832. [Google Scholar] [PubMed]
- Woo, M.; Kim, M. Insulin sensitization causes accelerated sinus nodal dysfunction through autophagic dysregulation in hypertensive mice. Transl. Clin. Pharmacol. 2021, 29, 92–106. [Google Scholar] [CrossRef]
- Colca, J.R.; McDonald, W.G.; Waldon, D.J.; Leone, J.W.; Lull, J.M.; Bannow, C.A.; Lund, E.T.; Mathews, W.R. Identification of a novel mitochondrial protein (“mitoNEET”) cross-linked specifically by a thiazolidinedione photoprobe. Am. J. Physiol. 2004, 286, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Mons, C.; Botzanowski, T.; Nikolaev, A.; Hellwig, P.; Cianférani, S.; Lescop, E.; Bouton, C.; Golinelli-Cohen, M.P. The H2O2-resistant Fe-S redox switch mitoNEET acts as a pH sensor to repair stress-damaged Fe-S protein. Biochemistry 2018, 57, 5616–5628. [Google Scholar] [CrossRef] [Green Version]
- Tamir, S.; Paddock, M.L.; Darash-Yahana-Baram, M.; Holt, S.H.; Sohn, Y.S.; Agranat, L.; Michaeli, D.; Stofleth, J.T.; Lipper, C.H.; Morcos, F.; et al. Structure-function analysis of NEET proteins uncovers their role as key regulators of iron and ROS homeostasis in health and disease. Biochim. Biophys. Acta 2015, 1853, 1294–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.; Li, X.; Zhang, X.; Kang, R.; Tang, D. CISD1 inhibits ferroptosis by protection against mitochondrial lipid peroxidation. Biochem. Biophys. Res. Commun. 2016, 478, 838–844. [Google Scholar] [CrossRef]
- Vernay, A.; Marchetti, A.; Sabra, A.; Jauslin, T.N.; Rosselin, M.; Scherer, P.E.; Demaurex, N.; Orci, L.; Cosson, P. MitoNEET-dependent formation of intermitochondrial junctions. Proc. Natl. Acad. Sci. USA 2017, 114, 8277–8282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, S.E.; Murphy, A.N.; Ross, S.A.; van der Geer, P.; Dixon, J.E. MitoNEET is an iron-containing outer mitochondrial membrane protein that regulates oxidative capacity. Proc. Natl. Acad. Sci. USA 2007, 104, 5318–5323. [Google Scholar] [CrossRef] [Green Version]
- Lipper, C.H.; Karmi, O.; Sohn, Y.S.; Darash-Yahana, M.; Lammert, H.; Song, L.; Liu, A.; Mittler, R.; Nechushtai, R.; Onuchic, J.N.; et al. Structure of the human monomeric NEET protein MiNT and its role in regulating iron and reactive oxygen species in cancer cells. Proc. Natl. Acad. Sci. USA 2018, 115, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Sohn, Y.S.; Tamir, S.; Song, L.; Michaeli, D.; Matouk, I.; Conlan, A.R.; Harir, Y.; Holt, S.H.; Shulaev, V.; Paddock, M.L.; et al. NAF-1 and mitoNEET are central to human breast cancer proliferation by maintaining mitochondrial homeostasis and promoting tumor growth. Proc. Natl. Acad. Sci. USA 2013, 110, 14676–14681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusminski, C.M.; Chen, S.; Ye, R.; Sun, K.; Wang, Q.A.; Spurgin, S.B.; Sanders, P.E.; Brozinick, J.T.; Geldenhuys, W.J.; Li, W.H.; et al. MitoNEET-Parkin effects in pancreatic alpha- and beta-cells, cellular survival, and intrainsular cross talk. Diabetes 2016, 65, 1534–1555. [Google Scholar] [CrossRef] [Green Version]
- Kusminski, C.M.; Holland, W.L.; Sun, K.; Park, J.; Spurgin, S.B.; Lin, Y.; Askew, G.R.; Simcox, J.A.; McClain, D.A.; Li, C.; et al. MitoNEET-driven alterations in adipocyte mitochondrial activity reveal a crucial adaptive process that preserves insulin sensitivity in obesity. Nat. Med. 2012, 18, 1539–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusminski, C.M.; Park, J.; Scherer, P.E. MitoNEET-mediated effects on browning of white adipose tissue. Nat. Commun. 2014, 5, 3962. [Google Scholar] [CrossRef] [Green Version]
