In Vitro Antibacterial and Antiproliferative Potential of Echinops lanceolatus Mattf. (Asteraceae) and Identification of Potential Bioactive Compounds


Abstract
1. Introduction
2. Results
2.1. Antibacterial Potential
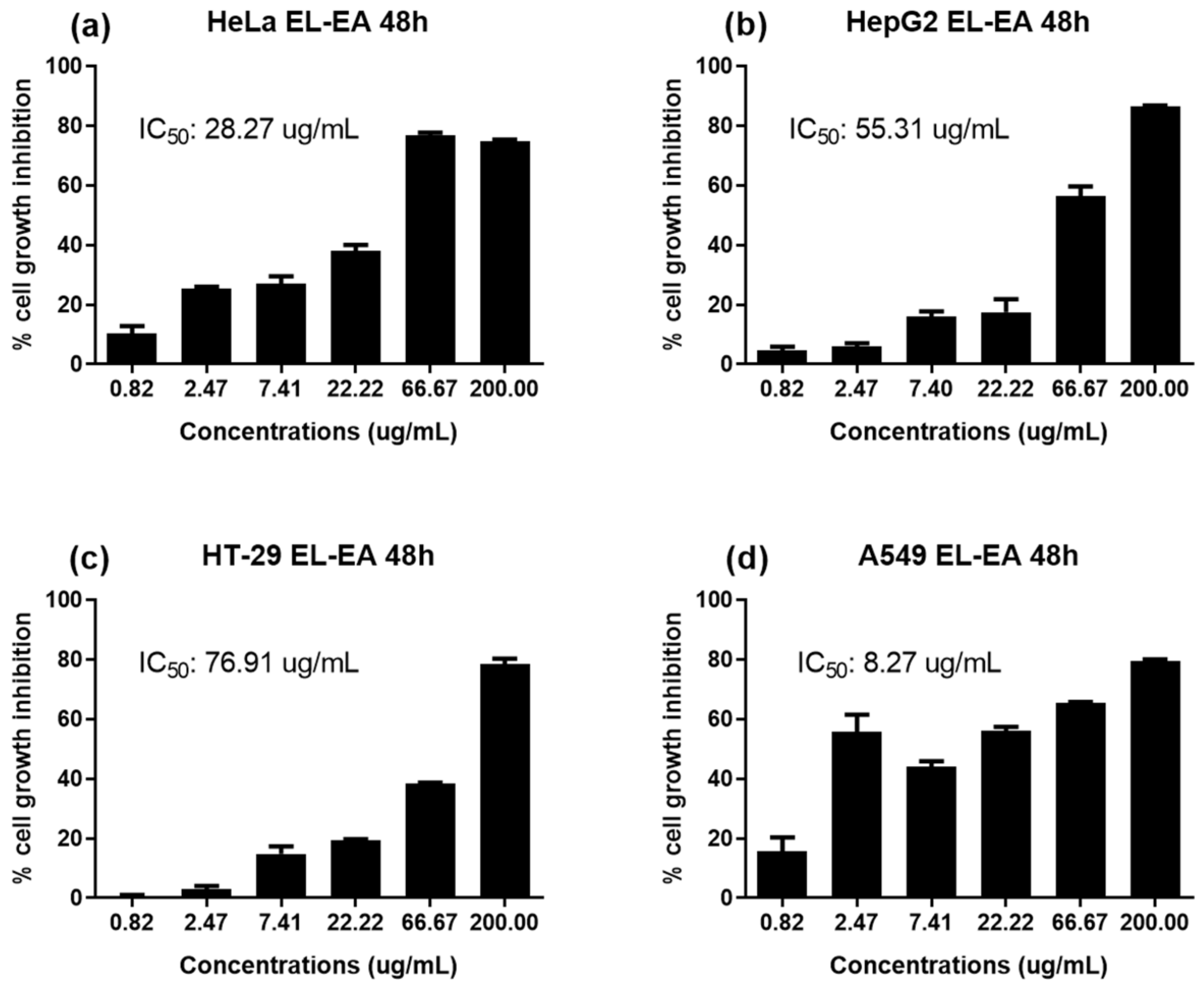
2.2. The Antiproliferative Potential of E. lanceolatus
2.3. UPLC–ESI–MS/MS Analysis of EL-EA Fraction
2.3.1. Phenolic Acids
2.3.2. Polyphenols
Flavonoids
Lignan
2.3.3. Organic Acids and Fatty Acids
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
4.3. Extraction Procedure
4.4. UPLC–ESI–MS/MS Analysis
4.5. Antibacterial Assay
4.5.1. Bacterial Strains
4.5.2. INT Colorimetric Assay for MIC and MBC Determinations
4.6. In Vitro Antiproliferative Assays
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis; WHO/EMP/IAU/2017.12; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Yu, Z.; Tang, J.; Khare, T.; Kumar, V. The alarming antimicrobial resistance in ESKAPEE pathogens: Can essential oils come to the rescue? Fitoterapia 2020, 140, 104433. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huang, B.X.; Guo, M. Current advances in screening for bioactive components from medicinal plants by affinity ultrafiltration mass spectrometry. Phytochem. Anal. 2018, 29, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.D.; Nahar, L. Computational Phytochemistry, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2018; p. 364. [Google Scholar]
- Bitew, H.; Hymete, A. The genus Echinops: Phytochemistry and biological activities: A review. Front. Pharm. 2019, 10, 1234. [Google Scholar] [CrossRef]
- Royal Botanical Garden (KEW). Echinops mildbraedii (Echinops lanceolatus). Available online: http://www.plantsoftheworldonline.org/taxon/urn:lsid:ipni.org:names:202586–1 (accessed on 13 December 2019).
- Roskov, Y.; Ower, G.; Orrell, T.; Nicolson, D.; Bailly, N.; Kirk, P.M.; Bourgoin, T.; DeWalt, R.E.; Decock, W.; Nieukerken, E.; et al. Species 2000 & ITIS Catalogue of Life, 2019 Annual Checklist; Species 2000: Naturalis, Leiden, The Netherlands, 2019. [Google Scholar]
- Liu, W.P.; Li, C.Y.; Huang, J.; Liao, J.Z.; Ma, W.J.; Chen, H.Y.; Rui, W. Identification of biomarkers in urine of rats with spleen Qi deficiency and biological significance. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Med. 2017, 42, 4855–4863. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, S.; Matei, M.F.; Jaiswal, R.; Bassil, B.S.; Kortz, U.; Kuhnert, N. Synthesis, structure, and tandem mass spectrometric characterization of the diastereomers of quinic acid. J. Agric. Food Chem. 2016, 64, 7298–7306. [Google Scholar] [CrossRef]
- Gruz, J.; Novák, O.; Strnad, M. Rapid analysis of phenolic acids in beverages by UPLC–MS/MS. Food Chem. 2008, 111, 789–794. [Google Scholar] [CrossRef]
