
Sequence Effect on the Activity of DNAzyme with Covalently Attached Hemin and Their Potential Bioanalytical Application

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
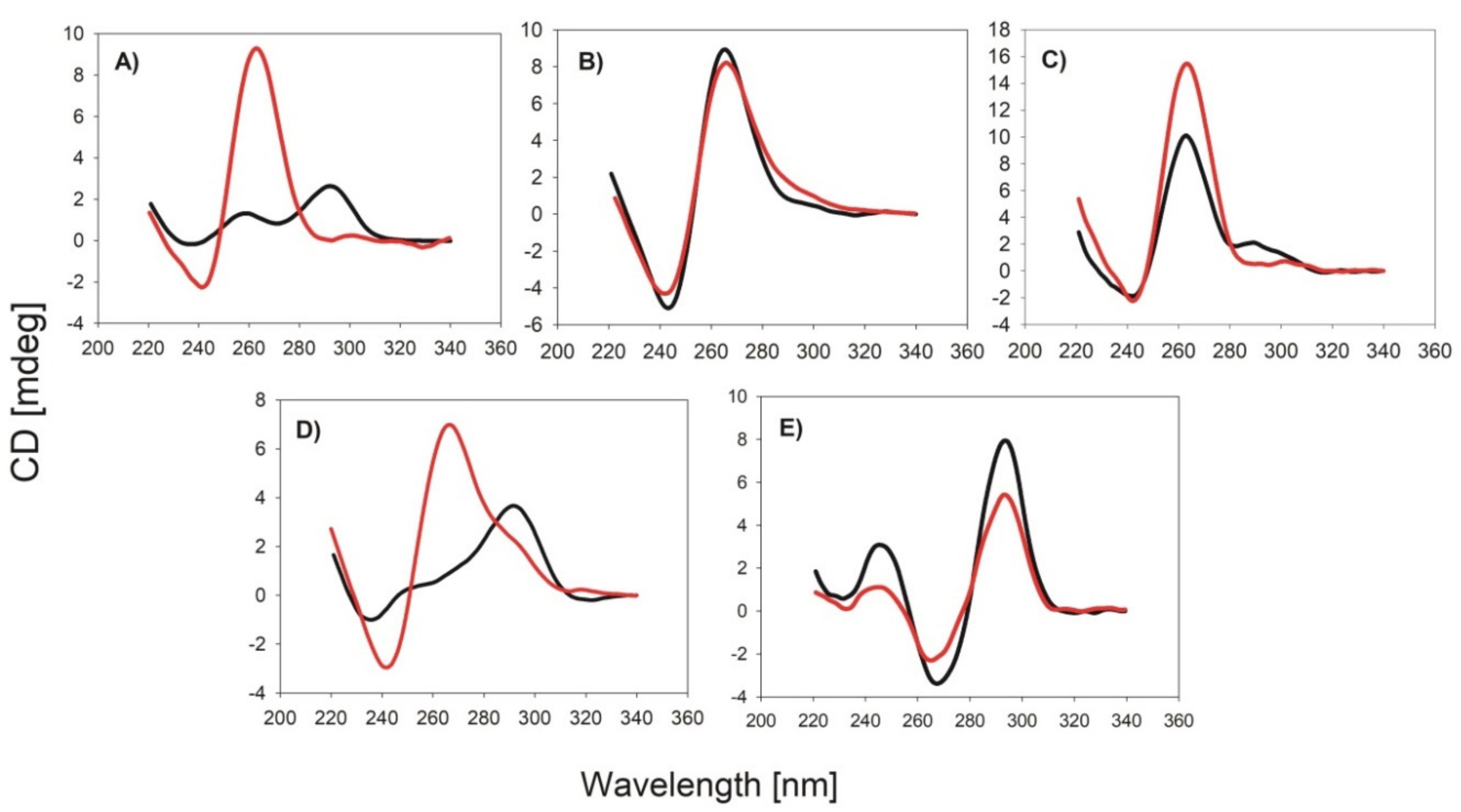
2.2. Circular Dichrosim (CD) Spectra
2.3. Melting Profiles
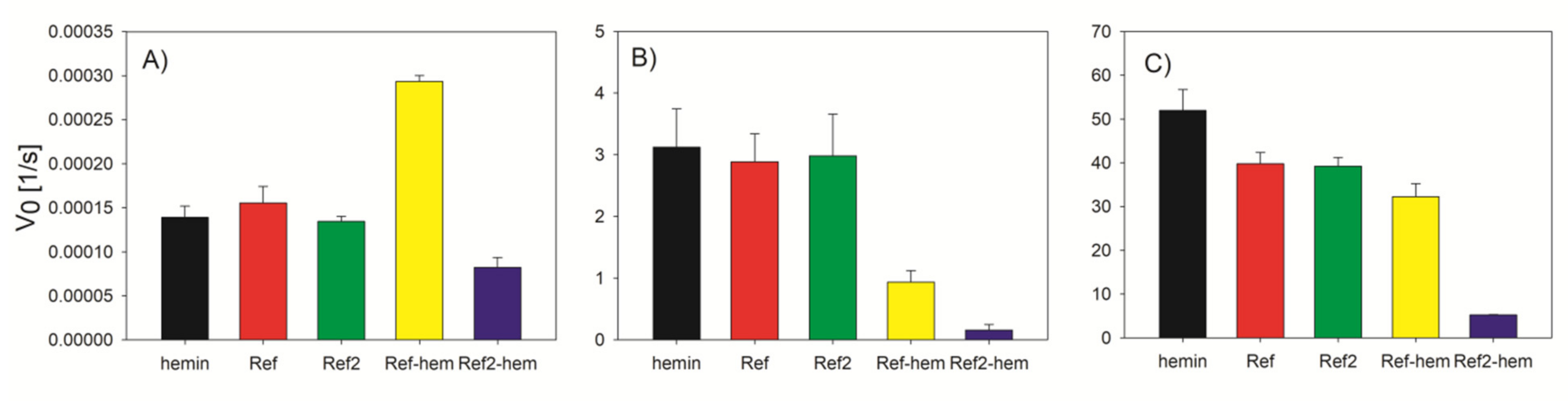
2.4. Activity Measurment
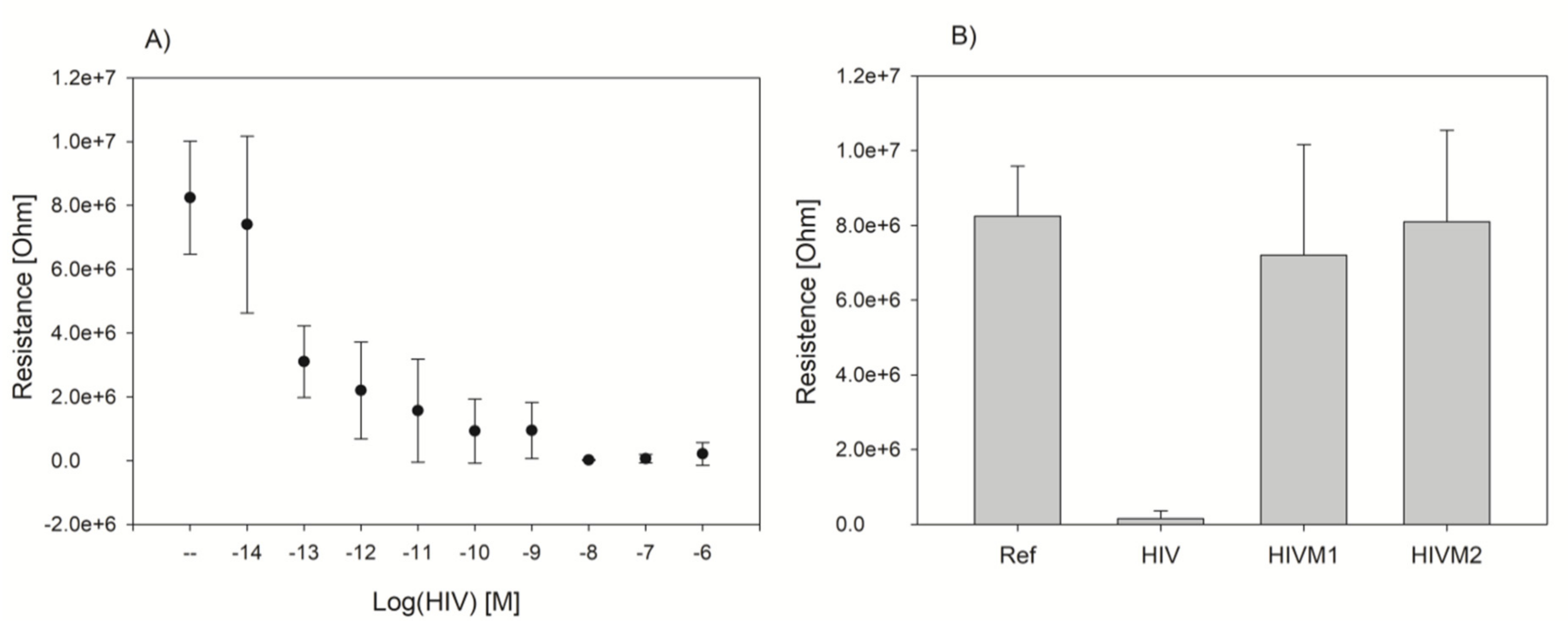
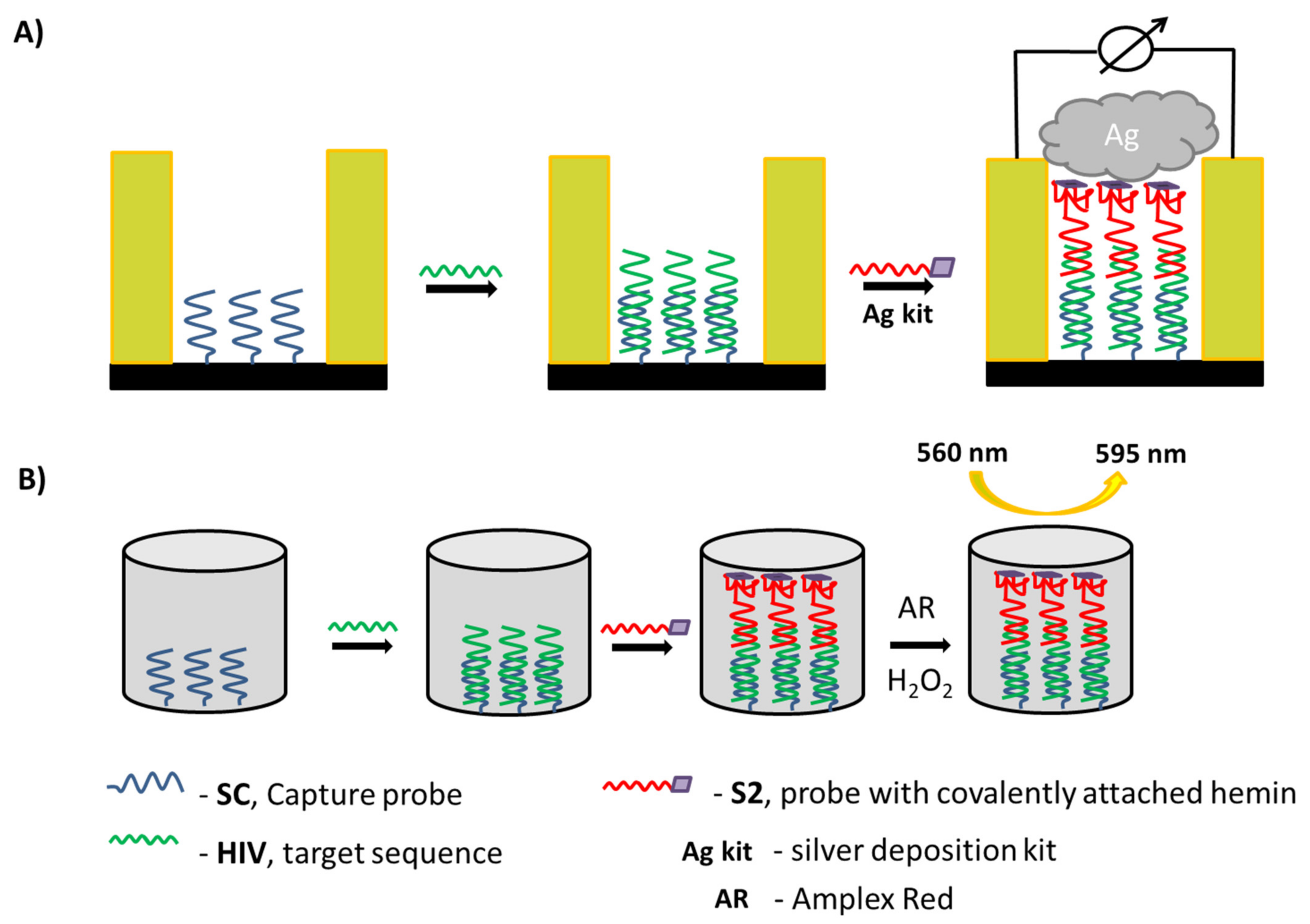
2.5. DNA Detection Procedure with Electric Chip
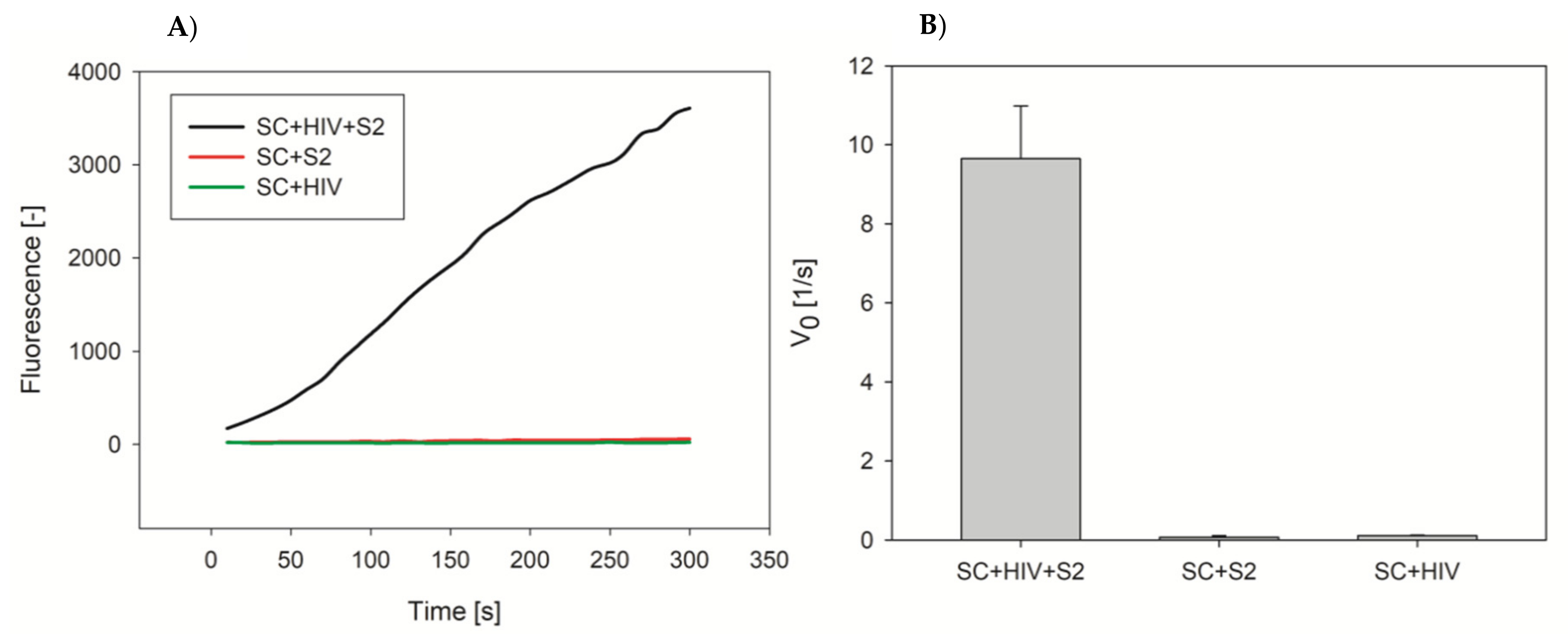
2.6. DNA Detection Procedure with Microplate Approach
3. Results
3.1. Effect of the DNA Sequence on the DNAzyme Properties
3.2. G4-Hemin Conjugates for Bioassay Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zimmermann, A.C.; White, I.M.; Kahn, J.D. Nucleic acid-cleaving catalytic DNA for sensing and therapeutics. Talanta 2020, 211, 120709. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Burciu, B.; Filipe, C.D.M.; Li, Y.; Dellinger, K.; Didar, T.F. DNAzyme-Based Biosensors: Immobilization Strategies, Applications, and Future Prospective. ACS Nano 2021, 15, 13943–13969. [Google Scholar] [CrossRef] [PubMed]
- Silverman, S.K. In vitro selection, characterization, and application of deoxyribozymes that cleave RNA. Nucleic Acids Res. 2005, 33, 6151–6163. [Google Scholar] [CrossRef] [Green Version]
