
Insect-Inspired Robots: Bridging Biological and Artificial Systems

 ,
,  ,
,  , , , ,
, , , ,  and
and 
Abstract
1. Introduction
- Section 2.1. Robotic leg design;
- Section 2.2. From legs to robots;
- Section 2.3. Hexapod robots’ accessibility criteria for academics.
- Section 3.1. Insect locomotion control;
- Section 3.2. Robot locomotion control.
- Section 4.1. The fly brain and cognition;
- Section 4.2. The insect brain structure;
- Section 4.3. Insect brain functional models, implementations and robotic experiments.
2. Biomechanics in Hexapod Robotics
- How can we improve the level of performance of hexapod robots in term of traveled distance?
- Is the study of insects helpful in the design of highly efficient hexapod mobility?
- Are robotic legs as intelligent as insect legs over complex terrain?
2.1. Robotic Leg Design
2.1.1. Design and Morphology
- Number of degrees of freedom (DOF): determines the operating space of the robot. By increasing of DOFs, the robot can achieve more complex trajectories. The number of DOFs also has a direct impact on robot characteristics, such as its autonomy, mass, and cost. Therefore, the number of DOFs should not be neglected in the design process. Currently, insect-inspired robotic legs are designed with between one and five DOFs per leg.With one DOF per leg, the robot’s maneuverability is highly limited. Depending on both leg and control designs, a one DOF per leg robot can perform a straight line walk [55] and also achieve simple rough terrain navigation if is equipped with whegs [46,56,57], comprising elements of both wheels and legs. These whegs equipped robots cannot really be considered as insect-like robots regarding their body structure. Their development tends to target navigation tasks over locomotion studies.With two DOFs per leg, a simplified hexapod robot can be built [58]. This choice is a good compromise between energetic cost and mobility. However, this type of robot walks mainly over flat terrains and can only perform curved leg trajectories, generating body oscillations.Previously, insect-based hexapod robots were built with three DOFs per leg (Table 1, in which the trochanter segment joint is merged with the femur and each joint only comprises one DOF, see Figure 2). Reflecting the standard insect leg model [59], this type of leg permits roaming in a slightly rough or slopped terrain in addition to a flat terrain walk.More DOFs in leg morphology improve maneuverability and adaptation to challenging terrains [39,60]. Additional actuators help to adjust robot orientation according to the slope in order to increase stability [26]. Experiments have shown that 4 or 5 joints per leg enable robots to cope with high gradient slopes in any orientation (e.g., up to slopes, see [26,61], or up to slopes, see [14]). Unfortunately, these improvements increase the level of complexity of control commands and the robot’s price and weight (Table 2), they also concomitantly, reduce autonomy due to the high power consumption of numerous actuators.To sum up, from the large number of robots based on three-DOF legs, this appears to be the right compromise to walk on a flat terrain. Despite the three DOFs per leg trend, from a biological point of view, an insect possesses more than three joints with one DOF per joint [62,63], allowing it to overcome large obstacles and cross sloped and rough terrain (e.g., up to see [64]). More complex models based more closely on insect leg kinematics are being developed [65]. Dung beetle like legs were built in 2018, the leg design was based on micro-CT scans of a real dung beetle [66]. A pair of beetle-like legs comprising four DOFs, allowing both manipulation and transportation was tested [66].In 2017, a hexapod robot, called Cassino Hexapod III (∼3 kg), composed of hybrid legs on a modular anthropomorphic architecture with omni-wheels, as feet at the extremities, was designed and built [67]. Each hybrid leg was built with three DOFs with the third being dedicated to rotating the wheel at the tip of the leg. This kind of hybrid locomotion is relevant for efficient rolling mobility on moderate terrain and walking mobility on extreme terrain, such as non-terrestrial exploration [68]. Hybrid locomotion by walking or by rolling allows hexapod robots to save energy, and this hybrid locomotion is a combination of an engineering approach and a bio-inspired approach. Hybrid locomotion has not been developed in this review, which is focused on the biomimetic approach.
- Structure type: this describes how leg joints are linked to each other. Two major leg designs are used on inspect-inspired robots: serial multi-shaped legs or single shape legs.Serial multi-shaped legs are the most common structures encountered for locomotion and navigation. By definition, an insect leg is composed of five segments (coxa, trochanter, femur, tibia, and tarsus), arranged in a particular toggled zig-zag shape, forming a sprawled posture, reducing and distributing the forces on every joint of the leg [69,70]. However, in most robotic cases, this structure is simplified to three segments per leg (coxa, femur, tibia) comprising three joints per leg, each one with only one DOF (see previous point). In this arrangement, the trochanter segment of the leg is merged with the femur, and the tarsus is generally removed. However, the tarsus makes an important contribution to the insect’s walk, serving as an adhesive pad [62,71] and allowing a better ground forces transmission with a passive spring effect. Moreover, some insects (e.g., leafhoppers) possess particular tarsal structures allowing them to jump from smooth surfaces [72]. Looking over the last decade of hexapod robots, presented in Table 1, the tarsus is often neglected, even though it represents more than of the leg length [73]. Currently, few artificial tarsus designs have been developed to efficiently walk on complex terrains [74].Single shape legs are used on whegs robots, origami, and compliant joint robots. The specifications of these types of robots, require the absence of most tiny mechanical parts such as bearings, shafts, screws, and nuts, and involves a simplicity of manufacturing, scale and cost reduction and, backlash and structural robustness improvements. The development of single shape legs follows the advances in new materials and manufacturing techniques such as multi-material 3D printing, which allows the building of soft joint robots [75]. Particularly, the 3D printing of legs appears to be a good way to develop and simplify standard joint designs by using properties of these new materials, such as flexibility or heat deformation [76]. In this way, hexapod robot legs are tending to become closer to real insect legs, in terms of relative dimensions and mass. An important point to notice for insects, e.g., cockroaches, is that a leg corresponds to approximately of the body mass [77], allowing them low inertia, high frequency strides during a walk. In comparison, insect-like robot legs represent at least of the overall mass (estimated for a 2 kg robot, from Table 1). Apart from 3D printing, some materials could take over from standard aluminum or molded plastic legs, e.g., chitosan–fibroin material, inspired by insect cuticle structure [78].Furthermore, some other original structures have been designed; they were mainly developed when a specific animal behavior, such as jumping [79,80], is to be replicated or to satisfy some sought after design specifications like posture changes [81].At first glance, leg design is highly dependent on actuator technologies. However, an impressive number of improvements are still possible through subtle structural modifications, allowing huge performance improvements. Independently of the structure type, observing the current state of the art in leg design, a question presents itself: why are all the legs of a hexapod robots the same? Insect legs are different in size (Figure 3, see [82]), and not built like robot legs, wherein the six are often identical, except for a few robots mimicking insect morphology in detail (Drosophibot [18,19] and MantisBot [30,31]). In response to this question and with the technologies now available, in the 2020s, leg design is likely to become increasingly based on available micro-CT scans of real insects (e.g., [66,83] dung beetles, [84] flies, or [65] ants) in order to improve the level of complexity, fidelity, and bio-inspiration.
2.1.2. Cost of Transport
2.1.3. Actuation of the Legs
- Servomotors: the main issues with servomotors are their weight and energy efficiency. A servomotor heats up easily until it surpasses its maximum operating temperature of 70 °C, then it stops working. In addition, a servomotor is composed by definition, of a motor with high ratio gearing used to make it as stiff as possible. In this sense, such servo-based actuators differ significantly from biological actuators that may have variable stiffness and adaptable compliance. One way to implement variable stiffness is to use springs to make variable impedance actuators (VIA). As summarized in [91], VIA actuators can be classified into three categories: spring pre-loaded variation, transmission ratio changing and spring physical property alteration. VIA is certainly an approach of great interest for the design of future hexapod robots able to dynamically change the stiffness of their joints.
- Brushless motors: recent developments in smart rotating actuators based on brushless motors will permit the design of direct drive joints without gearing. The maximum specific power of electric motors with permanent magnets is 300 W/kg, which is about the same order as biological muscle [92]. Companies, such as HEBI Robotics [17] or IQ Motion, have developed integrated rotating actuators for robotic applications and for the development of mobile robots of various sizes. As the electronic driver and angular sensor are integrated into the motor, it drastically simplifies the wiring and complexity of the overall hardware architecture, which can be crucial when designing robots like hexapods that require the control of 18 actuators.
- Artificial muscles: the design of future insect-inspired robots will certainly depend on the availability of actuators able to mimic the functionalities of biological muscles. Their properties of viscoelasticity and energy dissipation leading to high compliance is the holy grail of insect-inspired actuators. Among the broad repertoire of new artificial actuators for robots (see review by [93]), non-conventional actuators like pneumatic artificial muscles (PAMs), shape memory alloys (SMAs), and electroactive polymers (EAPs) are of great interest. One particular case is HASEL actuators, which are composed of a series of pouches made of a flexible and inextensible shell that is filled with a liquid dielectric. Electrodes cover a portion of each pouch so as to progressively close when a voltage is applied thus squeezing the pouches to increase their volume [94]. HASEL actuators can be implemented in different ways and can feature a bandwidth as high as 126 Hz for the quadrant-donut HASEL and even a specific energy twice as high as mammalian skeletal muscles for the planar HASEL actuator [94]. HASEL actuators mimic the muscle-like performance of dielectric actuators (DEAs), which can be highly effective for robotic applications. They can lift more than 200 times their weight and have a peak specific power of 585 W/kg [95]. Moreover, it is worth noting that a toolkit has been developed to aid designs using HASEL actuators [96]. In addition, electro-ribbon technology, with an ability of lifting 1000 times its own weight and a contraction by 99.8% of its length, is also very promising [97]. Finally, five-DOF soft dielectric elastomer actuators have been shown to be very useful in the implementation of soft legged robots [98] which are able to walk with an alternative tripod gait as fast as 52 mm/s for a 7 Hz actuation frequency.
2.1.4. Force Sensing in Robotic Legs
- Leg tip/TARSE sensing: sensing at the end of the leg can be done by a tactile sensor, a pressure sensor, a three-axis force/torque sensor [100], or a compliant force sensor made with a spring [15,39]. Leg tip sensing can be easily implemented by adding an attachment point to the leg tip without requiring any modification of the robot’s structure. The cost of these tip sensors can be expensive depending on the chosen technology, but recent research has developed low-cost designs [100]. Tip force sensing is useful because it provides the robot with a terrain description. Force measurements allow the robot to understand which of its legs are in contact with the ground, or to evaluate the terrain slope, in order to both adjust its gait and plan its path [101].
- Force sensing in actuators: sensing coming from the state of actuators by current measurement [26,29,37,43] or dedicated sensors in the joints. This category of sensors simplifies the robot’s design, since the sensor is integrated within the actuator forming a compact structure. The complexity of the estimation of forces from the currents generated by the actuators is based on the robot’s leg model. To obtain an accurate leg movement, the mathematical model should reflect the robot as closely as possible, and take account of any structural deformation under various loads since no material is perfectly rigid.
- Legs with compliant structures: compliant mechanisms exploit the deformation properties of the leg segments, deformations that could be a disadvantage in other legs. Stiffer legs appear to narrow the region of stable gaits while preventing tripod contact with the ground. However, compliant legs are more capable of absorbing energy even if the leg touches down early, thus minimizing the severity of ground reaction on legs. This solution had been developed for one-joint C-shaped legs [44,45,54]. Whegs do not possess any force sensors on their legs. Compliant legs offer the possibility of placing the force sensors along the segments (such as the femur or the tibia) [101,102]. This type of sensor placement mimics the force measurements in insects, as done by campaniform sensilla mechanoreceptors [103,104].
2.2. From Legs to Robots
2.2.1. Body Morphology
2.2.2. Scale Effect on Level of Performance
2.3. Hexapod Robot Accessibility Criteria for Academics
3. From Biomechanics to Locomotion
3.1. Insect Locomotion Control
- Interlimb coordination: biological studies have revealed rules for interlimb coordination of insect locomotion. For instance, Wilson [119] proposed five rules. Rule 1: a wave of swing runs from hind (posterior) to front (anterior) legs. Rule 2: contralateral legs of the same segment alternate in phase. Rule 3: protraction (swing) time is constant. Rule 4: frequency varies (stance decreases as frequency increases). Rule 5: the intervals between steps of the hind leg and middle leg and between the middle leg and fore leg are constant, while the interval between the foreleg and hind leg steps varies inversely with frequency. These rules have been translated to neural mechanisms for hexapod locomotion control, which can generate various insect-like gaits [141,142].Subsequent research by Cruse et al. [116] introduced six rules for insect walking (called WalkNet). The rules were derived from behavioral experiments with stick insects. Rule 1: posterior swing inhibits start of anterior swing. Rule 2: start of posterior stance excites anterior swing (posterior reaches a given anterior extreme position (AEP)). The AEP is the anterior transition point from swing to stance in a forward walking animal. Rule 3: caudal positions of anterior stance excite start of posterior swing (anterior reaches a given posterior extreme position (PEP)). The PEP is the posterior transition point from stance to swing. Rule 4: end position of anterior stance influences end position of posterior swing (called targeting). Rule 5: increased resistance increases force and increased load prolongs stance phase. Rule 6: the information from the anterior leg’s reflex stimulation is passed on to the posterior leg. Recently, Schilling and Cruse [143] introduced the realization of these rules as an artificial neuronal network with an antagonistic structure (called neuroWalknet controller). The controller can generate diverse robot walking behaviors including different gait patterns emerging from different velocities, curve negotiation, and backward walking.In addition to the aforementioned rules for insect locomotion, a recent study from Leung et al. [144] analyzed and identified four underlying rules for interlimb coordination of dung beetle ball rolling gaits. Rule 1: front legs alternately step on the ground. The rule describes the relationship between the two front legs in the gait. Rule 2: each middle leg steps similarly to its contralateral hind leg. The rule describes the synchronization of the contralateral middle and hind legs. Rule 3: an ipsilateral pair of middle and hind legs seldom lift together. Rule 4: a contralateral pair of middle or hind legs rarely lift together. In principle, a pair of legs following the third and fourth rules tend not to lift together. A partial implementation of the rules as modular neural control with a CPG was performed and tested on a simulated dung beetle-like robot [4]. The controller can generate four different robot behaviors including forward walking, backward walking, level-ground ball rolling, and sloped-ground ball rolling.