- Raddock, M.L.; Wiley, S.E.; Axelrod, H.L.; Cohen, A.E.; Roy, M.; Abresch, E.C.; Capraro, D.; Murphy, A.N.; Nechushtai, R.; Dixon, J.E.; et al. MitoNEET is a uniquely folded 2Fe 2S outer mitochondrial membrane protein stabilized by pioglitazone. Proc. Natl. Acad. Sci. USA 2007, 104, 14342–14347. [Google Scholar] [CrossRef] [Green Version]
- Zuris, J.A.; Harir, Y.; Conlan, A.R.; Shvartsman, M.; Michaeli, D.; Tamir, S.; Paddock, M.L.; Onuchic, J.N.; Mittler, R.; Cabantchik, Z.I.; et al. Facile transfer of [2Fe-2S] clusters from the diabetes drug target mitoNEET to an apo-acceptor protein. Proc. Natl. Acad. Sci. USA 2011, 108, 13047–13052. [Google Scholar] [CrossRef] [Green Version]
- Tarze, A.; Deniaud, A.; Le Bras, M.; Maillier, E.; Molle, D.; Larochette, N.; Zamzami, N.; Jan, G.; Kroemer, G.; Brenner, C. GAPDH, a novel regulator of the pro-apoptotic mitochondrial membrane permeabilization. Oncogene 2007, 26, 2606–2620. [Google Scholar] [CrossRef] [Green Version]
- Bénit, P.; Pelhaître, A.; Saunier, E.; Bortoli, S.; Coulibaly, A.; Rak, M.; Schiff, M.; Kroemer, G.; Zeviani, M.; Rustin, P.E. Paradoxical inhibition of glycolysis by pioglitazone opposes the mitochondriopathy caused by AIF deficiency. BioMedicine 2017, 17, 75–87. [Google Scholar] [CrossRef] [Green Version]
- García-Ruiz, I.; Solís-Muñoz, P.; Fernández-Moreira, D.; Muñoz-Yagüe, T.; Solís-Herruzo, J.A. Pioglitazone leads to an inactivation and disassembly of complex I of the mitochondrial respiratory chain. BMC Biol. 2013, 11, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz, M.N.; Sánchez-Martín, C.; Detaille, D.; Vial, G.; Rigoulet, M.; El-Mir, M.Y.; Rodríguez-Villanueva, G. Acute mitochondrial actions of glitazones on the liver: A crucial parameter for their antidiabetic properties. Cell Physiol. Biochem. 2011, 28, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Geldenhuys, W.J.; Funk, M.O.; Barnes, K.F.; Carroll, R.T. Structure-based design of a thiazolidinedione which targets the mitochondrial protein mitoNEET. Bioorg. Med. Chem. Lett. 2010, 20, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Takemura, A.; Ito, K. Role of respiratory uncoupling in drug-induced mitochondrial permeability transition. Toxicol. Appl. Pharmacol. 2021, 15, 115659. [Google Scholar] [CrossRef]
- Pruimboom-Brees, I.M.; Francone, O.; Pettersen, J.C.; Kerlin, R.L.; Will, Y.; Amacher, D.E.; Boucher, G.G.; Morton, D. The development of subcutaneous sarcomas in rodents exposed to peroxisome proliferators agonists: Hypothetical mechanisms of action and de-risking attitude. Toxicol. Pathol. 2012, 40, 810–818. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E.; Daniele, G.; Galindo, C.; Abdul-Ghani, M.A.; DeFronzo, R.A.; Norton, L. Pioglitazone inhibits mitochondrial pyruvate metabolism and glucose production in hepatocytes. FEBS J. 2017, 284, 451–465. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, M.J. The Merck Index—An Encyclopedia of Chemicals, Drugs, and Biologicals, 15th ed.; Royal Society of Chemistry: Cambridge, UK, 2013; p. 1477. [Google Scholar]
- McLaughlin, S.G.; Dilger, J.P. Transport of protons across membranes by weak acids. Physiol. Rev. 1980, 60, 825–863. [Google Scholar] [CrossRef]
- Dubinin, M.V.; Khoroshavina, E.I.; Samartsev, V.N. Lithocholic acid induces two different calcium-dependent inner membrane permeability systems in liver mitochondria. Biochemistry 2017, 11, 231–236. [Google Scholar] [CrossRef]
- Kharechkina, E.S.; Nikiforova, A.B.; Teplova, V.V.; Odinokova, I.V.; Krestinina, O.V.; Baburina, Y.L.; Kruglova, S.A.; Kruglov, A.G. Regulation of permeability transition pore opening in mitochondria by external NAD(H). Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 771–783. [Google Scholar] [CrossRef]
