
In Situ Real-Time Monitoring of Glutamate and Electrophysiology from Cortex to Hippocampus in Mice Based on a Microelectrode Array

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
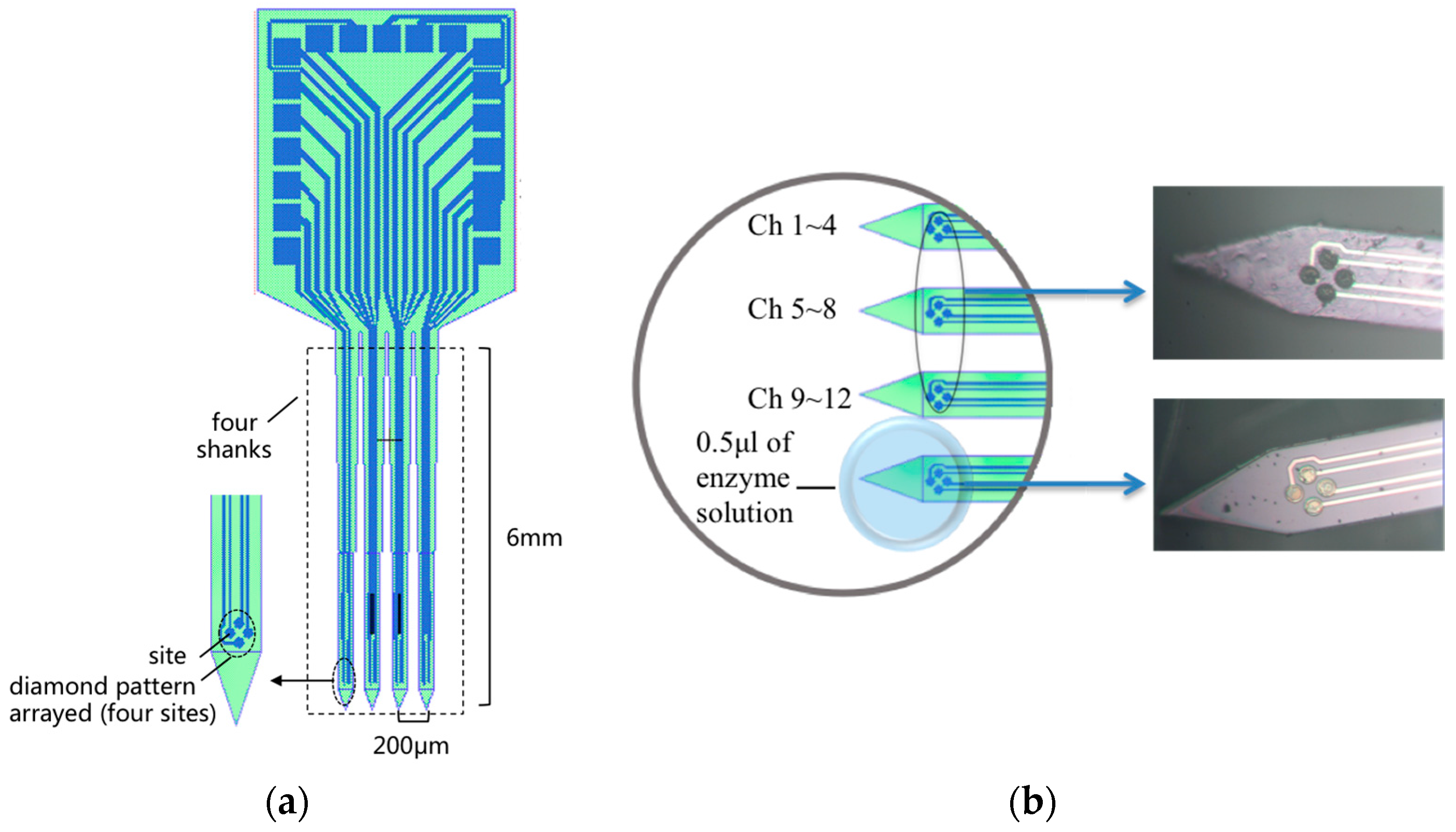
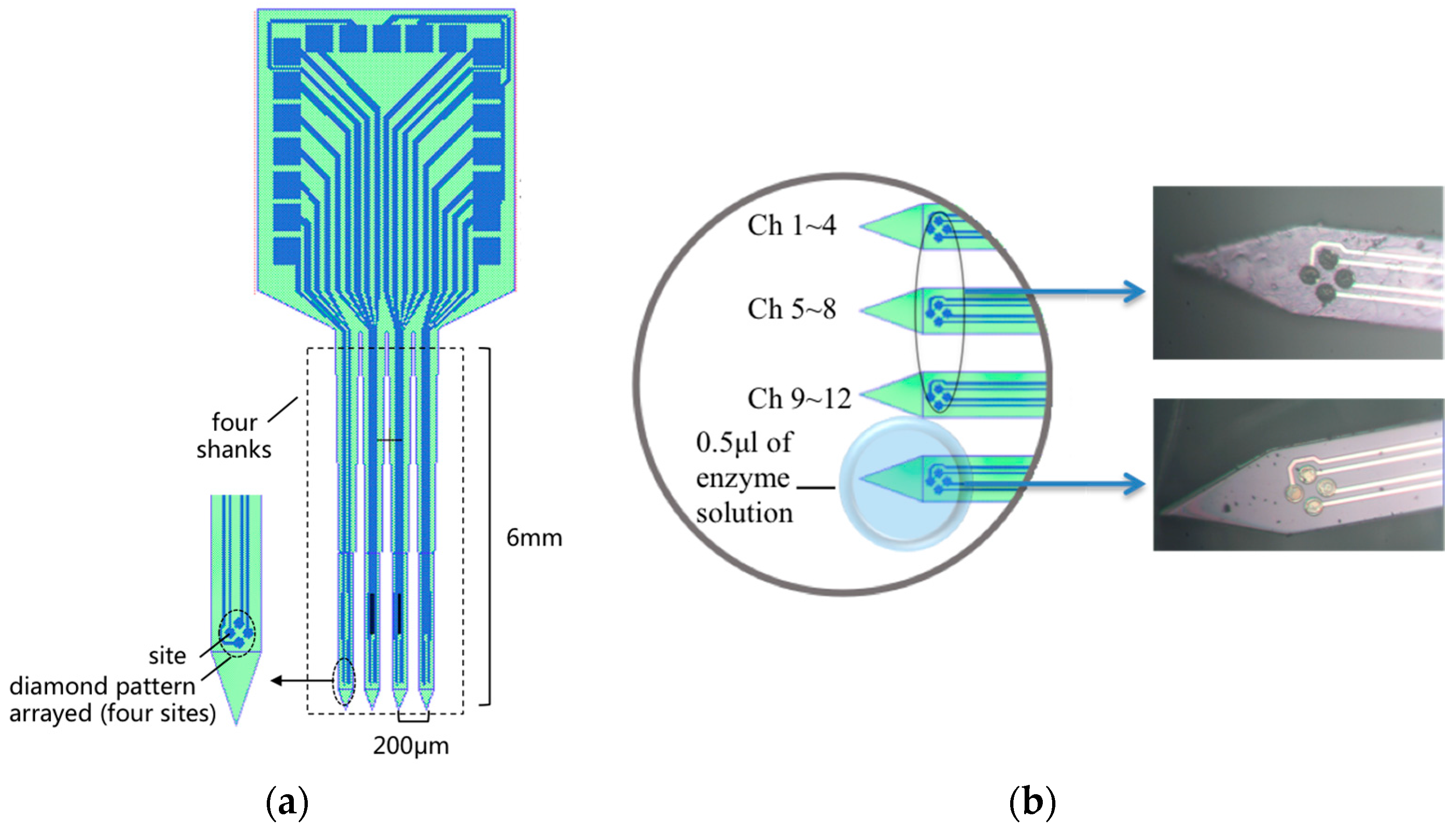
2.1. MEA Probe Fabrication and Preparation
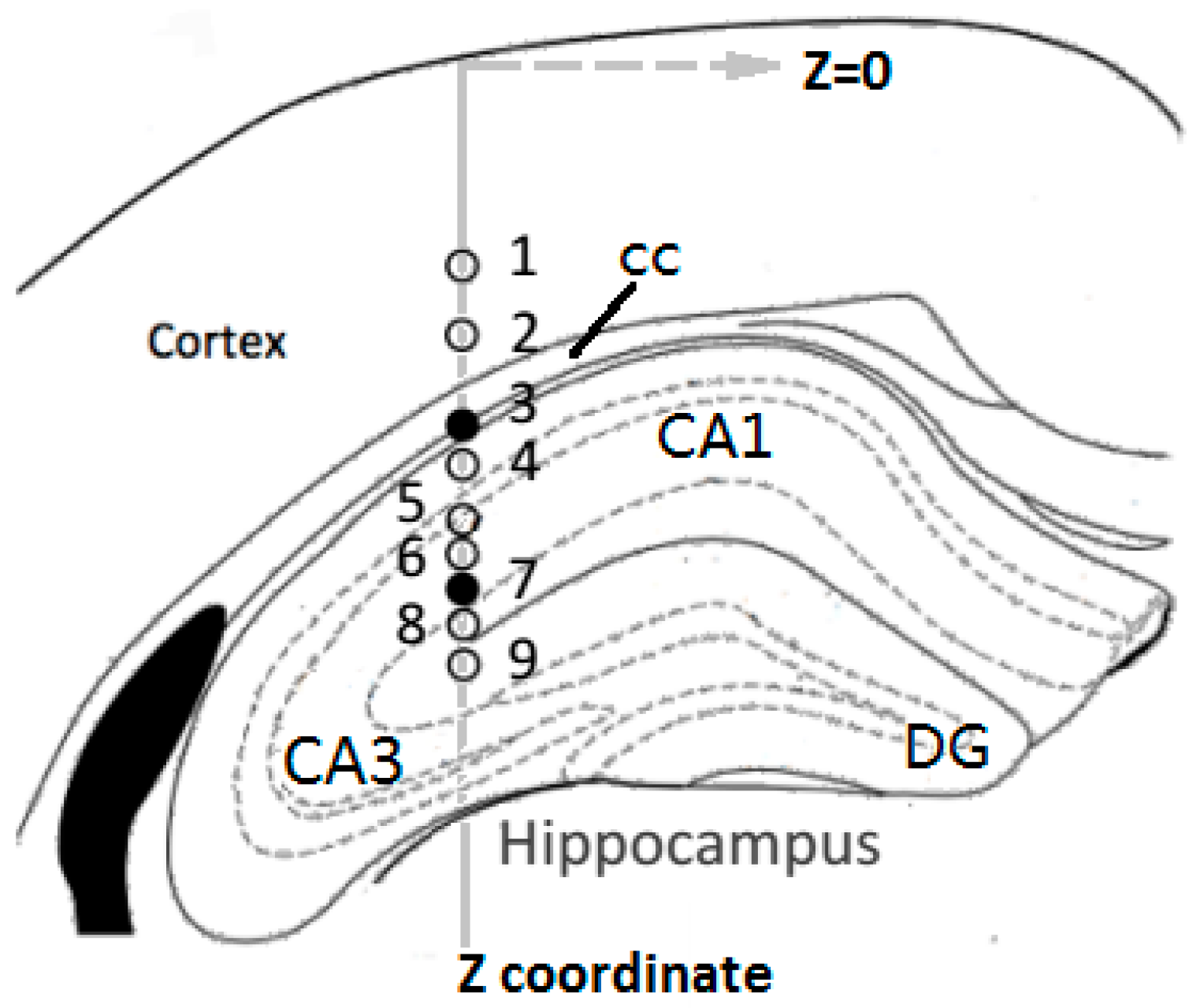
2.2. In Vivo Experiments
3. Results and Discussion
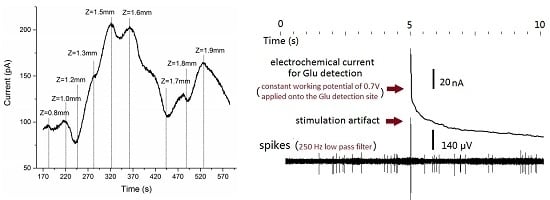
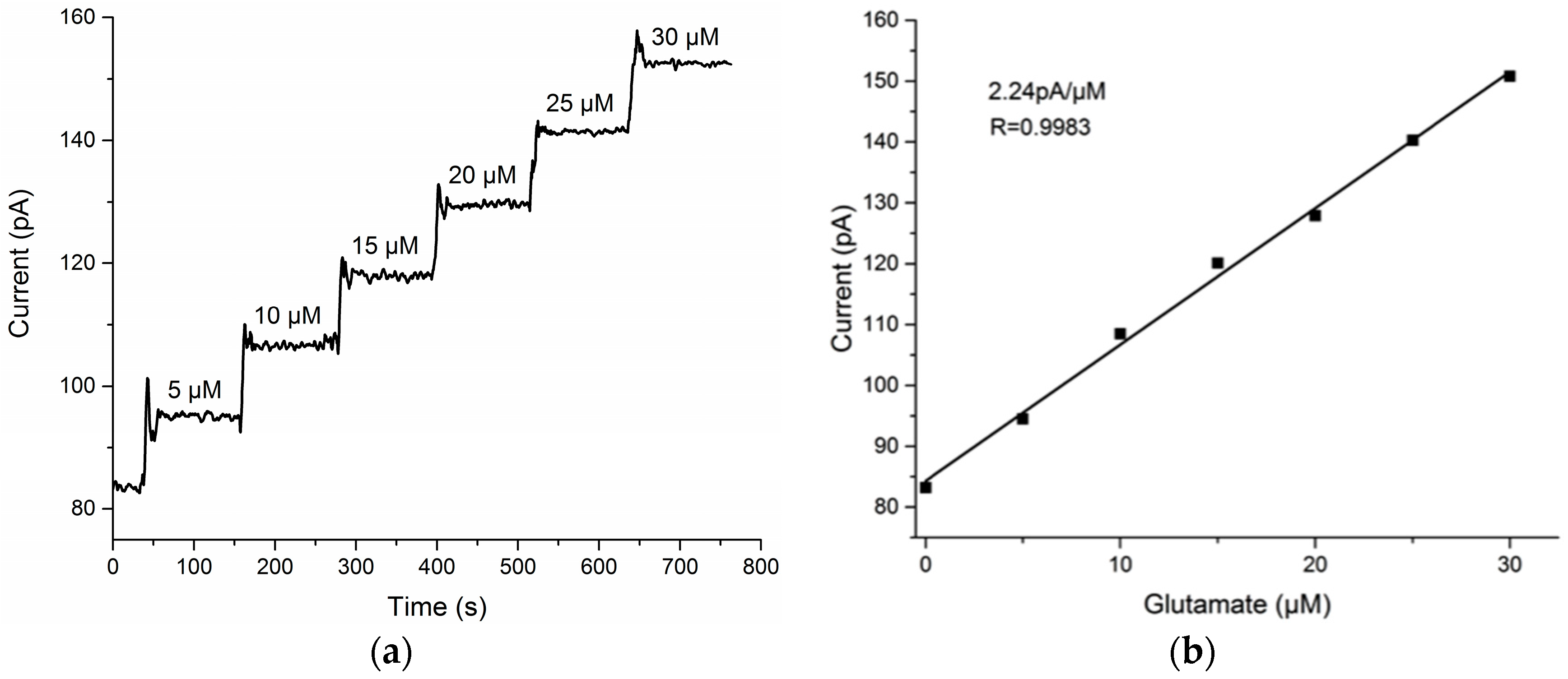
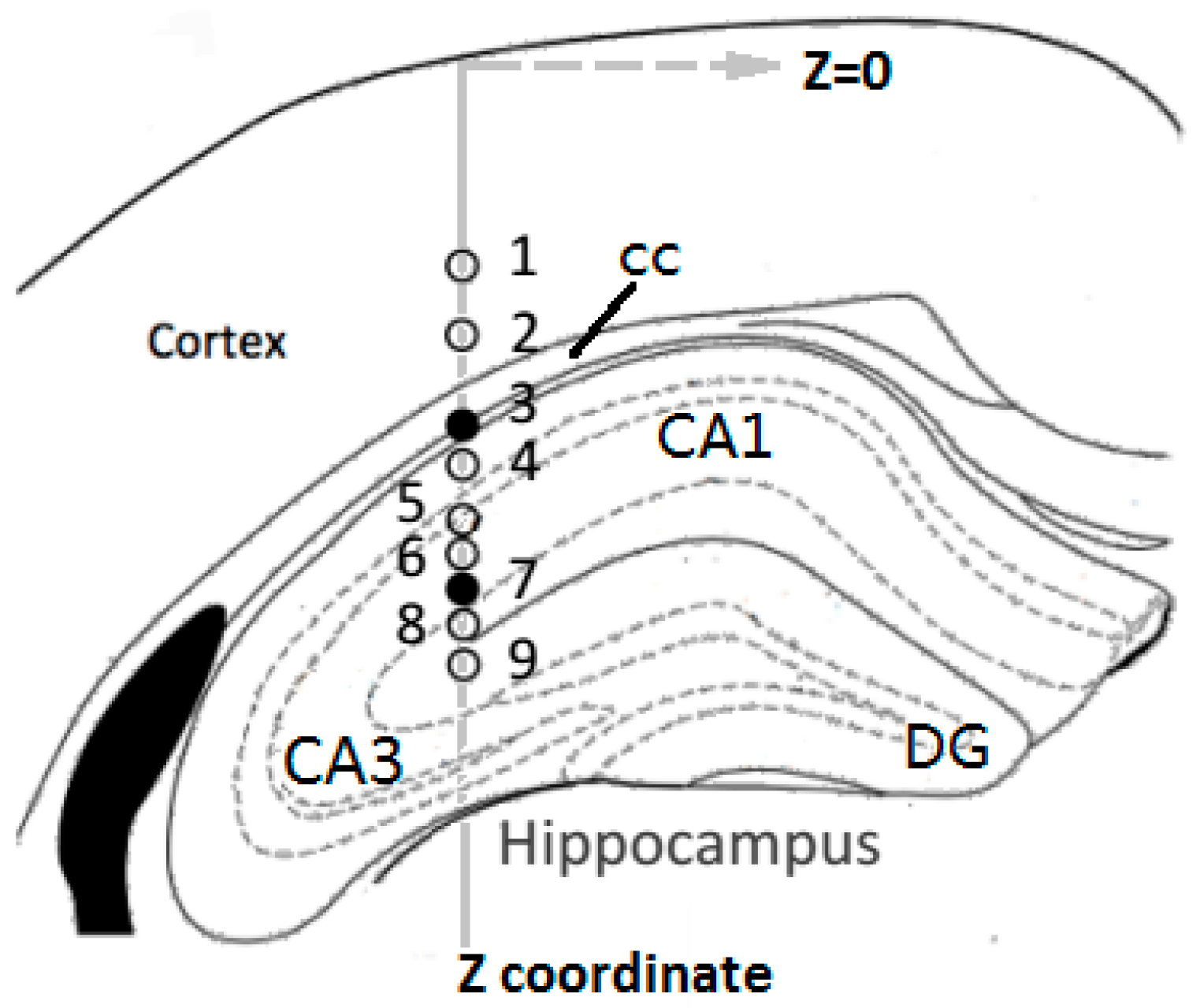
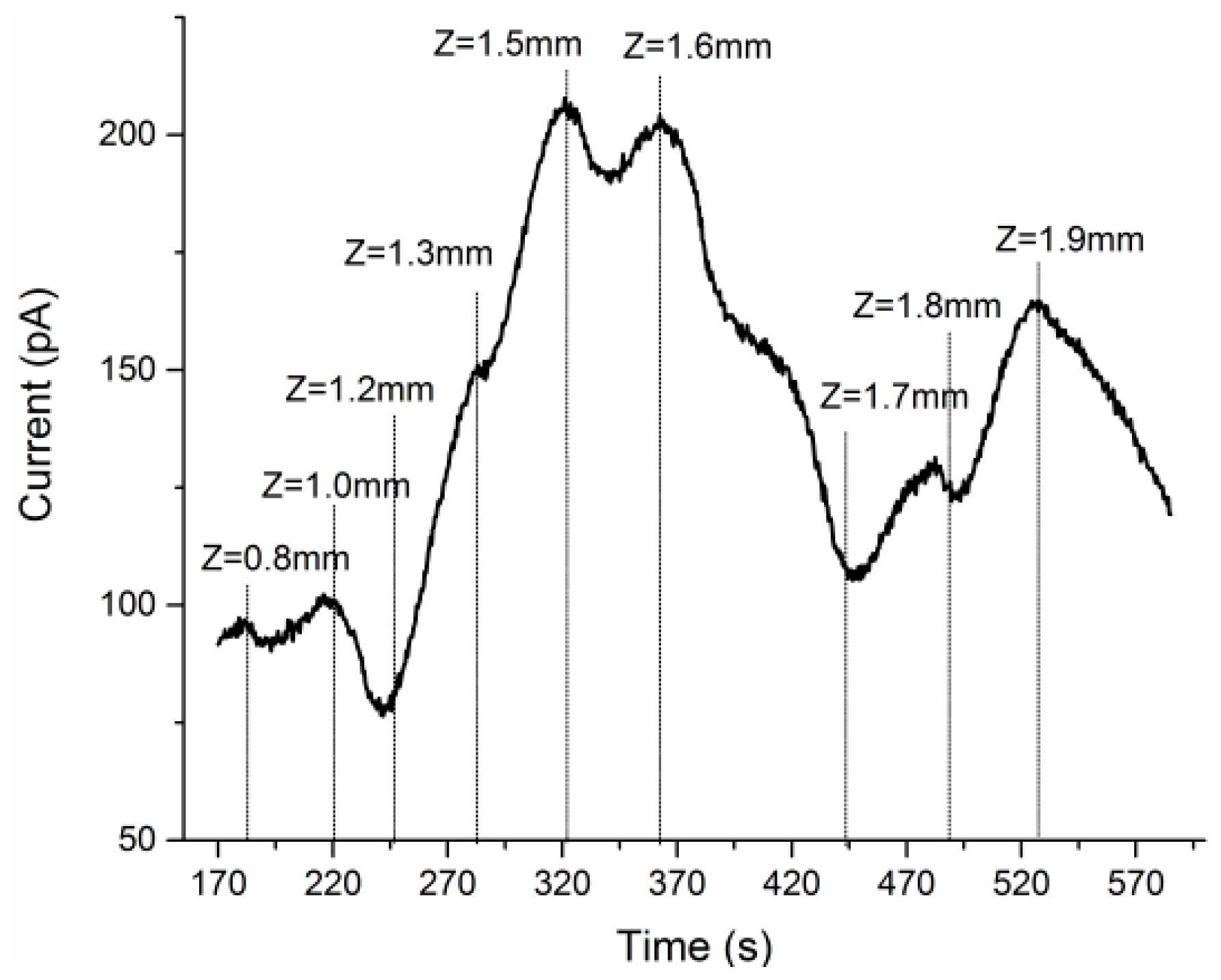
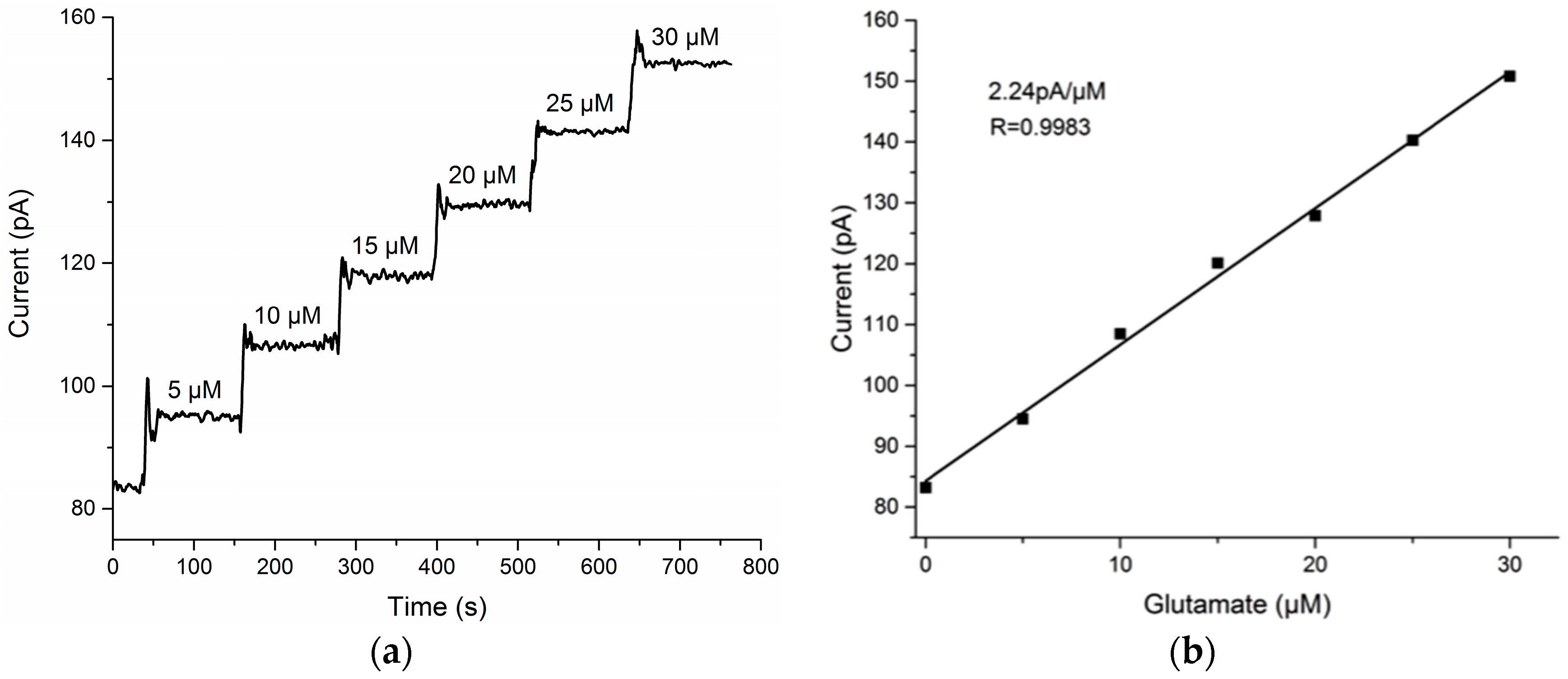
3.1. Glutamate Recordings from Cortex to Hippocampus
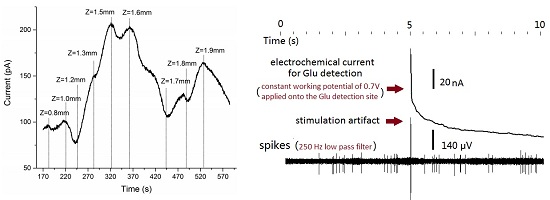
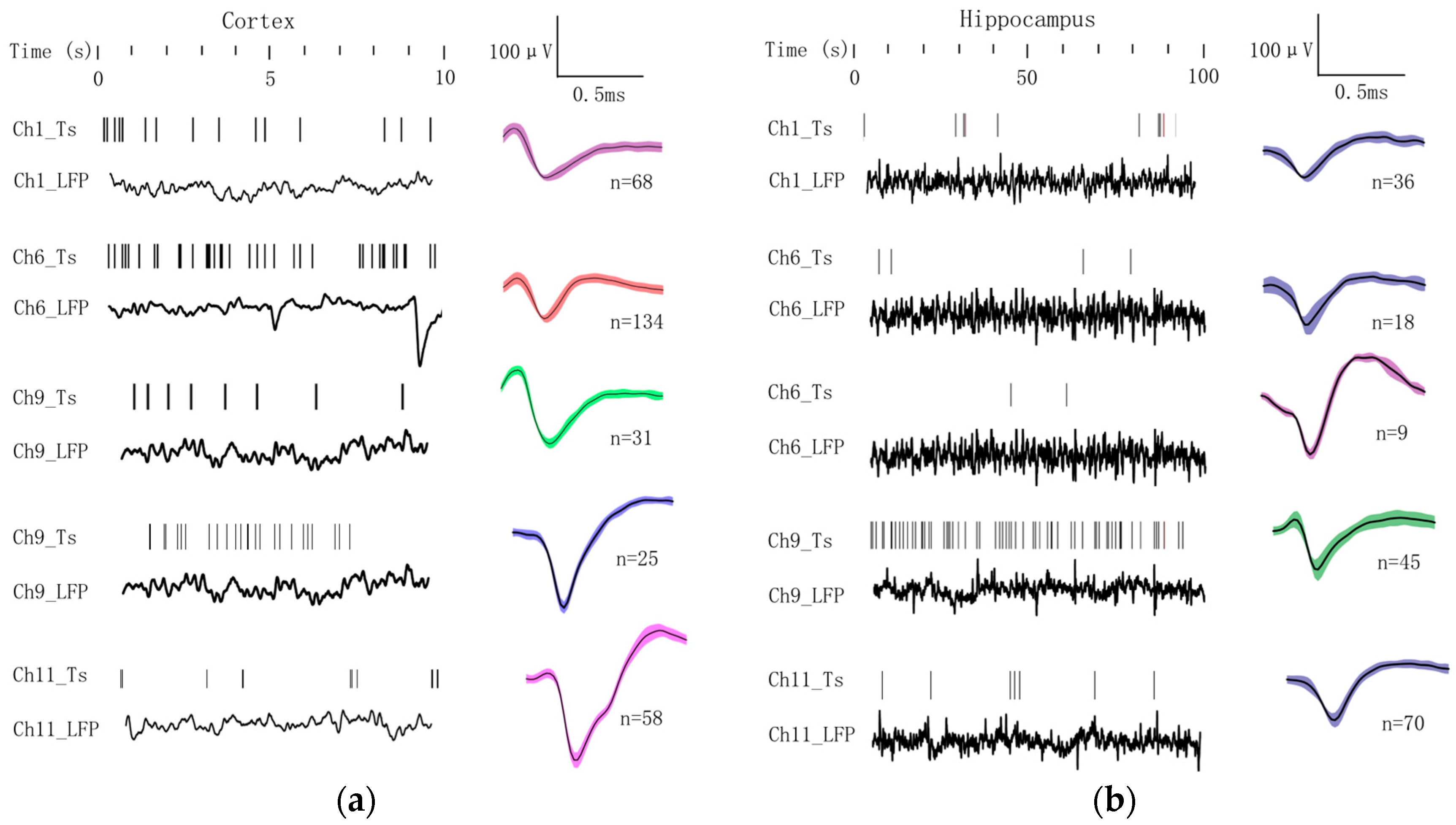
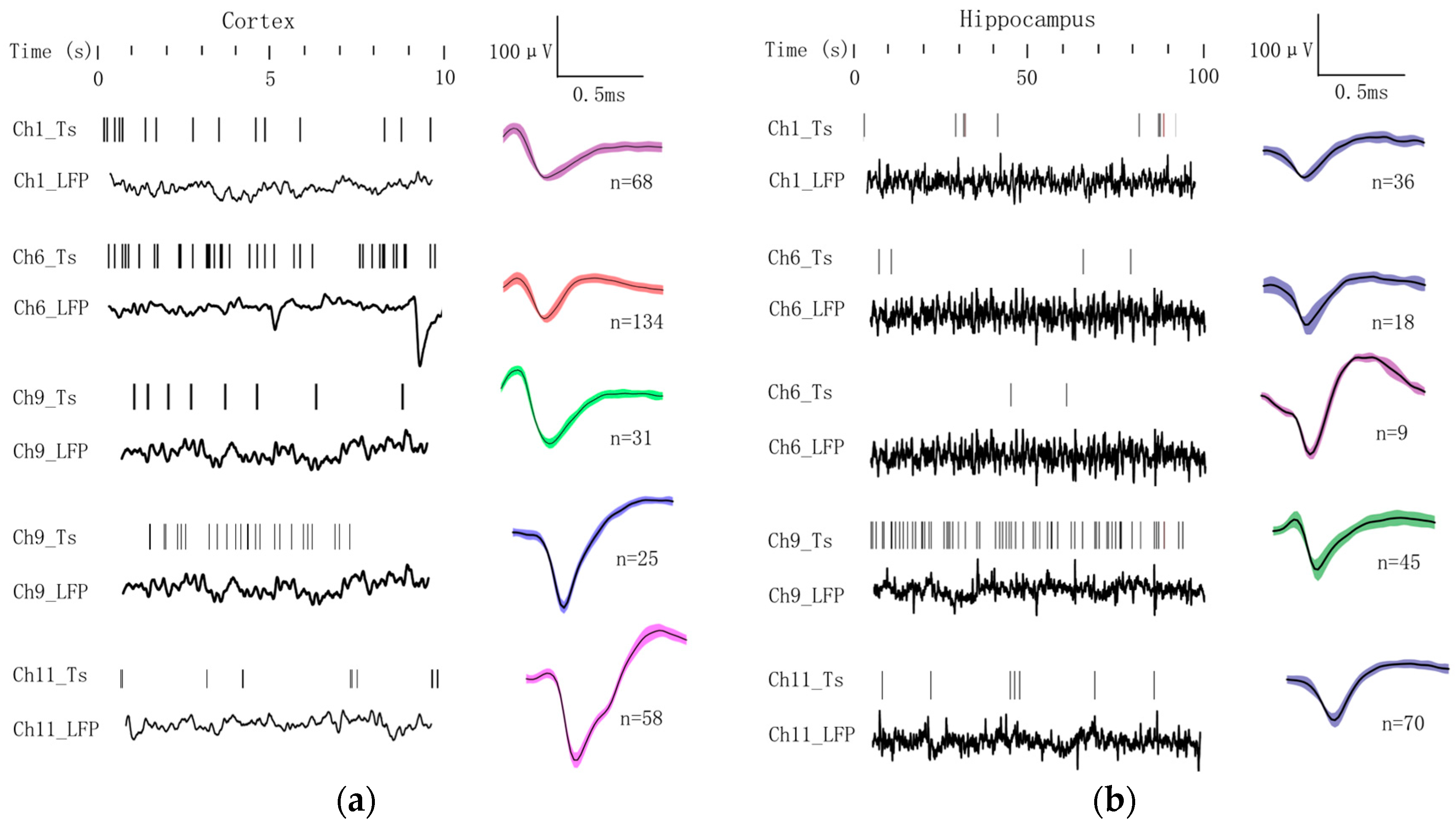
3.2. Electrophysiological Recordings from Cortex to Hippocampus
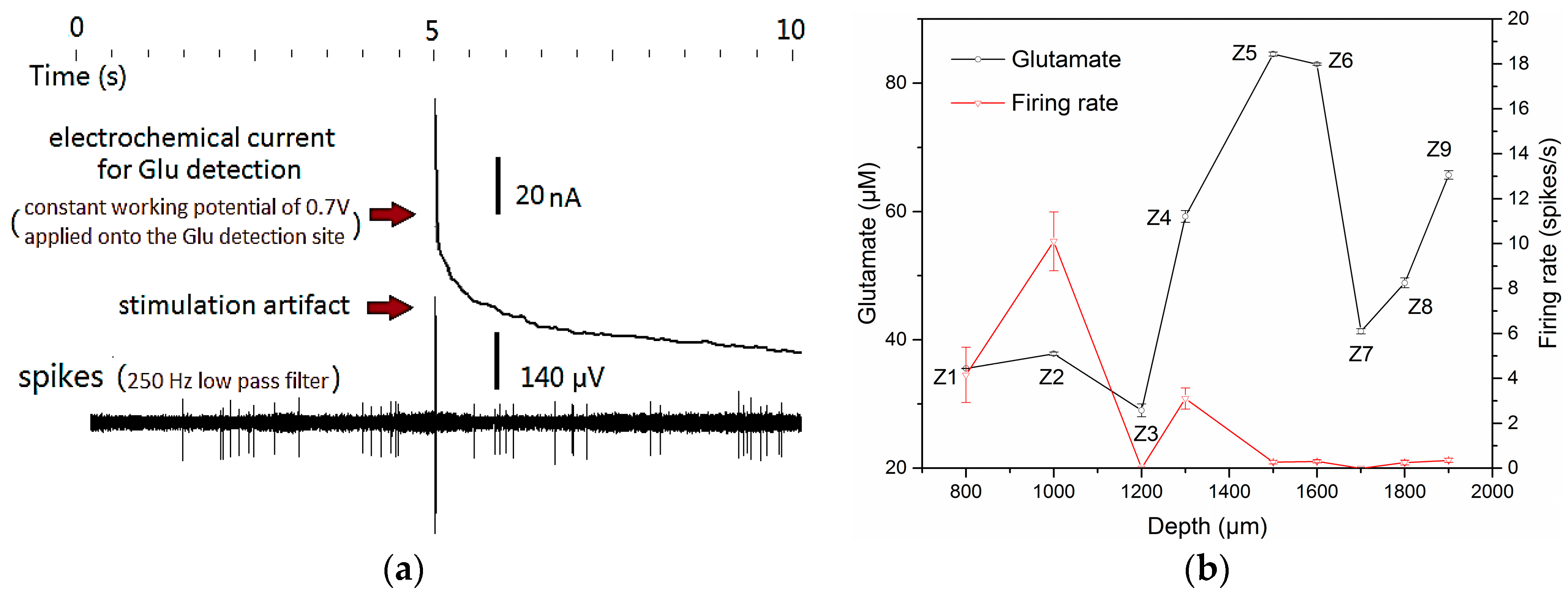
