The Amazonian Formative: Crop Domestication and Anthropogenic Soils

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
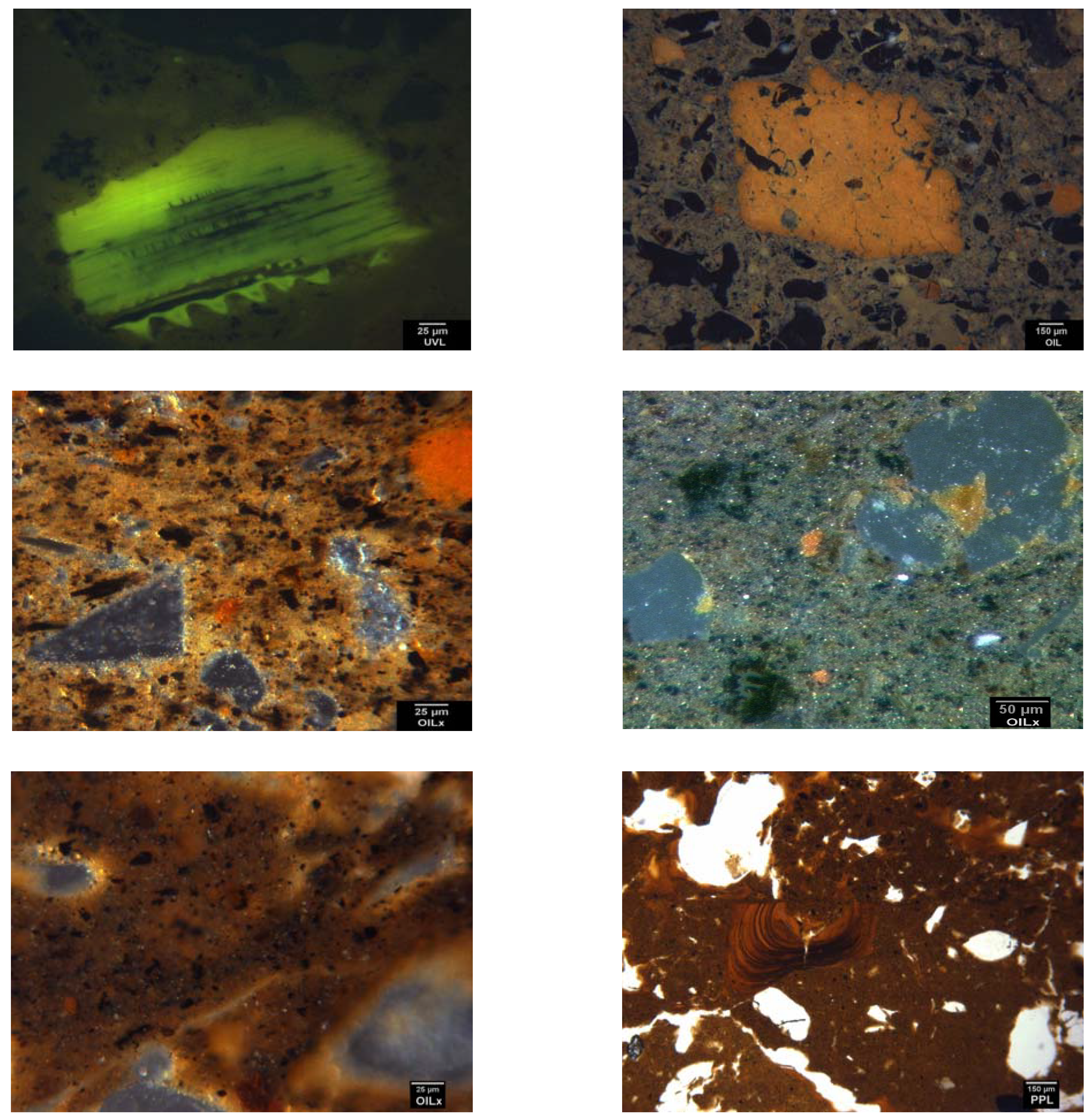