Probing the Effects of the FGFR-Inhibitor Derazantinib on Vascular Development in Zebrafish Embryos

Abstract
1. Introduction
2. Results
2.1. Binding of Compounds in Cell Culture and to Zebrafish Proteins
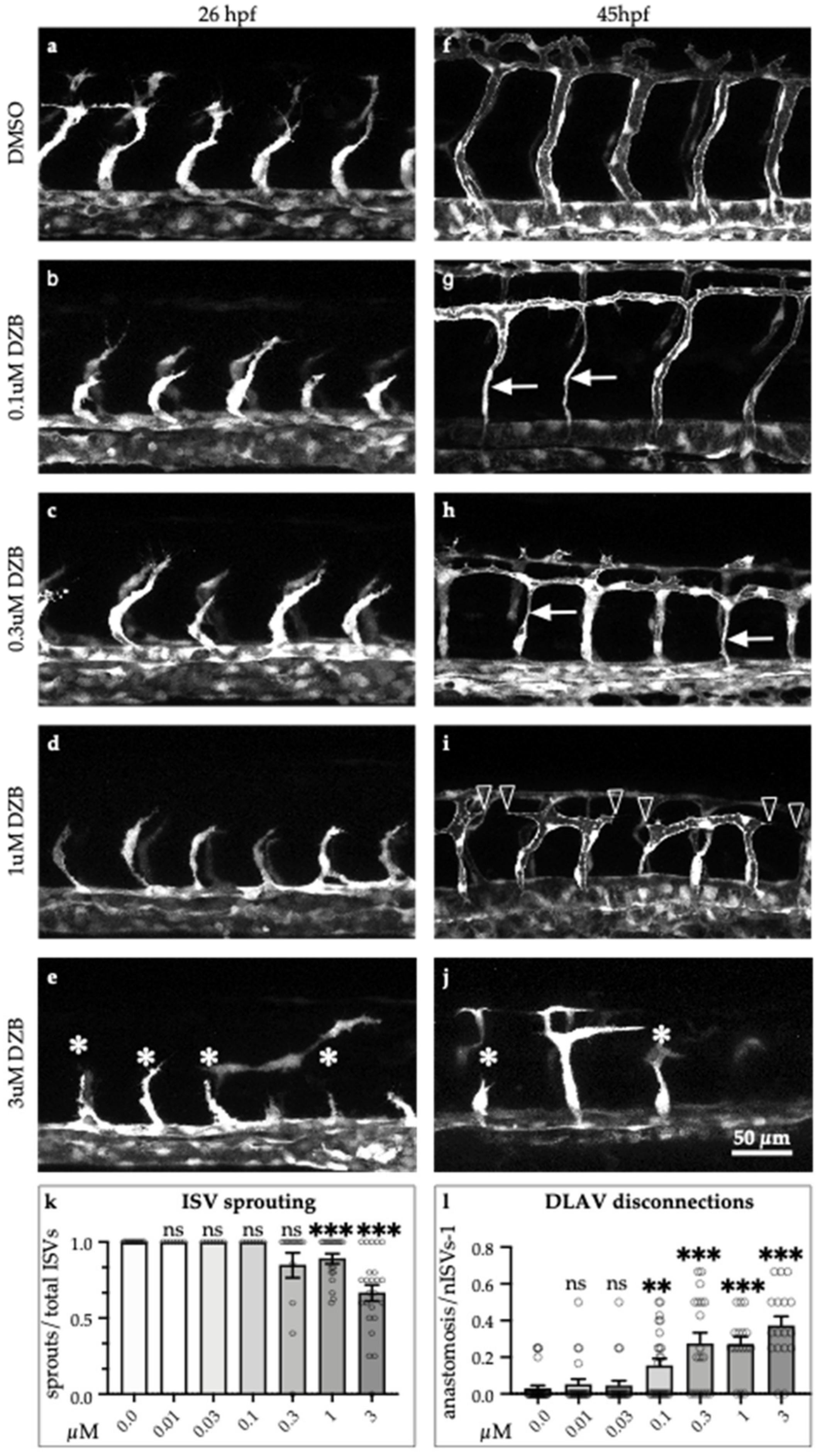
2.2. DZB Affects Vascular Development In Vivo in a Dose-Dependent Manner
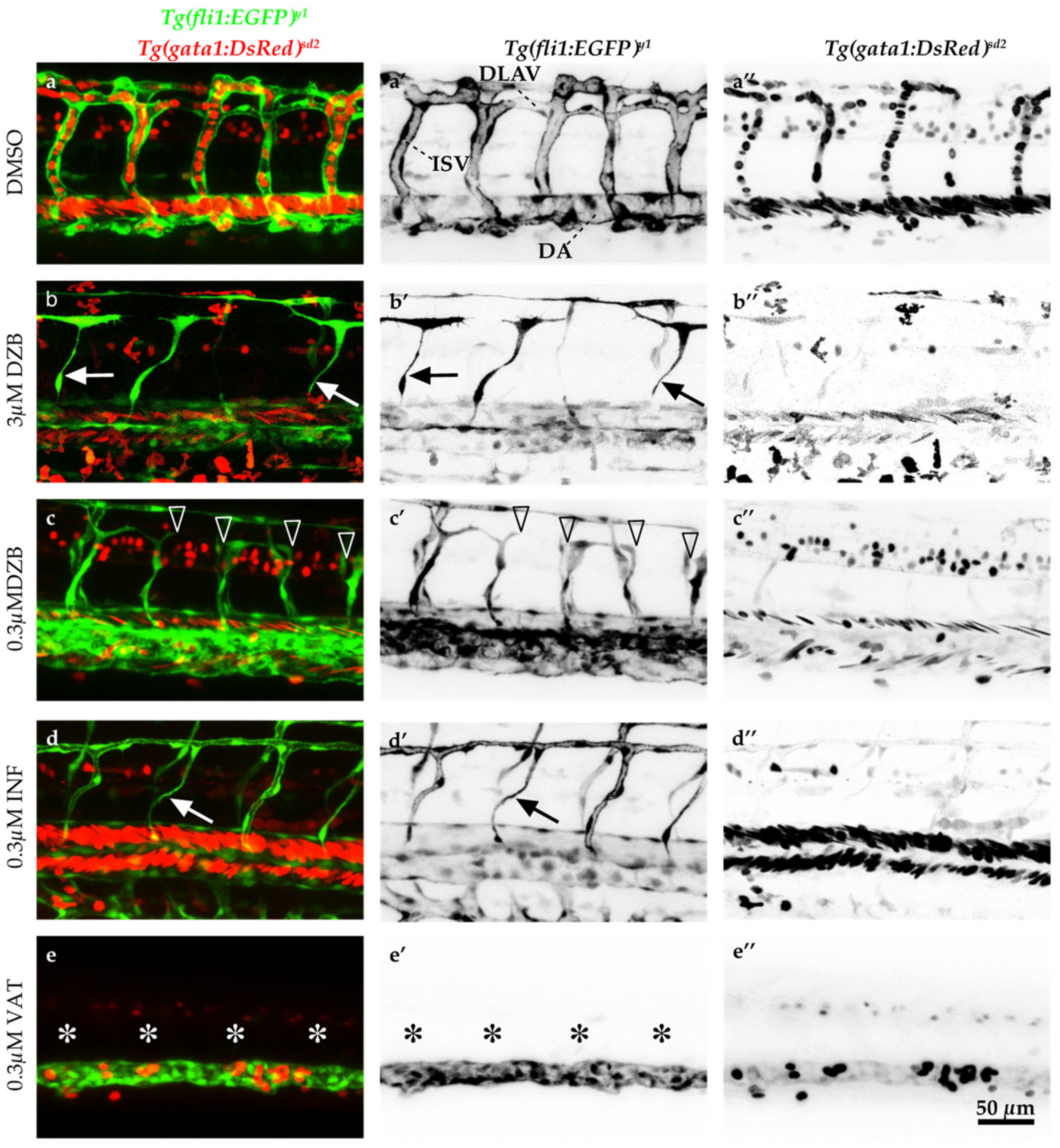
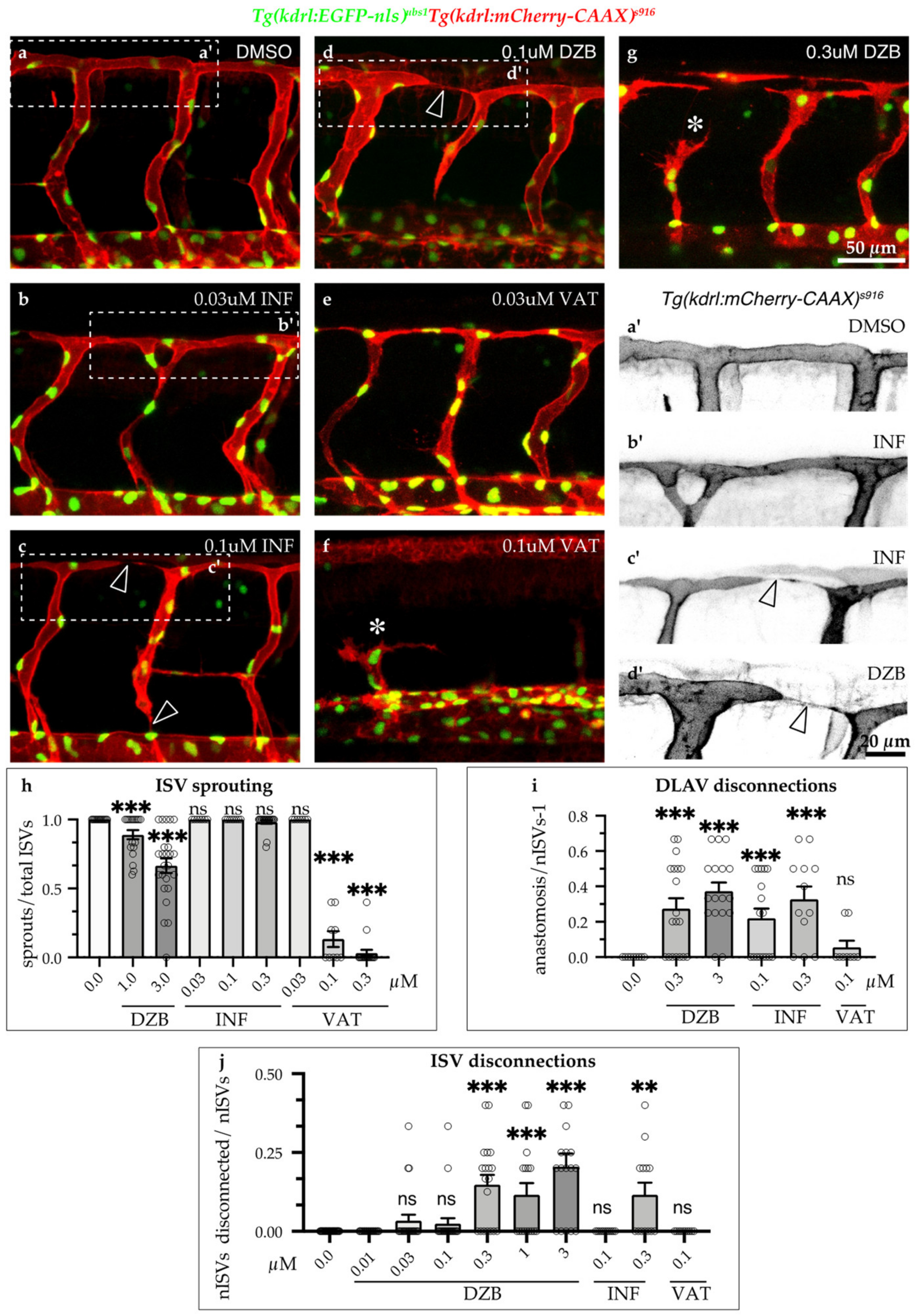
2.3. Comparisons of DZB with the FGFR Inhibitor Infigratinib and VEGFR Inhibitor Vatalanib with Regard to Their Effects on Vascular Development
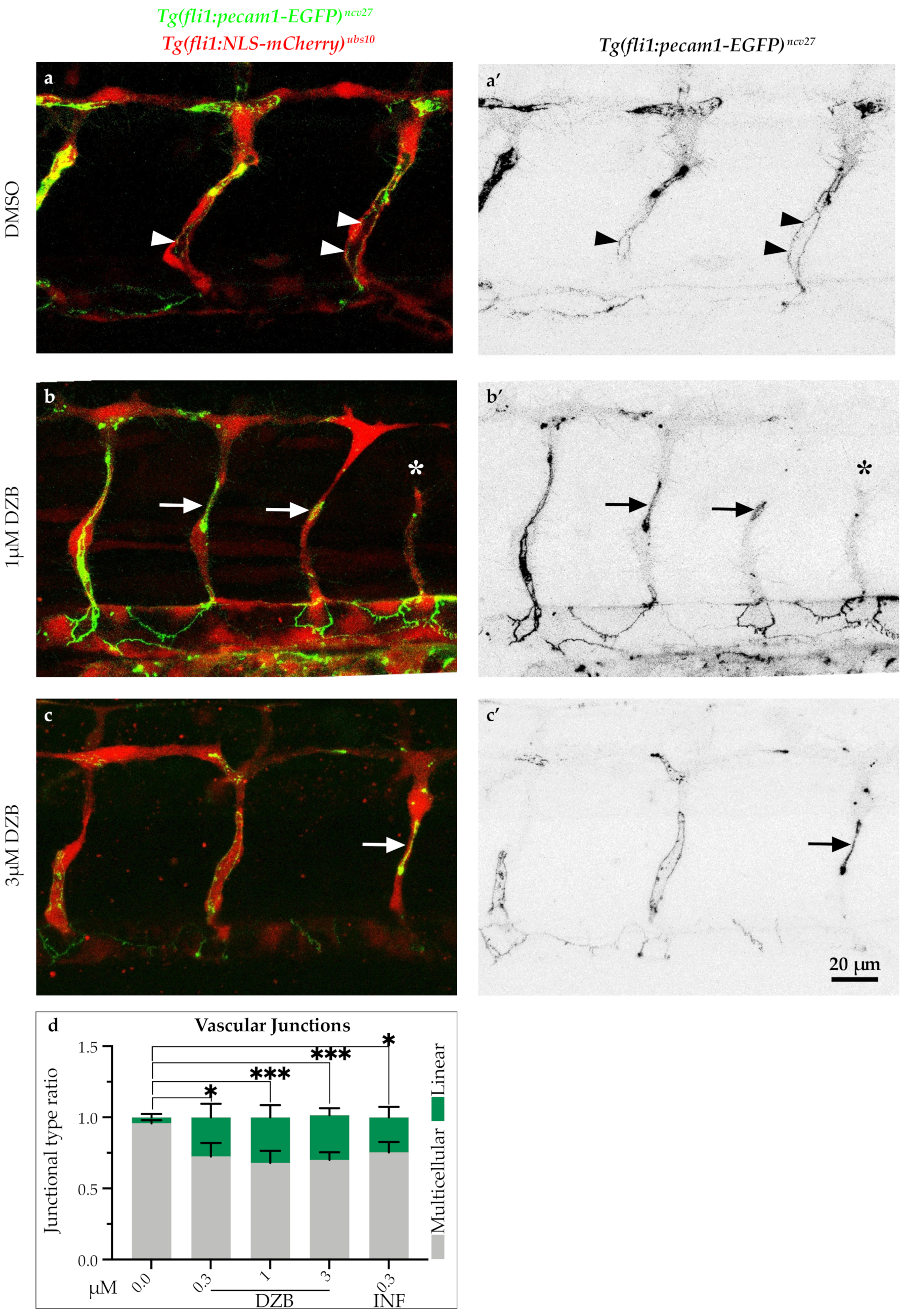
2.4. DZB Impairs Vascular Architecture
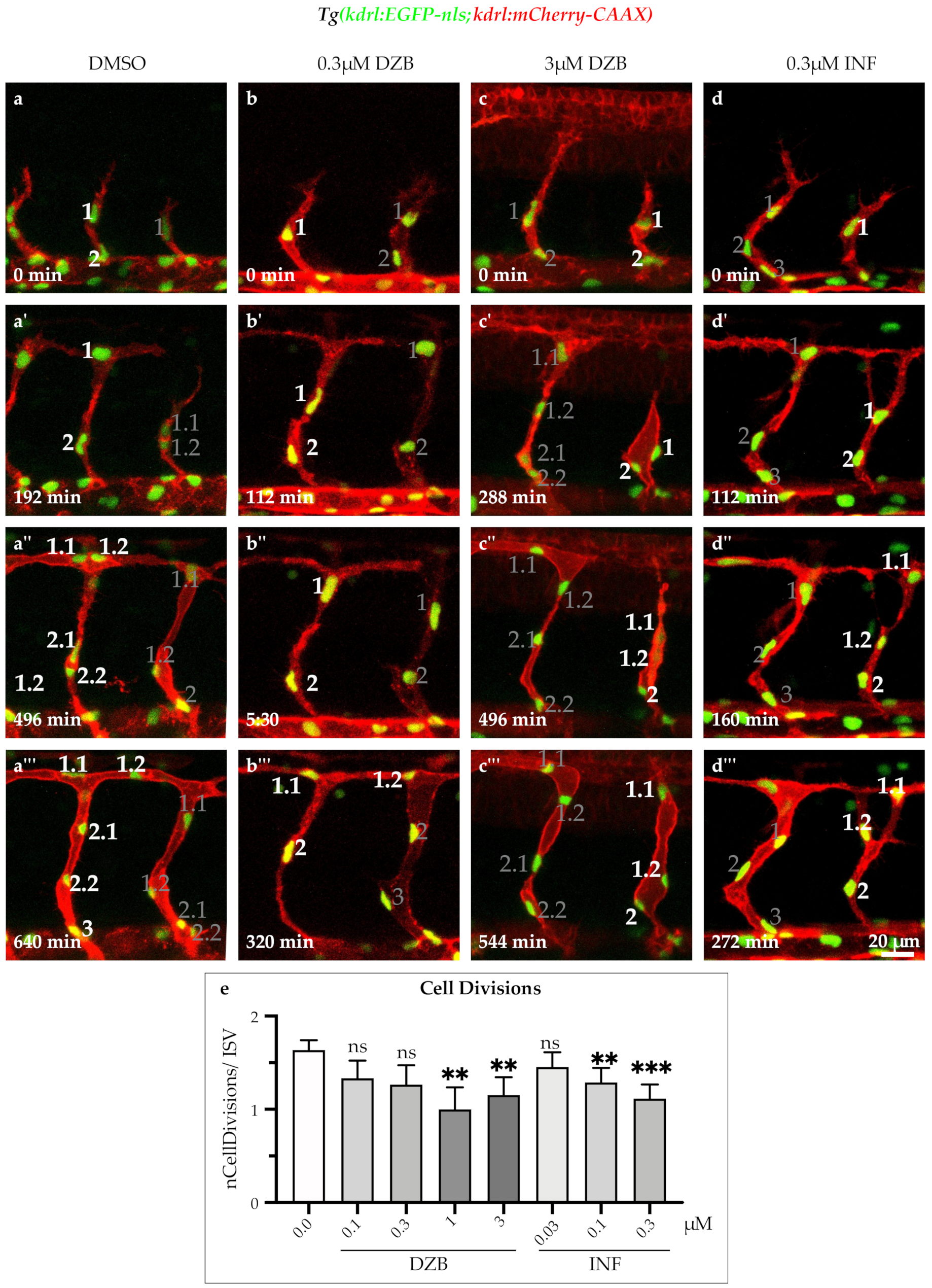
2.5. DZB Impedes the Endothelial Cell Cycle
3. Discussion
3.1. Concentration Range Effect of DZB Reveals Dual Role as Anti-Angiogenic Drug
3.2. Implications for Anastomosis and FGFR Inhibition
3.3. Implications for Sprouting
4. Materials and Methods
4.1. Measurement of Compound Binding to Mouse Plasma Protein and Zebrafish Protein
4.2. Zebrafish Maintenance and Transgenic Lines/Strains
4.3. In Vivo Time-Lapse Microscopy and Image Analysis
4.4. Pharmacological Treatment
4.5. Statistical Analysis and Graphs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Rao, N.; Lee, Y.F.; Ge, R. Novel endogenous angiogenesis inhibitors and their therapeutic potential. Acta Pharmacol. Sin. 2015, 36, 1177–1190. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.H.; Kim, M.R.; Jang, J.H.; Na, H.J.; Lee, S. A review of anti-angiogenic targets for monoclonal antibody cancer therapy. Int. J. Mol. Sci. 2017, 18, 1786. [Google Scholar] [CrossRef] [PubMed]