- Bondia-Pons, I.; Barri, T.; Hanhineva, K.; Juntunen, K.; Dragsted, L.O.; Mykkänen, H.; Poutanen, K. UPLC-QTOF/MS metabolic profiling unveils urinary changes in humans after a whole grain rye versus refined wheat bread intervention. Mol. Nutr. Food Res. 2013, 57, 412–422. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhner, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Kuhnert, N.; Jaiswal, R.; Matei, M.F.; Sovdat, T.; Deshpande, S. How to distinguish between feruloyl quinic acids and isoferuloyl quinic acids by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 1575–1582. [Google Scholar] [CrossRef]
- Zhong, R.F.; Xu, G.B.; Wang, Z.; Wang, A.M.; Guan, H.Y.; Li, J.; He, X.; Liu, J.H.; Zhou, M.; Li, Y.J.; et al. Identification of anti-inflammatory constituents from Kalimeris indica with UHPLC-ESI-Q-TOF-MS/MS and GC–MS. J. Ethnopharmacol. 2015, 165, 39–45. [Google Scholar] [CrossRef]
- Moqbel, H.; El Hawary, S.S.E.D.; Sokkar, N.M.; El-Naggar, E.M.B.; El Boghdady, N.; El Halawany, A.M. HPLC-ESI-MS/MS characterization of phenolics in Prunus amygdalus, cultivar “umm alfahm” and its antioxidant and hepatoprotective activity. J. Food Meas. Charact. 2018, 12, 808–819. [Google Scholar] [CrossRef]
- Kammerer, B.; Kahlich, R.; Biegert, C.; Gleiter, C.H.; Heide, L. HPLC-MS/MS analysis of willow bark extracts contained in pharmaceutical preparations. Phytochem. Anal. 2005, 16, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Song, H.P.; Zhang, H.; Fu, Y.; Mo, H.Y.; Zhang, M.; Chen, J.; Li, P. Screening for selective inhibitors of xanthine oxidase from Flos chrysanthemum using ultrafiltration LC–MS combined with enzyme channel blocking. J. Chromatogr. B 2014, 961, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Mena, P.; Sánchez-Salcedo, E.M.; Tassotti, M.; Martínez, J.J.; Hernández, F.; Del Rio, D. Phytochemical evaluation of eight white (Morus alba L.) and black (Morus nigra L.) mulberry clones grown in Spain based on UHPLC-ESI-MSn metabolomic profiles. Food Res. Int. 2016, 89, 1116–1122. [Google Scholar] [CrossRef]
- Barreca, D.; Bellocco, E.; Caristi, C.; Leuzzi, U.; Gattuso, G. Flavonoid composition and antioxidant activity of juices from chinotto (Citrus myrtifolia Raf.) fruits at different ripening stages. J. Agric. Food Chem. 2010, 58, 3031–3036. [Google Scholar] [CrossRef]
- Arivalagan, M.; Roy, T.K.; Yasmeen, A.M.; Pavithra, K.C.; Jwala, P.N.; Shivasankara, K.S.; Manikantan, M.R.; Hebbar, K.B.; Kanade, S.R. Extraction of phenolic compounds with antioxidant potential from coconut (Cocos nucifera L.) testa and identification of phenolic acids and flavonoids using UPLC coupled with TQD-MS/MS. LWT 2018, 92, 116–126. [Google Scholar] [CrossRef]