{kind=link}
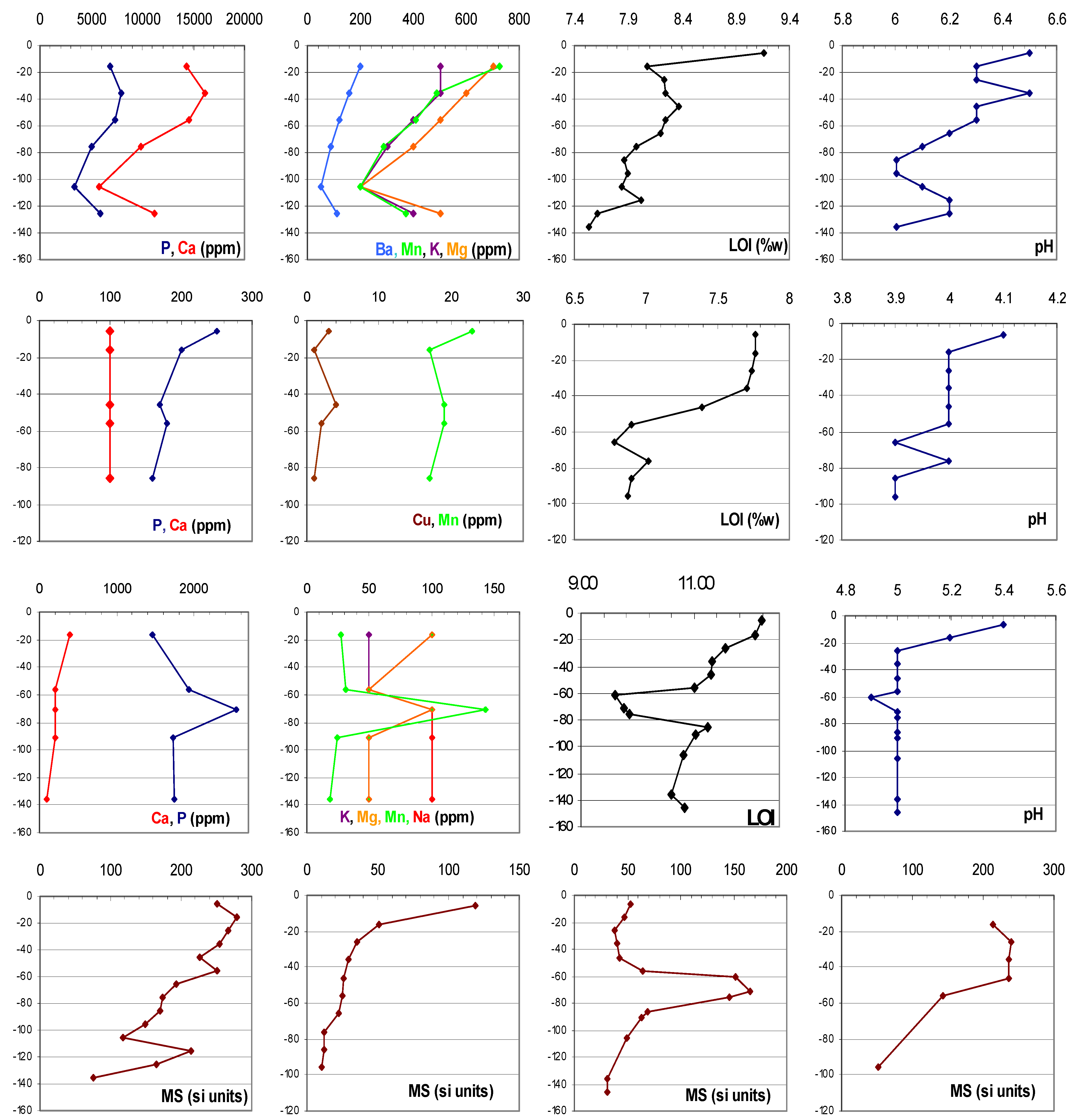{kind=link}
