
Investigating the Neural Mechanisms of Self-Controlled and Externally Controlled Movement with a Flexible Exoskeleton Using EEG Source Localization

 , , , , ,
, , , , ,
Abstract
Highlights
- Self-controlled tasks with flexEXO enhance activation in motor-related brain areas.
- Externally controlled tasks activate sensory feedback and error-monitoring regions.
- Self-controlled movement may better support motor learning and neurorehabilitation.
- flexEXO enables active engagement in motor training for users with limited hand function.
Abstract
1. Introduction
- A comprehensive review of the neural mechanisms underlying the differences between self-controlled and externally controlled movements, highlighting the potential benefits of self-controlled movements for motor learning and neurorehabilitation.
- An analysis of the role of motor imagery in enhancing neural plasticity and motor function in patients with neurological disorders, emphasizing the need for advanced assistive devices to optimize the benefits of motor imagery training.
- A discussion of the potential of advanced assistive devices, such as exoskeleton robots, in facilitating self-controlled movements and providing appropriate sensory feedback for neurorehabilitation.
2. Materials and Methods
2.1. Participants
2.2. Method
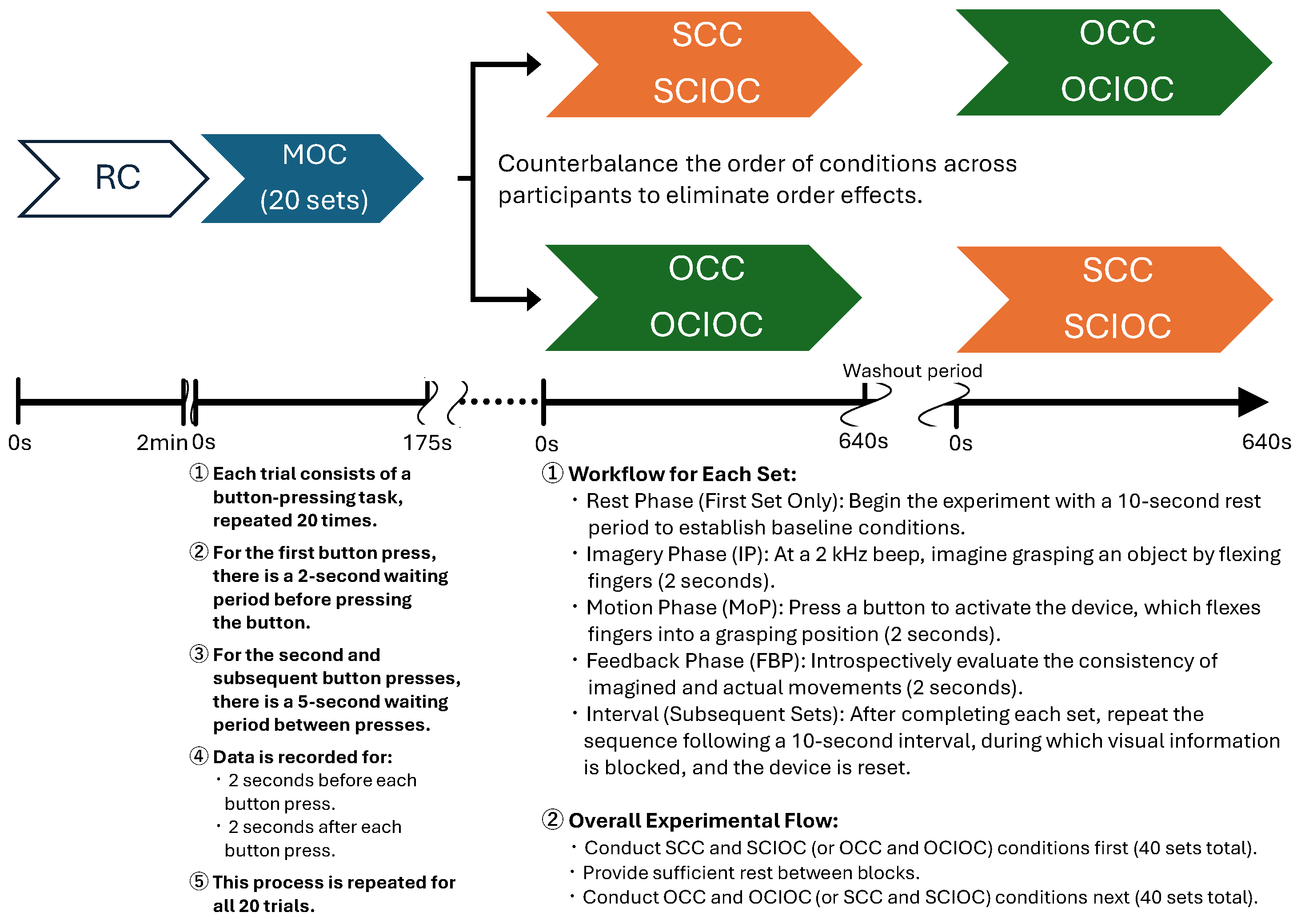
2.2.1. Experimental Procedure
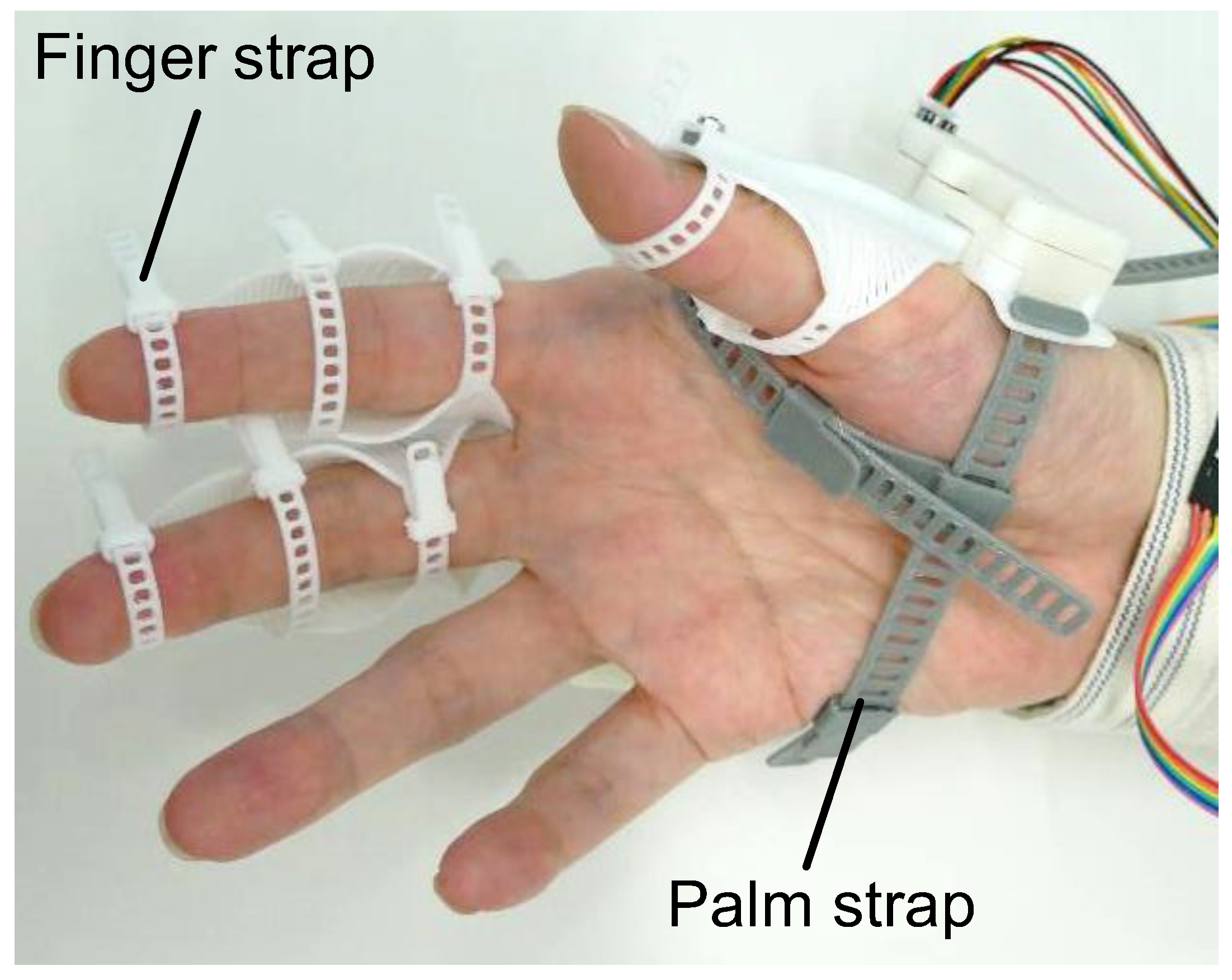
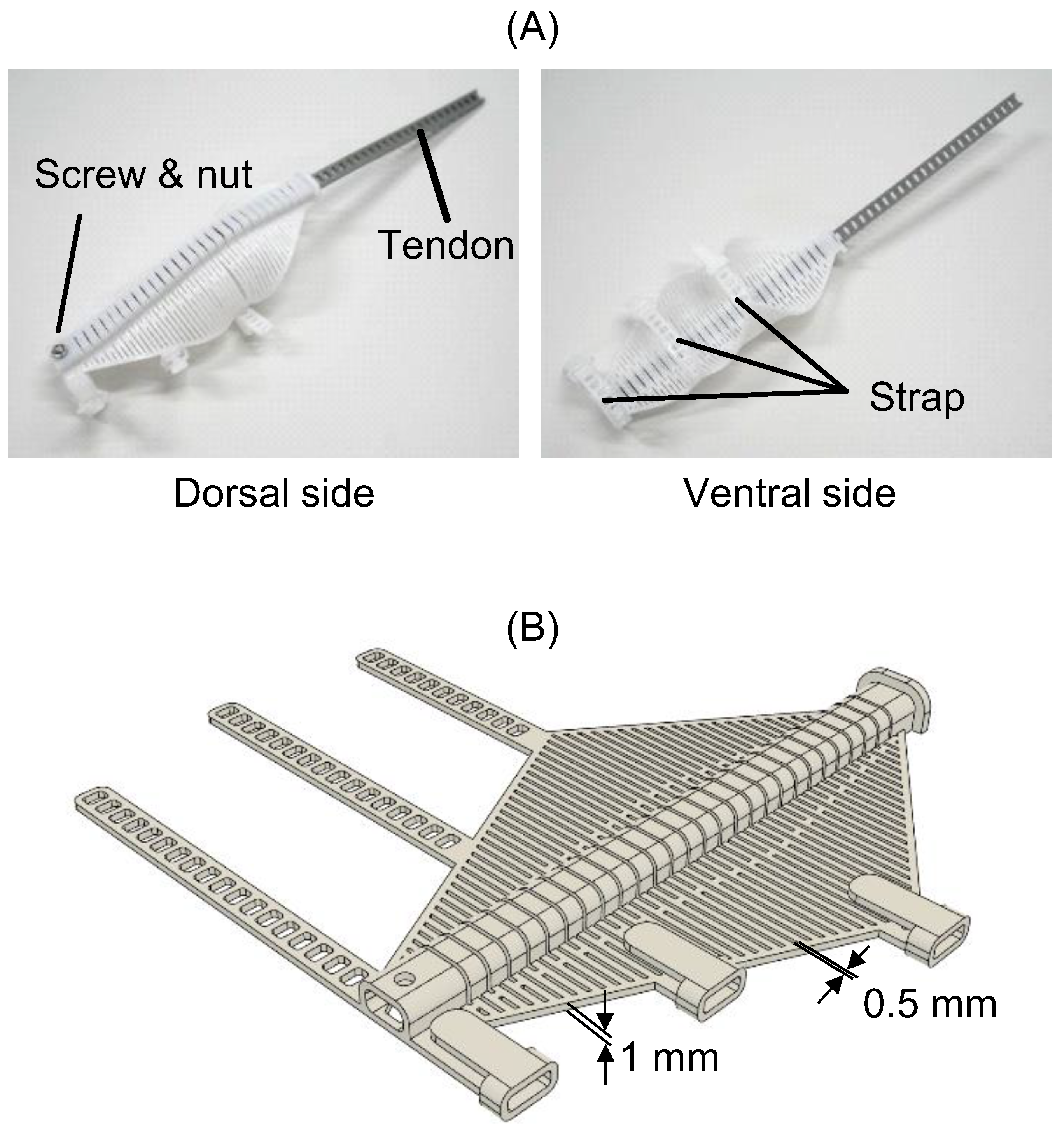
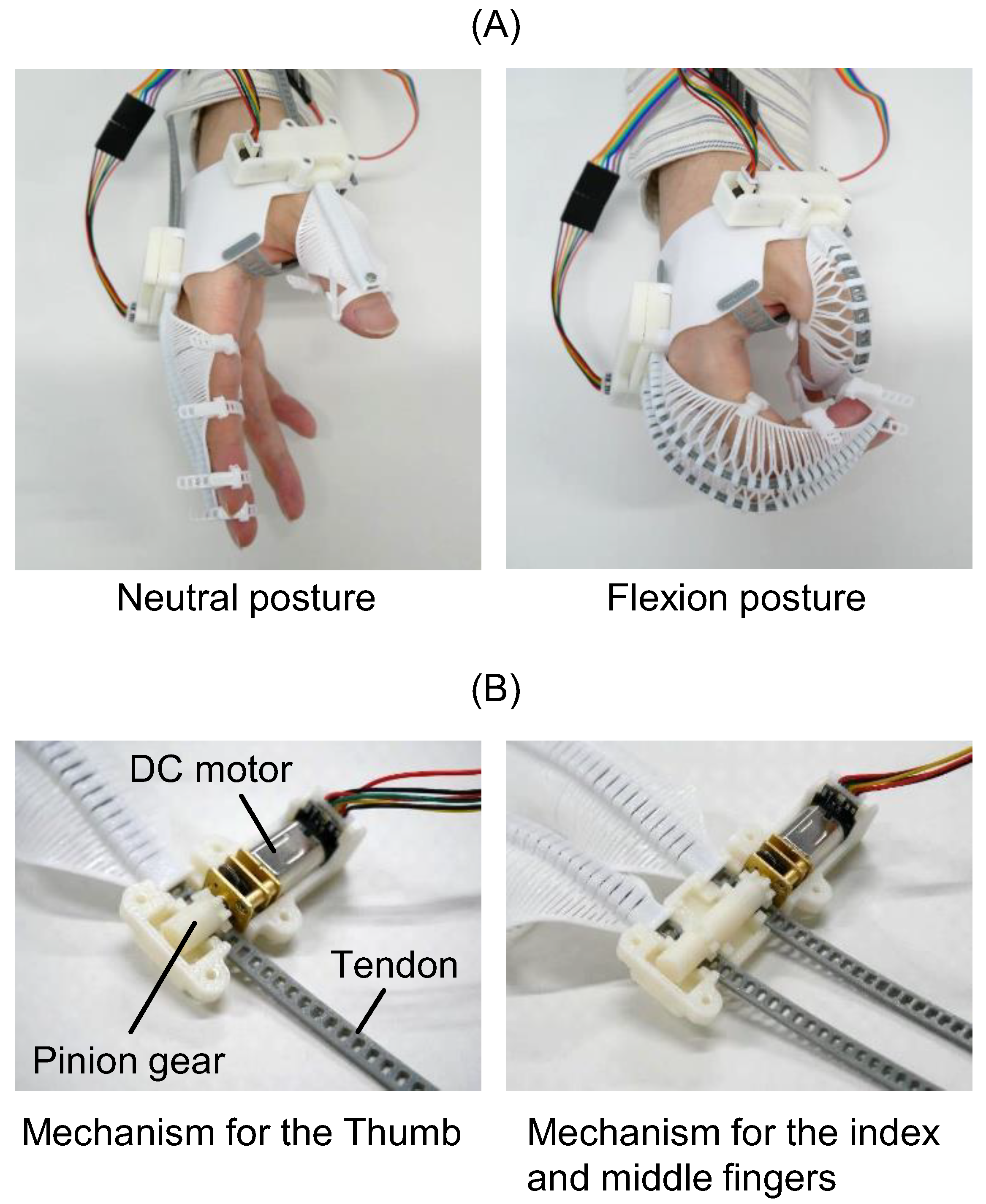
2.2.2. flexEXO
2.2.3. Experimental Conditions
- Rest Condition (RC)
- 2.