- Seukep, A.J.; Kuete, V.; Nahar, L.; Sarker, S.D.; Guo, M. Plant-derived secondary metabolites as the main source of efflux pump inhibitors and methods for identification. J. Pharm. Anal. 2019. [Google Scholar] [CrossRef]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, Y.; Wang, T.; Dong, Q.; Li, J.; Niu, C. Detection of 12 common food-borne bacterial pathogens by TaqMan Real-Time PCR using a single set of reaction conditions. Front. Microbiol. 2019, 10, 222. [Google Scholar] [CrossRef]
- Nikaido, H. Prevention of drug access to bacterial targets: Permeability barriers and active efflux. Science 1994, 264, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005, 10, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Efferth, T. African flora has the potential to fight multidrug resistance of cancer. Biomed Res. Int. 2015, 2015, 914813. [Google Scholar] [CrossRef] [PubMed]
- Bruneton, J. Pharmacognosie: Phytochimie, Plantes Médicinales, 3rd ed.; Tec & Doc: Paris, France, 1999; pp. 263–309. [Google Scholar]
- Seukep, A.J.; Fankam, A.G.; Djeussi, D.E.; Voukeng, I.K.; Tankeo, S.B.; Noumedem, J.A.K.; Kuete, A.H.; Kuete, V. Antibacterial activities of the methanol extracts of seven Cameroonian dietary plants against bacteria expressing MDR phenotypes. Springerplus 2013, 2, 363. [Google Scholar] [CrossRef]
- Seukep, A.J.; Ngadjui, B.T.; Kuete, V. Antibacterial activities of Fagara macrophylla, Canarium schweinfurthii, Myrianthus arboreus, Dischistocalyx grandifolius and Tragia benthamii against multi-drug resistant Gram-negative bacteria. Springerplus 2015, 4, 567. [Google Scholar] [CrossRef]
- Seukep, A.J.; Sandjo, L.P.; Ngadjui, B.T.; Kuete, V. Antibacterial and antibiotic-resistance modifying activity of the extracts and compounds from Nauclea pobeguinii against Gram-negative multi-drug resistant phenotypes. BMC Complement. Altern. Med. 2016, 16, 193. [Google Scholar] [CrossRef]
- Seukep, A.J.; Sandjo, L.P.; Ngadjui, B.T.; Kuete, V. Antibacterial activities of the methanol extracts and compounds from Uapaca togoensis against Gram-negative multi-drug resistant phenotypes. S. Afr. J. Bot. 2016, 103, 1–5. [Google Scholar] [CrossRef]
- Vandal, J.; Abou-Zaid, M.M.; Ferroni, G.; Leduc, L.G. Antimicrobial activity of natural products from the flora of Northern Ontario, Canada. Pharm. Biol. 2015, 53, 800–806. [Google Scholar] [CrossRef]
- Kima, J.E.; Seob, J.H.; Baec, M.S.; Baed, C.-S.; Yooe, J.C.; Bangf, M.A.; Choa, S.S.; Park, D.H. Antimicrobial constituents from Allium hookeri Root. Nat. Prod. Commun. 2016, 11, 237–238. [Google Scholar] [CrossRef]
- Purohit, A.; Mohan, A. Antimicrobial effects of pyruvic and succinic acids on Salmonella survival in ground chicken. LWT 2019, 116, 108596. [Google Scholar] [CrossRef]