- El Alaoui-Lasmaili, K.; Faivre, B. Antiangiogenic therapy: Markers of response, “normalization” and resistance. Crit. Rev. Oncol. Hematol. 2018, 128, 118–129. [Google Scholar] [CrossRef]
- McDermott, D.F.; Huseni, M.A.; Atkins, M.B.; Motzer, R.J.; Rini, B.I.; Escudier, B.; Fong, L.; Joseph, R.W.; Pal, S.K.; Reeves, J.A.; et al. Clinical activity and molecular correlates of response to atezolizumab alone or in combination with bevacizumab versus sunitinib in renal cell carcinoma. Nat. Med. 2018, 24, 749–757. [Google Scholar] [CrossRef]
- Rahma, O.E.; Hodi, F.S. The intersection between tumor angiogenesis and immune suppression. Clin. Cancer Res. 2019, 25, 5449–5457. [Google Scholar] [CrossRef]
- Anandappa, A.J.; Wu, C.J.; Ott, P.A. Directing traffic: How to effectively drive t cells into tumors. Cancer Discov. 2020, 10, 185–197. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Jia, W.; Deng, H.; Li, G.; Deng, W.; Chen, J.; Kim, B.Y.S.; Jiang, W.; Liu, Q.; et al. Low-dose anti-angiogenic therapy sensitizes breast cancer to PD-1 blockade. Clin. Cancer Res. 2020, 26, 1712–1724. [Google Scholar] [CrossRef]
- Hall, T.G.; Yu, Y.; Eathiraj, S.; Wang, Y.; Savage, R.E.; Lapierre, J.M.; Schwartz, B.; Abbadessa, G. Preclinical activity of ARQ 087, a novel inhibitor targeting FGFR dysregulation. PLoS ONE 2016, 11, e0162594. [Google Scholar] [CrossRef]