- Self-Controlled Condition (SCC)
- 3.
- Other-Controlled Condition (OCC)
- 4.
- Self-Controlled Imagery-Only Condition (SCIOC)
- 5.
- Other-Controlled Imagery-Only Condition (OCIOC)
- 6.
- Motion-Only Condition (MOC)
2.2.4. Evaluation of Brain Function Activity Using EEG
Electroencephalography
EEG Data Analysis
2.2.5. Statistical Analysis
2.3. Ethical Considerations
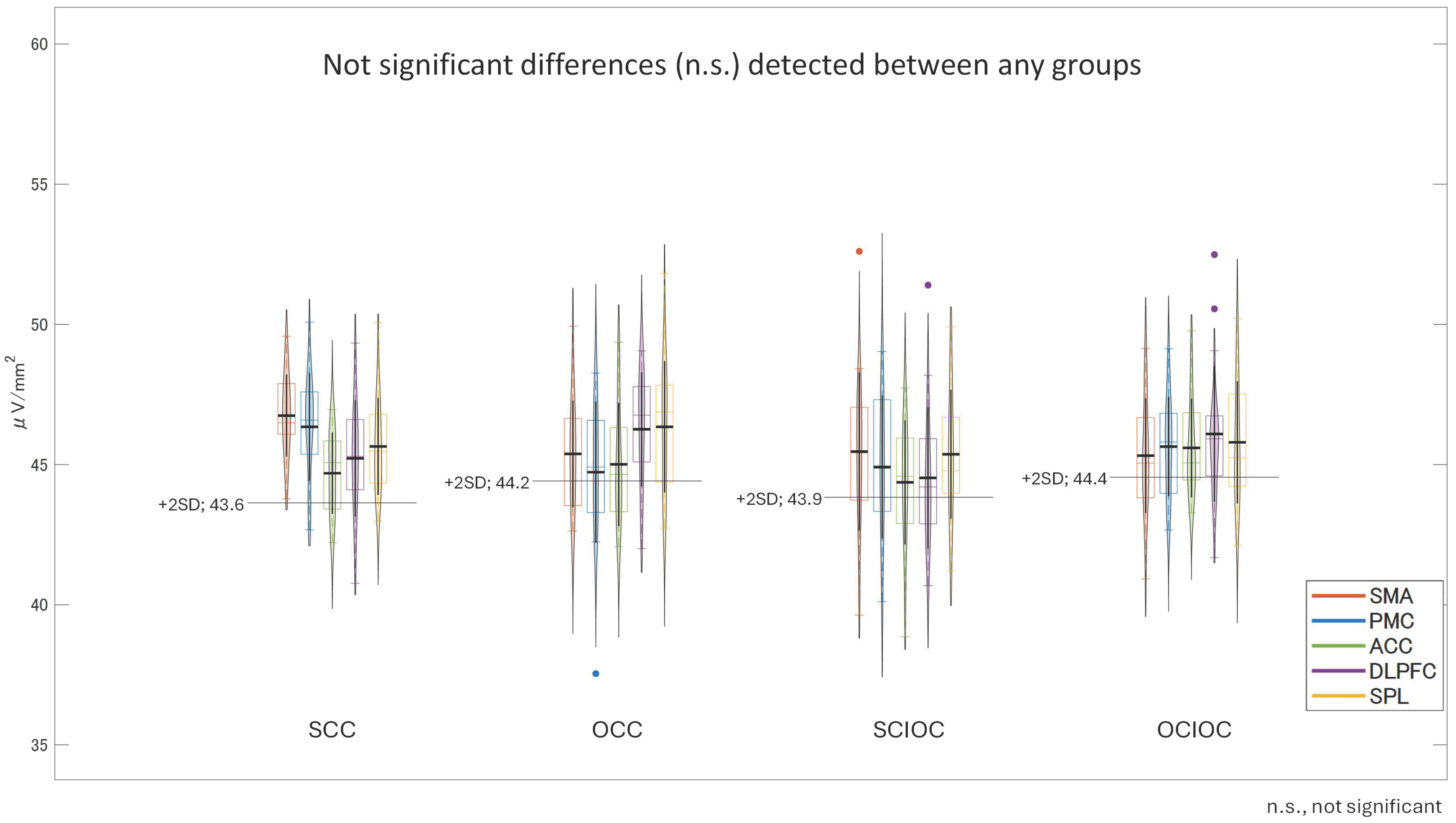
3. Results
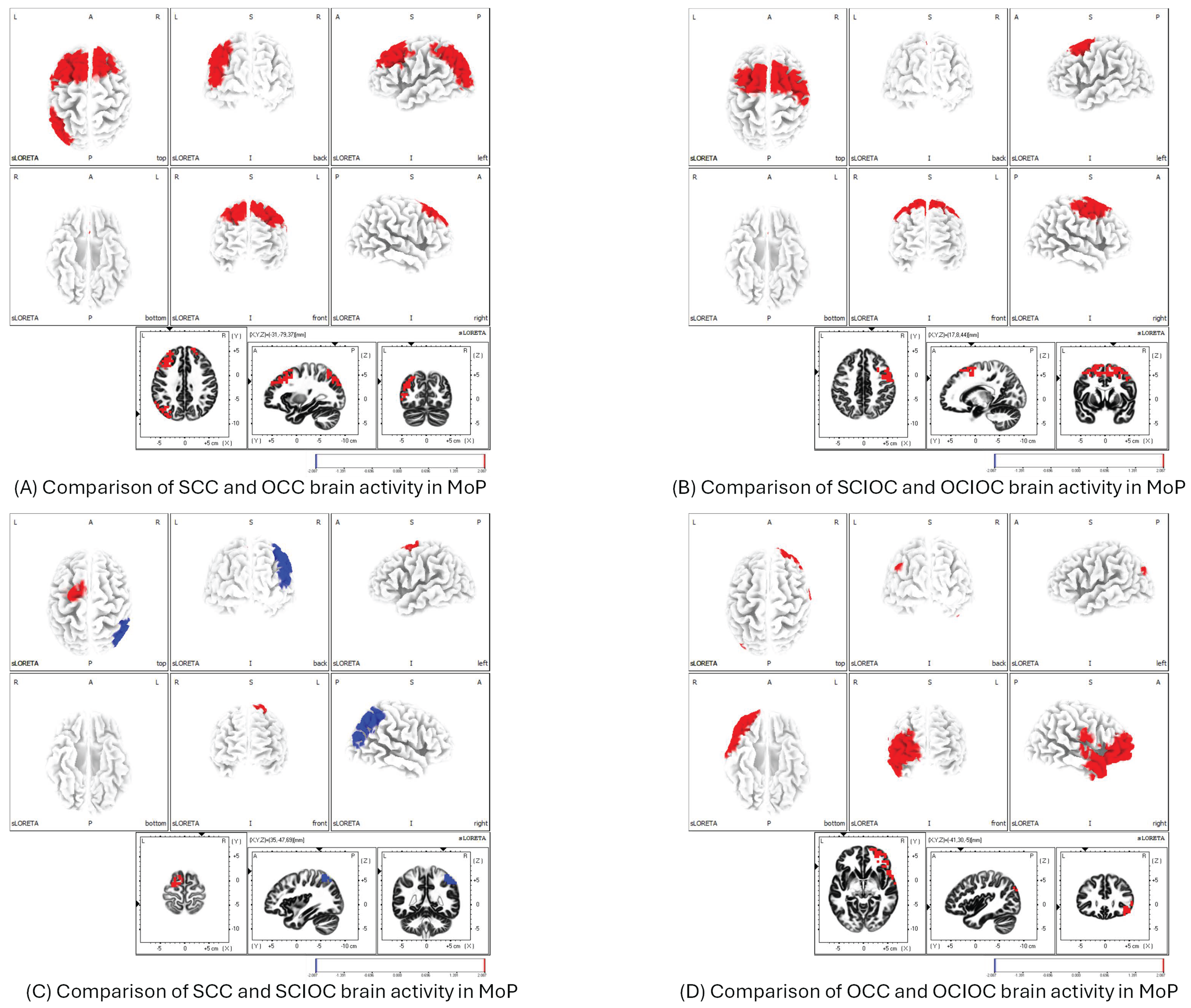
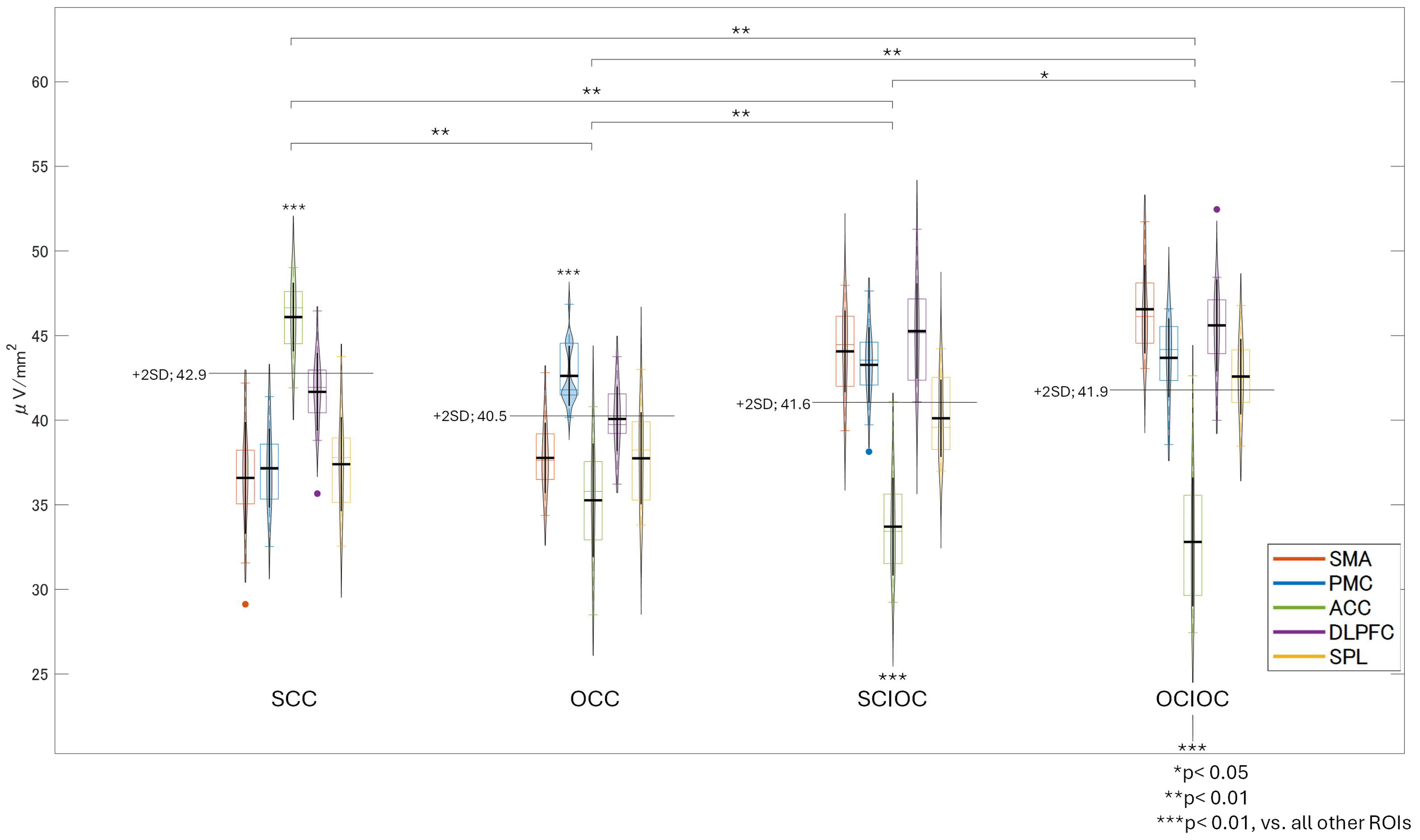
3.1. ERD and ERS During MI and ME
3.2. Brain Functional Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Abbreviation | Definition |
| MI | Motor Imagery |
| ME | Motor Execution |
| SCC | Self-Controlled Condition |
| OCC | Other-Controlled Condition |
| SCIOC | Self-Controlled Imagery-Only Condition |
| OCIOC | Other-Controlled Imagery-Only Condition |
| MOC | Motion-Only Condition |
| MoP | Motion Phase |
| IP | Imagery Phase |
| FBP | Feedback Phase |
| ERD | Event-Related Desynchronization |
| ERS | Event-Related Synchronization |
| EEG | Electroencephalography |
| M1 | Primary Motor Cortex |
| SMA | Supplementary Motor Area |
| PMC | Premotor Cortex |
| IPL | Inferior Parietal Lobule |
| S2 | Secondary Somatosensory Cortex |
| VLPFC | Ventrolateral Prefrontal Cortex |
| KVIQ | Kinesthetic and Visual Imagery Questionnaire |
| S/N | Signal-to-Noise |
| flexEXO | Flexible Exoskeleton |
References
- Jeannerod, M. Mental imagery in the motor context. Neuropsychologia 1995, 33, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Decety, J.; Grezes, J. Neural mechanisms subserving the perception of human actions. Trends Cogn. Sci. 1999, 3, 172–178. [Google Scholar] [CrossRef]
- Mulder, T. Motor imagery and action observation: Cognitive tools for rehabilitation. J. Neural Transm. 2007, 114, 1265–1278. [Google Scholar] [CrossRef]
- Schuster, C.; Hilfiker, R.; Amft, O.; Scheidhauer, A.; Andrews, B.; Butler, J.; Kischka, U.; Ettlin, T. Best practice for motor imagery: A systematic literature review on motor imagery training elements in five different disciplines. BMC Med. 2011, 9, 75. [Google Scholar] [CrossRef]
- Guillot, A.; Collet, C. Construction of the motor imagery integrative model in sport: A review and theoretical investigation of motor imagery use. Int. Rev. Sport Exerc. Psychol. 2008, 1, 31–44. [Google Scholar] [CrossRef]
- Lotze, M.; Halsband, U. Motor imagery. J. Physiol. 2006, 99, 386–395. [Google Scholar] [CrossRef]
- Sharma, N.; Pomeroy, V.M.; Baron, J.C. Motor imagery: A backdoor to the motor system after stroke? Stroke 2006, 37, 1941–1952. [Google Scholar] [CrossRef]
- Grezes, J.; Decety, J. Functional anatomy of execution, mental simulation, observation, and verb generation of actions: A meta-analysis. Hum. Brain Mapp. 2001, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hanakawa, T.; Immisch, I.; Toma, K.; Dimyan, M.A.; Van Gelderen, P.; Hallett, M. Functional properties of brain areas associated with motor execution and imagery. J. Neurophysiol. 2003, 89, 989–1002. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Flanagan, J.R. Motor prediction. Curr. Biol. 2001, 11, R729–R732. [Google Scholar] [CrossRef]
- Shadmehr, R.; Holcomb, H.H. Neural correlates of motor memory consolidation. Science 1997, 277, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, R.M.; Rottschy, C.; Miall, R.C.; Eickhoff, S.B. A quantitative meta-analysis and review of motor learning in the human brain. NeuroImage 2013, 67, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Nudo, R.J.; Wise, B.M.; SiFuentes, F.; Milliken, G.W. Neural substrates for the effects of rehabilitative training on motor recovery after ischemic infarct. Science 1996, 272, 1791–1794. [Google Scholar] [CrossRef]