- Kumar, R.; Chandar, B.; Parani, M. Use of succinic and oxalic acid in reducing the dosage of colistin against New Delhi metallo-β-lactamase- bacteria. Indian J. Med. Res. 2018, 147, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Gaglione, M.; Malgieri, G.; Pacifico, S.; Severino, V.; D’Abrosca, B.; Russo, L.; Fiorentino, A.; Messere, A. Synthesis and biological properties of caffeic acid-PNA dimers containing guanine. Molecules 2013, 18, 9147–9162. [Google Scholar] [CrossRef] [PubMed]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Ezzat, S.M.; Dine, R.S.; Abdel-Sattar, E.; Abdel-Naim, A.B. Protective effect of Echinops galalensis against CCl4-induced injury on the human hepatoma cell line (Huh7). Phytochem. Lett. 2013, 6, 73–78. [Google Scholar] [CrossRef]
- Fraisse, D.; Felgines, C.; Texier, O.; Lamaison, J.L. Caffeoyl derivatives: Major antioxidant compounds of some wild herbs of the Asteraceae family. Food Nutr. Sci. 2011, 2, 181–192. [Google Scholar] [CrossRef]
- Mijangos-Ramos, I.F.; Zapata-Estrellaa, H.E.; Ruiz-Vargas, J.A.; Escalante-Erosa, F.; Gómez-Ojeda, N.; García-Sosa, K.; Cechinel-Filho, V.; Meira-Quintão, N.L.; Pena-Rodríguez, L.M. Bioactive dicaffeoylquinic acid derivatives from the root extract of Calea urticifolia. Rev. Bras. Farm. 2018, 28, 339–343. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Pereira, C.; Tzortzakis, N.; Barros, L.; Ferreira, I.C.F.R. Nutritional value and bioactive compounds characterization of plant parts from Cynara cardunculus L. (Asteraceae) cultivated in Central Greece. Front. Plant Sci. 2018, 9, 459. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, G.; Sun, C.; Li, H.; Fu, Y.; Xu, W. Apoptosis effects of dihydrokaempferol isolated from Bauhinia championii on Synoviocytes. Evid. Based Complement. Altern. Med. 2018, 2018, 10. [Google Scholar] [CrossRef]
- Harborne, J.B. The Flavonoids Advances in Research Since 1986, 1st ed.; Routledge, Chapman and Hall/CRC: Abingdon-on-Thames, UK, 2017; p. 676. [Google Scholar]
- Wang, Y.; Xu, Z.; Huang, Y.; Wen, X.; Wu, Y.; Zhao, Y.; Ni, Y. Extraction, purification, and hydrolysis behavior of Apigenin-7-O-glucoside from Chrysanthemum morifolium tea. Molecules 2018, 23, 2933. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.L.; She, W.Z.; Guo, G.Q.; Jiang, Z.Y.; Cen, Y.Z.; Fan, Z.Y. Studies on antiproliferative effect of flavones compounds isolated from Yao herb medicines. Zhong Yao Cai 2007, 30, 980–984. [Google Scholar] [PubMed]