- Camden, A.J.; Walsh, S.M.; Suk, S.H.; Silverman, S.K. DNA Oligonucleotide 3′-Phosphorylation by a DNA Enzyme. Biochemistry 2016, 55, 2671–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Zhou, Y.; Liu, J. Porphyrin metalation catalyzed by DNAzymes and nanozymes. Inorg. Chem. Front. 2021, 8, 2183–2199. [Google Scholar] [CrossRef]
- Kosman, J.; Juskowiak, B. Bioanalytical Application of Peroxidase-Mimicking DNAzymes: Status and Challenges. Adv. Biochem. Eng. Biotechnol. 2017, 170, 59–84. [Google Scholar] [CrossRef]
- Travascio, P.; Li, Y.; Sen, D. DNA-enhanced peroxidase activity of a DNA aptamer-hemin complex. Chem. Biol. 1998, 5, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Burge, S.; Parkinson, G.N.; Hazel, P.; Todd, A.K.; Neidle, S. Quadruplex DNA: Sequence, topology and structure. Nucleic Acids Res. 2006, 34, 5402–5415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardinelli, G.; Oloketuyi, S.; Werner, S.F.; Mazzega, E.; Högberg, B.; de Marco, A. A compact nanobody-DNAzyme conjugate enables antigen detection and signal amplification. New Biotechnol. 2020, 56, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fruk, L.; Niemeyer, C.M. Covalent Hemin–DNA Adducts for Generating a Novel Class of Artificial Heme Enzymes. Angew. Chem. Int. Ed. 2005, 44, 2603–2606. [Google Scholar] [CrossRef] [PubMed]
- Fruk, L.; Kuhlmann, J.; Niemeyer, C.M. Analysis of heme-reconstitution of apoenzymes by means of surface plasmon resonance. Chem. Commun. 2009, 230–232. [Google Scholar] [CrossRef]