- Kleim, J.A.; Jones, T.A. Principles of experience-dependent neural plasticity: Implications for rehabilitation after brain damage. J. Speech Lang. Hear. Res. 2008, 51, S225–S239. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, N.; Sasaki, A.; Yokoyama, H.; Masugi, Y.; Nakazawa, K. Effects of action observation and motor imagery of walking on the corticospinal and spinal motoneuron excitability and motor imagery ability in healthy participants. PLoS ONE 2022, 17, e0266000. [Google Scholar] [CrossRef]
- Marchal-Crespo, L.; Reinkensmeyer, D.J. Review of control strategies for robotic movement training after neurologic injury. J. NeuroEng. Rehabil. 2009, 6, 20. [Google Scholar] [CrossRef]
- Francis, S.; Lin, X.; Aboushoushah, S.; White, T.P.; Phillips, M.; Bowtell, R.; Constantinescu, C.S. fMRI analysis of active, passive and electrically stimulated ankle dorsiflexion. NeuroImage 2009, 44, 469–479. [Google Scholar] [CrossRef]
- Luft, A.R.; McCombe Waller, S.; Whitall, J.; Forrester, L.W.; Macko, R.; Sorkin, J.D.; Schulz, J.B.; Goldberg, A.P.; Hanley, D.F. Repetitive bilateral arm training and motor cortex activation in chronic stroke: A randomized controlled trial. JAMA 2004, 292, 1853–1861. [Google Scholar] [CrossRef]
- Hummel, F.C.; Cohen, L.G. Non-invasive brain stimulation: A new strategy to improve neurorehabilitation after stroke? Lancet Neurol. 2006, 5, 708–712. [Google Scholar] [CrossRef]
- Teasell, R.; Mehta, S.; Pereira, S.; McIntyre, A.; Janzen, S.; Allen, L.; Lobo, L.; Viana, R. Time to rethink long-term rehabilitation management of stroke patients. Top. Stroke Rehabil. 2012, 19, 457–462. [Google Scholar] [CrossRef]
- Derosiere, G.; Zénon, A.; Alamia, A.; Duque, J. Primary motor cortex contributes to the implementation of implicit value-based rules during motor decisions. NeuroImage 2017, 146, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Abbruzzese, G.; Marchese, R.; Avanzino, L.; Pelosin, E. Rehabilitation for Parkinson’s disease: Current outlook and future challenges. Park. Relat. Disord. 2016, 22, 60–64. [Google Scholar] [CrossRef]
- Lambert, K.J.; Hoar, C.; Houle, J.; Motley, C.; Ball, N.; Leung, A.W. Motor imagery as an intervention to improve activities of daily living post-stroke: A systematic review of randomized controlled trials. Br. J. Occup. Ther. 2023, 86, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Murguialday, A.; Broetz, D.; Rea, M.; Läer, L.; Yilmaz, O.; Brasil, F.L.; Liberati, G.; Curado, M.R.; Garcia-Cossio, E.; Vyziotis, A.; et al. Brain-machine interface in chronic stroke rehabilitation: A controlled study. Ann. Neurol. 2013, 74, 100–108. [Google Scholar] [CrossRef]
- Page, S.J.; Levine, P.; Leonard, A.C. Effects of mental practice on affected limb use and function in chronic stroke. Arch. Phys. Med. Rehabil. 2005, 86, 399–402. [Google Scholar] [CrossRef]
- Blakemore, S.J.; Wolpert, D.M.; Frith, C.D. Abnormalities in the awareness of action. Trends Cogn. Sci. 2002, 6, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Farrer, C.; Frey, S.H.; Van Horn, J.D.; Tunik, E.; Turk, D.; Inati, S.; Grafton, S.T. The angular gyrus computes action awareness representations. Cereb. Cortex 2008, 18, 254–261. [Google Scholar] [CrossRef]
- Miall, R.C.; Wolpert, D.M. Forward models for physiological motor control. Neural Netw. 1996, 9, 1265–1279. [Google Scholar] [CrossRef]
- Friston, K. The free-energy principle: A unified brain theory? Nat. Rev. Neurosci. 2010, 11, 127–138. [Google Scholar] [CrossRef]
- Grush, R. The emulation theory of representation: Motor control, imagery, and perception. Behav. Brain Sci. 2004, 27, 377–396. [Google Scholar] [CrossRef]
- Dobkin, B.H. Strategies for stroke rehabilitation. Lancet Neurol. 2004, 3, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.; Zhang, X.; Wang, J. Hand Rehabilitation Robotics on Poststroke Motor Recovery. Behav. Neurol. 2017, 2017, 3908135. [Google Scholar] [CrossRef] [PubMed]
