Behavioral and Neurochemical Consequences of Pentylenetetrazol-Induced Kindling in Young and Middle-Aged Rats

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
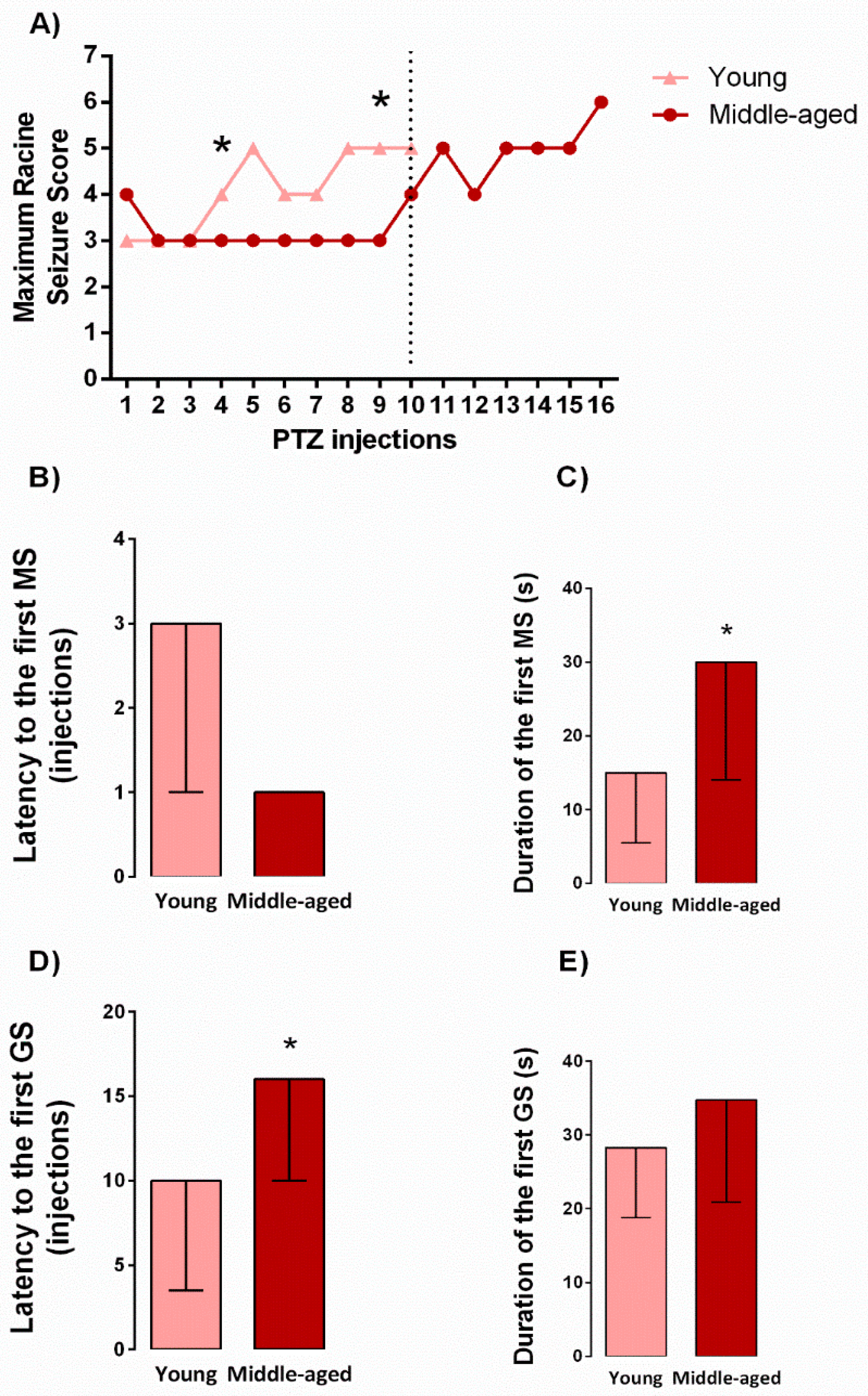
2.1. Behavioral Seizure Threshold during Kindling Induction
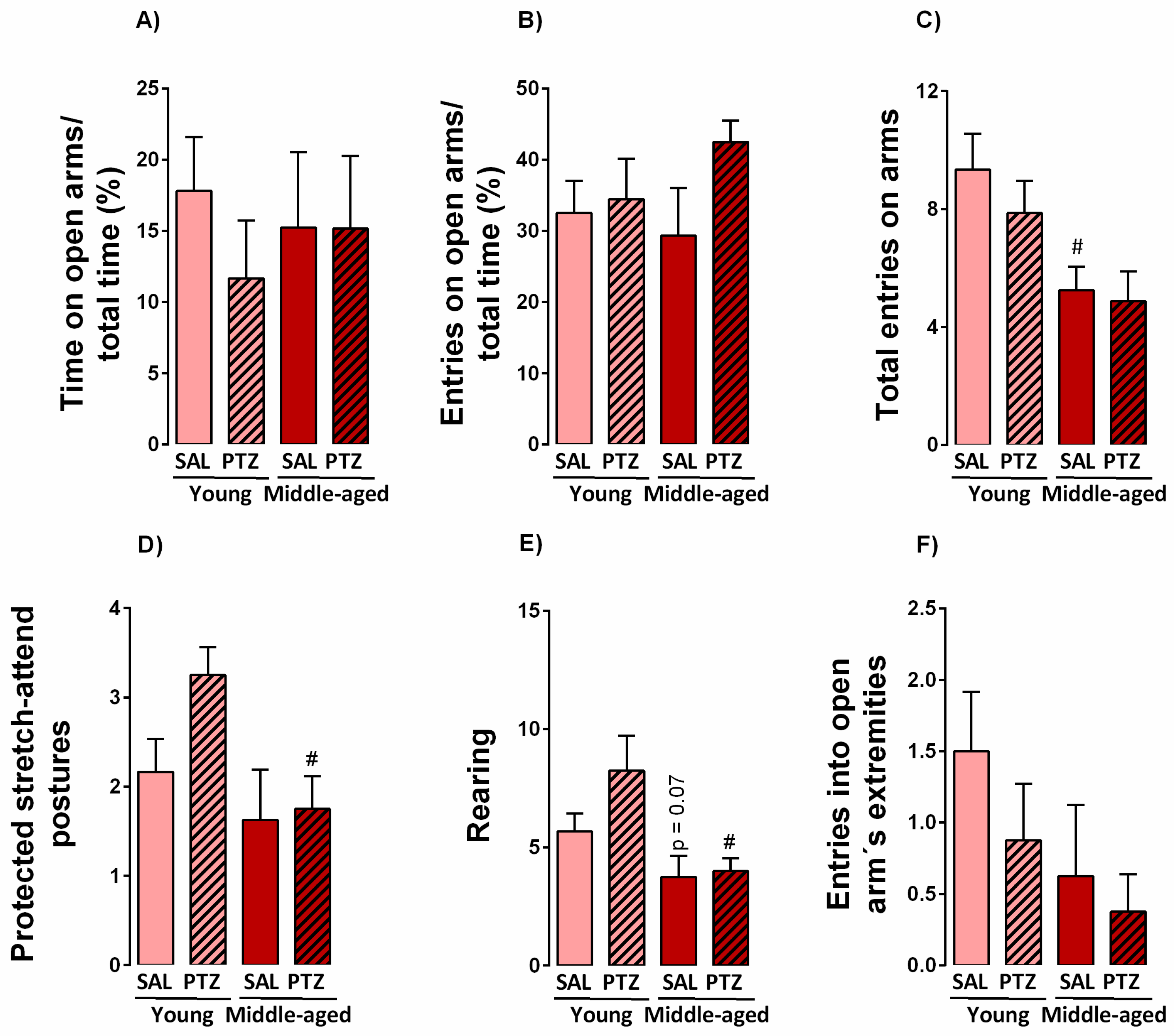
2.2. Anxiety-Like Behavior Following Kindling Induction in Middle-Aged Rats
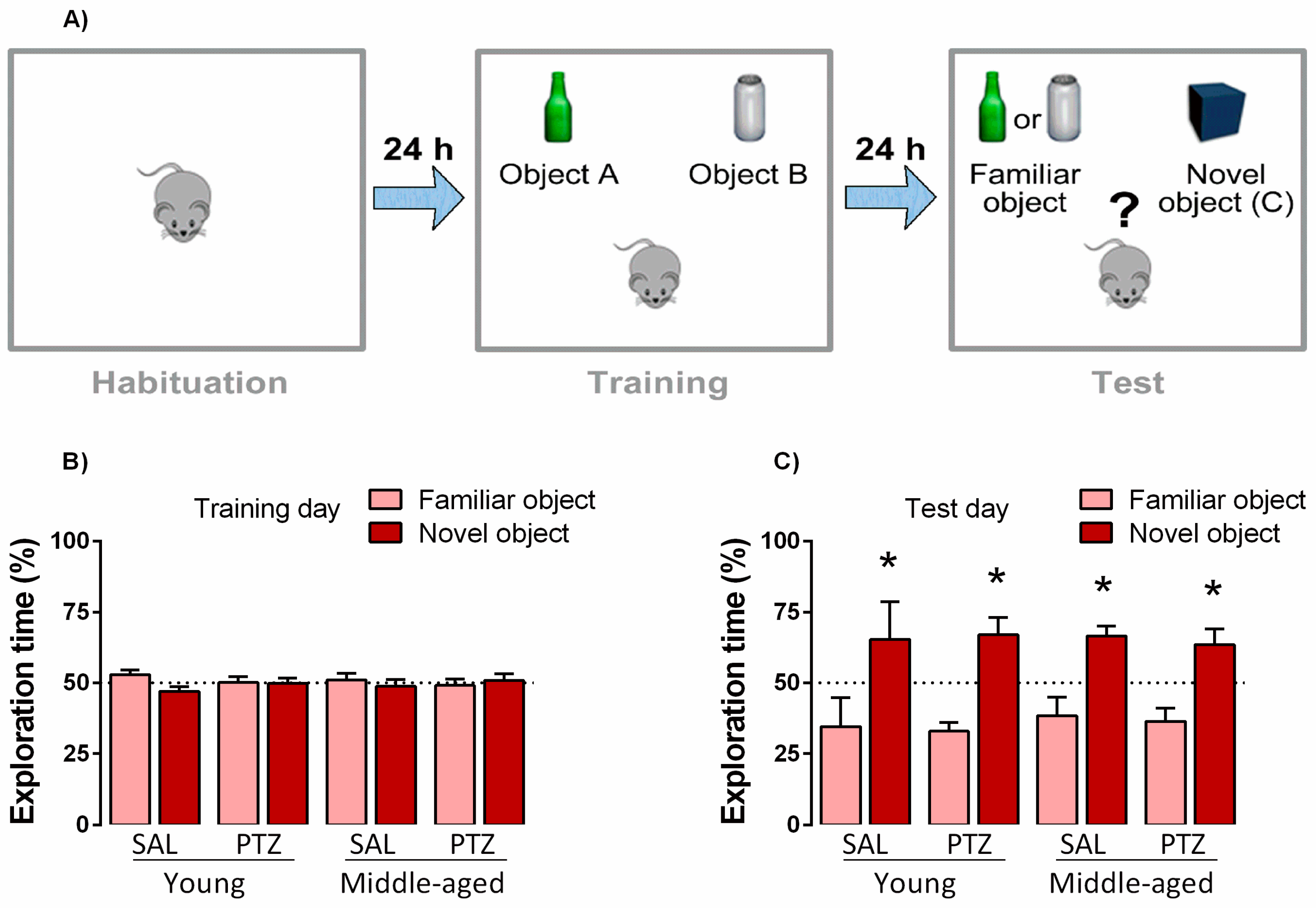
2.3. Object Recognition Memory Following Kindling Induction in Middle-Aged Rats
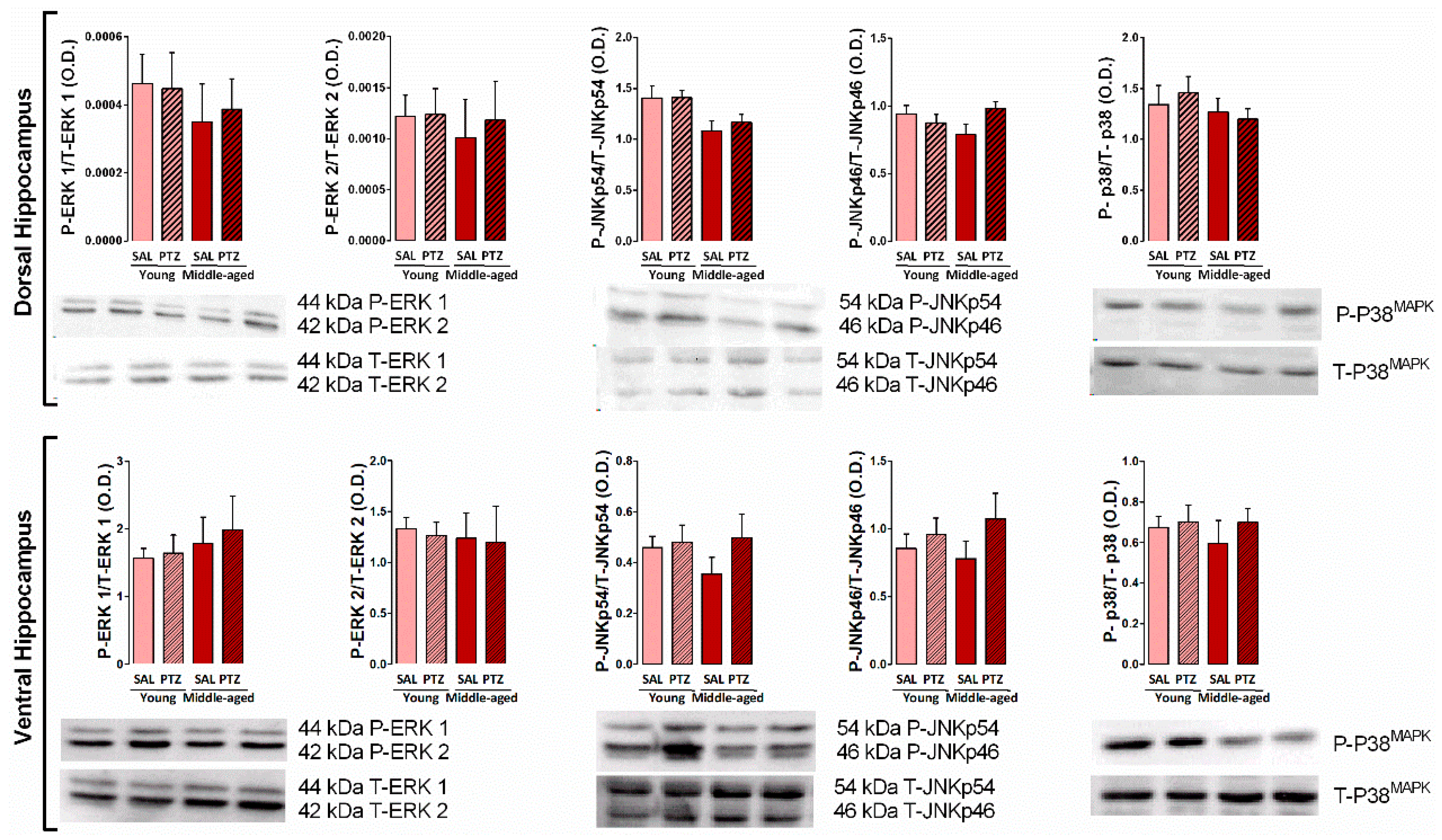
2.4. Hippocampal MAPKs Levels Following Kindling Induction in Middle-Aged Rats
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Drugs and Treatments
4.3. Experimental Procedures
4.3.1. Induction of Kindling
4.3.2. Elevated Plus-Maze (EPM)
4.3.3. The Object Recognition Task
4.3.4. Western Blotting
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fisher, R.S.; Boas, W.V.E.; Blume, W.; Elger, C.; Genton, P.; Lee, P.; Engel, J. Epileptic seizures and epilepsy: definitions proposed by the International League against Epilepsy (ILAE) and the International Bureau for Epilepsy (IBE). Epilepsia 2005, 46, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Hirtz, D.; Thurman, D.J.; Gwinn-Hardy, K.; Mohamed, M.; Chaudhuri, A.R.; Zalutsky, R. How common are the “common” neurologic disorders? Neurology 2007, 68, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Brodie, M.J.; Elder, A.T.; Kwan, P. Epilepsy in later life. Lancet Neurol. 2009, 8, 1019–1030. [Google Scholar] [CrossRef]
- Kelly, K.M. Aging Models of Acute Seizures and Epilepsy. Epilepsy Curr. 2010, 10, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Q.; Bell-Horner, C.L.; Dibas, M.I.; Covey, D.F.; Drewe, J.A.; Dillon, G.H. Pentylenetetrazole-induced inhibition of recombinant gamma-aminobutyric acid type A (GABA(A)) receptors: Mechanism and site of action. J. Pharmacol. Exp. Ther. 2001, 298, 986–995. [Google Scholar] [PubMed]