- Fruk, L.; Kuo, C.-H.; Torres, E.; Niemeyer, C.M. Apoenzyme Reconstitution as a Chemical Tool for Structural Enzymology and Biotechnology. Angew. Chem. Int. Ed. 2009, 48, 1550–1574. [Google Scholar] [CrossRef] [PubMed]
- Thirstrup, D.; Baird, G.S. Histochemical Application Of A Peroxidase DNAzyme with a Covalently Attached Hemin Cofactor. Anal. Chem. 2010, 82, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, S.; Wang, J.; Sintim, H.O. DNA-Based Peroxidation Catalyst-What Is the Exact Role of Topology on Catalysis and Is There a Special Binding Site for Catalysis? Chem. Eur. J. 2011, 17, 5691–5698. [Google Scholar] [CrossRef] [PubMed]
- Fapyane, D.; Kékedy-Nagy, L.; Sakharov, I.Y.; Ferapontova, E.E. Electrochemistry and electrocatalysis of covalent hemin-G4 complexes on gold. J. Electroanal. Chem. 2018, 812, 174–179. [Google Scholar] [CrossRef]
- Kosman, J.; Stanislawska, A.; Gluszynska, A.; Juskowiak, B. Conjugation of hemin to G-quadruplex forming oligonucleotide using click chemistry. Int. J. Biol. Macromol. 2017, 101, 799–804. [Google Scholar] [CrossRef]
- Kosman, J.; Żukowski, K.; Juskowiak, B. Comparison of Characteristics and DNAzyme Activity of G4–Hemin Conjugates Obtained via Two Hemin Attachment Methods. Molecules 2018, 23, 1400. [Google Scholar] [CrossRef] [Green Version]
- Fasman, G.D. Handbook of Biochemistry and Molecular Biology, 3rd ed.; Volume 1: Nucleic Acids; CRC Press: Boca Raton, FL, USA, 1975; ISBN 978-1-315-89328-0. [Google Scholar]