- Zich, C.; Debener, S.; Kranczioch, C.; Bleichner, M.G.; Gutberlet, I.; De Vos, M. Real-time EEG feedback during simultaneous EEG-fMRI identifies the cortical signature of motor imagery. NeuroImage 2015, 114, 438–447. [Google Scholar] [CrossRef]
- Pfurtscheller, G.; Lopes da Silva, F.H. Event-related EEG/MEG synchronization and desynchronization: Basic principles. Clin. Neurophysiol. 1999, 110, 1842–1857. [Google Scholar] [CrossRef]
- Neuper, C.; Wörtz, M.; Pfurtscheller, G. ERD/ERS patterns reflecting sensorimotor activation and deactivation. Prog. Brain Res. 2006, 159, 211–222. [Google Scholar] [PubMed]
- Cunnington, R.; Windischberger, C.; Deecke, L.; Moser, E. The preparation and execution of self-initiated and externally-triggered movement: A study of event-related fMRI. NeuroImage 2002, 15, 373–385. [Google Scholar] [CrossRef]
- Harwin, W.S.; Patton, J.L.; Edgerton, V.R. Challenges and opportunities for robot-mediated neurorehabilitation. Proc. IEEE 2006, 94, 1717–1726. [Google Scholar] [CrossRef]
- Mehrholz, J.; Hädrich, A.; Platz, T.; Kugler, J.; Pohl, M. Electromechanical and robot-assisted arm training for improving arm function and activities of daily living after stroke. Cochrane Database Syst. Rev. 2018, 9, CD006876. [Google Scholar] [CrossRef]
- Maciejasz, P.; Eschweiler, J.; Gerlach-Hahn, K.; Jansen-Toy, L.B.; Leonhardt, S. A survey on robotic devices for upper limb rehabilitation. J. NeuroEng. Rehabil. 2014, 11, 3. [Google Scholar] [CrossRef]
- Polygerinos, P.; Galloway, K.C.; Savage, E.; Herman, M.; O’Donnell, K.; Walsh, C.J. Soft robotic glove for hand rehabilitation and task specific training. In Proceedings of the IEEE International Conference on Robotics and Automation (ICRA), Seattle, WA, USA, 26–30 May 2015; pp. 2913–2919. [Google Scholar]
- Polygerinos, P.; Wang, Z.; Galloway, K.C.; Wood, R.J.; Walsh, C.J. Soft robotic glove for combined assistance and at-home rehabilitation. Robot. Auton. Syst. 2015, 73, 135–143. [Google Scholar] [CrossRef]
- Yap, H.K.; Lim, J.H.; Nasrallah, F.; Goh, J.C.H.; Yeow, R.C.H. A soft exoskeleton for hand assistive and rehabilitation application using pneumatic actuators with variable stiffness. In Proceedings of the IEEE International Conference on Robotics and Automation (ICRA), Seattle, WA, USA, 26–30 May 2015; pp. 4967–4972. [Google Scholar]
- Hampson, E. A brief guide to the menstrual cycle and oral contraceptive use for researchers in behavioral endocrinology. Horm. Behav. 2020, 119, 104655. [Google Scholar] [CrossRef] [PubMed]
- Malouin, F.; Richards, C.L.; Jackson, P.L.; Lafleur, M.F.; Durand, A.; Doyon, J. The Kinesthetic and Visual Imagery Questionnaire (KVIQ) for Assessing Motor Imagery in Persons with Physical Disabilities: A reliability and Construct Validity Study. J. Neurol. Phys. Ther. 2007, 31, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Vigouroux, L.; Domalain, M.; Berton, E. Effect of object width on muscle and joint forces during thumb-index finger grasping. J. Appl. Biomech. 2011, 27, 173–180. [Google Scholar] [CrossRef]
- Kong, Y.-K.; Lowe, B.D. Optimal cylindrical handle diameter for grip force tasks. Int. J. Ind. Ergon. 2005, 35, 495–507. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Ogawa, K.; Yamanaka, S.; Kawashima, N. Finch: Prosthetic Arm with Three Opposing Fingers Controlled by a Muscle Bulge. IEEE Trans. Neural Syst. Rehab. Eng. 2022, 31, 377–386. [Google Scholar] [CrossRef]
- Minamii, K.; Yoshikawa, M. A Hand Exoskeleton with 3D-Printed Compliant Mechanisms to Assist Grasping. In Proceedings of the 2024 46th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Orlando, FL, USA, 15–19 July 2024; pp. 1–4. [Google Scholar]