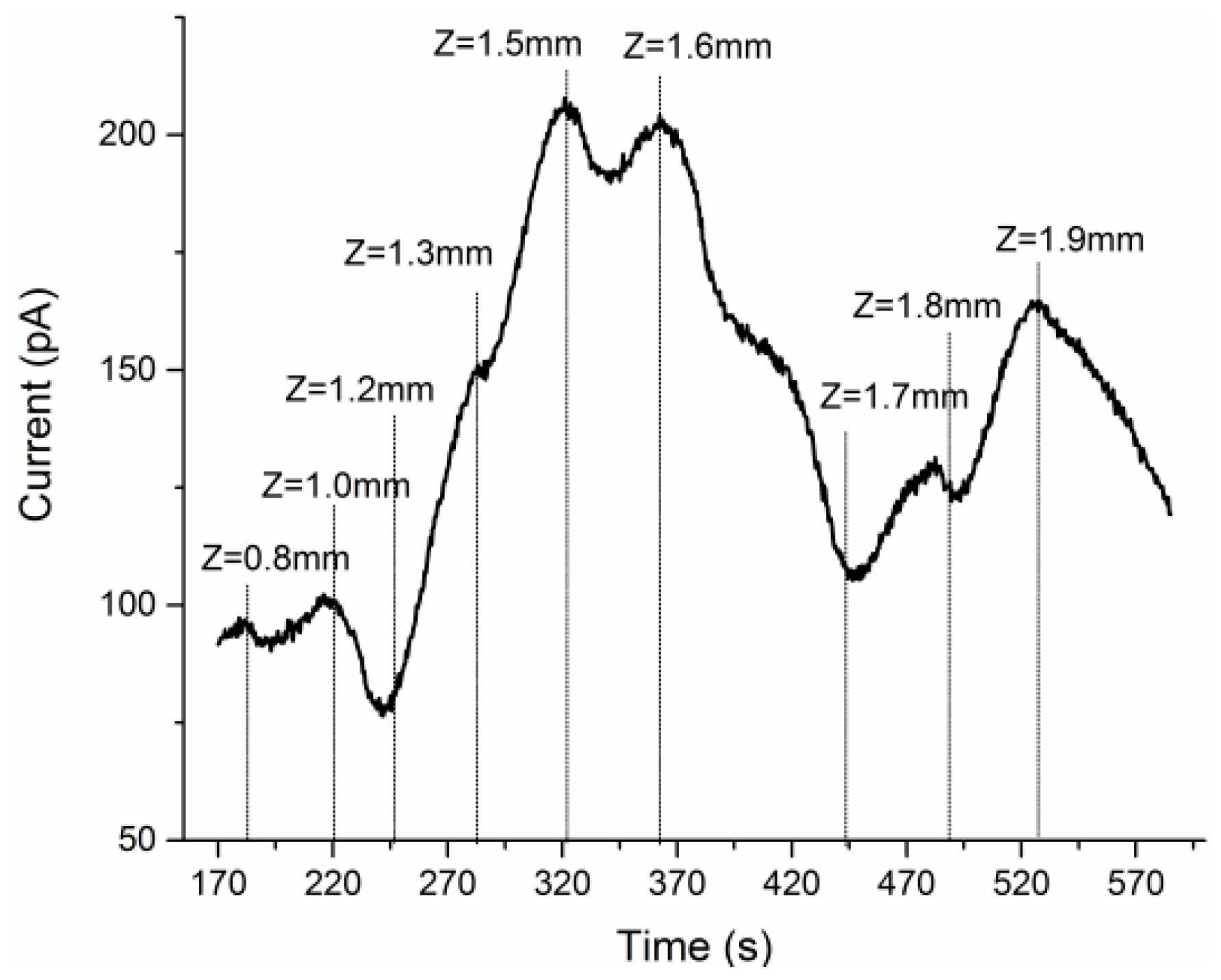
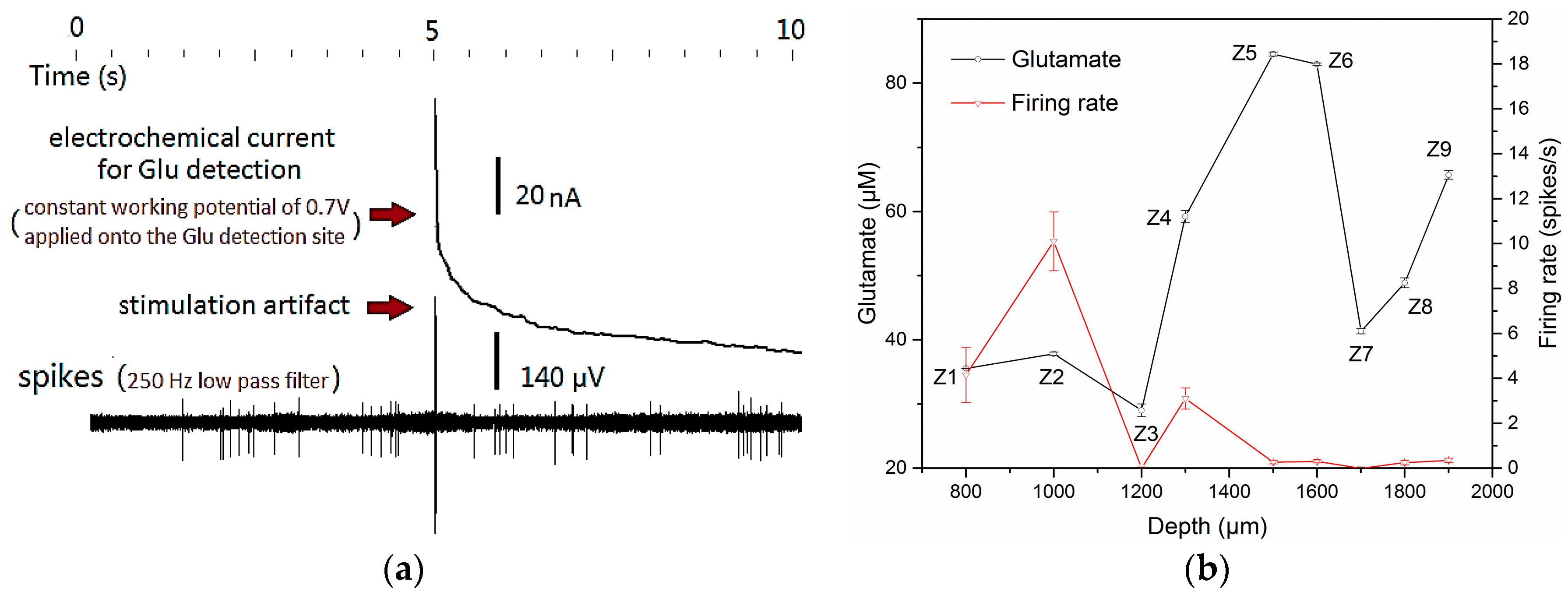
3.3. Concurrent Analysis of Glutamate Distribution and Spike Firing Rate Changes from Cortex to Hippocampus
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hascup, K.N.; Hascup, E.R.; Pomerleau, F.; Huettl, P.; Gerhardt, G.A. Second-by-second measures of L-glutamate in the prefrontal cortex and striatum of freely moving mice. J. Pharmacol. Exp. Ther. 2008, 324, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Lin, N.; Liu, C.; Jiang, H.; Xing, G.; Cai, X. A novel dual mode microelectrode array for neuroelectrical and neurochemical recording in vitro. Biosens. Bioelectron. 2012, 38, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Tamminga, C.A.; Stan, A.D.; Wagner, A.D. The hippocampal formation in schizophrenia. Am. J. Psychiatry 2010, 167, 1178–1193. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.; Bliss, T.V.P.; Skrede, K.K. Unit analysis of hippocampal population spikes. Exp. Brain Res. 1971, 13, 208–221. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, K. Disinhibition reduces extracellular glutamine and elevates extracellular glutamate in rat hippocampus in vivo. Epilepsy Res. 2015, 114, 32–46. [Google Scholar] [CrossRef] [PubMed]
- El-Khoury, R.; Panayotis, N.; Matagne, V.; Ghata, A.; Villard, L.; Roux, J.C. GABA and glutamate pathways are spatially and developmentally affected in the brain of Mecp2-deficient mice. PLoS ONE 2014, 9, e92169. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, L.; Ballini, C.; Colivicchi, M. Investigation on acetylcholine, aspartate, glutamate and gaba extracellular levels from ventral hippocampus during repeated exploratory activity in the rat. Neurochem. Res. 2003, 28, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Mizuseki, K.; Royer, S.; Diba, K. Activity dynamics and be havioral correlates of CA3 and CA1 hippocampal pyramidal neurons. Hippocampus 2012, 22, 1659–1680. [Google Scholar] [CrossRef] [PubMed]
- Blanche, T.; Spacek, M.; Hetke, J. Polytrodes: High-density silicon electrode arrays for large-scale multiunit recording. J. Neurophysiol. 2005, 93, 2987–3000. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.L.; Pomerleau, F.; Huettl, P.; Gerhardt, G.A.; Zhang, Z. Real-time glutamate measurements in the putamen of awake rhesus monkeys using an enzyme-based human microelectrode array prototype. J. Neurosci. Methods 2010, 185, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.D.; Franklin, R.K.; Gibson, M.D.; Brown, R.B.; Kipke, D.R. Implantable microelectrode arrays for simultaneous electrophysiological and