- Kong, D.-M.; Cai, L.-L.; Guo, J.-H.; Wu, J.; Shen, H.-X. Characterization of the G-quadruplex structure of a catalytic DNA with peroxidase activity. Biopolymers 2009, 91, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Girvan, A.C.; Teng, Y.; Casson, L.K.; Thomas, S.D.; Jüliger, S.; Ball, M.W.; Klein, J.B.; Pierce, W.M.; Barve, S.S.; Bates, P.J. AGRO100 inhibits activation of nuclear factor-κB (NF-κB) by forming a complex with NF-κB essential modulator (NEMO) and nucleolin. Mol. Cancer Ther. 2006, 5, 1790–1799. [Google Scholar] [CrossRef] [Green Version]
- Bock, L.C.; Griffin, L.C.; Latham, J.A.; Vermaas, E.H.; Toole, J.J. Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature 1992, 355, 564–566. [Google Scholar] [CrossRef]
- Wang, Y.; Patel, D.J. Solution structure of the human telomeric repeat d[AG3(T2AG3)3] G-tetraplex. Structure 1993, 1, 263–282. [Google Scholar] [CrossRef]
- Möller, R.; Powell, R.D.; Hainfeld, J.F.; Fritzsche, W. Enzymatic Control of Metal Deposition as Key Step for a Low-Background Electrical Detection for DNA Chips. Nano Lett. 2005, 5, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Dong, S.; Wang, E. G-Quadruplex Aptamers with Peroxidase-Like DNAzyme Functions: Which Is the Best and How Does it Work? Chem. Asian J. 2009, 4, 918–922. [Google Scholar] [CrossRef]
- Kosman, J.; Juskowiak, B. Hemin/G-quadruplex structure and activity alteration induced by magnesium cations. Int. J. Biol. Macromol. 2016, 85, 555–564. [Google Scholar] [CrossRef]