- McSheehy, P.; Bachmann, F.; Forster-Gross, N.; Lecoultre, M.; Shemerly, M.E.; Roceri, M.; Reinelt, S.; Kellenberger, L.; Walker, P.R.; Lane, H. Derazantinib (DZB): A dual FGFR/CSF1R-inhibitor active in PDX-models of urothelial cancer. In Proceedings of the International Conference on Molecular Targets and Cancer Therapeutics, Boston, MA, USA, 26–30 October 2019; Volume 18 (Suppl. 12), p. 18. [Google Scholar]
- Mazzaferro, V.; El-Rayes, B.F.; Droz dit Busset, M.; Cotsoglou, C.; Harris, W.P.; Damjanov, N.; Masi, G.; Rimassa, L.; Personeni, N.; Braiteh, F.; et al. Derazantinib (ARQ 087) in advanced or inoperable FGFR2 gene fusion-positive intrahepatic cholangiocarcinoma. Br. J. Cancer 2019, 120, 165–171. [Google Scholar] [CrossRef]
- Wu, J.Q.; Fan, R.Y.; Zhang, S.R.; Li, C.Y.; Shen, L.Z.; Wei, P.; He, Z.H.; He, M.F. A systematical comparison of anti-angiogenesis and anti-cancer efficacy of ramucirumab, apatinib, regorafenib and cabozantinib in zebrafish model. Life Sci. 2020, 247, 117402. [Google Scholar] [CrossRef] [PubMed]
- De Smet, F.; Tembuyser, B.; Lenard, A.; Claes, F.; Zhang, J.; Michielsen, C.; Van Schepdael, A.; Herbert, J.M.; Bono, F.; Affolter, M.; et al. Fibroblast growth factor signaling affects vascular outgrowth and is required for the maintenance of blood vessel integrity. Chem. Biol. 2014, 21, 1310–1317. [Google Scholar] [CrossRef]
- Hogan, B.M.; Schulte-Merker, S. How to Plumb a Pisces: Understanding Vascular Development and Disease Using Zebrafish Embryos. Dev. Cell 2017, 42, 567–583. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Dong, L.; Ahn, J.; Dao, D.; Hammerschmidt, M.; Chen, J.N. FoxH1 negatively modulates flk1 gene expression and vascular formation in zebrafish. Dev. Biol. 2007, 304, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 2003, 161, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Betz, C.; Lenard, A.; Belting, H.-G.; Affolter, M. Cell behaviors and dynamics during angiogenesis. Development 2016, 143, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M.; Bold, G.; Buchdunger, E.; Cozens, R.; Ferrari, S.; Frei, J.; Hofmann, F.; Mestan, J.; Mett, H.; O’Reilly, T.; et al. PTK787/ZK 222584, a novel and potent inhibitor of vascular endothelial growth factor receptor tyrosine kinases, impairs vascular endothelial growth factor-induced responses and tumor growth after oral administration. Cancer Res. 2000, 60, 2178–2189. [Google Scholar]
- Guagnano, V.; Kauffmann, A.; Wöhrle, S.; Stamm, C.; Ito, M.; Barys, L.; Pornon, A.; Yao, Y.; Li, F.; Zhang, Y.; et al. FGFR genetic alterations predict for sensitivity to NVP-BGJ398, a selective Pan-FGFR inhibitor. Cancer Discov. 2012, 2, 1118–1133. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, C.; Shi, W.; Yang, L.; Zhang, Q.; Cui, J.; Fang, Y.; Li, Y.; Ren, G.; Yang, S.; et al. The novel VEGF receptor 2 inhibitor YLL545 inhibits angiogenesis and growth in breast cancer. Oncotarget 2016, 7, 41067–41080. [Google Scholar] [CrossRef]
- Cannon, J.E.; Upton, P.D.; Smith, J.C.; Morrell, N.W. Intersegmental vessel formation in zebrafish: Requirement for VEGF but not BMP signalling revealed by selective and non-selective BMP antagonists. Br. J. Pharmacol. 2010, 161, 140–149. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Angulo-Urarte, A.; Casado, P.; Castillo, S.D.; Kobialka, P.; Kotini, M.P.; Figueiredo, A.M.; Castel, P.; Rajeeve, V.; Milà-Guasch, M.; Millan, J.; et al. Endothelial cell rearrangements during vascular patterning require PI3-kinase-mediated inhibition of actomyosin contractility. Nat. Commun. 2018, 9, 4826. [Google Scholar] [CrossRef] [PubMed]
- Sauteur, L.; Krudewig, A.; Herwig, L.; Ehrenfeuchter, N.; Lenard, A.; Affolter, M.; Belting, H.G. Cdh5/VE-cadherin promotes endothelial cell interface elongation via cortical actin polymerization during angiogenic sprouting. Cell Rep. 2014, 9, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Ornitz, D.M.; Itoh, N. The fibroblast growth factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef]
- Katoh, M.; Nakagama, H. FGF Receptors: Cancer Biology and Therapeutics. Med. Res. Rev. 2014, 34, 280–300. [Google Scholar] [CrossRef]
- Lobjois, V.; Benazeraf, B.; Bertrand, N.; Medevielle, F.; Pituello, F. Specific regulation of cyclins D1 and D2 by FGF and Shh signaling coordinates cell cycle progression, patterning, and differentiation during early steps of spinal cord development. Dev. Biol. 2004, 273, 195–209. [Google Scholar] [CrossRef]
- Koziczak, M.; Holbro, T.; Hynes, N.E. Blocking of FGFR signaling inhibits breast cancer cell proliferation through downregulation of D-type cyclins. Oncogene 2004, 23, 3501–3508. [Google Scholar] [CrossRef]