- Intralimb coordination: In addition to the biological studies of the relationship between legs (interlimb coordination) in insects, some studies have further investigated individual leg movements and adaptations during normal and rough terrain walking in insects. The leg movements basically reflect intralimb coordination. For instance, Pearson and Franklin [117] proposed locusts’ reflex strategies for leg movements when walking over rough and complex terrain. As described by them, the strategies include (1) rhythmic searching movements; (2) local searching movements; and (3) elevator reflex.Based on [117], the rhythmic searching movements are to search for a ground contact, if the animal has not located it by the end of its swing phase. The searching movements show rhythmic patterns including fast elevation and depression movements of the leg. While searching, the animal also extends the search range from the body to explore the supporting points around the leg, e.g., up to eight searching cycles. The searching typically stops either when the animal stops walking, the leg gets stuck, or ground contact is found. The local searching movements are small rhythmic leg movements from point to point on a potential supporting ground. These movements occur either at the beginning of a stance phase, after the rhythmic searching movements and/or an elevator reflex (described below). The local searching movements are required if the potential support surface is smooth where the leg action needs to push the animal forward. The elevator reflex is a rapid elevation and extension of the leg to step over an obstacle, followed by placing the leg where the obstacle can be used as a support. The elevator reflex can be activated when the leg gets stuck during the swing phase. It can also occur during searching movements. In rough terrain walking experiments on locusts, the elevator reflex was mostly observed in the fore and middle legs while the hind legs moved behind the animal. This makes it difficult to distinguish between an elevator reflex and a passive pulling of the hind legs up onto the obstacle while moving forward. Examples of the implementation of these reflex strategies for adaptive hexapod robot locomotion on rough and complex terrain can be seen at [118,142].
- Joint (mechanical) compliance: this is the interactive relationship between kinematic changes and the resulting dynamics of joints [145]. One of its key components, stiffness, refers to the ratio between joint torque and angle changes, which are related to muscle activation. Compliance control of insect muscles is very important in facilitating adaptive and robust locomotion over natural terrain [146,147]. Computerized (computational) muscle models can be used to enhance the understanding of neuromechanical control principles underlying insect locomotion [148]. The Hill muscle is one of the most influential ‘seed’ models that has inspired many successors [149]. For instance, Proctor and Holmes built a neuromechanical model to study feedback effects of perturbated insect locomotion [150], in which 24 neural oscillators and 48 pairs of Hill muscles were used. Guo et al. proposed a neuro–musculo–skeletal model to reproduce gait pattern in virtual insects [151]. Naris et al. analyzed a closed-loop neuromechanical simulation of insect joint control driven by a pair of Hill muscles [152]. However, most of these studies were limited to numerical simulations, because a greater number of parameters needed to be offline optimized based on nonlinear differential equations. Therefore, they failed to account for intrinsically delayed feedback in real insect locomotion dynamics. This failure may cause a misinterpretation of the neuromechanical control principles in insect locomotion. Therefore, questions remain open whose answers may decode insect muscle intelligence in dynamic robust locomotion.
- -
- Multifunctional muscles: insects exhibit different muscle functions in flying and walking [153], which are characterized by the work loop technique [154]. These functions may facilitate the decoding of muscle compliance in dynamic insect locomotion. Interestingly, some preliminary results show that muscles act as brakes and springs when their passive stiffness and damping are tuned in computational simulations [155]. Tuning muscle stiffness and damping properties based on the work-loop technique, can be a key to understanding and translating muscle intelligence between engineering applications and neuromechanical models [115,156].
- -
- Predictive muscle tuning: muscle compliance can be tuned in terms of sensory feedback. However, this feedback is intrinsically subject to noise and delays owing to high levels of dynamics of insect locomotion. Therefore, it may be assumed that insects, and their robot counterparts use internal models to predict sensory outputs for tuning insect muscle compliance [114].
- -
- Engineering-inspired muscle intelligence: biological muscle control principles have been borrowed to enhance robot designs and control for many years. This research approach can be flipped, i.e., robots as tools for decoding muscle compliance in insect locomotion [157]. For instance, an insect-like robot was used to test a simplified muscle control hypothesis, i.e., proximodistal gradient [114]. It showed that this gradient reduces the number of controlled variables and enhances walking stability. Engineering-inspired methods can close the research loop of insect muscle intelligence, providing new hypotheses for biological experiments on insect locomotion.
3.2. Robot Locomotion Control
3.2.1. Bio-Inspired Control
3.2.2. Engineering-Based Control
3.2.3. Machine Learning-Based Control
4. From Locomotion to Cognition
4.1. The Fly Brain and Cognition
4.2. The Insect Brain Structure
4.3. Insect Brain Functional Models, Implementations and Robotic Experiments
4.3.1. MB Models
4.3.2. CX Models
4.3.3. Models Involving MB-CX Interaction
5. Lessons Learned from This Review
5.1. Future Directions in Biomechanics
5.2. Future Directions in Locomotion Control
5.3. Future Directions in Insect-Inspired Robotic Cognition
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AEP | anterior extreme position |
| AFO | adaptive frequency oscillator |
| AFDC | adaptation through fast dynamical coupling |
| AHM | artificial hormone mechanism |
| CoT | cost of transport |
| CPG | central pattern generator |
| CX | central complex |
| DOF | degree of freedom |
| EB | ellipsoid body |
| FB | fan-shaped body |
| GRF | ground reaction force |
| KCs | Kenyon cells |
| IK | inverse kinematics |
| MBs | mushroom bodies |
| PB | protocerebral bridge |
| PEP | posterior extreme position |
| PM | phase modulation |
| PR | phase resetting |
| STDP | spike-timing-dependent plasticity |
References
- Ijspeert, A.J. Biorobotics: Using robots to emulate and investigate agile locomotion. Science 2014, 346, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, J.; Zhang, T.; Qian, F.; Kingsbury, M.; McInroe, B.; Mazouchova, N.; Li, C.; Maladen, R.; Gong, C.; Travers, M. A review on locomotion robophysics: The study of movement at the intersection of robotics, soft matter and dynamical systems. Rep. Prog. Phys. 2016, 79, 110001. [Google Scholar] [CrossRef] [PubMed]
- Dupeyroux, J.; Serres, J.R.; Viollet, S. AntBot: A six-legged walking robot able to home like desert ants in outdoor environments. Sci. Robot. 2019, 4. [Google Scholar] [CrossRef]
- Leung, B.; Thor, M.; Manoonpong, P. Modular neural control for bio-inspired walking and ball rolling of a dung beetle-like robot. In ALIFE 2018: The 2018 Conference on Artificial Life; MIT Press: Cambridge, MA, USA, 2018; pp. 335–342. [Google Scholar]
- Xiong, X.; Manoonpong, P. No Need for Landmarks: An Embodied Neural Controller for Robust Insect-like Navigation Behaviors. IEEE Trans. Cybern. 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Beer, R.D.; Ritzmann, R.E.; McKenna, T.M. Biological Neural Networks in Invertebrate Neuroethology and Robotics; Academic Press: Cambridge, MA, USA, 1993. [Google Scholar]
- Mocci, U.; Petternella, M.; Salinari, S. Experiments with Six Legged Walking Machines with Fixed Gait; Technical Report, Report 2–12; Institute of Automation, University of Rome: Rome, Italy, 1972. [Google Scholar]
- Schneider, A.Y.; Gurfinkel, E.; Kanaev, E.; Ostapchuk, V. A System for Controlling the Extremities of an Artificial Walking Apparatus; Technical Report, Report No. 5, General and Molecular Physics Series; Physio-Technical Institute: Moscow, Russia, 1974. [Google Scholar]
- Gurfinkel, V.; Gurfinkel, E.; Shneider, A.; Devjanin, E.; Lensky, A.; Shtilman, L. Walking robot with supervisory control. Mech. Mach. Theory 1981, 16, 31–36. [Google Scholar] [CrossRef]
- McGhee, R.B.; Iswandhi, G.I. Adaptive locomotion of a multilegged robot over rough terrain. IEEE Trans. Syst. Man Cybern. 1979, 9, 176–182. [Google Scholar] [CrossRef]
- Kessis, J.; Penne, J.; Rambant, J. Six legged walking robot has brains in its legs. Sens. Rev. 1982, 2, 30–32. [Google Scholar] [CrossRef]
- Carbone, G.; Ceccarelli, M. Legged robotic systems. In Cutting Edge Robotics; IntechOpen: London, UK, 2005. [Google Scholar]
- Tedeschi, F.; Carbone, G. Design issues for hexapod walking robots. Robotics 2014, 3, 181–206. [Google Scholar] [CrossRef]
- Čížek, P.; Zoula, M.; Faigl, J. Design, Construction, and Rough-Terrain Locomotion Control of Novel Hexapod Walking Robot With Four Degrees of Freedom Per Leg. IEEE Access 2021, 9, 17866–17881. [Google Scholar] [CrossRef]
- Thor, M. MORF—Modular Robot Framework. Master’s Thesis, Maersk Mc-Kinney Moller Inst., Univ. Southern Denmark, Odense, Denmark, 2019. [Google Scholar]
- Thor, M.; Manoonpong, P. A fast online frequency adaptation mechanism for CPG-based robot motion control. IEEE Robot. Autom. Lett. 2019, 4, 3324–3331. [Google Scholar] [CrossRef]
- HEBI Robotics. Daisy Hexapod Robot Kit (18-DoF), Ref. A-2049-01. 2019. Available online: https://www.hebirobotics.com/; https://docs.hebi.us/resources/kits/outOfTheBox/A-2049-01_Box_Manual.pdf (accessed on 19 November 2019).
- Goldsmith, C.; Szczecinski, N.; Quinn, R. Drosophibot: A fruit fly inspired bio-robot. In Conference on Biomimetic and Biohybrid Systems; Springer International Publishing: Cham, Switzerland, 2019; pp. 146–157. [Google Scholar]
- Goldsmith, C.A.; Szczecinski, N.S.; Quinn, R.D. Neurodynamic modeling of the fruit fly Drosophila melanogaster. Bioinspir. Biomim. 2020, 15, 065003. [Google Scholar] [CrossRef]
- Dupeyroux, J.; Viollet, S.; Serres, J.R. An ant-inspired celestial compass applied to autonomous outdoor robot navigation. Robot. Auton. Syst. 2019, 117, 40–56. [Google Scholar] [CrossRef]
- Cheah, W.; Khalili, H.H.; Arvin, F.; Green, P.; Watson, S.; Lennox, B. Advanced motions for hexapods. Int. J. Adv. Robot. Syst. 2019, 16, 1729881419841537. [Google Scholar] [CrossRef]
- Zhong, B.; Zhang, S.; Xu, M.; Zhou, Y.; Fang, T.; Li, W. On a CPG-based hexapod robot: AmphiHex-II with variable stiffness legs. IEEE/ASME Trans. Mechatron. 2018, 23, 542–551. [Google Scholar] [CrossRef]
- Silva, O.A.; Sigel, P.; Eaton, W.; Osorio, C.; Valdivia, E.; Frois, N.; Vera, F. CRABOT: A Six-Legged Platform for Environmental Exploration and Object Manipulation. In Proceedings of the CRoNe2018: 4th Congress on Robotics and Neuroscience, Valparaíso, Chile, 8–10 November 2018; pp. 46–51. [Google Scholar]
- Dupeyroux, J.; Passault, G.; Ruffier, F.; Viollet, S.; Serres, J. Hexabot: A Small 3D-Printed Six-Legged Walking Robot Designed for Desert Ant-Like Navigation Tasks. In Proceedings of the 20th IFAC Word Congress 2017, Toulouse, France, 9–14 July 2017; Volume 2017, pp. 1628–1631. [Google Scholar]
- Trossen Robotics. PhantomX AX Metal Hexapod MK-III Kit, Ref. KIT-PXC-HEX-MK3-AX12. 2020. Available online: https://www.trossenrobotics.com/phantomx-ax-hexapod.aspx (accessed on 3 June 2021).
- Bjelonic, M.; Kottege, N.; Beckerle, P. Proprioceptive control of an over-actuated hexapod robot in unstructured terrain. In Proceedings of the 2016 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Daejeon, Korea, 9–14 October 2016; pp. 2042–2049. [Google Scholar]
- Halvorsen, K. MX Phoenix, Zenta Robotic Creations. 2016. Available online: http://zentasrobots.com/ (accessed on 10 June 2021).