- Grecksch, G.; Becker, A.; Rauca, C. Effect of age on pentylenetetrazol-kindling and kindling-induced impairments of learning performance. Pharmacol. Biochem. Behav. 1997, 56, 595–601. [Google Scholar] [CrossRef]
- De Toledo-Morrell, L.; Morrell, F.; Fleming, S. Age-dependent deficits in spatial memory are related to impaired hippocampal kindling. Behav. Neurosci. 1984, 98, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, H.; Ramboiu, S.; Platt, D.; Herndon, J.G.; Kessler, C.; Popa-Wagner, A. Popa-Wagner. Age influences the expression of GAP-43 in the rat hippocampus following seizure. Gerontology 2005, 51, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, J.L.; Dell’Acqua, M.L. AKAP signaling complexes in regulation of excitatory synaptic plasticity. Neuroscientist 2011, 17, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.M.; Huganir, R.L. MAPK cascade signalling and synaptic plasticity. Nat. Rev. Neurosci. 2004, 5, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.W.; Soares, F.M.S.; de Mello, N.; Nunes, J.C.; de Cordova, F.M.; Walz, R.; Leal, R.B. Time-dependent modulation of mitogen activated protein kinases and AKT in rat hippocampus and cortex in the pilocarpine model of epilepsy. Neurochem. Res. 2012, 37, 1868–1878. [Google Scholar] [CrossRef] [PubMed]
- Ben, J.; Marques Gonçalves, F.; Alexandre Oliveira, P.; Vieira Peres, T.; Hohl, A.; Bainy Leal, R.; Abrão Cavalheiro, E.; Daniel Schroder Prediger, R.; Walz, R. Brain MAPKs levels are differentially associated with seizures threshold and severity progression in pentylenetetrazole-kindled mice. CNS Neurosci. Ther. 2013, 19, 726–729. [Google Scholar] [CrossRef] [PubMed]