- Filgueiras, A.; Conde, E.F.Q.; Hall, C.R. The neural basis of kinesthetic and visual imagery in sports: An ALE meta−analysis. Brain Imaging Behav. 2018, 12, 1513–1523. [Google Scholar] [CrossRef]
- Munzert, J.; Lorey, B.; Zentgraf, K. Cognitive motor processes: The role of motor imagery in the study of motor representations. Brain Res. Rev. 2009, 60, 306–326. [Google Scholar] [CrossRef] [PubMed]
- Papaxanthis, C.; Pozzo, T.; Skoura, X.; Schieppati, M. Does order and timing in performance of imagined and actual movements affect the motor imagery process? The duration of walking and writing task. Behav. Brain Res. 2002, 134, 209–215. [Google Scholar] [CrossRef]
- Nakayashiki, K.; Saeki, M.; Takata, Y.; Hayashi, Y.; Kondo, T. Modulation of event-related desynchronization during kinematic and kinetic hand movements. J. NeuroEng. Rehabil. 2014, 11, 90. [Google Scholar] [CrossRef]
- Jasper, H.H. The ten-twenty electrode system of the International Federation. Electroencephalogr. Clin. Neurophysiol. 1958, 10, 370–375. [Google Scholar]
- Luck, S.J. An Introduction to the Event-Related Potential Technique; MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.P.; Makeig, S.; Westerfield, M.; Townsend, J.; Courchesne, E.; Sejnowski, T.J. Removal of eye activity artifacts from visual event-related potentials in normal and clinical subjects. Clin. Neurophysiol. 2000, 111, 1745–1758. [Google Scholar] [CrossRef] [PubMed]
- Makeig, S.; Jung, T.P.; Bell, A.J.; Ghahremani, D.; Sejnowski, T.J. Blind separation of auditory event-related brain responses into independent components. Proc. Natl. Acad. Sci. USA 1997, 94, 10979–10984. [Google Scholar] [CrossRef] [PubMed]
- Urigüen, J.A.; Garcia-Zapirain, B. EEG artifact removal—State-of-the-art and guidelines. J. Neural Eng. 2015, 12, 031001. [Google Scholar] [CrossRef]
- Pfurtscheller, G.; Neuper, C. Motor imagery activates primary sensorimotor area in humans. Neurosci. Lett. 1997, 239, 65–68. [Google Scholar] [CrossRef]
- Fatourechi, M.; Bashashati, A.; Ward, R.K.; Birch, G.E. EMG and EOG artifacts in brain computer interface systems: A survey. Clin. Neurophysiol. 2007, 118, 480–494. [Google Scholar] [CrossRef]
- Keinrath, C.; Wriessnegger, S.; Müller-Putz, G.R.; Pfurtscheller, G. Post-movement beta synchronization after kinesthetic illusion, active and passive movements. Int. J. Psychophysiol. 2006, 62, 321–327. [Google Scholar] [CrossRef]
- Pfurtscheller, G. Functional brain imaging based on ERD/ERS. Vis. Res. 2001, 41, 1257–1260. [Google Scholar] [CrossRef]
- Formaggio, E.; Storti, S.F.; Cerini, R.; Fiaschi, A.; Manganotti, P. Brain oscillatory activity during motor imagery in EEG-fMRI coregistration. Magn. Reson. Imaging 2010, 28, 1403–1412. [Google Scholar] [CrossRef]
- Kranczioch, C.; Mathews, S.; Dean, P.J.; Sterr, A. On the equivalence of executed and imagined movements: Evidence from lateralized motor and nonmotor potentials. Hum. Brain Mapp. 2009, 30, 3275–3286. [Google Scholar] [CrossRef]
- Alegre, M.; Labarga, A.; Gurtubay, I.G.; Iriarte, J.; Malanda, A.; Artieda, J. Beta electroencephalograph changes during passive movements: Sensory afferences contribute to beta event-related desynchronization in humans. Neurosci. Lett. 2002, 331, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.R.; Neuper, C.; Rupp, R.; Keinrath, C.; Gerner, H.J.; Pfurtscheller, G. Event-related beta EEG changes during wrist movements induced by functional electrical stimulation of forearm muscles in man. Neurosci. Lett. 2003, 340, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.K.; Fries, P. Beta-band oscillations—Signalling the status quo? Curr. Opin. Neurobiol. 2010, 20, 156–165. [Google Scholar] [CrossRef]
- Spitzer, B.; Haegens, S. Beyond the status quo: A role for beta oscillations in endogenous content (re)activation. eNeuro 2017, 4, ENEURO.0170-17.2017. [Google Scholar] [CrossRef]