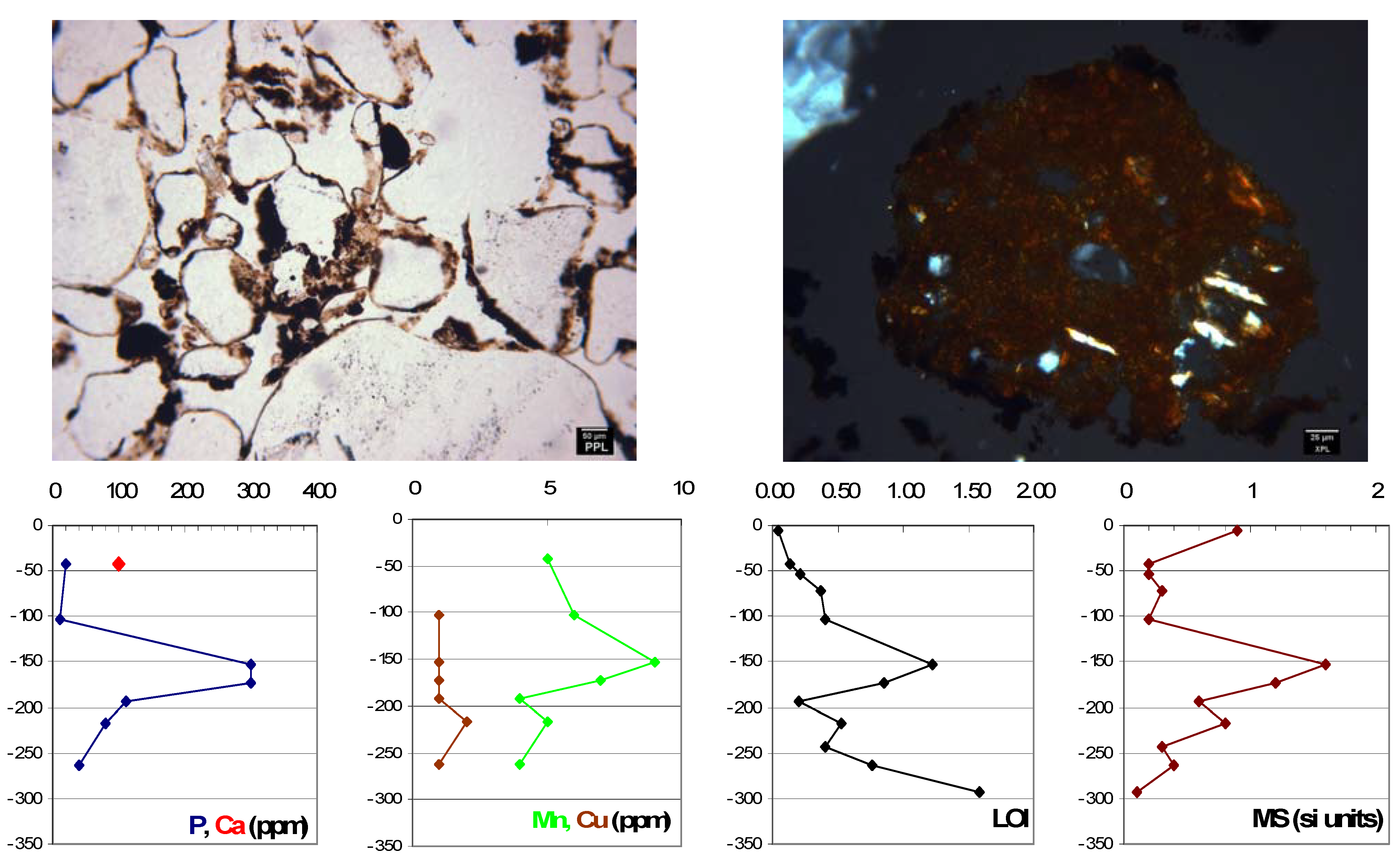{kind=link}
{kind=link}
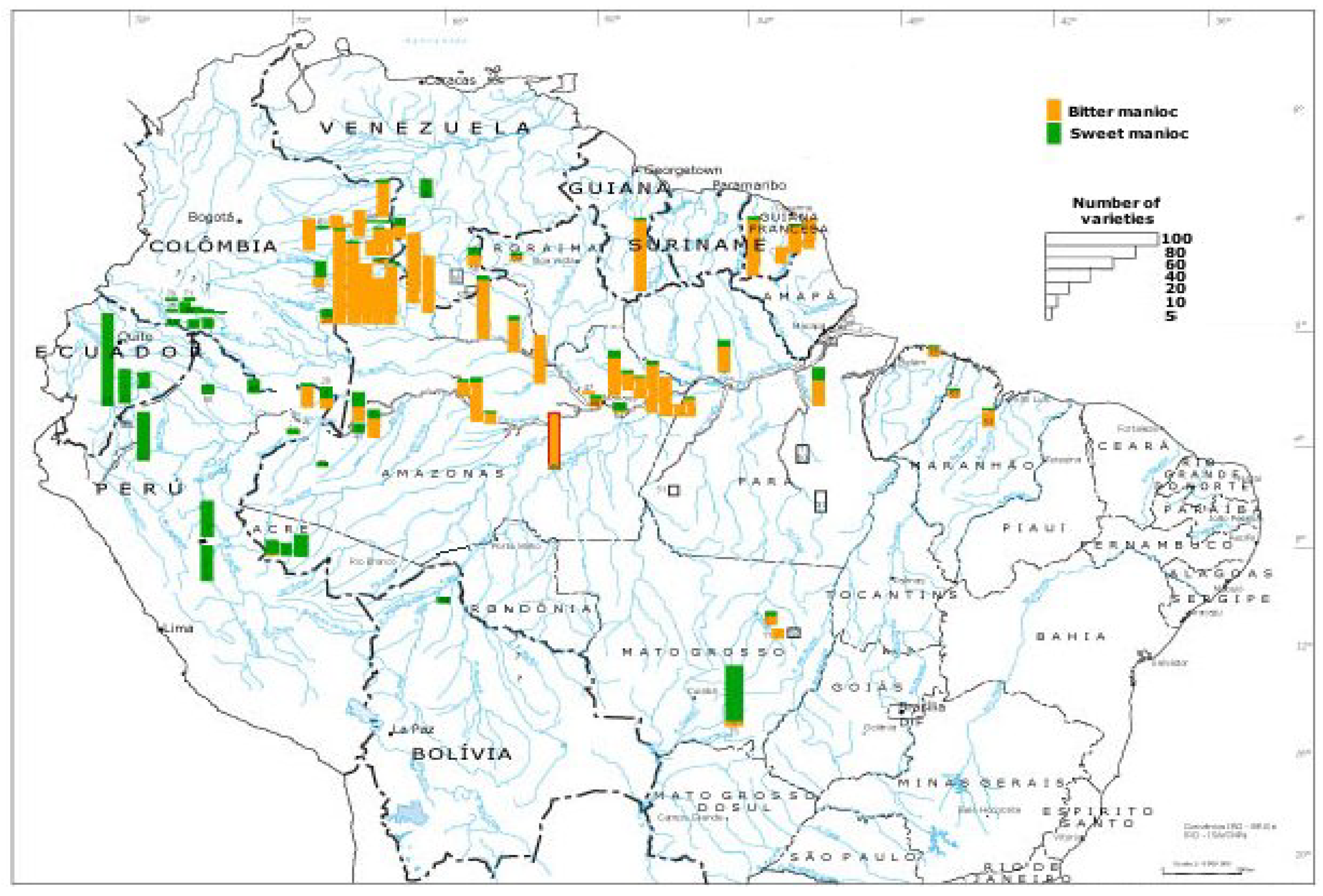