- Deore, P.S.; Manderville, R.A. Ratiometric fluorescent sensing of the parallel G-quadruplex produced by PS2.M: Implications for K+ detection. Analyst 2020, 145, 1288–1293. [Google Scholar] [CrossRef]
- Kong, D.-M. Factors influencing the performance of G-quadruplex DNAzyme-based sensors. Methods 2013, 64, 199–204. [Google Scholar] [CrossRef]
- Travascio, P.; Witting, P.K.; Mauk, A.G.; Sen, D. The Peroxidase Activity of a Hemin−DNA Oligonucleotide Complex: Free Radical Damage to Specific Guanine Bases of the DNA. J. Am. Chem. Soc. 2001, 123, 1337–1348. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Chen, H.; Zhou, P.; Tao, H.; Wu, Y. Oligonucleotides and pesticide regulated peroxidase catalytic activity of hemin for colorimetric detection of isocarbophos in vegetables by naked eyes. Anal. Bioanal. Chem. 2019, 411, 7857–7868. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Zhan, S.; Liu, L.; Luo, Y.; Zhou, P. Regulation of hemin peroxidase catalytic activity by arsenic-binding aptamers for the colorimetric detection of arsenic(iii). RSC Adv. 2013, 3, 25614. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Function |
|---|---|---|
| PS2.M | 5′ GTG GGT AGG GCG GGT TGG 3′ | Hemin aptamer [7] |
| PS2.M-hemin | hemin-5′ GTG GGT AGG GCG GGT TGG 3′ | DNAzyme |
| CatG4 | 5′ TGG GTA GGG CGG GTT GGG AAA 3′ | Hemin aptamer [19] |