- Hagen, T.; Lagace, C.J.; Modica-Napolitano, J.S.; Aprille, J.R. Permeability transition in rat liver mitochondria is modulated by the ATP-Mg/Pi carrier. Am. J. Physiol. 2003, 285, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Andreyev, A.Y.; Bondareva, T.O.; Dedukhova, V.I.; Mokhova, E.N.; Skulachev, V.P.; Tsofina, L.M.; Volkov, N.I.; Vygodina, T.V. The ATP/ADP-antiporter is involved in the uncoupling effect of fatty acids on mitochondria. Eur. J. Biochem. 1989, 182, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Žuna, K.; Jovanović, O.; Khailova, L.S.; Škulj, S.; Brkljača, Z.; Kreiter, J.; Kotova, E.A.; Vazdar, M.; Antonenko, Y.N.; Pohl, E.E. Mitochondrial Uncoupling Proteins (UCP1-UCP3) and Adenine Nucleotide Translocase (ANT1) Enhance the Protonophoric Action of 2,4-Dinitrophenol in Mitochondria and Planar Bilayer Membranes. Biomolecules 2021, 11, 1178. [Google Scholar] [CrossRef]
- Berardi, J.M.; Chou, J.J. Fatty acid flippase activity of UCP2 is essential for its proton transport in mitochondria. Cell Metab. 2014, 20, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seedher, N.; Kanojia, M. Co-solvent solubilization of some poorly-soluble antidiabetic drugs. Pharm Dev. Technol. 2009, 14, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.H.; Gatta, A.T.; Levine, T.P. Lipid transfer proteins: The lipid commute via shuttles, bridges and tubes. Nat. Rev. Mol. Cell Biol. 2019, 20, 85–101. [Google Scholar] [CrossRef]
- Geisler, J.G. 2,4 Dinitrophenol as Medicine. Cells 2019, 8, 280. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.A.; Johnson, R.A.; Corkey, B.; Kamp, F. Fatty acid transport: The diffusion mechanism in model and biological membranes. J. Mol. Neurosci. 2001, 16, 99–108. [Google Scholar] [CrossRef]
- Brustovetsky, N.N.; Dedukhova, V.I.; Egorova, M.V.; Mokhova, E.N.; Skulachev, V.P. Inhibitors of the ATP/ADP antiporter suppress stimulation of mitochondrial respiration and H+ permeability by palmitate and anionic detergents. FEBS 1990, 272, 187–189. [Google Scholar] [CrossRef] [Green Version]
- Brustovetsky, N.; Klingenberg, M. The reconstituted ADP/ATP carrier can mediate H+ transport by free fatty acids, which is further stimulated by mersalyl. J. Biol. Chem. 1994, 269, 27329–27336. [Google Scholar] [CrossRef]
- Bertholet, A.M.; Kirichok, Y. UCP1: A transporter for H+ and fatty acid anions. Biochimie 2017, 134, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardi, P.; Broekemeier, K.M.; Pfeiffer, D.R. Recent progress on regulation of the mitochondrial permeability transition pore; a cyclosporin-sensitive pore in the inner mitochondrial membrane. J. Bioenerg. Biomembr. 1994, 26, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Traba, J.; del Arco, A.; Duchen, M.R.; Szabadkai, G.; Satrústegui, J. SCaMC-1 promotes cancer cell survival by desensitizing mitochondrial permeability transition via ATP/ADP-mediated matrix Ca(2+) buffering. Cell Death Differ. 2012, 19, 650–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishna, K.; Rebecca Shiffali, D.; Pani Kumar, A.D. Estimation of Pioglitazone Hydrochloride in Bulk Drug and Pharmaceutical Dosage Forms by Hydrotropy Technique and Oxidative-Coupling Reaction. Asian J. Res. Chem. 2013, 6, 172–176. [Google Scholar]
- Khailova, L.S.; Firsov, A.M.; Kotova, E.A.; Antonenko, Y.N. Interaction of Potent Mitochondrial Uncouplers with Thiol-Containing Antioxidants. Antioxidants 2019, 8, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saremi, A.; Schwenke, D.C.; Buchanan, T.A.; Hodis, H.N.; Mack, W.J.; Banerji, M.; Bray, G.A.; Clement, S.C.; Henry, R.R.; Kitabchi, A.E.; et al. Pioglitazone slows progression of atherosclerosis in prediabetes independent of changes in cardiovascular risk factors. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 393–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldeira da Silva, C.C.; Cerqueira, F.M.; Barbosa, L.F.; Medeiros, M.H.; Kowaltowski, A.J. Mild mitochondrial uncoupling in mice affects energy metabolism, redox balance and longevity. Aging Cell 2008, 7, 552–560. [Google Scholar] [CrossRef]