- Zaepffel, M.; Trachel, R.; Kilavik, B.E.; Brochier, T. Modulations of EEG beta power during planning and execution of grasping movements. PLoS ONE 2013, 8, e60060. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find. Exp. Clin. Pharmacol. 2002, 24 (Suppl. D), 5–12. [Google Scholar] [PubMed]
- Pascual-Marqui, R.D. Discrete, 3D distributed, linear imaging methods of electric neuronal activity. Part 1: Exact, zero error localization. arXiv 2007, arXiv:0710.3341. [Google Scholar]
- Olbrich, S.; Mulert, C.; Karch, S.; Trenner, M.; Leicht, G.; Pogarell, O.; Hegerl, U. EEG vigilance and BOLD effect during simultaneous EEG/fMRI measurement. Neuroimage 2009, 45, 319–332. [Google Scholar] [CrossRef]
- Michel, C.M.; Brunet, D. EEG Source Imaging: A Practical Review of the Analysis Steps. Front. Neurol. 2019, 10, 325. [Google Scholar] [CrossRef]
- Nichols, T.E.; Holmes, A.P. Nonparametric permutation tests for functional neuroimaging: A primer with examples. Hum. Brain Mapp. 2002, 15, 1–25. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 1988. [Google Scholar]
- Pascual-Marqui, R.D.; Esslen, M.; Kochi, K.; Lehmann, D. Functional imaging with low-resolution brain electromagnetic tomography (LORETA): A review. Methods Find. Exp. Clin. Pharmacol. 2002, 24 (Suppl. C), 91–95. [Google Scholar] [PubMed]
- Espenhahn, S.; de Berker, A.O.; van Wijk, B.C.M.; Rossiter, H.E.; Ward, N.S. Movement-related beta oscillations show high intra-individual reliability. Neuroimage 2017, 147, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Haggard, P.; Clark, S.; Kalogeras, J. Voluntary action and conscious awareness. Nat. Neurosci. 2002, 5, 382–385. [Google Scholar] [CrossRef]
- Neuper, C.; Pfurtscheller, G. Event-related dynamics of cortical rhythms: Frequency-specific features and functional correlates. Int. J. Psychophysiol. 2001, 43, 41–58. [Google Scholar] [CrossRef]
- Haggard, P. Human volition: Towards a neuroscience of will. Nat. Rev. Neurosci. 2008, 9, 934–946. [Google Scholar] [CrossRef]
- Nachev, P.; Kennard, C.; Husain, M. Functional role of the supplementary and pre-supplementary motor areas. Nat. Rev. Neurosci. 2008, 9, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Blakemore, S.J.; Sirigu, A. Action prediction in the cerebellum and in the parietal lobe. Exp. Brain Res. 2003, 153, 239–245. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Amunts, K.; Mohlberg, H.; Zilles, K. The human parietal operculum. II. Stereotaxic maps and correlation with functional imaging results. Cereb. Cortex 2008, 16, 268–279. [Google Scholar] [CrossRef]
- Tanji, J.; Hoshi, E. Role of the lateral prefrontal cortex in executive behavioral control. Physiol. Rev. 2008, 88, 37–57. [Google Scholar] [CrossRef]
- Crucianelli, L.; Krahé, C.; Jenkinson, P.M.; Fotopoulou, A. Interoceptive ingredients of body ownership: Affective touch and cardiac awareness in the rubber hand illusion. Cortex 2018, 104, 180–192. [Google Scholar] [CrossRef]
- Marchal-Crespo, L.; Rappo, N.; Riener, R. The effectiveness of robotic training depends on motor task characteristics. Exp. Brain Res. 2017, 235, 3799–3816. [Google Scholar] [CrossRef]
- Ullsperger, M.; Cramon, D.Y.; Müller, N.G. Interactions of focal cortical lesions with error processing: Clinical and experimental evidence. Front. Hum. Neurosci. 2014, 8, 410. [Google Scholar]
- Craig, A.D. How do you feel—Now? The anterior insula and human awareness. Nat. Rev. Neurosci. 2009, 10, 59–70. [Google Scholar] [CrossRef]
- Botvinick, M.M.; Cohen, J.D.; Carter, C.S. Conflict monitoring and anterior cingulate cortex: An update. Trends Cogn. Sci. 2004, 8, 539–546. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, A.W.; Cohen, J.D.; Stenger, V.A.; Carter, C.S. Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science 2000, 288, 1835–1838. [Google Scholar] [CrossRef]