- McSheehy, P.; Boult, J.; Robinson, S.; Bachmann, F.; El Shemerly, M.; Kellenberger, L.; Lane, H. Derazantinib, an oral fibroblast growth factor receptor inhibitor, in phase-2 clinical development, shows anti-angiogenic activity in pre-clinical models. Eur. J. Cancer 2020, 138, S25–S26. [Google Scholar] [CrossRef]
- Lin, R.Z.; Melero-Martin, J.M. Fibroblast growth factor-2 facilitates rapid anastomosis formation between bioengineered human vascular networks and living vasculature. Methods 2012, 56, 440–451. [Google Scholar] [CrossRef][Green Version]
- Murakami, M.; Nguyen, L.T.; Zhang, Z.W.; Moodie, K.L.; Carmeliet, P.; Stan, R.V.; Simons, M. The FGF system has a key role in regulating vascular integrity. J. Clin. Investig. 2008, 118, 3355–3366. [Google Scholar] [CrossRef]
- Aleström, P.; D’Angelo, L.; Midtlyng, P.J.; Schorderet, D.F.; Schulte-Merker, S.; Sohm, F.; Warner, S. Zebrafish: Housing and husbandry recommendations. Lab. Anim. 2020, 54, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Corti, P.; Young, S.; Chen, C.Y.; Patrick, M.J.; Rochon, E.R.; Pekkan, K.; Roman, B.L. Interaction between alk1 and blood flow in the development of arteriovenous malformations. Development 2011, 138, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Blum, Y.; Belting, H.G.; Ellertsdottir, E.; Herwig, L.; Lüders, F.; Affolter, M. Complex cell rearrangements during intersegmental vessel sprouting and vessel fusion in the zebrafish embryo. Dev. Biol. 2008, 316, 312–322. [Google Scholar] [CrossRef]
- Hogan, B.M.; Herpers, R.; Witte, M.; Heloterä, H.; Alitalo, K.; Duckers, H.J.; Schulte-Merker, S. Vegfc/Flt4 signalling is suppressed by Dll4 in developing zebrafish intersegmental arteries. Development 2009, 136, 4001–4009. [Google Scholar] [CrossRef]