- Smiljkovic, M.; Stanisavljevic, D.; Stojkovic, D.; Petrovic, I.; Vicentic, J.M.; Popovic, J.; Grdadolnik, S.G.; Markovic, D.; Sankovic-Babice, S.; Glamoclija, J.; et al. Apigenin-7-O-glucoside versus apigenin: Insight into the modes of anticandidal and cytotoxic actions. EXCLI J. 2017, 16, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Qi, Q.L.; Wang, M.T.; Li, Q.Y. Therapeutic potential of naringin: An overview. Pharm. Biol. 2016, 54, 3203–3210. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kregiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The therapeutic potential of apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed]
- Refaat, J.; Desoukey, Y.S.; Ramadan, M.A.; Kamel, M.S. Rhoifolin: A review of sources and biological activities. IJP 2015, 2, 102–109. [Google Scholar] [CrossRef]
- Xu, J.P. Cancer Inhibitors from Chinese Natural Medicines, 1st ed.; CRC Press, Francis and Taylor Group: Abingdon, UK, 2016; p. 787. [Google Scholar]
- Céspedes, C.L.; Avila, J.G.; García, A.M.; Becerra, J.; Flores, C.; Aqueveque, P.; Bittner, M.; Hoeneisen, M.; Martinez, M.; Silva, M. Antifungal and antibacterial activities of Araucaria araucana (Mol.) K. Koch heartwood lignans. Zeitschrift Naturforschung C 2006, 61, 35–43. [Google Scholar] [CrossRef]
- Kezimana, P.; Dmitriev, A.A.; Kudryavtseva, A.V.; Romanova, E.V.; Melnikova, N.V. Secoisolariciresinol diglucoside of flaxseed and its metabolites: Biosynthesis and potential for nutraceuticals. Front. Genet. 2018, 9, 641. [Google Scholar] [CrossRef]
- Dilika, F.; Bremner, P.D.; Meyer, J.J. Antibacterial activity of linoleic and oleic acids isolated from Helichrysum pedunculatum: A plant used during circumcision rites. Fitoterapia 2000, 71, 450–452. [Google Scholar] [CrossRef]
- Parsaee, H.; Asilib, J.; Mousavic, S.H.; Soofi, H.; Emami, S.A.; Tayarani-Najarane, Z. Apoptosis induction of Salvia chorassanica root extract on human cervical cancer cell line. Iran J. Pharm. Res. 2013, 12, 75–83. [Google Scholar] [PubMed]
- Chen, G.; Li, X.; Saleri, F.; Guo, M. Analysis of flavonoids in Rhamnus davurica and its antiproliferative activities. Molecules 2016, 21, 1275. [Google Scholar] [CrossRef]
- Dzoyem, J.P.; Guru, S.K.; Pieme, C.A.; Kuete, V.; Sharma, A.; Khan, I.A.; Saxena, A.K.; Vishwakarma, R.A. Cytotoxic and antimicrobial activity of selected Cameroonian edible plants. BMC Complement. Altern. Med. 2013, 13, 78. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria Strains | MIC and MBC (µg/mL) of Echinops lanceolatus Extracts * | ATB | |||||
|---|---|---|---|---|---|---|---|
| MeOH | Hex | DCM | EA | BuOH | STR | ||
| Gram-negative | Salmonella enterica | 1024 | 512 | 512 | 256 | 1024 | 4 |
| Escherichia coli | >1024 | >1024 | 1024 | 1024 | 512 | 8 | |
| Enterobacter cloacae | 1024 | 1024 | 1024 | 512 | 1024 | 64 | |
| Klebsiella pneumoniae | 1024 | 1024 | 1024 | >1024 | >1024 | >256 | |