{kind=link}
1. Introduction
2. Sedentism and Amazonian Anthropogenic Soils
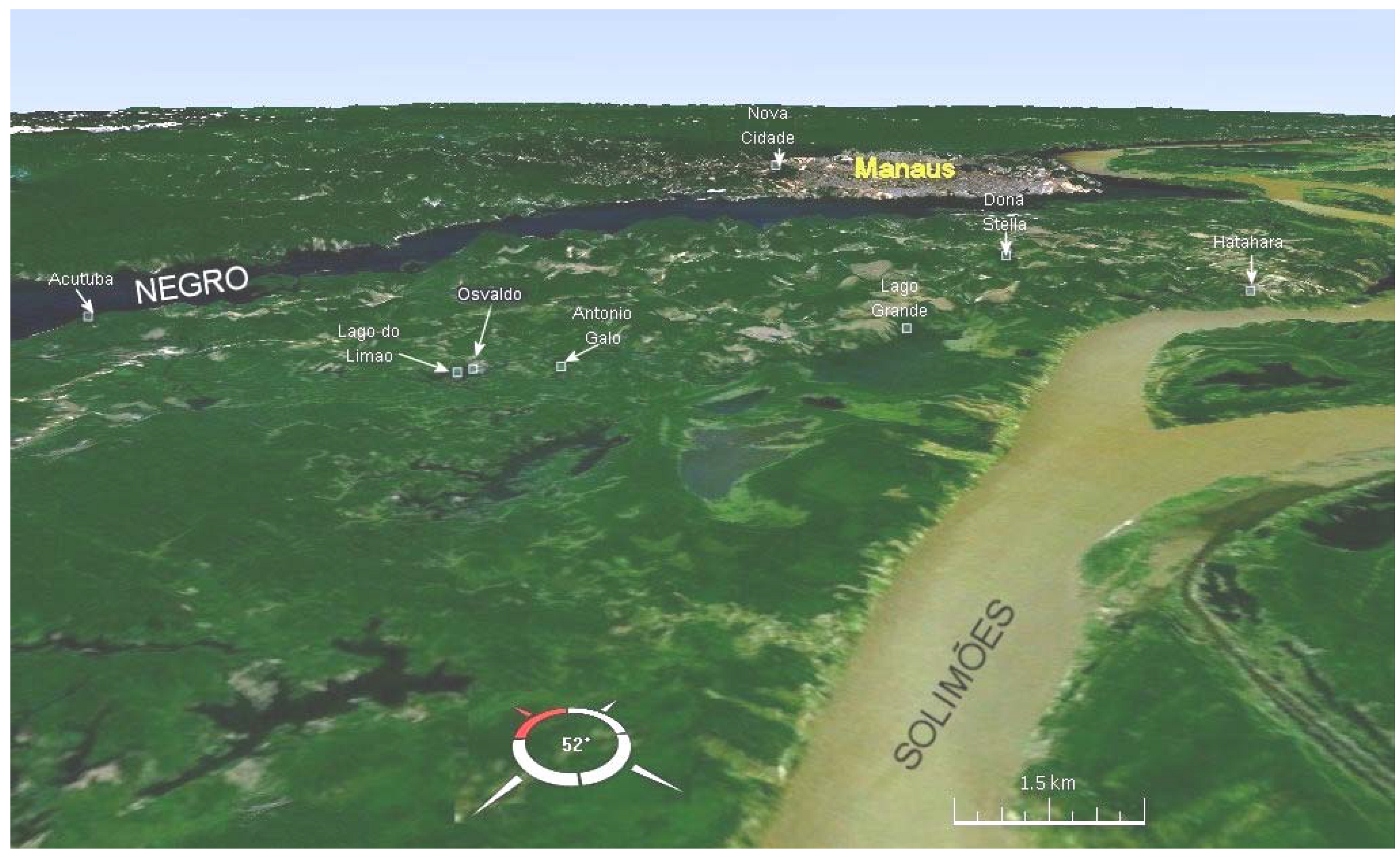
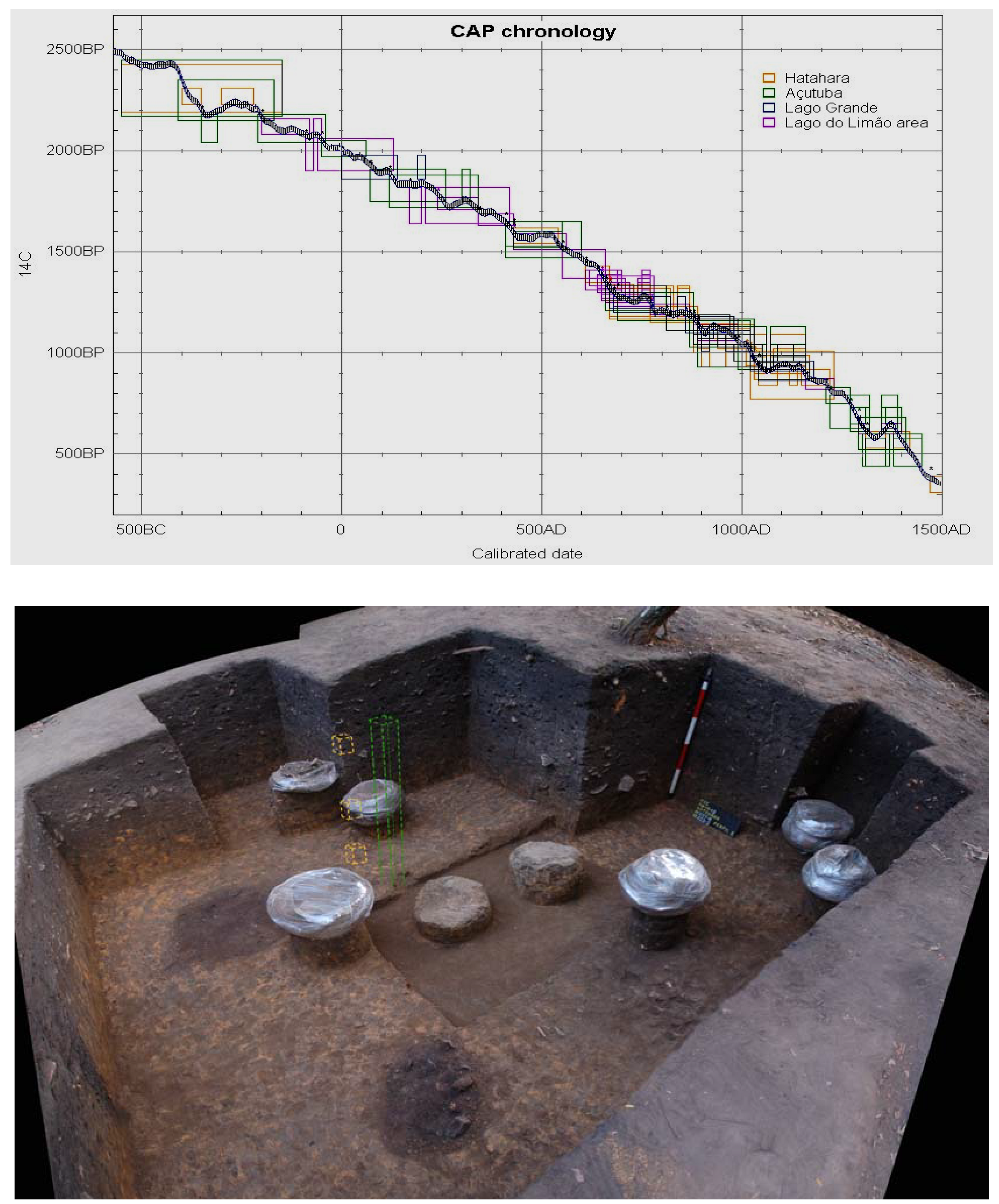