- Ando, K.; Fukuhara, S.; Izumi, N.; Nakajima, H.; Fukui, H.; Kelsh, R.N.; Mochizuki, N. Clarification of mural cell coverage of vascular endothelial cells by live imaging of zebrafish. Development 2016, 143, 1328–1339. [Google Scholar] [CrossRef]
- Bussmann, J.; Bos, F.L.; Urasaki, A.; Kawakami, K.; Duckers, H.J.; Schulte-Merker, S. Arteries provide essential guidance cues for lymphatic endothelial cells in the zebrafish trunk. Development 2010, 137, 2653–2657. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, L.; Liu, X.; Chen, C.; Wang, B.; Wang, W.; Hu, C.; Yu, K.; Qi, Z.; Liu, Q.; et al. Discovery of a highly selective VEGFR2 kinase inhibitor CHMFL-VEGFR2-002 as a novel anti-angiogenesis agent. Acta Pharm. Sin. B 2020, 10, 488–497. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | %PB-ZF | %Fu-ZF 1 | Ratio in ZF (Fu) | %PPB-Mouse | %Fu-PPB | Ratio in Mice (Fu) |
|---|---|---|---|---|---|---|
| derazantinib | 99.93 | 0.067 ± 0.003 | 1 | 99.96 | 0.05 ± 0.01 | 1 |
| infigratinib | 95.03 | 4.97 ± 0.82 | 71 | 98.7 | 1.3 ± 0.00 | 29 |
| vatalanib | 90.05 | 9.95 ± 1.31 | 142 | 95.87 | 4.1 ± 0.18 | 92 |
| Developmental Process | Derazantinib | Derazantinib | Infigratinib | Vatalanib |
|---|---|---|---|---|
| Concentration range 1 | 0.1–0.3 μM | 1–3 μM | 0.1–0.3 μM | 0.1–0.3 μM |
| Calculated Fu in zebrafish 2 | 0.07–0.2 nM | 0.7–2 nM | 5–15 nM | 10–30 nM |
| ISV sprouting angiogenesis | No/mild defects | Strong defects | No defects | Strong defects |