- Perry, R.J.; Zhang, D.; Zhang, X.M.; Boyer, J.L.; Shulman, G.I. Controlled-release mitochondrial protonophore reverses diabetes and steatohepatitis in rats. Science 2015, 347, 1253–1256. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; Lardy, H.A. Isolation of liver or kidney mitochondria. Methods Enzymol. 1967, 10, 94–96. [Google Scholar] [CrossRef]
- Nikiforova, A.B.; Saris, N.E.; Kruglov, A.G. External mitochondrial NADH-dependent reductase of redox cyclers: VDAC1 or Cyb5R3? Free Radic. Biol. Med. 2014, 74, 74–84. [Google Scholar] [CrossRef]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar] [CrossRef]
- Chinopoulos, C.; Vajda, S.; Csanády, L.; Mándi, M.; Mathe, K.; Adam-Vizi, V. A novel kinetic assay of mitochondrial ATP-ADP exchange rate mediated by the ANT. Biophys. J. 2009, 96, 2490–2504. [Google Scholar] [CrossRef] [Green Version]
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Penkov, N.V.; Nedopekina, D.A.; Sharapov, V.A.; Khoroshavina, E.I.; Davletshin, E.V.; Belosludtseva, N.V.; Spivak, A.Y.; et al. Mitochondria-targeted prooxidant effects of betulinic acid conjugated with delocalized lipophilic cation F16. Free Radic Biol. Med. 2021, 168, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Schindler, M.; Simon, S.M. A mechanism for tamoxifen-mediated inhibition of acidification. J. Biol. Chem. 1999, 274, 18364–18373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denisov, S.S.; Kotova, E.A.; Khailova, L.S.; Korshunova, G.A.; Antonenko, Y.N. Tuning the hydrophobicity overcomes unfavorable deprotonation making octylamino-substituted 7-nitrobenz-2-oxa-1,3-diazole (n-octylamino-NBD) a protonophore and uncoupler of oxidative phosphorylation in mitochondria. Bioelectrochemistry 2014, 98, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Brierley, G.P.; Jurkowitz, M.; Scott, K.M.; Merola, A.J. Ion transport by heart mitochondria. XX. Factors affecting passive osmotic swelling of isolated mitochondria. J. Biol. Chem. 1970, 245, 5404–5411. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kharechkina, E.S.; Nikiforova, A.B.; Belosludtsev, K.N.; Rokitskaya, T.I.; Antonenko, Y.N.; Kruglov, A.G. Pioglitazone Is a Mild Carrier-Dependent Uncoupler of Oxidative Phosphorylation and a Modulator of Mitochondrial Permeability Transition. Pharmaceuticals 2021, 14, 1045. https://doi.org/10.3390/ph14101045
Kharechkina ES, Nikiforova AB, Belosludtsev KN, Rokitskaya TI, Antonenko YN, Kruglov AG. Pioglitazone Is a Mild Carrier-Dependent Uncoupler of Oxidative Phosphorylation and a Modulator of Mitochondrial Permeability Transition. Pharmaceuticals. 2021; 14(10):1045. https://doi.org/10.3390/ph14101045
Chicago/Turabian StyleKharechkina, Ekaterina S., Anna B. Nikiforova, Konstantin N. Belosludtsev, Tatyana I. Rokitskaya, Yuri N. Antonenko, and Alexey G. Kruglov. 2021. "Pioglitazone Is a Mild Carrier-Dependent Uncoupler of Oxidative Phosphorylation and a Modulator of Mitochondrial Permeability Transition" Pharmaceuticals 14, no. 10: 1045. https://doi.org/10.3390/ph14101045
APA StyleKharechkina, E. S., Nikiforova, A. B., Belosludtsev, K. N., Rokitskaya, T. I., Antonenko, Y. N., & Kruglov, A. G. (2021). Pioglitazone Is a Mild Carrier-Dependent Uncoupler of Oxidative Phosphorylation and a Modulator of Mitochondrial Permeability Transition. Pharmaceuticals, 14(10), 1045. https://doi.org/10.3390/ph14101045