- Stouffer, E.M.; Yoder, J.E. Middle-aged (12 month old) male rats show selective latent learning deficit. Neurobiol. Aging 2011, 32, e2311–e2324. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Goel, R.K. Age dependent learning and memory deficit in Pentylenetetrazol kindled mice. Eur. J. Pharmacol. 2012, 674, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Welsh, K.A.; Gold, P.E. Attenuation of epileptogenesis: Proactive effect of a single epinephrine injection of amygdaloid kindling. Behav. Neural. Biol. 1984, 40, 179–185. [Google Scholar] [CrossRef]
- File, S.E.; Mabbutt, P.S.; Becker, A.; Gonzalez, L.E.; Fluck, E. Short-term rebound anxiolytic effects and long-term changes in platelet benzodiazepine binding after pentylenetetrazole-kindling in two strains of rat. Anxiety 1996, 2, 109–116. [Google Scholar] [CrossRef]
- Szyndler, J.; Rok, P.; Maciejak, P.; Walkowiak, J.; Członkowska, A.I.; Sienkiewicz-Jarosz, H.; Wisłowska, A.; Zienowicz, M.; Lehner, M.; Bidziński, A.; et al. Effects of pentylenetetrazol-induced kindling of seizures on rat emotional behavior and brain monoaminergic systems. Pharmacol. Biochem. Behav. 2002, 73, 851–861. [Google Scholar] [CrossRef]
- Becker, A.; Grecksch, G.; Matthies, H. The influence of diazepam on learning processes impaired by pentylenetetrazol kindling. Naunyn Schmiedebergs Arch. Pharmacol. 1994, 349, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Hannesson, D.K.; Pollock, M.S.; Howland, J.G.; Mohapel, P.; Wallace, A.E.; Corcoran, M.E. Amygdaloid kindling is anxiogenic but fails to alter object recognition or spatial working memory in rats. Epilepsy Behav. 