| Pseudomonas aeruginosa | >1024 | >1024 | >1024 | 1024 | >1024 | >256 | |
| Acinetobacter baumannii | >1024 | >1024 | >1024 | 1024 | >1024 | 256 | |
| Gram-positive | Staphylococcus aureus | 1024 | 1024 | 512 | 512 (1024) B | 1024 | 4 |
| Enterococcus faecalis | >1024 | >1024 | >1024 | >1024 | >1024 | 256 | |
| Peak | Rt (min) | [M-H]− (m/z) | MF | MS2 (m/z) | Identification * | Ref |
|---|---|---|---|---|---|---|
| 1 | 1.16 | 117.0180 | C4H6O4 | 117.0179, 99.0074, 73.0282 | Succinic acid | [9] |
| 2 | 1.22 | 191.0551 | C7H12O6 | 191.0552, 173.0453, 127.0388, 93.0333, 85.0282 | Quinic acid | [10] |
| 3 | 1.34 | 153.0181 | C7H6O4 | 152.9947, 109.0282, 108.0201 | Dihydroxybenzoic acid | [11] |
| 4 | 1.53 | 167.0338 | C8H8O4 | 167.0334, 152.0104, 123.0440, 108.0204, 91.0177 | Hydroxy-methoxy-benzoic acid isomer 1 | [11] |
| 5 | 1.69 | 179.0339 | C9H8O4 | 179.0553, 135.0442 | Dihydroxycinnamic acid isomer 1 | [11] |
| 6 | 2.15 | 167.0338 | C8H8O4 | 167.0334, 152.0104, 123.0440, 108.0204, 91.0177 | Hydroxy-methoxy-benzoic acid isomer 2 | [11] |
| 7 | 2.44 | 179.0338 | C9H8O4 | 179.0553, 135.0442 | Dihydroxycinnamic acid isomer 2 | [11] |
| 8 | 3.08 | 163.0389 | C9H8O3 | 163.0390, 120.0523, 119.0490 | Coumaric acid isomer 1 | [11] |
| 9 | 3.29 | 163.0389 | C9H8O3 | 163.0390, 120.0523, 119.0490 | Coumaric acid isomer 2 | [11] |
| 10 | 4.58 | 187.0965 | C9H16O4 | 187.0963, 125.0960, 97.0643 | Nonanedioic acid | [12] |
| 11 | 5.70 | 137.0231 | C7H6O3 | 137.0232, 93.0333, 71.4599 | 3-Hydroxybenzoic acid | [11] |
| 12 | 5.97 | 337.0924 | C16H18O8 | 191.0552, 173.0444, 163.0389, 119.0489, 93.0332 | 5-Coumaroylquinic acid | [13] |
| 13 | 6.67 | 367.1027 | C17H20O9 | 367.1055, 193.0497, 191.0553, 173.0444, 134.0361, 93.0330 | 5-Feruloyl quinic acid | [14] |
| 14 | 7.67 | 515.1187 | C25H24O12 | 353.0876, 191.0553, 179.0340, 161.0234, 135.0439, 93.0332, 85.0282 | Dicaffeoylquinicacid isomer 1 | [15] |
| 15 | 7.94 | 515.1186 | C25H24O12 | 353.0874, 191.0552, 179.0340, 173.0445, 135.0439, 93.0332, 85.0282 | Dicaffeoylquinicacid isomer 2 | [15] |
| 16 | 8.29 | 515.1189 | C25H24O12 | 515.1190, 353.0875, 191.0552, 179.0340, 173.0445, 161.0233, 135.0439 | Dicaffeoylquinicacid isomer 3 | [15] |
| 17 | 8.72 | 515.1189 | C25H24O12 | 515.1190, 353.0875, 191.0552, 179.0340, 173.0445, 135.0439, 93.0333 | Dicaffeoylquinicacid isomer 4 | [15] |
| 18 | 9.04 | 287.0556 | C15H12O6 | 287.0921, 259.0608, 243.0658, 201.0549, 177.0547, 151.0025, 125.0231 | Dihydrokaempferol | [16] |
| 19 | 9.33 | 433.1134 | C21H22O10 | 433.1120, 313.0716, 271.0608, 177.0181, 151.0025, 119.0489, 107.0125 | Naringenin-7-O-glucoside | [17] |