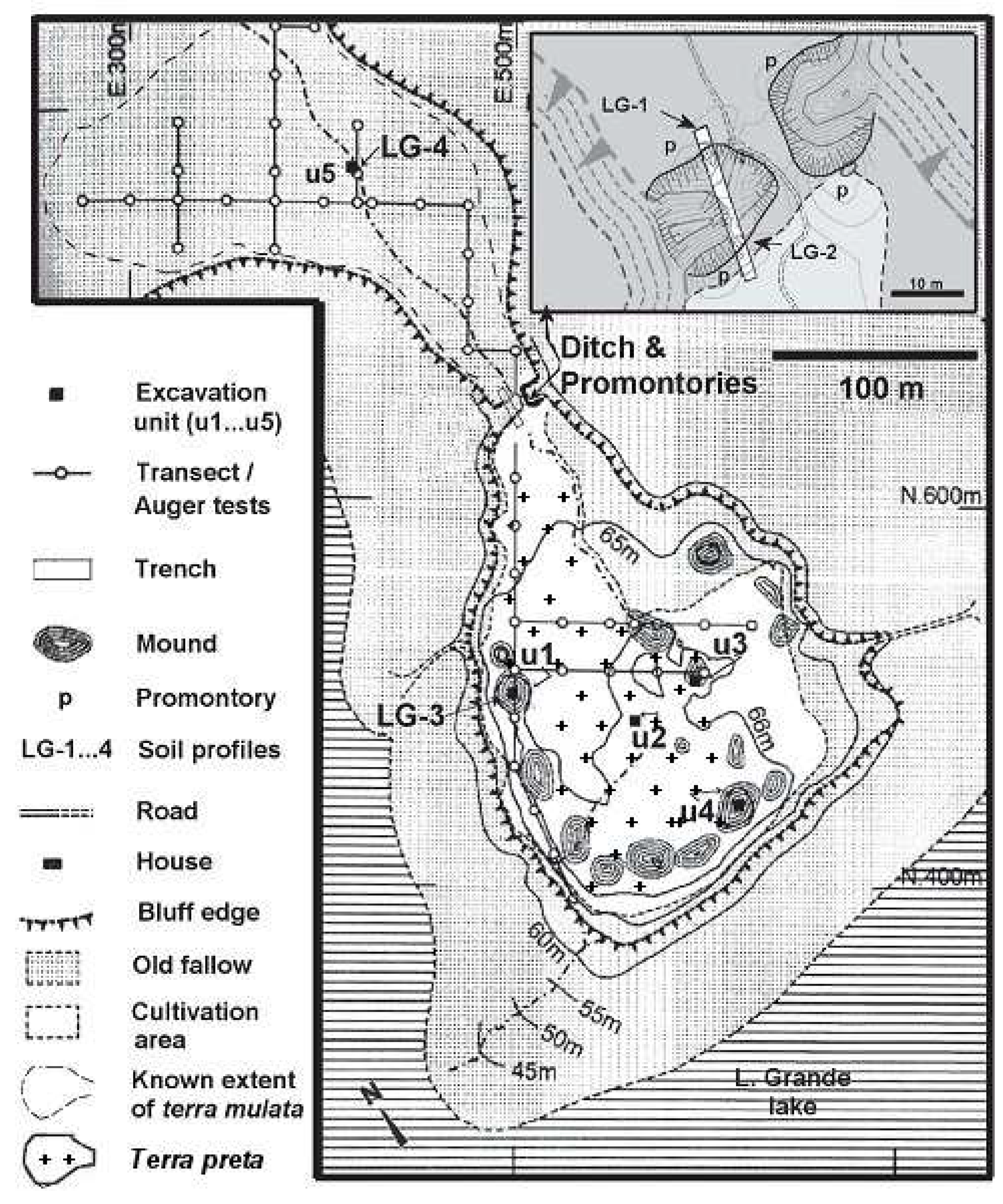
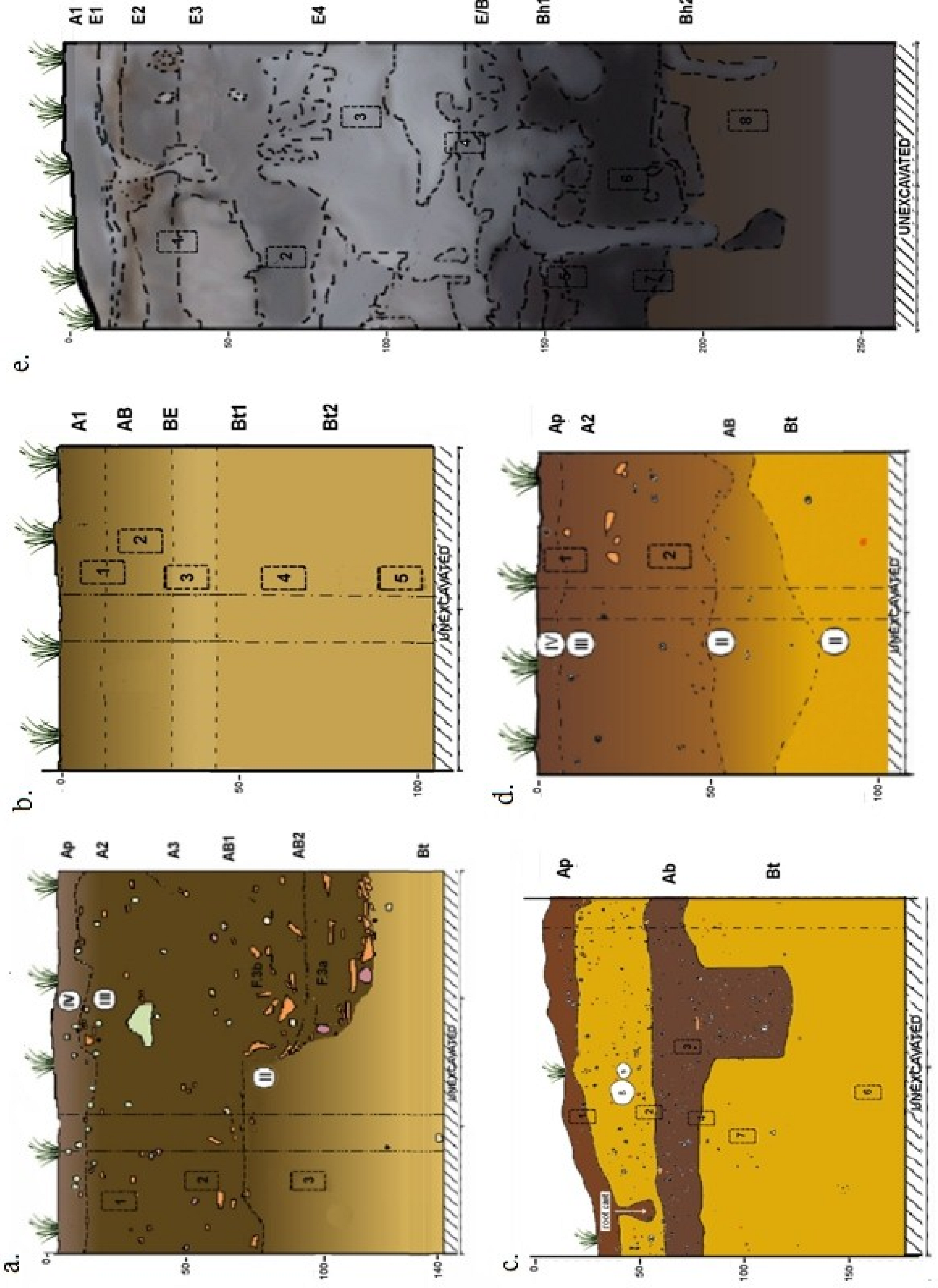