2008, 13, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.D.; Wang, Y.L.; Liang, S.F.; Shaw, F.Z. Rapid Amygdala Kindling Causes Motor Seizure and Comorbidity of Anxiety- and Depression-Like Behaviors in Rats. Front. Behav. Neurosci. 2016, 10, 129. [Google Scholar] [CrossRef]
- Altun, M.; Bergman, E.; Edström, E.; Johnson, H.; Ulfhake, B. Behavioral impairments of the aging rat. Physiol. Behav. 2007, 92, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.; Ackermann, R.F.; Engel, J. Chronic and single administration of pentylenetetrazol modifies benzodiazepine receptor-binding: an autoradiographic study. Epilepsy Res. 1996, 24, 65–72. [Google Scholar] [CrossRef]
- Gernert, M.; Richter, A.; Löscher, W. Subconvulsive dose of pentylenetetrazole increases the firing rate of substantia nigra pars reticulata neurons in dystonic but not in nondystonic hamsters. Synapse 1999, 33, 259–267. [Google Scholar] [CrossRef]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- Gámiz, F.; Gallo, M. Spontaneous object recognition memory in aged rats: Complexity versus similarity. Learn. Mem. 2012, 19, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Rushaidhi, M.; Zhang, H.; Liu, P. Effects of prolonged agmatine treatment in aged male Sprague-Dawley rats. Neuroscience 2013, 234, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.N.; Wallace, J.L.; Nematollahi, S.; Uprety, A.R.; Barnes, C.A. Pattern separation deficits may contribute to age-associated recognition impairments. Behav. Neurosci. 2010, 124, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Lamberty, Y.; Klitgaard, H. Consequences of pentylenetetrazole kindling on spatial memory and emotional responding in the rat. Epilepsy Behav. 