- Barron-Zambrano, J.H.; Torres-Huitzil, C.; Girau, B. Perception-driven adaptive CPG-based locomotion for hexapod robots. Neurocomputing 2015, 170, 63–78. [Google Scholar] [CrossRef]
- Palankar, M.; Palmer, L. A force threshold-based position controller for legged locomotion. Auton. Robot. 2015, 38, 301–316. [Google Scholar] [CrossRef]
- Szczecinski, N.S.; Chrzanowski, D.M.; Cofer, D.W.; Moore, D.R.; Terrasi, A.S.; Martin, J.P.; Ritzmann, R.E.; Quinn, R.D. MantisBot: A platform for investigating mantis behavior via real-time neural control. In Conference on Biomimetic and Biohybrid Systems; Springer International Publishing: Cham, Switzerland, 2015; pp. 175–186. [Google Scholar]
- Szczecinski, N.S.; Getsy, A.P.; Martin, J.P.; Ritzmann, R.E.; Quinn, R.D. Mantisbot is a robotic model of visually guided motion in the praying mantis. Arthropod Struct. Dev. 2017, 46, 736–751. [Google Scholar] [CrossRef] [PubMed]
- Kalouche, S.; Rollinson, D.; Choset, H. Modularity for maximum mobility and manipulation: Control of a reconfigurable legged robot with series-elastic actuators. In Proceedings of the 2015 IEEE International Symposium on Safety, Security, and Rescue Robotics (SSRR), West Lafayette, IN, USA, 18–20 October 2015; pp. 1–8. [Google Scholar]
- Festo. BionicANT. 2015. Available online: https://www.festo.com/group/en/cms/10157.htm (accessed on 11 June 2021).
- Schneider, A.; Paskarbeit, J.; Schilling, M.; Schmitz, J. HECTOR, a bio-inspired and compliant hexapod robot. In Conference on Biomimetic and Biohybrid Systems; Springer International Publishing: Cham, Switzerland, 2014; pp. 427–429. [Google Scholar]
- Dürr, V.; Arena, P.P.; Cruse, H.; Dallmann, C.J.; Drimus, A.; Hoinville, T.; Krause, T.; Mátéfi-Tempfli, S.; Paskarbeit, J.; Patanè, L. Integrative biomimetics of autonomous hexapedal locomotion. Front. Neurorobot. 2019, 13, 88. [Google Scholar] [CrossRef]
- Meyer, H.G.; Klimeck, D.; Paskarbeit, J.; Rückert, U.; Egelhaaf, M.; Porrmann, M.; Schneider, A. Resource-efficient bio-inspired visual processing on the hexapod walking robot HECTOR. PLoS ONE 2020, 15, e0230620. [Google Scholar] [CrossRef]
- Belter, D.; Walas, K. A compact walking robot–flexible research and development platform. In Recent Advances in Automation, Robotics and Measuring Techniques; Springer International Publishing: Cham, Switzerland, 2014; pp. 343–352. [Google Scholar]
- Belter, D.; Skrzypczyński, P.; Walas, K.; Wlodkowic, D. Affordable multi-legged robots for research and STEM education: A case study of design and technological aspects. In Progress in Automation, Robotics and Measuring Techniques; Springer International Publishing: Cham, Switzerland, 2015; pp. 23–34. [Google Scholar]
- Rönnau, A.; Heppner, G.; Nowicki, M.; Dillmann, R. LAURON V: A versatile six-legged walking robot with advanced maneuverability. In Proceedings of the 2014 IEEE/ASME International Conference on Advanced Intelligent Mechatronics, Besançon, France, 8–11 July 2014; pp. 82–87. [Google Scholar]
- Buettner, T.; Roennau, A.; Heppner, G.; Dillmann, R. Design of an exchangeable, compact and modular bio-inspired leg for six-legged walking robots. In Proceedings of the CLAWAR 2017: 20th International Conference on Climbing and Walking Robots and the Support Technologies for Mobile Machines, Porto, Portugal, 11–13 September 2017; pp. 89–96. [Google Scholar] [CrossRef]
- Roehr, T.M.; Cordes, F.; Kirchner, F. Reconfigurable integrated multirobot exploration system (RIMRES): Heterogeneous modular reconfigurable robots for space exploration. J. Field Robot. 2014, 31, 3–34. [Google Scholar] [CrossRef]
- Von Twickel, A.; Hild, M.; Siedel, T.; Patel, V.; Pasemann, F. Neural control of a modular multi-legged walking machine: Simulation and hardware. Robot. Auton. Syst. 2012, 60, 227–241. [Google Scholar] [CrossRef]
- Asif, U.; Iqbal, J. Motion planning using an impact-based hybrid control for trajectory generation in adaptive walking. Int. J. Adv. Robot. Syst. 2011, 8, 53. [Google Scholar] [CrossRef]
- Galloway, K.C.; Clark, J.E.; Yim, M.; Koditschek, D.E. Experimental investigations into the role of passive variable compliant legs for dynamic robotic locomotion. In Proceedings of the 2011 IEEE International Conference on Robotics and Automation, Shanghai, China, 9–13 May 2011; pp. 1243–1249. [Google Scholar]
- Galloway, K.C.; Clark, J.E.; Koditschek, D.E. Variable stiffness legs for robust, efficient, and stable dynamic running. J. Mech. Robot. 2013, 5, 011009. [Google Scholar]
- Galloway, K.C.; Haynes, G.C.; Ilhan, B.D.; Johnson, A.M.; Knopf, R.; Lynch, G.A.; Plotnick, B.N.; White, M.; Koditschek, D.E. X-RHex: A Highly Mobile Hexapedal Robot for Sensorimotor Tasks; Technical Report; University of Pennsylvania, Department of Electrical & Systems Engineering: Philadelphia, PA, USA, 2010. [Google Scholar]
- Gorner, M.; Wimbock, T.; Baumann, A.; Fuchs, M.; Bahls, T.; Grebenstein, M.; Borst, C.; Butterfass, J.; Hirzinger, G. The DLR-Crawler: A testbed for actively compliant hexapod walking based on the fingers of DLR-Hand II. In Proceedings of the 2008 IEEE/RSJ International Conference on Intelligent Robots and Systems, Nice, France, 22–26 September 2008; pp. 1525–1531. [Google Scholar]
- Manoonpong, P. Neural Preprocessing and Control of Reactive Walking Machines. Master’s Thesis, University of Siegen, Siegen, Germany, 2006. [Google Scholar]
- Manoonpong, P.; Pasemann, F.; Roth, H. Modular reactive neurocontrol for biologically inspired walking machines. Int. J. Robot. Res. 2007, 26, 301–331. [Google Scholar] [CrossRef]
- Pavone, M.; Arena, P.; Patané, L. An innovative mechanical and control architecture for a biomimetic hexapod for planetary exploration. Space Technol.-Abingdon 2006, 26, 13–24. [Google Scholar]
- Arena, P.; Fortuna, L.; Frasca, M.; Patané, L.; Pavone, M. Realization of a CNN-driven cockroach-inspired robot. In Proceedings of the 2006 IEEE International Symposium on Circuits and Systems, Island of Kos, Greece, 21–24 May 2006. [Google Scholar]
- William, A.L. Insect-Inspired, Actively Compliant Robotic Hexapod. Master’s Thesis, Case Western Reserve University, Cleveland, OH, USA, 2005. [Google Scholar]
- Lewinger, W.A.; Quinn, R.D. A hexapod walks over irregular terrain using a controller adapted from an insect’s nervous system. In Proceedings of the 2010 IEEE/RSJ International Conference on Intelligent Robots and Systems, Taipei, Taiwan, 18–22 October 2010; pp. 3386–3391. [Google Scholar]
- Saranli, U.; Buehler, M.; Koditschek, D.E. RHex: A simple and highly mobile hexapod robot. Int. J. Robot. Res. 2001, 20, 616–631. [Google Scholar]
- Tavolieri, C.; Ottaviano, E.; Ceccarelli, M.; Di Rienzo, A. Analysis and design of a 1-DOF leg for walking machines. Proc. RAAD 2006, 6, 63–71. [Google Scholar]
- Li, M.; He, Z.; Zha, F. Design and analysis of a semi-circular flexible hexapod robot miniRHex. In Proceedings of the 2018 13th World Congress on Intelligent Control and Automation (WCICA), Changsha, China, 4–8 July 2018; pp. 418–423. [Google Scholar] [CrossRef]
- Haldane, D.W.; Fearing, R.S. Running beyond the bio-inspired regime. IEEE Int. Conf. Robot. Autom. 2015, 2015, 4539–4546. [Google Scholar] [CrossRef]
- Soltero, D.E.; Julian, B.J.; Onal, C.D.; Rus, D. A lightweight modular 12-DOF print-and-fold hexapod. In Proceedings of the IEEE International Conference on Intelligent Robots and Systems, Tokyo, Japan, 3–7 November 2013; pp. 1465–1471. [Google Scholar] [CrossRef]
- Schilling, M.; Hoinville, T.; Schmitz, J.; Cruse, H. Walknet, a bio-inspired controller for hexapod walking. Biol. Cybern. 2013, 107, 397–419. [Google Scholar] [CrossRef]
- Bjelonic, M.; Kottege, N.; Homberger, T.; Borges, P.; Beckerle, P.; Chli, M. Weaver: Hexapod robot for autonomous navigation on unstructured terrain. J. Field Robot. 2018, 35, 1063–1079. [Google Scholar] [CrossRef]
- Žák, M.; Rozman, J.; Zbořil, F.V. Design and Control of 7-DOF Omni-directional Hexapod Robot. Open Comput. Sci. 2021, 11, 80–89. [Google Scholar] [CrossRef]
- Delcomyn, F.; Nelson, M.E.; Cocatre-Zilgien, J.H. Sense organs of insect legs and the selection of sensors for agile walking robots. Int. J. Robot. Res. 1996, 15, 113–127. [Google Scholar] [CrossRef]
- Roennau, A.; Kerscher, T.; Dillmann, R. Design and kinematics of a biologically-inspired leg for a six-legged walking machine. In Proceedings of the 2010 3rd IEEE RAS and EMBS International Conference on Biomedical Robotics and Biomechatronics, BioRob 2010, Tokyo, Japan, 26–29 September 2010; pp. 626–631. [Google Scholar] [CrossRef]
- Wohlgemuth, S.; Ronacher, B.; Wehner, R. Ant odometry in the third dimension. Nature 2001, 411, 795–798. [Google Scholar] [CrossRef]
- Drapin, J.; Arroyave-Tobon, S.; Linares, J.M.; Fourcassié, V.; Moretto, P. 3D kinematic of the leg joints at the ant scale. Comput. Methods Biomech. Biomed. Eng. 2021, (in press).