| CatG4-hemin | hemin-5′ TGG GTA GGG CGG GTT GGG AAA 3′ | DNAzyme |
| AGRO100 | 5′ GGT GG TGG TGG TTG TGG TGGTGGTGG 3′ | Nucleolin aptamer [20] |
| AGRO100-hemin | hemin-5′ GGT GG TGG TGG TTG TGG TGG TGG TGG 3′ | DNAzyme |
| TBA | 5′ GGT TGG TGT GGT TGG 3′ | Thrombin aptamer [21] |
| TBA-hemin | hemin-5′ GGT TGG TGT GGT TGG 3′ | DNAzyme |
| HT22 | 5′ A GGG TTA GGG TTA GGG TTA GGG 3′ | Human telomeric sequence [22] |
| HT22-hemin | hemin-5′ A GGG TTA GGG TTA GGG TTA GGG 3′ | DNAzyme |
| Ref | 5′ GAG TAG TTC GTG GCC TAG 3′ | Reference sequence |
| Ref-hemin | hemin-5′ GAG TAG TTC GTG GCC TAG 3′ | Reference conjugate |
| Ref2 | 5′ TTT ATT TAT TTA TTT ATT TAT T 3′ | Reference sequence |
| Ref2-hemin | hemin-5′ TTT ATT TAT TTA TTT ATT TAT T 3′ | Reference conjugate |
| SC | 5′-GTG TGG AAA ATC TCT AGC AGT TTT-3′-NH2/biotin | Capture probe |
| S2 | hemin-5′-AGG GTT AGG GTT AGG GTT AGG GTC CCT CAG ACC CTT TTA GTC-3′ | Detection probe |
| HIV | 5′-ACT GCT AGA GAT TTT CCA CAC TGA CTA AAG GGT CTG AGG GA-3′ | Analyte, sequence of virus HIV |
| HIVM1 | 5′-ACT GCT AGA GAT TCT CCA CAC TGA CTA AAG GGT CTG AGG GA-3′ | Analyte with 1 mismatch |
| HIVM2 | 5′-ACT GCT GGA GAT TCT CCA CAC TGA CTA AAG GGT CT GAG GGA-3′ | Analyte with 2 mismatches |
| G4 | G4/Hemin | G4-Hemin | ||||
|---|---|---|---|---|---|---|
| Na+ | K+ | Na+ | K+ | Na+ | K+ | |
| PS2.M | 46.0 | 56.5 | 45.2 | 64.6 | 58.4 | >80 |
| CatG4 | 41.8 | 79.2 | 42.2 | >80 | 53.0 | >80 |