2000, 1, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Adamec, R.; Shallow, T.; Burton, P. Anxiolytic and anxiogenic effects of kindling—Role of baseline anxiety and anatomical location of the kindling electrode in response to kindling of the right and left basolateral amygdala. Behav. Brain Res. 2005, 159, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Jokeit, H.; Ebner, A. Long term effects of refractory temporal lobe epilepsy on cognitive abilities: A cross sectional study. J. Neurol. Neurosurg. Psychiatry 1999, 67, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.D.; Dodrill, C.B.; Wilkus, R.J.; Ojemann, L.M.; Ojemann, G.A. Is partial epilepsy progressive? Ten-year follow-up of EEG and neuropsychological changes in adults with partial seizures. Epilepsia. 1998, 39, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Helmstaedter, C.; Kurthen, M.; Lux, S.; Reuber, M.; Elger, C.E. Chronic epilepsy and cognition: A longitudinal study in temporal lobe epilepsy. Ann. Neurol. 2003, 54, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Elger, C.E.; Helmstaedter, C.; Kurthen, M. Chronic epilepsy and cognition. Lancet Neurol. 2004, 3, 663–672. [Google Scholar] [CrossRef]
- Löscher, W. Animal models of drug-resistant epilepsy. Novartis Found. Symp. 2002, 243, 149–159. [Google Scholar] [PubMed]
- Wan, H.; Aggleton, J.P.; Brown, M.W. Different contributions of the hippocampus and perirhinal cortex to recognition memory. J. Neurosci. 1999, 19, 1142–1148. [Google Scholar] [PubMed]
- Schmoll, H.; Badan, I.; Grecksch, G.; Walker, L.; Kessler, C.; Popa-Wagner, A. Kindling status in sprague-dawley rats induced by pentylenetetrazole: Involvement of a critical development period. Am. J. Pathol. 2003, 162, 1027–1034. [Google Scholar] [CrossRef]