- Ignasov, J.; Kapilavai, A.; Filonenko, K.; Larsen, J.C.; Baird, E.; Hallam, J.; Büsse, S.; Kovalev, A.; Gorb, S.N.; Duggen, L. Bio-inspired design and movement generation of dung beetle-like legs. Artif. Life Robot. 2018, 23, 555–563. [Google Scholar] [CrossRef]
- Tedeschi, F.; Carbone, G. Design of a novel leg-wheel hexapod walking robot. Robotics 2017, 6, 40. [Google Scholar] [CrossRef]
- Wilcox, B.H.; Litwin, T.; Biesiadecki, J.; Matthews, J.; Heverly, M.; Morrison, J.; Townsend, J.; Ahmad, N.; Sirota, A.; Cooper, B. ATHLETE: A cargo handling and manipulation robot for the moon. J. Field Robot. 2007, 24, 421–434. [Google Scholar] [CrossRef]
- Chen, J.; Liang, Z.; Zhu, Y.; Zhao, J. Improving Kinematic Flexibility and Walking Performance of a Six-legged Robot by Rationally Designing Leg Morphology. J. Bionic Eng. 2019, 16, 608–620. [Google Scholar] [CrossRef]
- Weihmann, T. Survey of biomechanical aspects of arthropod terrestrialisation – Substrate bound legged locomotion. Arthropod Struct. Dev. 2020, 59, 100983. [Google Scholar] [CrossRef] [PubMed]
- Bullock, J.M.; Drechsler, P.; Federle, W. Comparison of smooth and hairy attachment pads in insects: Friction, adhesion and mechanisms for direction-dependence. J. Exp. Biol. 2008, 211, 3333–3343. [Google Scholar] [CrossRef]
- Clemente, C.J.; Goetzke, H.H.; Bullock, J.M.; Sutton, G.P.; Burrows, M.; Federle, W. Jumping without slipping: Leafhoppers (Hemiptera: Cicadellidae) possess special tarsal structures for jumping from smooth surfaces. J. R. Soc. Interface 2017, 14. [Google Scholar] [CrossRef]
- Dixey, L.; Gardiner, P. LXIII.—Heterogony in Messor barbarus L. var. capitatus Latreille. J. Nat. Hist. 1934, 13, 619–627. [Google Scholar] [CrossRef]
- Canio, G.D.; Stoyanov, S.; Larsen, J.C.; Hallam, J.; Kovalev, A.; Kleinteich, T.; Gorb, S.N.; Manoonpong, P. A robot leg with compliant tarsus and its neural control for efficient and adaptive locomotion on complex terrains. Artif. Life Robot. 2016, 21, 274–281. [Google Scholar] [CrossRef]
- DeMario, A.; Zhao, J. Development and analysis of a three-dimensional printed miniature walking robot with soft joints and links. J. Mech. Robot. 2018, 10, 1–10. [Google Scholar] [CrossRef]
- Sun, J.; Zhao, J. An Adaptive Walking Robot with Reconfigurable Mechanisms Using Shape Morphing Joints. IEEE Robot. Autom. Lett. 2019, 4, 724–731. [Google Scholar] [CrossRef]
- Kram, R.; Wong, B.; Full, R.J. Three-dimensional kinematics and limb kinetic energy of running cockroaches. J. Exp. Biol. 1997, 200, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.G.; Ingber, D.E. Unexpected strength and toughness in chitosan-fibroin laminates inspired by insect cuticle. Adv. Mater. 2012, 24, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Q.; Yang, Q.; Zhao, J.; Gui, S. Dynamic model and performance analysis of rigid-flexible coupling four-bar leg mechanism for small scale bio-inspired jumping robot. Microsyst. Technol. 2019, 25, 3269–3285. [Google Scholar] [CrossRef]
- Li, F.; Liu, W.; Fu, X.; Bonsignori, G.; Scarfogliero, U.; Stefanini, C.; Dario, P. Jumping like an insect: Design and dynamic optimization of a jumping mini robot based on bio-mimetic inspiration. Mechatronics 2012, 22, 167–176. [Google Scholar] [CrossRef]
- Kitano, S.; Hirose, S.; Horigome, A.; Endo, G. TITAN-XIII: Sprawling-type quadruped robot with ability of fast and energy-efficient walking. ROBOMECH J. 2016, 3, 1–16. [Google Scholar] [CrossRef]
- Merienne, H. Biomécanique et Energétique de la Locomotion et du Transport de Charge Chez la Fourmi. Ph.D. Thesis, University of Toulouse 3 (Paul Sabatier), Doctoral School “Science Ecologiques, Vétérinaires, Agronomiques et Bioingénieries”, Toulouse, France, 2019. [Google Scholar]
- Billeschou, P.; Bijma, N.N.; Larsen, L.B.; Gorb, S.N.; Larsen, J.C.; Manoonpong, P. Framework for developing bio-inspired morphologies for walking robots. Appl. Sci. 2020, 10, 6986. [Google Scholar] [CrossRef]
- Ríos, V.L.; Özdil, P.G.; Ramalingasetty, S.T.; Arreguit, J.; Ijspeert, A.J.; Ramdya, P. NeuroMechFly, a neuromechanical model of adult Drosophila melanogaster. bioRxiv 2021. [Google Scholar] [CrossRef]
- Nishii, J. An analytical estimation of the energy cost for legged locomotion. J. Theor. Biol. 2006, 238, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Kottege, N.; Parkinson, C.; Moghadam, P.; Elfes, A.; Singh, S.P. Energetics-informed hexapod gait transitions across terrains. In Proceedings of the 2015 IEEE International Conference on Robotics and Automation (ICRA), Seattle, WA, USA, 26–30 May 2015; pp. 5140–5147. [Google Scholar]
- Pontzer, H. A unified theory for the energy cost of legged locomotion. Biol. Lett. 2016, 12, 20150935. [Google Scholar] [CrossRef]
- Tucker, V.A. The energetic cost of moving about: Walking and running are extremely inefficient forms of locomotion. Much greater efficiency is achieved by birds, fish—And bicyclists. Am. Sci. 1975, 63, 413–419. [Google Scholar]
- Alexander, R.M. Models and the scaling of energy costs for locomotion. J. Exp. Biol. 2005, 208, 1645–1652. [Google Scholar] [CrossRef] [PubMed]
- Seok, S.; Wang, A.; Chuah, M.Y.; Otten, D.; Lang, J.; Kim, S. Design principles for highly efficient quadrupeds and implementation on the MIT Cheetah robot. In Proceedings of the 2013 IEEE International Conference on Robotics and Automation, Karlsruhe, Germany, 6–10 May 2013; pp. 3307–3312. [Google Scholar]
- Vanderborght, B.; Albu-Schäffer, A.; Bicchi, A.; Burdet, E.; Caldwell, D.G.; Carloni, R.; Catalano, M.; Eiberger, O.; Friedl, W.; Ganesh, G. Variable impedance actuators: A review. Robot. Auton. Syst. 2013, 61, 1601–1614. [Google Scholar] [CrossRef]
- Hunter, I.W.; Lafontaine, S. A comparison of muscle with artificial actuators. In Proceedings of the Technical Digest IEEE Solid-State Sensor and Actuator Workshop, Hilton Head, SC, USA, 22–25 June 1992; pp. 178–185. [Google Scholar]
- Liang, W.; Liu, H.; Wang, K.; Qian, Z.; Ren, L.; Ren, L. Comparative study of robotic artificial actuators and biological muscle. Adv. Mech. Eng. 2020, 12, 1687814020933409. [Google Scholar] [CrossRef]
- Rothemund, P.; Kellaris, N.; Mitchell, S.K.; Acome, E.; Keplinger, C. HASEL Artificial Muscles for a New Generation of Lifelike Robots—Recent Progress and Future Opportunities. Adv. Mater. 2021, 33, 2003375. [Google Scholar] [CrossRef] [PubMed]
- El-Atab, N.; Mishra, R.B.; Al-Modaf, F.; Joharji, L.; Alsharif, A.A.; Alamoudi, H.; Diaz, M.; Qaiser, N.; Hussain, M.M. Soft Actuators for Soft Robotic Applications: A Review. Adv. Intell. Syst. 2020, 2, 2000128. [Google Scholar] [CrossRef]
- Mitchell, S.K.; Wang, X.; Acome, E.; Martin, T.; Ly, K.; Kellaris, N.; Venkata, V.G.; Keplinger, C. An Easy-to-Implement Toolkit to Create Versatile and High-Performance HASEL Actuators for Untethered Soft Robots. Adv. Sci. 2019, 6, 1900178. [Google Scholar] [CrossRef]
- Taghavi, M.; Helps, T.; Rossiter, J. Electro-ribbon actuators and electro-origami robots. Sci. Robot. 2018, 3. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Phung, H.; Hoang, P.T.; Nguyen, T.D.; Jung, H.; Moon, H.; Koo, J.C.; Choi, H.R. A novel bioinspired hexapod robot developed by soft dielectric elastomer actuators. In Proceedings of the 2017 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Vancouver, BC, Canada, 24–28 September 2017; pp. 6233–6238. [Google Scholar]
- Schneider, A.; Schmucker, U. Force Sensing for Multi-Legged Walking Robots: Theory and Experiments Part 1: Overview and Force Sensing. In Mobile Robotics, Moving Intelligence; IntechOpen: London, UK, 2006; pp. 447–470. [Google Scholar]
- Billeschou, P.; Albertsen, C.; Larsen, J.C.; Manoonpong, P. A Low-Cost, Compact, Sealed, Three-Axis Force/Torque Sensor for Walking Robots. IEEE Sens. J. 2021, 21, 8916–8926. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, R.; Li, C.; Zang, X.; Zhang, X.; Jin, H.; Zhao, J. A force-sensing system on legs for biomimetic hexapod robots interacting with unstructured terrain. Sensors 2017, 17, 1514. [Google Scholar] [CrossRef] [PubMed]
- Bachega, R.P.; Pires, R.; Campo, A.B. Force sensing to control a bio-inspired walking robot. IFAC Proc. Vol. IFAC-Pap. 2013, 46, 105–109. [Google Scholar] [CrossRef]
- Dallmann, C.J.; Hoinville, T.; Dürr, V.; Schmitz, J. A load-based mechanism for inter-leg coordination in insects. Proc. R. Soc. B Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed]
- Kaliyamoorthy, S.; Quinn, R.D.; Zill, S.N. Force sensors in hexapod locomotion. Int. J. Robot. Res. 2005, 24, 563–574. [Google Scholar] [CrossRef]
- Yang, M.; Kang, R.; Chen, Y. A highly mobile crawling robot inspired by hexapod insects. In Proceedings of the 2019 IEEE International Conference on Robotics and Biomimetics, ROBIO 2019, Dali, China, 6–8 December 2019; pp. 1797–1802. [Google Scholar] [CrossRef]
- Nonami, K.; Barai, R.K.; Irawan, A.; Daud, M.R. Hydraulically Actuated Hexapod Robots: Design, Implementation and Control; Springer: Tokyo, Japan, 2014; Volume 66. [Google Scholar] [CrossRef]
- Clark, J.E.; Goldman, D.I.; Lin, P.C.; Lynch, G.; Chen, T.S.; Komsuoglu, H.; Full, R.J.; Koditschek, D. Design of a bio-inspired dynamical vertical climbing robot. Robot. Sci. Syst. 2008, 3, 9–16. [Google Scholar] [CrossRef]
- Jayaram, K.; Shum, J.; Castellanos, S.; Helbling, E.F.; Wood, R.J. Scaling down an insect-size microrobot, HAMR-VI into HAMR-Jr. In Proceedings of the 2020 IEEE International Conference on Robotics and Automation, Paris, France, 31 May–31 August 2020; pp. 10305–10311. [Google Scholar] [CrossRef]
- Stopp, S.; Wolff, T.; Irlinger, F.; Lueth, T. A new method for printer calibration and contour accuracy manufacturing with 3D-print technology. Rapid Prototyp. J. 2008, 14, 167–172. [Google Scholar] [CrossRef]
- Zhu, Z.; Kin Tam, T.; Sun, F.; You, C.; Percival Zhang, Y.H. A high-energy-density sugar biobattery based on a synthetic enzymatic pathway. Nat. Commun. 2014, 5, 3026. [Google Scholar] [CrossRef]
- Yang, X.; Chang, L.; Pérez-Arancibia, N.O. An 88-milligram insect-scale autonomous crawling robot driven by a catalytic artificial muscle. Sci. Robot. 2020, 5, 1–14. [Google Scholar] [CrossRef]
- Aoi, S.; Manoonpong, P.; Ambe, Y.; Matsuno, F.; Wörgötter, F. Adaptive control strategies for interlimb coordination in legged robots: A review. Front. Neurorobot. 2017, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Huerta, C.V.; Xiong, X.; Billeschou, P.; Manoonpong, P. Adaptive Neuromechanical Control for Robust Behaviors of Bio-Inspired Walking Robots. In International Conference on Neural Information Processing; Springer: Cham, Switzerland, 2020; pp. 775–786. [Google Scholar]