| AGRO100 | 35.7 | 64.9 | 36.9 | 65.7 | 38.6 | >80 |
| HT22 | 53.9 | 62.7 | 52.9 | 63.4 | 80.0 | >80 |
| TBA | 23.3 | 50.7 | 21.5 | 49.0 | 57.8 | >80 |
| Name | pKa | |
|---|---|---|
| G4/Hemin | G4-Hemin | |
| PS2.M | >10 1 | 8.71 |
| CatG4 | >10 1 | 8.78 |
| AGRO100 | 9.6 | 9.73 |
| HT22 | 7.91 | >10 1 |
| TBA | 8.9 | >10 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosman, J.; Żukowski, K.; Csáki, A.; Fritzsche, W.; Juskowiak, B. Sequence Effect on the Activity of DNAzyme with Covalently Attached Hemin and Their Potential Bioanalytical Application. Sensors 2022, 22, 500. https://doi.org/10.3390/s22020500
Kosman J, Żukowski K, Csáki A, Fritzsche W, Juskowiak B. Sequence Effect on the Activity of DNAzyme with Covalently Attached Hemin and Their Potential Bioanalytical Application. Sensors. 2022; 22(2):500. https://doi.org/10.3390/s22020500
Chicago/Turabian StyleKosman, Joanna, Krzysztof Żukowski, Andrea Csáki, Wolfgang Fritzsche, and Bernard Juskowiak. 2022. "Sequence Effect on the Activity of DNAzyme with Covalently Attached Hemin and Their Potential Bioanalytical Application" Sensors 22, no. 2: 500. https://doi.org/10.3390/s22020500
APA StyleKosman, J., Żukowski, K., Csáki, A., Fritzsche, W., & Juskowiak, B. (2022). Sequence Effect on the Activity of DNAzyme with Covalently Attached Hemin and Their Potential Bioanalytical Application. Sensors, 22(2), 500. https://doi.org/10.3390/s22020500