neurochemical recordings. J. Neurosci. Methods 2008, 174, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Jason, J.; Karen, M.; Greg, G. Ceramic-Based Multisite Microelectrodes for Electrochemical Recordings. Anal. Chem. 2000, 72, 187–192. [Google Scholar]
- Jason, J.; Greg, G. Self-Referencing Ceramic-Based Multisite Microelectrodes for the Detection and Elimination of Interferences from the Measurement of L-Glutamate and Other Analytes. Anal. Chem. 2001, 73, 1037–1042. [Google Scholar]
- Wei, W.; Song, Y.; Wang, L.; Zhang, S.; Luo, J.; Xu, S.; Cai, X. An implantable microelectrode array for simultaneous L-glutamate and electrophysiological recordings in vivo. Microsyst. Nanoeng. 2015, 1, 15002. [Google Scholar] [CrossRef]
- Wei, W.J.; Song, Y.L.; Shi, W.T.; Liu, C.X.; Jiang, T.J.; Cai, X.X. A novel microelectrode array probe integrated with electrophysiology reference electrode for neural recording. Key Eng. Mater. 2013, 562, 67–73. [Google Scholar] [CrossRef]
- Wei, W.; Song, Y.; Fan, X.; Zhang, S.; Wang, L.; Xu, S.; Wang, M.; Cai, X. Simultaneous recording of brain extracellular glucose, spike and local field potential in real time using an implantable microelectrode array with nano-materials. Nanotechnology 2016, 27, 114001. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Song, Y.; Wang, M.; Zhang, Z.; Fan, X.; Song, X.; Zhuang, P.; Yue, F.; Chan, P.; Cai, X. A silicon based implantable microelectrode array for electrophysiological and dopamine recording from cortex to striatum in the non-human primate brain. Biosen. Bioelectron. 2016, 85, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Mizuseki, K.; Diba, K.; Pastalkova, E. Hippocampal CA1 pyramidal cells form functionally distinct sublayers. Nat. Neurosci. 