- Xiong, X.; Wörgötter, F.; Manoonpong, P. Adaptive and energy efficient walking in a hexapod robot under neuromechanical control and sensorimotor learning. IEEE Trans. Cybern. 2015, 46, 2521–2534. [Google Scholar] [CrossRef]
- Xiong, X.; Wörgötter, F.; Manoonpong, P. Virtual agonist-antagonist mechanisms produce biological muscle-like functions. Ind. Robot. Int. J. 2014, 41, 340–346. [Google Scholar] [CrossRef]
- Cruse, H.; Kindermann, T.; Schumm, M.; Dean, J.; Schmitz, J. Walknet—A biologically inspired network to control six-legged walking. Neural Netw. 1998, 11, 1435–1447. [Google Scholar] [CrossRef]
- Pearson, K.; Franklin, R. Characteristics of leg movements and patterns of coordination in locusts walking on rough terrain. Int. J. Robot. Res. 1984, 3, 101–112. [Google Scholar] [CrossRef]
- Espenschied, K.S.; Quinn, R.D.; Beer, R.D.; Chiel, H.J. Biologically based distributed control and local reflexes improve rough terrain locomotion in a hexapod robot. Robot. Auton. Syst. 1996, 18, 59–64. [Google Scholar] [CrossRef]
- Wilson, D.M. Insect walking. Annu. Rev. Entomol. 1966, 11, 103–122. [Google Scholar] [CrossRef]
- Ferrell, C. A comparison of three insect-inspired locomotion controllers. Robot. Auton. Syst. 1995, 16, 135–159. [Google Scholar] [CrossRef]
- Graham, D. A behavioural analysis of the temporal organisation of walking movements in the 1st instar and adult stick insect (Carausius morosus). J. Comp. Physiol. 1972, 81, 23–52. [Google Scholar] [CrossRef]
- Cruse, H. What mechanisms coordinate leg movement in walking arthropods? Trends Neurosci. 1990, 13, 15–21. [Google Scholar] [CrossRef]
- Delcomyn, F. The locomotion of the cockroach Periplaneta americana. J. Exp. Biol. 1971, 54, 443–452. [Google Scholar] [CrossRef]
- Wosnitza, A.; Bockemühl, T.; Dübbert, M.; Scholz, H.; Büschges, A. Inter-leg coordination in the control of walking speed in Drosophila. J. Exp. Biol. 2013, 216, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Ambe, Y.; Aoi, S.; Nachstedt, T.; Manoonpong, P.; Wörgötter, F.; Matsuno, F. Simple analytical model reveals the functional role of embodied sensorimotor interaction in hexapod gaits. PLoS ONE 2018, 13, e0192469. [Google Scholar] [CrossRef] [PubMed]
- Cruse, H.; Dürr, V.; Schilling, M.; Schmitz, J. Principles of insect locomotion. In Spatial Temporal Patterns for Action-Oriented Perception in Roving Robots; Springer: Berlin/Heidelberg, Germany, 2009; pp. 43–96. [Google Scholar]
- Büschges, A.; Akay, T.; Gabriel, J.P.; Schmidt, J. Organizing network action for locomotion: Insights from studying insect walking. Brain Res. Rev. 2008, 57, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Bässler, U.; Wegner, U. Motor output of the denervated thoracic ventral nerve cord in the stick insect Carausius morosus. J. Exp. Biol. 1983, 105, 127–145. [Google Scholar] [CrossRef]
- Mantziaris, C.; Bockemühl, T.; Büschges, A. Central pattern generating networks in insect locomotion. Dev. Neurobiol. 2020, 80, 16–30. [Google Scholar] [CrossRef]
- Bässler, U.; Büschges, A. Pattern generation for stick insect walking movements—multisensory control of a locomotor program. Brain Res. Rev. 1998, 27, 65–88. [Google Scholar] [CrossRef]
- Berg, E.M.; Hooper, S.L.; Schmidt, J.; Büschges, A. A leg-local neural mechanism mediates the decision to search in stick insects. Curr. Biol. 2015, 25, 2012–2017. [Google Scholar] [CrossRef]
- Bucher, D.; Akay, T.; DiCaprio, R.A.; Büschges, A. Interjoint coordination in the stick insect leg-control system: The role of positional signaling. J. Neurophysiol. 2003, 89, 1245–1255. [Google Scholar] [CrossRef]
- Akay, T.; Ludwar, B.C.; Göritz, M.L.; Schmitz, J.; Büschges, A. Segment specificity of load signal processing depends on walking direction in the stick insect leg muscle control system. J. Neurosci. 2007, 27, 3285–3294. [Google Scholar] [CrossRef]
- Daun-Gruhn, S.; Tóth, T.I. An inter-segmental network model and its use in elucidating gait-switches in the stick insect. J. Comput. Neurosci. 2011, 31, 43–60. [Google Scholar] [CrossRef]
- Szczecinski, N.S.; Quinn, R.D. MantisBot changes stepping speed by entraining CPGs to positive velocity feedback. In Conference on Biomimetic and Biohybrid Systems; Springer International Publishing: Cham, Switzerland, 2017; pp. 440–452. [Google Scholar]
- Niemeier, M.; Jeschke, M.; Dürr, V. Effect of thoracic connective lesion on inter-leg coordination in freely walking stick insects. Front. Bioeng. Biotechnol. 2021, 9, 628998. [Google Scholar] [CrossRef] [PubMed]
- Webb, B. Neural mechanisms for prediction: Do insects have forward models? Trends Neurosci. 2004, 27, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Bläsing, B.; Cruse, H. Mechanisms of stick insect locomotion in a gap-crossing paradigm. J. Comp. Physiol. A 2004, 190, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Möhl, B. ‘Biological noise’and plasticity of sensorimotor pathways in the locust flight system. J. Comp. Physiol. A 1989, 166, 75–82. [Google Scholar] [CrossRef]
- Heisenberg, M.; Wolf, R. Reafferent control of optomotor yaw torque inDrosophila melanogaster. J. Comp. Physiol. A 1988, 163, 373–388. [Google Scholar] [CrossRef]
- Steingrube, S.; Timme, M.; Wörgötter, F.; Manoonpong, P. Self-organized adaptation of a simple neural circuit enables complex robot behaviour. Nat. Phys. 2010, 6, 224–230. [Google Scholar] [CrossRef]
- Manoonpong, P.; Parlitz, U.; Wörgötter, F. Neural control and adaptive neural forward models for insect-like, energy-efficient, and adaptable locomotion of walking machines. Front. Neural Circuits 2013, 7, 12. [Google Scholar] [CrossRef]
- Schilling, M.; Cruse, H. Decentralized control of insect walking: A simple neural network explains a wide range of behavioral and neurophysiological results. PLoS Comput. Biol. 2020, 16, e1007804. [Google Scholar] [CrossRef]
- Leung, B.; Bijma, N.; Baird, E.; Dacke, M.; Gorb, S.; Manoonpong, P. Rules for the Leg Coordination of Dung Beetle Ball Rolling Behaviour. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Mizrahi, J. Review: Mechanical Impedance and Its Relations to Motor Control, Limb Dynamics, and Motion Biomechanics. J. Med. Biol. Eng. 2015, 35, 1–20. [Google Scholar] [CrossRef]
- Bressler, B.H.; Clinch, N.F. The compliance of contracting skeletal muscle. J. Physiol. 1974, 237, 477–493. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cruse, H.; Kühn, S.; Park, S.; Schmitz, J. Adaptive control for insect leg position: Controller properties depend on substrate compliance. J. Comp. Physiol. A 2004, 190, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Knoll, A.; Röhrbein, F.; Kuhn, A.; Akl, M.; Sharma, K. Neurorobotics. Informatik-Spektrum 2017, 40, 161–164. [Google Scholar] [CrossRef]
- Hill, A.V. The mechanics of active muscle. Proc. R. Soc. Lond. Ser. B—Biol. Sci. 1953, 141, 104–117. [Google Scholar] [CrossRef]
- Proctor, J.L.; Holmes, P. The effects of feedback on stability and maneuverability of a phase-reduced model for cockroach locomotion. Biol. Cybern. 2018, 112, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Lin, J.; Wöhrl, T.; Liao, M. A Neuro-Musculo-Skeletal Model for Insects With Data-driven Optimization. Sci. Rep. 2018, 8, 2129. [Google Scholar] [CrossRef]
- Naris, M.; Szczecinski, N.S.; Quinn, R.D. A neuromechanical model exploring the role of the common inhibitor motor neuron in insect locomotion. Biol. Cybern. 2020, 114, 23–41. [Google Scholar] [CrossRef]
- Dickinson, M.H.; Farley, C.T.; Full, R.J.; Koehl, M.A.R.; Kram, R.; Lehman, S. How Animals Move: An Integrative View. Science 2000, 288, 100–106. [Google Scholar] [CrossRef]
- Ahn, A.N. How muscles function—The work loop technique. J. Exp. Biol. 2012, 215, 1051–1052. [Google Scholar] [CrossRef]
- Xiong, X.; Manoonpong, P. Muscles Can be Brakes: The Work Loop Technique for Stable Muscle-like Control. In Proceedings of the International Symposium on Adaptive Motion of Animals and Machines (AMAM 2019), Lausanne, Switzerland, 20–23 August 2019. [Google Scholar] [CrossRef]
- Xiong, X.; Wörgötter, F.; Manoonpong, P. Neuromechanical control for hexapedal robot walking on challenging surfaces and surface classification. Robot. Auton. Syst. 2014, 62, 1777–1789. [Google Scholar] [CrossRef]
- Braun, J.M.; Manoonpong, P.; Xiong, X. Editorial: Biology-Inspired Engineering and Engineering-Inspired Biology. Front. Neurorobot. 2020, 14, 614683. [Google Scholar] [CrossRef] [PubMed]
- Owaki, D.; Goda, M.; Miyazawa, S.; Ishiguro, A. A minimal model describing hexapedal interlimb coordination: The Tegotae-based approach. Front. Neurorobot. 2017, 11, 29. [Google Scholar] [CrossRef]
- Miguel-Blanco, A.; Manoonpong, P. General distributed neural control and sensory adaptation for self-organized locomotion and fast adaptation to damage of walking robots. Front. Neural Circuits 2020, 14, 46. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Xiong, X.; Dai, Z.; Owaki, D.; Manoonpong, P. A Comparative Study of Adaptive Interlimb Coordination Mechanisms for Self-Organized Robot Locomotion. Front. Robot. AI 2021, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Cruse, H.; Dürr, V.; Schmitz, J. Insect walking is based on a decentralized architecture revealing a simple and robust controller. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2007, 365, 221–250. [Google Scholar] [CrossRef]
- Ayali, A.; Borgmann, A.; Büschges, A.; Couzin-Fuchs, E.; Daun-Gruhn, S.; Holmes, P. The comparative investigation of the stick insect and cockroach models in the study of insect locomotion. Curr. Opin. Insect Sci. 2015, 12, 1–10. [Google Scholar] [CrossRef]
- Bidaye, S.S.; Bockemühl, T.; Büschges, A. Six-legged walking in insects: How CPGs, peripheral feedback, and descending signals generate coordinated and adaptive motor rhythms. J. Neurophysiol. 2018, 119, 459–475. [Google Scholar] [CrossRef]
- Bjelonic, M.; Homberger, T.; Kottege, N.; Borges, P.; Chli, M.; Beckerle, P. Autonomous navigation of hexapod robots with vision-based controller adaptation. In Proceedings of the 2017 IEEE International Conference on Robotics and Automation (ICRA), Singapore, 29 May–3 June 2017; pp. 5561–5568. [Google Scholar]
- Hafner, R.; Hertweck, T.; Klöppner, P.; Bloesch, M.; Neunert, M.; Wulfmeier, M.; Tunyasuvunakool, S.; Heess, N.; Riedmiller, M. Towards General and Autonomous Learning of Core Skills: A Case Study in Locomotion. arXiv 2020, arXiv:2008.12228. [Google Scholar]
- Pretorius, C.J.; du Plessis, M.C.; Gonsalves, J.W. Evolutionary robotics applied to hexapod locomotion: A comparative study of simulation techniques. J. Intell. Robot. Syst. 2019, 96, 363–385. [Google Scholar] [CrossRef]