| 20 | 9.59 | 431.0977 | C21H20O10 | 431.0980, 269.0441, 268.0373, 240.0422, 211.0392, 151.0025, 117.0333 | Apigenin-7-O-glucoside | [18] |
| 21 | 10.53 | 177.0546 | C10H10O3 | 177.0547, 162.0312, 145.0283, 133.0283, 123.0439, 121.0282, 117.0333 | Coniferyl aldehyde | [19] |
| 22 | 10.61 | 579.1503 | C27H32O14 | 579.1474, 307.0825, 271.0610, 145.0283, 119.0490, 117.0334 | Naringin | [20] |
| 23 | 10.86 | 577.1346 | C27H30O14 | 577.1351, 431.0982, 269.0453, 145.0284, 117.0333 | Rhoifolin | [20] |
| 24 | 11.02 | 269.0451 | C15H10O5 | 269.0451, 225.0551, 151.0025, 117.0332 | Apigenin | [21] |
| 25 | 11.12 | 329.2328 | C18H34O5 | 329.2331, 311.2234, 229.1438, 211.1332, 171.1016, 139.1117, 99.0802 | (-)-Pinellic acid | / |
| 26 | 11.87 | 361.0818 | C20H26O6 | 361.0820, 288.0636, 269.0451, 163.0390, 145.0283, 117.0333 | (-)-Secoisolariciresinol | / |
| 27 | 13.09 | 293.2117 | C18H30O3 | 293.2119, 265.2168 | 13-Keto-9Z,11E-octadecadienoic acid | / |
| 28 | 13.45 | 295.2273 | C18H32O3 | 295.2273, 251.2378 | 13S-Hydroxyoctadecadienoic acid | / |
| 29 | 13.69 | 275.1647 | C17H24O3 | 275.1648, 215.1434, 59.0125 | Cyclandelate | / |
| 30 | 14.17 | 277.2166 | C18H30O2 | 277.2166 | α-Linolenic acid | / |
| 31 | 14.63 | 255.2324 | C16H32O2 | 255.2326 | Ethyl myristate | / |
| 32 | 14.74 | 281.2479 | C18H34O2 | 281.1389 | Oleic acid | / |
| Time | Phase A (5mM Ammonium Acetate, pH 9) | Phase B (50% ACN, 10mM Ammonium Acetate, pH 9) |
|---|---|---|
| 0.00 | 98% | 2% |
| 1.50 | 98% | 2% |
| 12.00 | 0% | 100% |
| 14.00 | 0% | 100% |
| 14.10 | 98% | 2% |
| 16.00 | 98% | 2% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson Seukep, A.; Zhang, Y.-L.; Xu, Y.-B.; Guo, M.-Q. In Vitro Antibacterial and Antiproliferative Potential of Echinops lanceolatus Mattf. (Asteraceae) and Identification of Potential Bioactive Compounds. Pharmaceuticals 2020, 13, 59. https://doi.org/10.3390/ph13040059
Jackson Seukep A, Zhang Y-L, Xu Y-B, Guo M-Q. In Vitro Antibacterial and Antiproliferative Potential of Echinops lanceolatus Mattf. (Asteraceae) and Identification of Potential Bioactive Compounds. Pharmaceuticals. 2020; 13(4):59. https://doi.org/10.3390/ph13040059
Chicago/Turabian StyleJackson Seukep, Armel, Yong-Li Zhang, Yong-Bing Xu, and Ming-Quan Guo. 2020. "In Vitro Antibacterial and Antiproliferative Potential of Echinops lanceolatus Mattf. (Asteraceae) and Identification of Potential Bioactive Compounds" Pharmaceuticals 13, no. 4: 59. https://doi.org/10.3390/ph13040059
APA StyleJackson Seukep, A., Zhang, Y.-L., Xu, Y.-B., & Guo, M.-Q. (2020). In Vitro Antibacterial and Antiproliferative Potential of Echinops lanceolatus Mattf. (Asteraceae) and Identification of Potential Bioactive Compounds. Pharmaceuticals, 13(4), 59. https://doi.org/10.3390/ph13040059