- Ang, K.K.; Guan, C.; Phua, K.S.; Wang, C.; Zhou, L.; Tang, K.Y.; Ephraim Joseph, G.J.; Kuah, C.W.; Chua, K.S. Brain-computer interface-based robotic end effector system for wrist and hand rehabilitation: Results of a three-armed randomized controlled trial for chronic stroke. Front. Neuroeng. 2014, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Do, J.; Lim, W.T.; Kim, D.Y.; Ko, E.J.; Ko, M.H.; Kim, G.W.; Kim, J.H.; Kim, S.; Kim, H. Effects of high-intensity interval robot-assisted gait training on cardiopulmonary function and walking ability in chronic stroke survivors: A multicenter single-blind randomized controlled trial. J. Back Musculoskelet. Rehabil. 2024, 37, 1309–1319. [Google Scholar] [CrossRef]
- Cappello, L.; Meyer, J.T.; Galloway, K.C.; Peisner, J.D.; Granberry, R.; Wagner, D.A.; Engelhardt, S.; Paganoni, S.; Walsh, C.J. Assisting hand function after spinal cord injury with a fabric-based soft robotic glove. J. NeuroEng. Rehabil. 2018, 15, 59. [Google Scholar] [CrossRef]
- Yun, Y.; Dancausse, S.; Esmatloo, P.; Serrato, A.; Merring, C.A.; Agarwal, P.; Deshpande, A.D. Maestro: An EMG-driven assistive hand exoskeleton for spinal cord injury patients. In Proceedings of the 2017 IEEE International Conference on Robotics and Automation (ICRA), Singapore, 29 May–3 June 2017; pp. 2904–2910. [Google Scholar]
- Aarts, H.; van den Bos, K. On the foundations of beliefs in free will: Intentional binding and unconscious priming in self-agency. Psychol. Sci. 2012, 23, 532–537. [Google Scholar] [CrossRef]
- Bandura, A. Toward a psychology of human agency: Pathways and reflections. Perspect. Psychol. Sci. 2018, 13, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Yamashita, A.; Asama, H. The influence of performance on action-effect integration in sense of agency. Conscious. Cogn. 2017, 53, 89–98. [Google Scholar] [CrossRef] [PubMed]

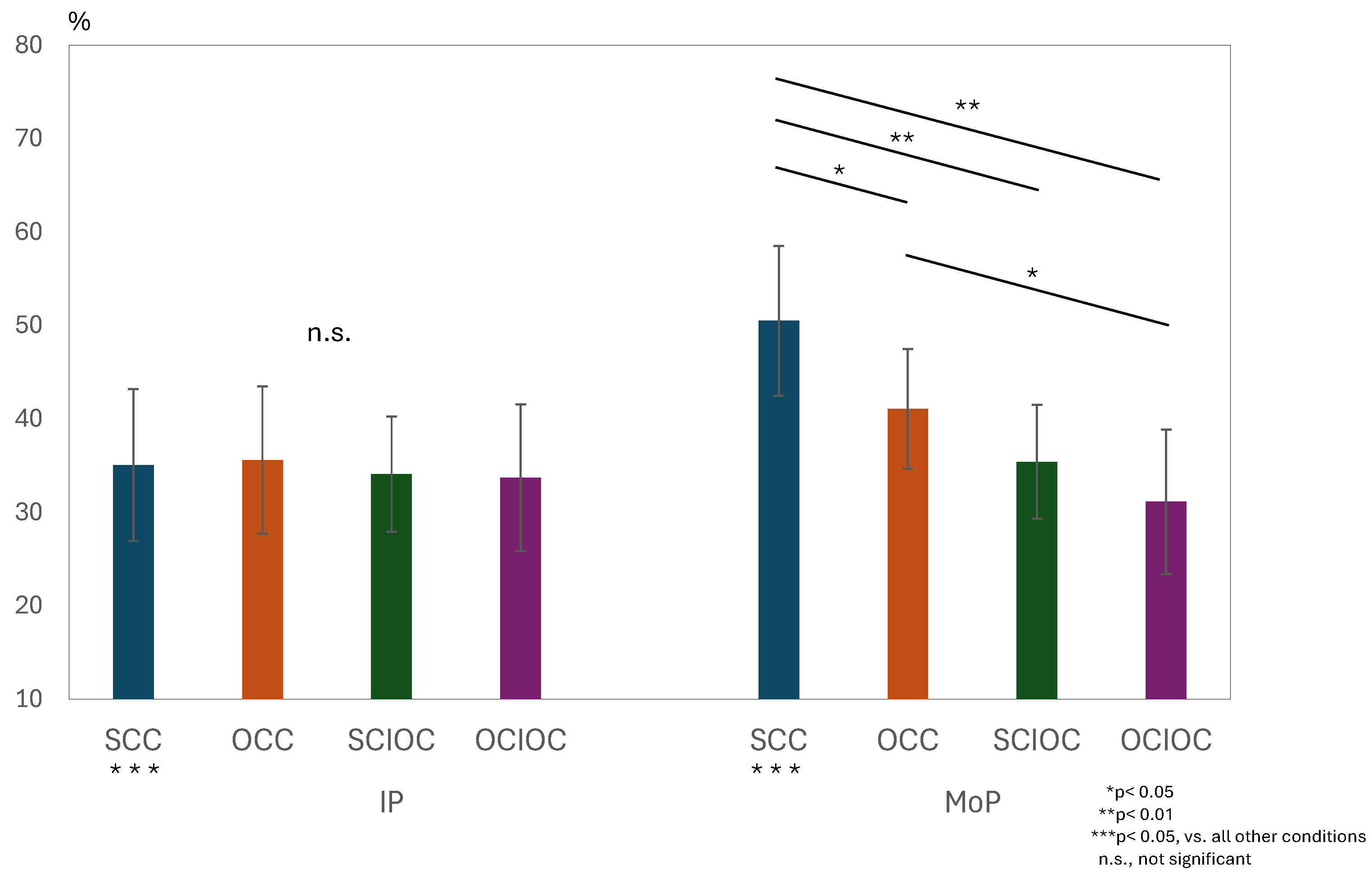
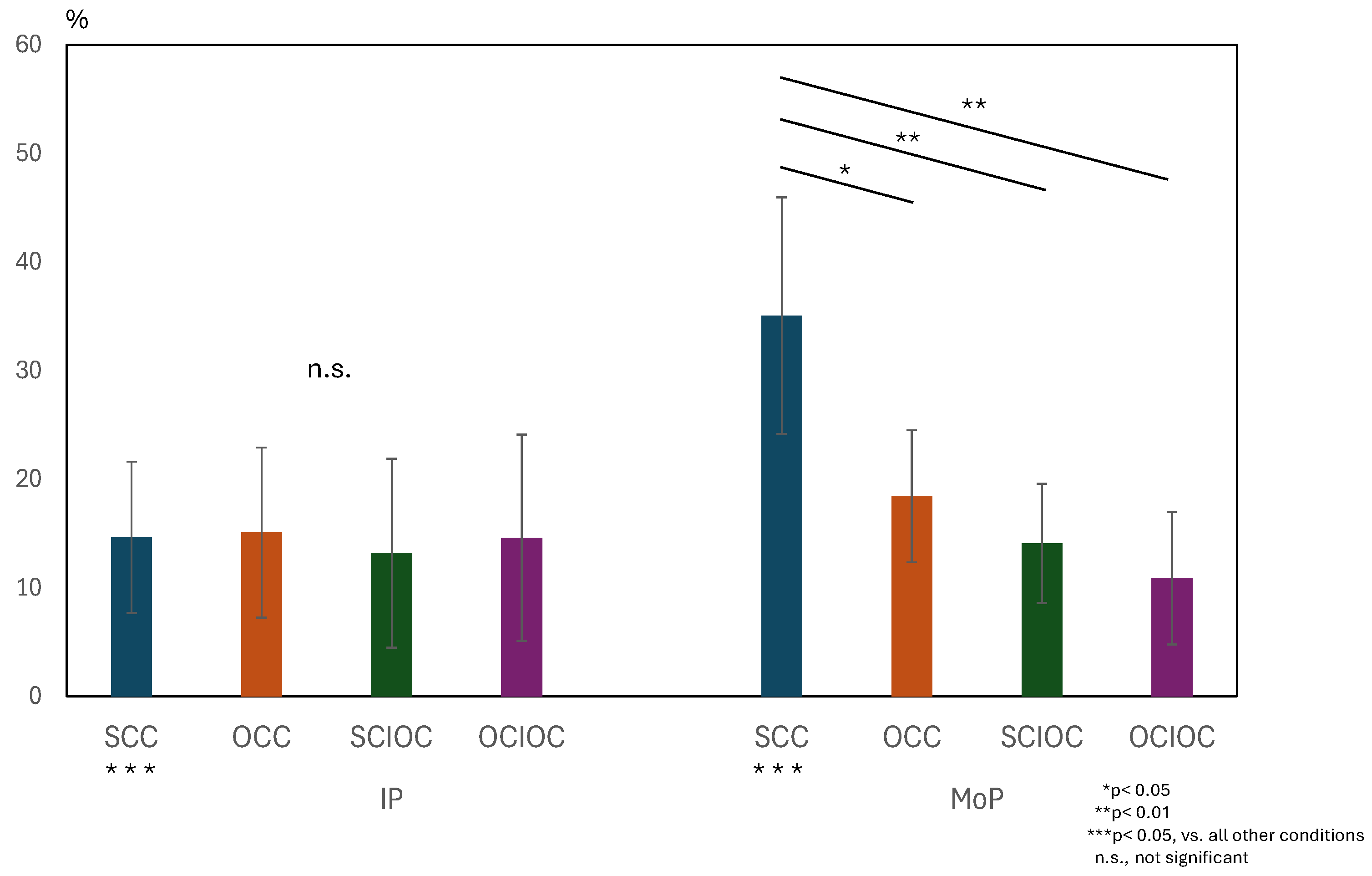
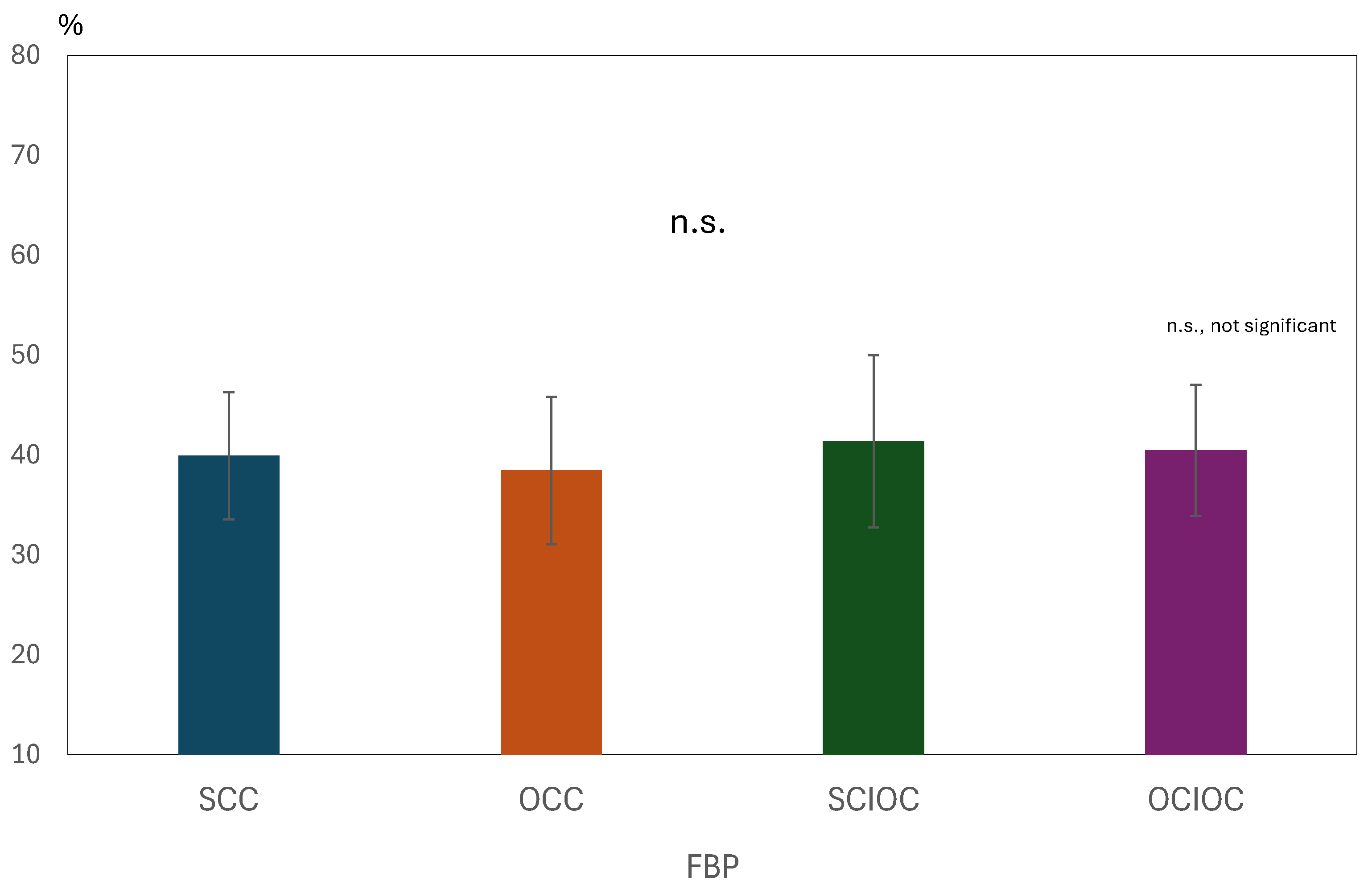

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Mean ± SD |
|---|---|
| Age (years) | 22.9 ± 2.0 |
| Height (cm) | 170.4 ± 3.8 |
| Weight (kg) | 63.2 ± 6.7 |
| Dominant hand (Right/Left) | 25/0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kodama, T.; Yoshikawa, M.; Minamii, K.; Nishimoto, K.; Kadowaki, S.; Inoue, Y.; Ito, H.; Shigeto, H.; Okuyama, K.; Maeda, K.; et al. Investigating the Neural Mechanisms of Self-Controlled and Externally Controlled Movement with a Flexible Exoskeleton Using EEG Source Localization. Sensors 2025, 25, 3527. https://doi.org/10.3390/s25113527
Kodama T, Yoshikawa M, Minamii K, Nishimoto K, Kadowaki S, Inoue Y, Ito H, Shigeto H, Okuyama K, Maeda K, et al. Investigating the Neural Mechanisms of Self-Controlled and Externally Controlled Movement with a Flexible Exoskeleton Using EEG Source Localization. Sensors. 2025; 25(11):3527. https://doi.org/10.3390/s25113527
Chicago/Turabian StyleKodama, Takayuki, Masahiro Yoshikawa, Kosuke Minamii, Kazuhei Nishimoto, Sayuna Kadowaki, Yuuki Inoue, Hiroki Ito, Hayato Shigeto, Kohei Okuyama, Kouta Maeda, and et al. 2025. "Investigating the Neural Mechanisms of Self-Controlled and Externally Controlled Movement with a Flexible Exoskeleton Using EEG Source Localization" Sensors 25, no. 11: 3527. https://doi.org/10.3390/s25113527
APA StyleKodama, T., Yoshikawa, M., Minamii, K., Nishimoto, K., Kadowaki, S., Inoue, Y., Ito, H., Shigeto, H., Okuyama, K., Maeda, K., Katayama, O., Murata, S., & Morita, K. (2025). Investigating the Neural Mechanisms of Self-Controlled and Externally Controlled Movement with a Flexible Exoskeleton Using EEG Source Localization. Sensors, 25(11), 3527. https://doi.org/10.3390/s25113527