- Ijspeert, A.J. Central pattern generators for locomotion control in animals and robots: A review. Neural Netw. 2008, 21, 642–653. [Google Scholar] [CrossRef]
- Fischer, J.; Pasemann, F.; Manoonpong, P. Neuro-controllers for walking machines—An evolutionary approach to robust behavior. In Climbing and Walking Robots; Springer: Berlin/Heidelberg, Germany, 2005; pp. 97–102. [Google Scholar]
- Bai, L.; Hu, H.; Chen, X.; Sun, Y.; Ma, C.; Zhong, Y. CPG-Based Gait Generation of the Curved-Leg Hexapod Robot with Smooth Gait Transition. Sensors 2019, 19, 3705. [Google Scholar] [CrossRef]
- Arena, P.; Fortuna, L.; Frasca, M.; Patane, L. A CNN-based chip for robot locomotion control. IEEE Trans. Circuits Syst. I Regul. Pap. 2005, 52, 1862–1871. [Google Scholar] [CrossRef]
- Ekeberg, Ö.; Blümel, M.; Büschges, A. Dynamic simulation of insect walking. Arthropod Struct. Dev. 2004, 33, 287–300. [Google Scholar] [CrossRef]
- Brown, T.G. On the nature of the fundamental activity of the nervous centres; together with an analysis of the conditioning of rhythmic activity in progression, and a theory of the evolution of function in the nervous system. J. Physiol. 1914, 48, 18–46. [Google Scholar] [CrossRef] [PubMed]
- Hellgren, J.; Grillner, S.; Lansner, A. Computer simulation of the segmental neural network generating locomotion in lamprey by using populations of network interneurons. Biol. Cybern. 1992, 68, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ekeberg, Ö. A combined neuronal and mechanical model of fish swimming. Biol. Cybern. 1993, 69, 363–374. [Google Scholar] [CrossRef]
- Arena, P. The central pattern generator: A paradigm for artificial locomotion. Soft Comput. 2000, 4, 251–266. [Google Scholar] [CrossRef]
- Pasemann, F.; Hild, M.; Zahedi, K. SO (2)-networks as neural oscillators. In International Work-Conference on Artificial Neural Networks; Springer: Berlin/Heidelberg, Germany, 2003; pp. 144–151. [Google Scholar]
- Van der Pol, B. LXXXVIII. On “relaxation-oscillations”. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1926, 2, 978–992. [Google Scholar] [CrossRef]
- Matsuoka, K. Sustained oscillations generated by mutually inhibiting neurons with adaptation. Biol. Cybern. 1985, 52, 367–376. [Google Scholar] [CrossRef]
- Taga, G.; Yamaguchi, Y.; Shimizu, H. Self-organized control of bipedal locomotion by neural oscillators in unpredictable environment. Biol. Cybern. 1991, 65, 147–159. [Google Scholar] [CrossRef]
- Righetti, L.; Buchli, J.; Ijspeert, A.J. Dynamic hebbian learning in adaptive frequency oscillators. Phys. D Nonlinear Phenom. 2006, 216, 269–281. [Google Scholar] [CrossRef]
- Aoi, S.; Egi, Y.; Sugimoto, R.; Yamashita, T.; Fujiki, S.; Tsuchiya, K. Functional roles of phase resetting in the gait transition of a biped robot from quadrupedal to bipedal locomotion. IEEE Trans. Robot. 2012, 28, 1244–1259. [Google Scholar] [CrossRef]
- Owaki, D.; Kano, T.; Nagasawa, K.; Tero, A.; Ishiguro, A. Simple robot suggests physical interlimb communication is essential for quadruped walking. J. R. Soc. Interface 2013, 10, 20120669. [Google Scholar] [CrossRef]
- Nachstedt, T.; Tetzlaff, C.; Manoonpong, P. Fast dynamical coupling enhances frequency adaptation of oscillators for robotic locomotion control. Front. Neurorobot. 2017, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Thor, M.; Manoonpong, P. Error-based learning mechanism for fast online adaptation in robot motor control. IEEE Trans. Neural Netw. Learn. Syst. 2019, 31, 2042–2051. [Google Scholar] [CrossRef]
- Chuthong, T.; Leung, B.; Tiraborisute, K.; Ngamkajornwiwat, P.; Manoonpong, P.; Dilokthanakul, N. Dynamical State Forcing on Central Pattern Generators for Efficient Robot Locomotion Control. In International Conference on Neural Information Processing; Springer: Cham, Switzerland, 2020; pp. 799–810. [Google Scholar]
- Buchli, J.; Righetti, L.; Ijspeert, A.J. Engineering entrainment and adaptation in limit cycle systems. Biol. Cybern. 2006, 95, 645–664. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, T.; Ewald, A.; von Twickel, A.; Bueschges, A. Controlling legs for locomotion—Insights from robotics and neurobiology. Bioinspir. Biomim. 2015, 10, 041001. [Google Scholar] [PubMed]
- Thor, M.; Kulvicius, T.; Manoonpong, P. Generic neural locomotion control framework for legged robots. IEEE Trans. Neural Netw. Learn. Syst. 2020, 32, 4013–4025. [Google Scholar] [CrossRef]
- Goldschmidt, D.; Wörgötter, F.; Manoonpong, P. Biologically-inspired adaptive obstacle negotiation behavior of hexapod robots. Front. Neurorobot. 2014, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Grinke, E.; Tetzlaff, C.; Wörgötter, F.; Manoonpong, P. Synaptic plasticity in a recurrent neural network for versatile and adaptive behaviors of a walking robot. Front. Neurorobot. 2015, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Goldschmidt, D.; Wörgötter, F.; Manoonpong, P. Distributed recurrent neural forward models with synaptic adaptation and CPG-based control for complex behaviors of walking robots. Front. Neurorobot. 2015, 9, 10. [Google Scholar] [CrossRef]
- Sun, X.; Yue, S.; Mangan, M. A decentralised neural model explaining optimal integration of navigational strategies in insects. eLife 2020, 9, e54026. [Google Scholar] [CrossRef] [PubMed]
- Roennau, A.; Kerscher, T.; Ziegenmeyer, M.; Zöllner, J.M.; Dillmann, R. Adaptation of a six-legged walking robot to its local environment. In Robot Motion and Control 2009; Springer: London, UK, 2009; pp. 155–164. [Google Scholar]
- Paskarbeit, J.; Schilling, M.; Schmitz, J.; Schneider, A. Obstacle crossing of a real, compliant robot based on local evasion movements and averaging of stance heights using singular value decomposition. In Proceedings of the 2015 IEEE International Conference on Robotics and Automation (ICRA), Seattle, WA, USA, 26–30 May 2015; pp. 3140–3145. [Google Scholar]
- Thor, M.; Strøm-Hansen, T.; Larsen, L.B.; Kovalev, A.; Gorb, S.N.; Baird, E.; Manoonpong, P. A dung beetle-inspired robotic model and its distributed sensor-driven control for walking and ball rolling. Artif. Life Robot. 2018, 23, 435–443. [Google Scholar] [CrossRef]
- Schilling, M.; Cruse, H. ReaCog, a minimal cognitive controller based on recruitment of reactive systems. Front. Neurorobot. 2017, 11, 3. [Google Scholar] [CrossRef]
- Tieck, J.C.V.; Rutschke, J.; Kaiser, J.; Schulze, M.; Buettner, T.; Reichard, D.; Roennau, A.; Dillmann, R. Combining spiking motor primitives with a behaviour-based architecture to model locomotion for six-legged robots. In Proceedings of the 2019 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Macau, China, 3–9 November 2019; pp. 4161–4168. [Google Scholar]
- von Twickel, A.; Büschges, A.; Pasemann, F. Deriving neural network controllers from neuro-biological data: Implementation of a single-leg stick insect controller. Biol. Cybern. 2011, 104, 95–119. [Google Scholar] [CrossRef] [PubMed]
- Lewinger, W.A.; Quinn, R.D. Neurobiologically-based control system for an adaptively walking hexapod. Ind. Robot. 2011, 38, 258–263. [Google Scholar] [CrossRef]
- Yu, H.; Gao, H.; Deng, Z. Enhancing adaptability with local reactive behaviors for hexapod walking robot via sensory feedback integrated central pattern generator. Robot. Auton. Syst. 2020, 124, 103401. [Google Scholar] [CrossRef]
- Szadkowski, R.; Prágr, M.; Faigl, J. Self-Learning Event Mistiming Detector Based on Central Pattern Generator. Front. Neurorobot. 2021, 15, 5. [Google Scholar] [CrossRef]
- Ngamkajornwiwat, P.; Homchanthanakul, J.; Teerakittikul, P.; Manoonpong, P. Bio-inspired adaptive locomotion control system for online adaptation of a walking robot on complex terrains. IEEE Access 2020, 8, 91587–91602. [Google Scholar] [CrossRef]
- Homchanthanakul, J.; Ngamkajornwiwat, P.; Teerakittikul, P.; Manoonpong, P. Neural control with an artificial hormone system for energy-efficient compliant terrain locomotion and adaptation of walking robots. In Proceedings of the 2019 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Macau, China, 3–8 November 2019; pp. 5475–5482. [Google Scholar]
- Cruse, H.; Bartling, C. Movement of joint angles in the legs of a walking insect, Carausius morosus. J. Insect Physiol. 1995, 41, 761–771. [Google Scholar] [CrossRef]
- Belter, D.; Skrzypczyński, P. Rough terrain mapping and classification for foothold selection in a walking robot. J. Field Robot. 2011, 28, 497–528. [Google Scholar] [CrossRef]
- Liu, T.; Chen, W.; Wang, J.; Wu, X. Terrain analysis and locomotion control of a hexapod robot on uneven terrain. In Proceedings of the 2014 9th IEEE Conference on Industrial Electronics and Applications, Hangzhou, China, 9–11 June 2014; pp. 1959–1964. [Google Scholar]
- Buchanan, R.; Bandyopadhyay, T.; Bjelonic, M.; Wellhausen, L.; Hutter, M.; Kottege, N. Walking posture adaptation for legged robot navigation in confined spaces. IEEE Robot. Autom. Lett. 2019, 4, 2148–2155. [Google Scholar] [CrossRef]
- Murata, Y.; Inagaki, S.; Suzuki, T. Development of an adaptive hexapod robot based on Follow-the-contact-point gait control and Timekeeper control. In Proceedings of the 2019 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Macau, China, 3–8 November 2019; pp. 3321–3327. [Google Scholar]
- Hu, N.; Li, S.; Zhu, Y.; Gao, F. Constrained model predictive control for a hexapod robot walking on irregular terrain. J. Intell. Robot. Syst. 2019, 94, 179–201. [Google Scholar] [CrossRef]
- Xiong, X.; Wörgötter, F.; Manoonpong, P. A Simplified Variable Admittance Controller Based on a Virtual Agonist-Antagonist Mechanism for Robot Joint Control. In Nature-Inspired Mobile Robotics; World Scientific: Singapore, 2013; pp. 281–288. [Google Scholar]
- Xiong, X.; Manoonpong, P. Adaptive Motor Control for Human-like Spatial-temporal Adaptation. In Proceedings of the 2018 IEEE International Conference on Robotics and Biomimetics (ROBIO), Kuala Lumpur, Malaysia, 12–15 December 2018; pp. 2107–2112. [Google Scholar]
- Holmes, P.; Full, R.; Koditschek, D.; Guckenheimer, J. The Dynamics of Legged Locomotion: Models, Analyses, and Challenges. SIAM Rev. 2006, 48, 207–304. [Google Scholar] [CrossRef]
- Sun, T.; Xiong, X.; Dai, Z.; Manoonpong, P. Small-Sized Reconfigurable Quadruped Robot With Multiple Sensory Feedback for Studying Adaptive and Versatile Behaviors. Front. Neurorobot. 2020, 14, 14. [Google Scholar] [CrossRef]