| Anastomosis | Mild defects | Moderate/strong defects | Moderate/strong defects | n.a. |
| ISV-Aorta Connections | Mild defects | Moderate defects | Moderate defects | No defects |
| Lumenization | Mild defects | Mild defects | Mild defects | Moderate defects |
| Blood flow | No defects | Mild defects | Mild defects | Moderate defects |
| Cell junctions | Mild defects | Moderate/strong defects | Mild defects | n.a. |
| Cell cycle | Mild defects | Moderate defects | Moderate defects | n.a. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotini, M.P.; Bachmann, F.; Spickermann, J.; McSheehy, P.M.; Affolter, M. Probing the Effects of the FGFR-Inhibitor Derazantinib on Vascular Development in Zebrafish Embryos. Pharmaceuticals 2021, 14, 25. https://doi.org/10.3390/ph14010025
Kotini MP, Bachmann F, Spickermann J, McSheehy PM, Affolter M. Probing the Effects of the FGFR-Inhibitor Derazantinib on Vascular Development in Zebrafish Embryos. Pharmaceuticals. 2021; 14(1):25. https://doi.org/10.3390/ph14010025
Chicago/Turabian StyleKotini, Maria P., Felix Bachmann, Jochen Spickermann, Paul M. McSheehy, and Markus Affolter. 2021. "Probing the Effects of the FGFR-Inhibitor Derazantinib on Vascular Development in Zebrafish Embryos" Pharmaceuticals 14, no. 1: 25. https://doi.org/10.3390/ph14010025
APA StyleKotini, M. P., Bachmann, F., Spickermann, J., McSheehy, P. M., & Affolter, M. (2021). Probing the Effects of the FGFR-Inhibitor Derazantinib on Vascular Development in Zebrafish Embryos. Pharmaceuticals, 14(1), 25. https://doi.org/10.3390/ph14010025