2011, 14, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Arnthjensen, N.; Jabaudon, D.; Scanziani, M. Cooperation between independent hippocampal synapses is controlled by glutamate uptake. Nat. Neurosci. 2002, 5, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Fedele, E.; Mazzone, P.; Stefani, A.; Bassi, A.; Ansaldo, M.; Raiteri, M. Microdialysis in parkinsonian patient basal ganglia: Acute apomorphine-induced clinical and electrophysiological effects not paralleled by changes in the release of neuroactive amino acids. Exp. Neurol. 2001, 167, 356–365. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Song, Y.; Ma, Y.; Zhang, S.; Xiao, G.; Yang, L.; Xu, H.; Zhang, D.; Cai, X. In Situ Real-Time Monitoring of Glutamate and Electrophysiology from Cortex to Hippocampus in Mice Based on a Microelectrode Array. Sensors 2017, 17, 61. https://doi.org/10.3390/s17010061
Fan X, Song Y, Ma Y, Zhang S, Xiao G, Yang L, Xu H, Zhang D, Cai X. In Situ Real-Time Monitoring of Glutamate and Electrophysiology from Cortex to Hippocampus in Mice Based on a Microelectrode Array. Sensors. 2017; 17(1):61. https://doi.org/10.3390/s17010061
Chicago/Turabian StyleFan, Xinyi, Yilin Song, Yuanlin Ma, Song Zhang, Guihua Xiao, Lili Yang, Huiren Xu, Dai Zhang, and Xinxia Cai. 2017. "In Situ Real-Time Monitoring of Glutamate and Electrophysiology from Cortex to Hippocampus in Mice Based on a Microelectrode Array" Sensors 17, no. 1: 61. https://doi.org/10.3390/s17010061
APA StyleFan, X., Song, Y., Ma, Y., Zhang, S., Xiao, G., Yang, L., Xu, H., Zhang, D., & Cai, X. (2017). In Situ Real-Time Monitoring of Glutamate and Electrophysiology from Cortex to Hippocampus in Mice Based on a Microelectrode Array. Sensors, 17(1), 61. https://doi.org/10.3390/s17010061