- Kukillaya, R.; Proctor, J.; Holmes, P. Neuromechanical models for insect locomotion: Stability, maneuverability, and proprioceptive feedback. Chaos Interdiscip. J. Nonlinear Sci. 2009, 19. [Google Scholar] [CrossRef]
- Szczecinski, N.S.; Goldsmith, C.A.; Young, F.R.; Quinn, R.D. Tuning a Robot Servomotor to Exhibit Muscle-Like Dynamics. In Biomimetic and Biohybrid Systems, Living Machines 2019; Springer: Cham, Switzerland, 2019; pp. 254–265. [Google Scholar]
- Ribak, G. Insect-inspired jumping robots: Challenges and solutions to jump stability. Curr. Opin. Insect Sci. 2020, 42, 32–38. [Google Scholar] [CrossRef]
- Schilling, M.; Konen, K.; Ohl, F.W.; Korthals, T. Decentralized Deep Reinforcement Learning for a Distributed and Adaptive Locomotion Controller of a Hexapod Robot. In Proceedings of the 2020 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Las Vegas, NV, USA, 24–30 October 2020; pp. 5335–5342. [Google Scholar] [CrossRef]
- Ting, J.; Fang, Y.; Lele, A.; Raychowdhury, A. Bio-inspired gait imitation of hexapod robot using event-based vision sensor and spiking neural network. In Proceedings of the 2020 International Joint Conference on Neural Networks (IJCNN), Glasgow, UK, 19–24 July 2020; pp. 1–7. [Google Scholar]
- Cully, A.; Clune, J.; Tarapore, D.; Mouret, J.B. Robots that can adapt like animals. Nature 2015, 521, 503–507. [Google Scholar] [CrossRef]
- Parker, G.B. Evolving gaits for hexapod robots using cyclic genetic algorithms. Int. J. Gen. Syst. 2005, 34, 301–315. [Google Scholar] [CrossRef]
- Cully, A.; Mouret, J.B. Evolving a behavioral repertoire for a walking robot. Evol. Comput. 2016, 24, 59–88. [Google Scholar] [CrossRef]
- Azayev, T.; Zimmerman, K. Blind Hexapod Locomotion in Complex Terrain with Gait Adaptation Using Deep Reinforcement Learning and Classification. J. Intell. Robot. Syst. 2020, 99, 1–13. [Google Scholar] [CrossRef]
- Milicka, P.; Cizek, P.; Faigl, J. On Chaotic Oscillator-Based Central Pattern Generator for Motion Control of Hexapod Walking Robot. ITAT 2016, 1649, 131–137. [Google Scholar]
- Chen, W.; Ren, G.; Wang, J.; Liu, D. An adaptive locomotion controller for a hexapod robot: CPG, kinematics and force feedback. Sci. China Inf. Sci. 2014, 57, 1–18. [Google Scholar] [CrossRef]
- Fu, H.; Tang, K.; Li, P.; Zhang, W.; Wang, X.; Deng, G.; Wang, T.; Chen, C. Deep Reinforcement Learning for Multi-contact Motion Planning of Hexapod Robots. In Proceedings of the Thirtieth International Joint Conference on Artificial Intelligence (IJCAI), Montréal, QC, Canada, 21 August 2021; pp. 2381–2388. [Google Scholar]
- Ouyang, W.; Chi, H.; Pang, J.; Liang, W.; Ren, Q. Adaptive Locomotion Control of a Hexapod Robot via Bio-Inspired Learning. Front. Neurorobot. 2021, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Lillicrap, T.P.; Hunt, J.J.; Pritzel, A.; Heess, N.; Erez, T.; Tassa, Y.; Silver, D.; Wierstra, D. Continuous control with deep reinforcement learning. arXiv 2015, arXiv:1509.02971. [Google Scholar]
- Cruse, H.; Schilling, M. Mental States as Emergent Properties From Walking to Consciousness. In Open Mind; MIND Group: Frankfurt am Main, Germany, 2015; pp. 335–373. [Google Scholar] [CrossRef]
- Perry, C.J.; Barron, A.B.; Chittka, L. The frontiers of insect cognition. Curr. Opin. Behav. Sci. 2017, 16, 111–118. [Google Scholar] [CrossRef]
- Chittka, L.; Niven, J. Are Bigger Brains Better? Curr. Biol. 2009, 19, R995–R1008. [Google Scholar] [CrossRef]
- Wolf, R.; Voss, A.; Hein, S.; Heisenberg, M.; Sullivan, G.; Barlow, H.B.; Frisby, J.P.; Horridge, G.A.; Jeeves, M.A. Can a fly ride a bicycle? Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1992, 337, 261–269. [Google Scholar] [CrossRef]
- Dacke, M.; Srinivasan, M. Evidence for counting in insect. Anim. Cogn. 2008, 11, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Srinivasan, M.V.; Zhu, H.; Wong, J. Grouping of visual objects by honeybees. J. Exp. Biol. 2004, 207, 3289–3298. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M.; Zhang, S.; Jenett, A.; Menzel, R.; Srinivasan, M. The concepts of ‘sameness’ and ‘difference’ in an insect. Nature 2001, 410, 930–933. [Google Scholar] [CrossRef]
- Ofstad, T.; Zuker, C.; Reiser, M. Visual Place Learning in Drosophila melanogaster. Nature 2011, 474, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M. Social learning in insects: A higher-order capacity? Front. Behav. Neurosci. 2012, 6, 57. [Google Scholar] [CrossRef]
- Collett, T.; Fry, S.; Fry, S. Sequence learning by honeybees. J. Comp. Physiol. A 1993, 172, 693–706. [Google Scholar] [CrossRef]
- Menzel, R. Serial Position Learning in Honeybees. PLoS ONE 2009, 4, 4694. [Google Scholar] [CrossRef]
- Menzel, R.; Greggers, U.; Smith, A.; Berger, S.; Brandt, R.; Brunke, S.; Bundrock, G.; Hülse, S.; Plümpe, T.; Schaupp, F.; et al. Honey bees navigate according to a map-like spatial memory. Proc. Natl. Acad. Sci. USA 2005, 102, 3040–3045. [Google Scholar] [CrossRef]
- Menzel, R.; Lehmann, K.; Manz, G.; Degen, J.; Koblofsky, M.; Greggers, U. Vector integration and novel shortcutting in honeybee navigation. Apidologie 2012, 43, 229–243. [Google Scholar] [CrossRef][Green Version]
- Menzel, R.; Kirbach, A.; Haass, W.D.; Fischer, B.; Degen, J.; Koblofsky, M.; Lehmann, K.; Reiter, L.; Meyer, H.; Nguyen, H.; et al. A Common Frame of Reference for Learned and Communicated Vectors in Honeybee Navigation. Curr. Biol. CB 2011, 21, 645–650. [Google Scholar] [CrossRef]
- Collett, T. Insect navigation en route to the goal: Multiple strategies for the use of landmarks. J. Exp. Biol. 1996, 199, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Bos, N.; Dreier, S.; Jørgensen, C.; Nielsen, J.; Guerrieri, F. Learning and perceptual similarity among cuticular hydrocarbons in ants. J. Insect Physiol. 2011, 58, 138–146. [Google Scholar] [CrossRef]
- Brembs, B.; Hempel de Ibarra, N. Different parameters support generalization and discrimination learning in Drosophila at the flight simulator. Learn. Mem. (Cold Spring Harb. N.Y.) 2006, 13, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Zentall, T.; Galizio, M.; Critchfield, T. Categorization, concept learning, and behavior analysis: An introduction. J. Exp. Anal. Behav. 2002, 78, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Avarguès-Weber, A.; Dyer, A.G.; Giurfa, M. Conceptualization of above and below relationships by an insect. Proc. R. Soc. B Biol. Sci. 2010, 278, 898–905. [Google Scholar] [CrossRef]
- Bell, A.T.A.; Niven, J.E. Strength of forelimb lateralization predicts motor errors in an insect. Biol. Lett. 2016, 12, 20160547. [Google Scholar] [CrossRef]
- Romano, D.; Benelli, G.; Kavallieratos, N.; Athanassiou, C.; Canale, A.; Stefanini, C. Beetle-robot hybrid interaction: Sex, lateralization and mating experience modulate behavioural responses to robotic cues in the larger grain borer Prostephanus truncatus (Horn). Biol. Cybern. 2020, 114. [Google Scholar] [CrossRef]
- Bockhorst, T.; Homberg, U. Interaction of compass sensing and object-motion detection in the locust central complex. J. Neurophysiol. 2017, 118, 496–506. [Google Scholar] [CrossRef]
- de Bivort, B.L.; van Swinderen, B. Evidence for selective attention in the insect brain. Curr. Opin. Insect Sci. 2016, 15, 9–15. [Google Scholar] [CrossRef]
- Sheehan, M.; Tibbetts, E. Specialized Face Learning Is Associated with Individual Recognition in Paper Wasps. Science 2011, 334, 1272–1275. [Google Scholar] [CrossRef]
- Coolen, I.; Dangles, O.; Casas, J. Social Learning in Noncolonial Insects? Curr. Biol. CB 2005, 15, 1931–1935. [Google Scholar] [CrossRef] [PubMed]
- Sachin, S.; Reuven, D. Social learning about egg-laying substrates in fruitflies. Proc. R. Soc. B Biol. Sci. 2009, 276, 4323–4328. [Google Scholar] [CrossRef]
- Romano, D.; Benelli, G.; Stefanini, C. Opposite valence social information provided by bio-robotic demonstrators shapes selection processes in the green bottle fly. J. R. Soc. Interface 2021, 18, 20210056. [Google Scholar] [CrossRef]
- Pfeiffer, B.D.; Jenett, A.; Hammonds, A.S.; Ngo, T.T.B.; Misra, S.; Murphy, C.; Scully, A.; Carlson, J.W.; Wan, K.H.; Laverty, T.R.; et al. Tools for neuroanatomy and neurogenetics in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 9715–9720. [Google Scholar] [CrossRef]
- Tang, S.; Guo, A. Choice Behavior of Drosophila Facing Contradictory Visual Cues. Science 2001, 294, 1543–1547. [Google Scholar] [CrossRef]
- Schröter, U.; Menzel, R. A new ascending sensory tract to the calyces of the honeybee mushroom body, the subesophageal-calycal tract. J. Comp. Neurol. 2003, 465, 168–178. [Google Scholar] [CrossRef]
- Patanè, L.; Strauss, R.; Arena, P. Biological Investigation of Neural Circuits in the Insect Brain. In Nonlinear Circuits and Systems for Neuro-inspired Robot Control; Springer: Cham, Switzerland, 2018; pp. 1–20. [Google Scholar]
- Turner-Evans, D.B.; Jayaraman, V. The insect central complex. Curr. Biol. 2016, 26, R453–R457. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Mangan, M.; Webb, B. Spatio-Temporal Memory for Navigation in a Mushroom Body Model. In Biomimetic and Biohybrid Systems; Vouloutsi, V., Mura, A., Tauber, F., Speck, T., Prescott, T.J., Verschure, P.F.M.J., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 415–426. [Google Scholar]
- Rapp, H.; Nawrot, M. A spiking neural program for sensorimotor control during foraging in flying insects. Proc. Natl. Acad. Sci. USA 2020, 117, 28412–28421. [Google Scholar] [CrossRef]
- Gerber, B.; Stocker, R. The Drosophila larva as a model for studying chemosensation and chemosensory learning: A review. Chem. Senses 2007, 32, 65–89. [Google Scholar] [CrossRef]
- Koenig, S.; Wolf, R.; Heisenberg, M. Visual Attention in Flies-Dopamine in the Mushroom Bodies Mediates the After-Effect of Cueing. PLoS ONE 2016, 11, 1412. [Google Scholar] [CrossRef] [PubMed]
- Arena, P.; Patanè, L.; Termini, P.S. Modeling attentional loop in the insect Mushroom Bodies. In Proceedings of the 2012 International Joint Conference on Neural Networks (IJCNN), Brisbane, QLD, Australia, 10–15 June 2012; pp. 1–7. [Google Scholar] [CrossRef]
- Zhao, C.; Widmer, Y.; Diegelmann, S.; Petrovici, M.; Sprecher, S.; Senn, W. Predictive olfactory learning in Drosophila. Sci. Rep. 2021, 11, 6795. [Google Scholar] [CrossRef]
- Arena, P.; Patanè, L.; Termini, P.S. Learning expectation in insects: A recurrent spiking neural model for spatio-temporal representation. Neural Netw. 2012, 32, 35–45. [Google Scholar] [CrossRef]
- Arena, P.; Patané, L.; Stornanti, V.; Termini, P.S.; Zäpf, B.; Strauss, R. Modeling the insect mushroom bodies: Application to a delayed match-to-sample task. Special Issue on Autonomous Learning. Neural Netw. 2013, 41, 202. [Google Scholar] [CrossRef]