- Buga, A.M.; Vintilescu, R.; Balseanu, A.T.; Pop, O.T.; Streba, C.; Toescu, E.; Popa-Wagner, A. Repeated PTZ treatment at 25-day intervals leads to a highly efficient accumulation of doublecortin in the dorsal hippocampus of rats. PLoS ONE 2012, 7, e39302. [Google Scholar] [CrossRef] [PubMed]
- Bozon, B.; Kelly, A.; Josselyn, S.A.; Silva, A.J.; Davis, S.; Laroche, S. MAPK, CREB and zif268 are all required for the consolidation of recognition memory. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Blum, S.; Moore, A.N.; Adams, F.; Dash, P.K. A mitogen-activated protein kinase cascade in the CA1/CA2 subfield of the dorsal hippocampus is essential for long-term spatial memory. J. Neurosci. 1999, 19, 3535–3544. [Google Scholar]
- Ben, J.; de Oliveira, P.A.; Gonçalves, F.M.; Peres, T.V.; Matheus, F.C.; Hoeller, A.A.; Leal, R.B.; Walz, R.; Prediger, R.D. Effects of pentylenetetrazole kindling on mitogen-activated protein kinases levels in neocortex and hippocampus of mice. Neurochem. Res. 2014, 39, 2492–2500. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.W.; Aggleton, J.P. Recognition memory: What are the roles of the perirhinal cortex and hippocampus? Nat. Rev. Neurosci. 2001, 2, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Lüttjohann, A.; Fabene, P.F.; van Luijtelaar, G. A revised Racine’s scale for PTZ-induced seizures in rats. Physiol. Behav. 2009, 98, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.C.; Rodgers, R.J. Ethological evaluation of the effects of acute and chronic buspirone treatment in the murine elevated plus-maze test: Comparison with haloperidol. Psychopharmacology 1994, 114, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Rossato, J.I.; Bevilaqua, L.R.; Myskiw, J.C.; Medina, J.H.; Izquierdo, I.; Cammarota, M. On the role of hippocampal protein synthesis in the consolidation and reconsolidation of object recognition memory. Learn. Mem. 2007, 14, 36–46. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoeller, A.A.; De Carvalho, C.R.; Franco, P.L.C.; Formolo, D.A.; Imthon, A.K.; Dos Santos, H.R.; Eidt, I.; Souza, G.R.; Constantino, L.C.; Ferreira, C.L.; et al. Behavioral and Neurochemical Consequences of Pentylenetetrazol-Induced Kindling in Young and Middle-Aged Rats. Pharmaceuticals 2017, 10, 75. https://doi.org/10.3390/ph10030075
Hoeller AA, De Carvalho CR, Franco PLC, Formolo DA, Imthon AK, Dos Santos HR, Eidt I, Souza GR, Constantino LC, Ferreira CL, et al. Behavioral and Neurochemical Consequences of Pentylenetetrazol-Induced Kindling in Young and Middle-Aged Rats. Pharmaceuticals. 2017; 10(3):75. https://doi.org/10.3390/ph10030075
Chicago/Turabian StyleHoeller, Alexandre Ademar, Cristiane Ribeiro De Carvalho, Pedro Leite Costa Franco, Douglas Affonso Formolo, Alexandre Kracker Imthon, Henrique Rodighero Dos Santos, Ingrid Eidt, Gabriel Roman Souza, Leandra Celso Constantino, Camila Leite Ferreira, and et al. 2017. "Behavioral and Neurochemical Consequences of Pentylenetetrazol-Induced Kindling in Young and Middle-Aged Rats" Pharmaceuticals 10, no. 3: 75. https://doi.org/10.3390/ph10030075
APA StyleHoeller, A. A., De Carvalho, C. R., Franco, P. L. C., Formolo, D. A., Imthon, A. K., Dos Santos, H. R., Eidt, I., Souza, G. R., Constantino, L. C., Ferreira, C. L., Prediger, R. D., Bainy Leal, R., & Walz, R. (2017). Behavioral and Neurochemical Consequences of Pentylenetetrazol-Induced Kindling in Young and Middle-Aged Rats. Pharmaceuticals, 10(3), 75. https://doi.org/10.3390/ph10030075