- Arena, P.; Calí, M.; Patanè, L.; Portera, A.; Strauss, R. A Fly-Inspired Mushroom Bodies Model for Sensory-Motor Control Through Sequence and Subsequence Learning. Int. J. Neural Syst. 2016, 26, 1650035. [Google Scholar] [CrossRef] [PubMed]
- Arena, P.; Calí, M.; Patané, L.; Portera, A.; Strauss, R. Modelling the insect Mushroom Bodies: Application to sequence learning. Neural Netw. 2015, 67, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Arena, P.; Calí, M.; Patané, L.; Portera, A.; Spinosa, A.G. A CNN-based neuromorphic model for classification and decision control. Nonlinear Dyn. 2019, 95, 1999–2017. [Google Scholar] [CrossRef]
- Arena, P.; Patanè, L.; Termini, P. Decision making processes in the fruit fly: A computational model. In Frontiers in Artificial Intelligence and Applications, Neural Nets WIRN11; IOS Press: Amsterdam, The Netherlands, 2011; Volume 234, pp. 284–291. [Google Scholar] [CrossRef]
- Kuntz, S.; Poeck, B.; Sokolowski, M.; Strauss, R. The visual orientation memory of Drosophila requires Foraging (PKG) upstream of Ignorant (RSK2) in ring neurons of the central complex. Learn. Mem. 2012, 19, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Arena, P.; Maceo, S.; Patanè, L.; Strauss, R. A spiking network for spatial memory formation: Towards a fly-inspired ellipsoid body model. In Proceedings of the 2013 International Joint Conference on Neural Networks (IJCNN), Dallas, TX, USA, 4–9 August 2013; pp. 1–6. [Google Scholar] [CrossRef]
- Kakaria, K.S.; de Bivort, B.L. Ring attractor dynamics emerge from a spiking model of the entire protocerebral bridge. Front. Behav. Neurosci. 2017, 11, 8. [Google Scholar] [CrossRef]
- Varga, A.G.; Kathman, N.D.; Martin, J.P.; Guo, P.; Ritzmann, R.E. Spatial Navigation and the Central Complex: Sensory Acquisition, Orientation, and Motor Control. Front. Behav. Neurosci. 2017, 11, 4. [Google Scholar] [CrossRef]
- Daltorio, K.; Mirletz, B.; Bender, J.; Webster-Wood, V.; Szczecinski, N.; Branicky, M.; Ritzmann, R.; Quinn, R. A model of exploration and goal-searching in the cockroach, Blaberus discoidalis. Adapt. Behav. 2013, 21, 404–420. [Google Scholar] [CrossRef]
- Krause, T.; Spindler, L.; Poeck, B.; Strauss, R. Drosophila Acquires a Long-Lasting Body-Size Memory from Visual Feedback. Curr. Biol. 2019, 29, 1833–1841.e3. [Google Scholar] [CrossRef] [PubMed]
- Arena, P.; Patané, L.; Sanalitro, D.; Vitanza, A. Insect-Inspired Body Size Learning Model on a Humanoid Robot. In Proceedings of the 2018 7th IEEE International Conference on Biomedical Robotics and Biomechatronics (Biorob), Enschede, The Netherlands, 26–29 August 2018; pp. 1127–1132. [Google Scholar] [CrossRef]
- Arena, P.; Mauro, G.D.; Krause, T.; Patanè, L.; Strauss, R. A spiking network for body size learning inspired by the fruit fly. In Proceedings of the 2013 International Joint Conference on Neural Networks (IJCNN), Dallas, TX, USA, 4–9 August 2013; pp. 1–7. [Google Scholar] [CrossRef]
- Goulard, R.; Buehlmann, C.; Niven, J.E.; Graham, P.; Webb, B. A unified mechanism for innate and learned visual landmark guidance in the insect central complex. PLoS Comput. Biol. 2021, 15, 1473–1478. [Google Scholar] [CrossRef]
- Pick, S.; Strauss, R. Goal-Driven Behavioral Adaptations in Gap-Climbing Drosophila. Curr. Biol. 2005, 15, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Arena, E.; Arena, P.; Strauss, R.; Patanè, L. Motor-Skill Learning in an Insect Inspired Neuro-Computational Control System. Front. Neurorobot. 2017, 11, 12. [Google Scholar] [CrossRef]
- Plath, J.A.; Entler, B.V.; Kirkerud, N.H.; Schlegel, U.; Galizia, C.G.; Barron, A.B. Different roles for honey bee mushroom bodies and central complex in visual learning of colored lights in an aversive conditioning assay. Front. Behav. Neurosci. 2017, 11, 98. [Google Scholar] [CrossRef]
- Izhikevich, E.; Desai, N. Relating stdp to bcm. Neural Comput. 2003, 15, 1511–1523. [Google Scholar] [CrossRef]
- Fiore, V.G.; Kottler, B.; Gu, X.; Hirth, F. In silico Interrogation of Insect Central Complex Suggests Computational Roles for the Ellipsoid Body in Spatial Navigation. Front. Behav. Neurosci. 2017, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Zill, S.; Schmitz, J.; Büschges, A. Load sensing and control of posture and locomotion. Arthropod Struct. Dev. 2004, 33, 273–286. [Google Scholar] [CrossRef]
- Manoonpong, P.; Tetzlaff, C. Neural computation in embodied closed-loop systems for the generation of complex behavior: From biology to technology. Front. Neurorobot. 2018, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Ritzmann, R.E.; Quinn, R.D.; Fischer, M.S. Convergent evolution and locomotion through complex terrain by insects, vertebrates and robots. Arthropod Struct. Dev. 2004, 33, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Wystrach, A.; Schwarz, S. Ants use a predictive mechanism to compensate for passive displacements by wind. Curr. Biol. 2013, 23, R1083–R1085. [Google Scholar] [CrossRef][Green Version]
- Manoonpong, P.; Petersen, D.; Kovalev, A.; Wörgötter, F.; Gorb, S.N.; Spinner, M.; Heepe, L. Enhanced locomotion efficiency of a bio-inspired walking robot using contact surfaces with frictional anisotropy. Sci. Rep. 2016, 6, 1–11. [Google Scholar]
- Collins, L. The case for emulating insect brains using anatomical “wiring diagrams” equipped with biophysical models of neuronal activity. Biol. Cybern. 2019, 113, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Niven, J.E.; Chittka, L. Reuse of identified neurons in multiple neural circuits. Behav. Brain Sci. 2010, 33, 285. [Google Scholar] [CrossRef]
- Simons, M.; Tibbetts, E. Insects as models for studying the evolution of animal cognition. Curr. Opin. Insect Sci. 2019, 34, 117–122. [Google Scholar] [CrossRef] [PubMed]










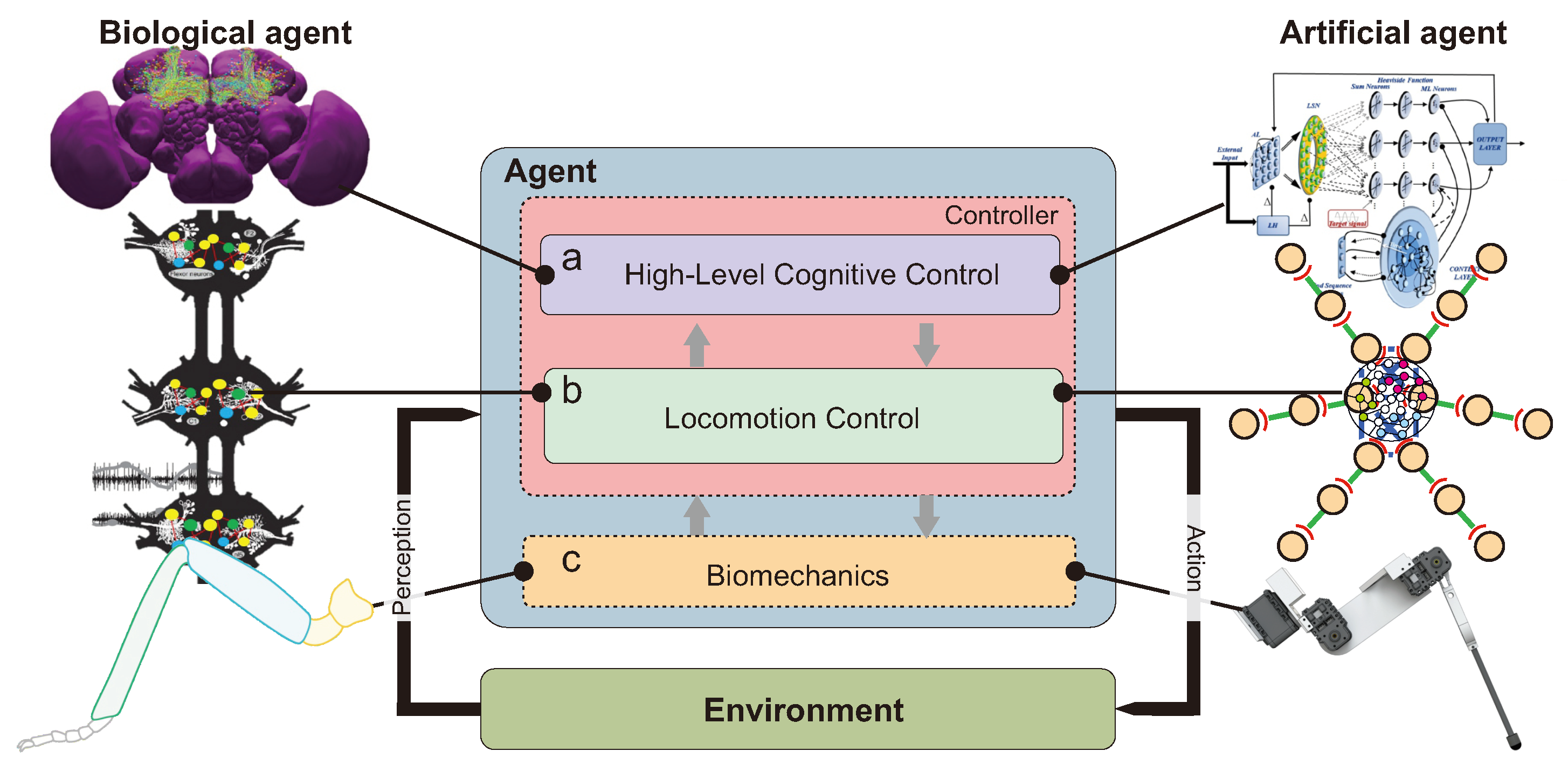{kind=link}
{kind=link}
{kind=link}
{kind=link}
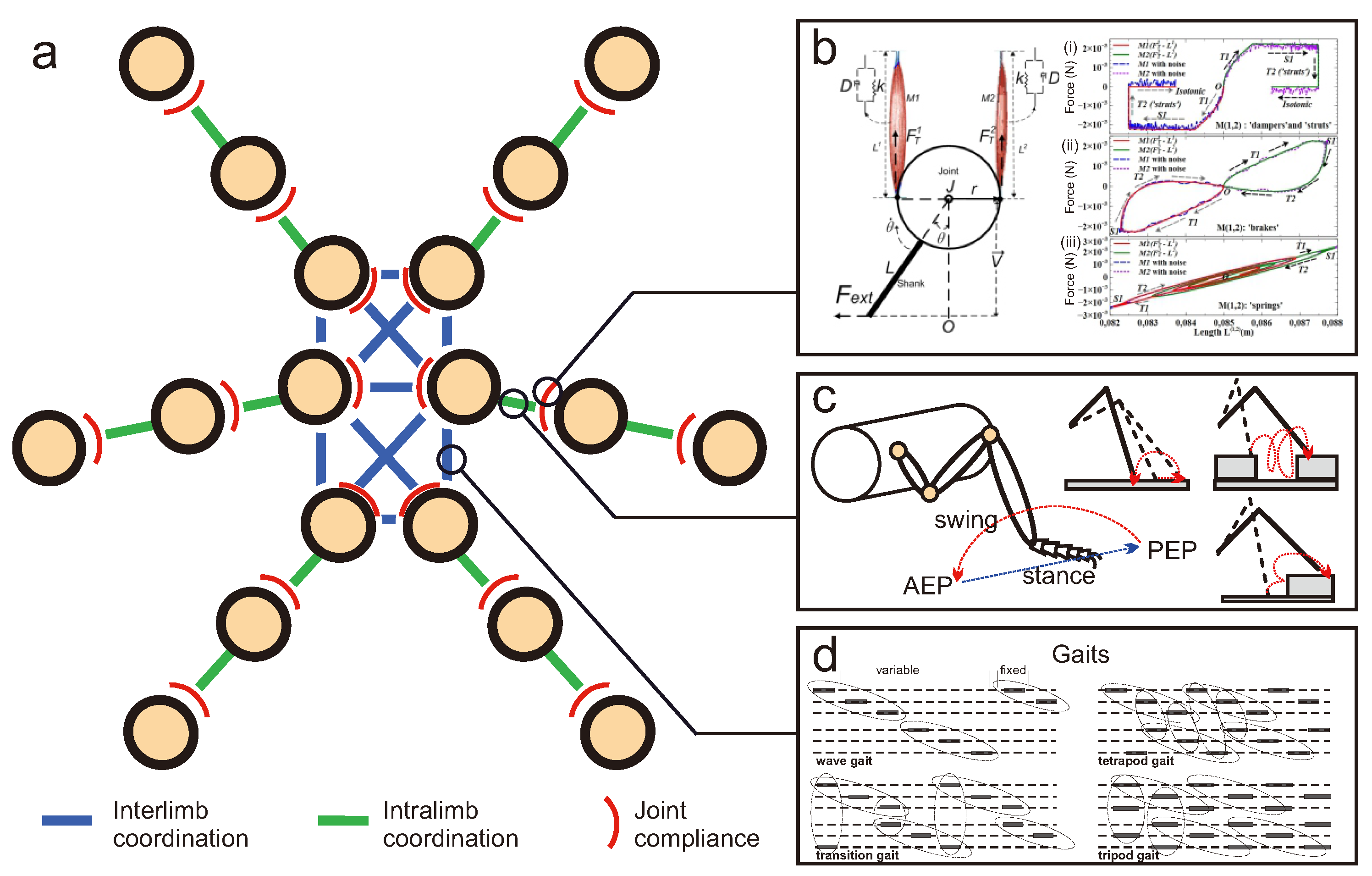{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Ref. | Hexapod | Size [m] | Mass [kg] | DOF | Compliant | Speed [m/s] | Task |
|---|---|---|---|---|---|---|---|---|
| 2021 | [14] | HAntR | 0.50 | 2.9 | 24 |  | 0.43 | Locomotion |
| 2019 | [15,16] | MORF | 0.60 | 4.2 | 18 |  | 0.70 | Locomotion |
| 2019 | [17] | Daisy | 1.10 | 21 | 18 | | 0.13 | Locomotion |
| 2019 | [18,19] | Drosophibot | 0.80 | 1 | 18 | | 0.05 | Locomotion |
| 2019 | [3,20] | AntBot | 0.45 | 2.3 | 18 | | 0.90 | Navigation |
| 2019 | [21] | Corin | 0.6 | 4.2 | 18 | | 0.10 | Locomotion |
| 2018 | [22] | AmphiHex-II | 0.51 | 14 | 6 | | 0.36 | Locomotion |
| 2018 | [23] | CRABOT | 0.70 | 2.5 | 24 | | 0.05 | Locomotion |
| 2017 | [24,25] | PhantomX AX | 0.50 | 2.6 | 18 | | 0.29 | Locomotion |
| 2017 | [20,24] | Hexabot | 0.36 | 0.68 | 18 | | 0.35 | Navigation |
| 2016 | [26] | Weaver | 0.35 | 7 | 30 | | 0.16 | Locomotion |
| 2016 | [27] | MX Phoenix | 0.80 | 4.8 | 18 | | 0.50 | Locomotion |
| 2015 | [28] | Phoenix 3DOF | 0.37 | 1.3 | 18 | | 0.25 | Locomotion |
| 2015 | [29] | HexaBull-1 | 0.53 | 3.4 | 18 | | - | Locomotion |
| 2015 | [30,31] | MantisBot | 0.74 | 6.1 | 28 | | - | Navigation |
| 2015 | [32] | Snake Monster | 0.70 | 4.6 | 18 | | - | Locomotion |
| 2015 | [33] | BionicANT | 0.15 | 0.105 | 18 | | - | Swarming |
| 2014 | [34,35,36] | HECTOR | 0.95 | 13 | 18 | | - | Navigation |
| 2014 | [37,38] | Messor II | 0.30 | 2.5 | 18 | | 0.09 | Locomotion |
| 2014 | [39,40] | LAURON V | 0.90 | 42 | 24 | | - | Navigation |
| 2014 | [41] | CREX | 1 | 27 | 24 | | 0.17 | Locomotion |
| 2012 | [42] | Octavio | 1 | 10.8 | 18 | | - | Locomotion |
| 2011 | [43] | - | 0.46 | 3 | 18 | | 0.03 | Navigation |
| 2011 | [44,45] | EduBot | 0.36 | 3.3 | 6 | | 2.50 | Locomotion |
| 2010 | [46] | X-RHex | 0.57 | 9.5 | 6 | | 1.54 | Locomotion |
| 2008 | [47] | DLR-crawler | 0.50 | 3.5 | 18 | | 0.20 | Locomotion |
| 2006 | [48,49] | AMOS-WD06 | 0.40 | 4.2 | 19 | | 0.07 | Locomotion |
| 2006 | [50,51] | Gregor I | 0.30 | 1.2 | 12 | | 0.03 | Locomotion |
| 2005 | [52,53] | BILL-Ant-a | 0.33 | 2.3 | 18 | | 0.03 | Locomotion |
| 2001 | [54] | RHex | 0.54 | 7 | 6 | | 0.55 | Locomotion |
| Robot | Actuators | #Actuators | Mass (kg) | Speed (m/s) | CoT |
|---|---|---|---|---|---|
| Daisy | X-series | 18 | 21 | 0.13 | 3.7 |
| X8-9 and X8-16 | |||||
| HAntR | Dynamixel AX-12A | 24 | 2.9 | 0.43 | 1.5 |
| AntBot | Dynamixel AX-18A | 18 | 2.3 | 0.90 | 6.2 |
| CRABOT | Dynamixel AX-18A | 24 | 2.5 | 0.05 | - |
| Hexabot | Dynamixel XL-320 | 18 | 0.93 | 0.35 | - |
| Weaver | Dynamixel | 30 | 7 | 0.16 | 1.5–1.8 |
| MX-64 and MX-106 | |||||
| EduBot | DC motor | 6 | 3.3 | 2.5 | 0.5–1.6 |
| Messor II | Dynamical RX-28 | 18 | 2.5 | 0.09 | - |
| BionicANT | Trimorphic piezo-ceramic | 18 | 0.105 | - | - |
| Insect Brain Areas | Functionalities | References |
|---|---|---|
| Mushroom bodies | Olfactory learning | [261], [262] |
| Attention | [263], [264] | |
| Expectation | [265], [266] | |
| Sameness | [234] [267] | |
| Sequence learning | [268] [269] | |
| Navigation and visual sequential memory | [260] | |
| Classification and decision making | [270], [271] | |
| Central complex | Navigation and detour paradigm | [272], [273], [274] |
| Goal-directed navigation | [275] [276] | |
| Spatial working memory in a water maze scenario | [235] | |
| Body-size model | [277], [35], [278], [279] | |
| Mixed | Navigation and landmark targeting behavior | [280], [192] |
| Motor-skill learning | [231], [281], [282] | |
| MBs and CX contribution to aversive visual learning | [283] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manoonpong, P.; Patanè, L.; Xiong, X.; Brodoline, I.; Dupeyroux, J.; Viollet, S.; Arena, P.; Serres, J.R. Insect-Inspired Robots: Bridging Biological and Artificial Systems. Sensors 2021, 21, 7609. https://doi.org/10.3390/s21227609
Manoonpong P, Patanè L, Xiong X, Brodoline I, Dupeyroux J, Viollet S, Arena P, Serres JR. Insect-Inspired Robots: Bridging Biological and Artificial Systems. Sensors. 2021; 21(22):7609. https://doi.org/10.3390/s21227609
Chicago/Turabian StyleManoonpong, Poramate, Luca Patanè, Xiaofeng Xiong, Ilya Brodoline, Julien Dupeyroux, Stéphane Viollet, Paolo Arena, and Julien R. Serres. 2021. "Insect-Inspired Robots: Bridging Biological and Artificial Systems" Sensors 21, no. 22: 7609. https://doi.org/10.3390/s21227609
APA StyleManoonpong, P., Patanè, L., Xiong, X., Brodoline, I., Dupeyroux, J., Viollet, S., Arena, P., & Serres, J. R. (2021). Insect-Inspired Robots: Bridging Biological and Artificial Systems. Sensors, 21(22), 7609. https://doi.org/10.3390/s21227609