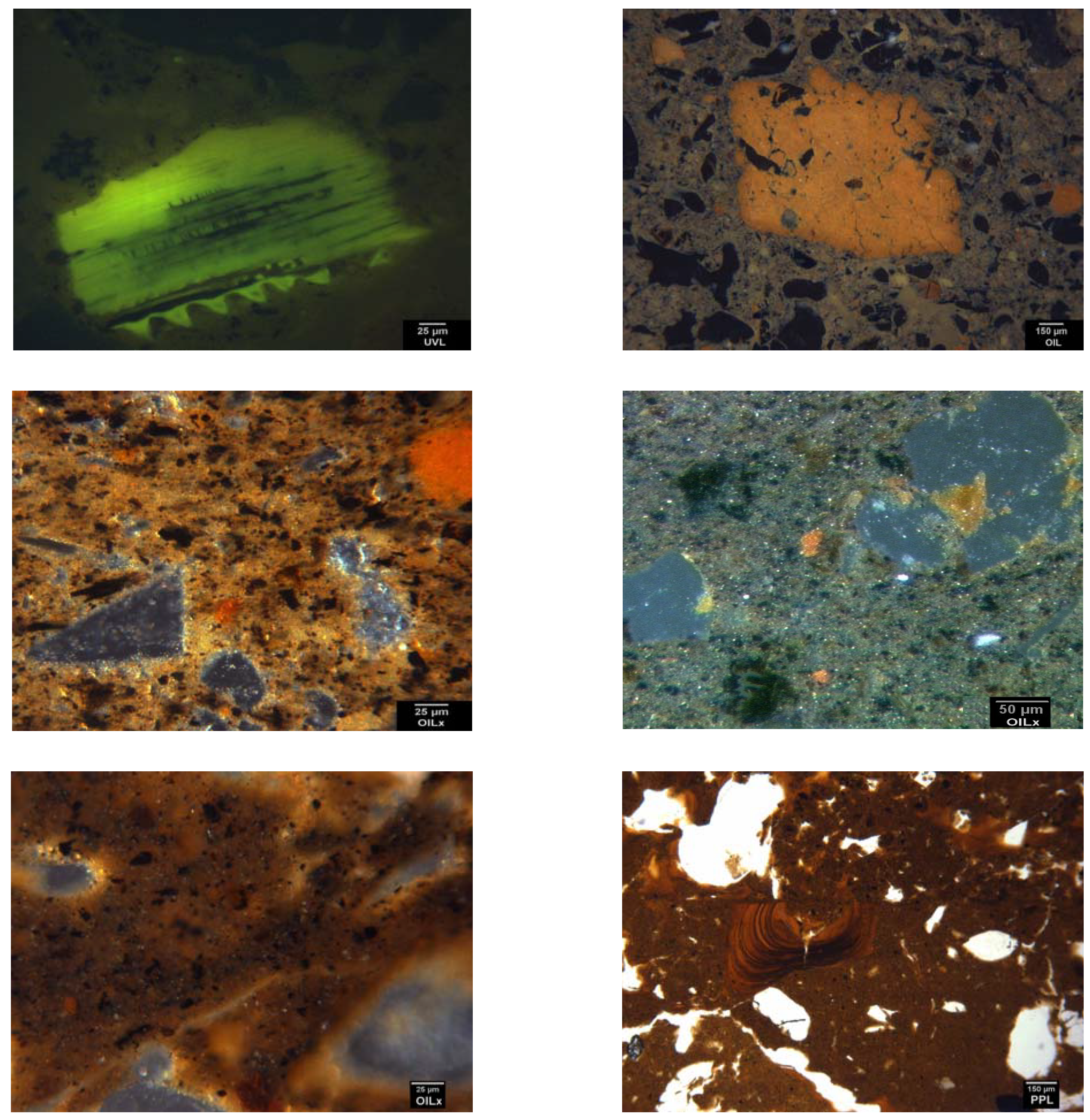
3. Domestication in the Landscape
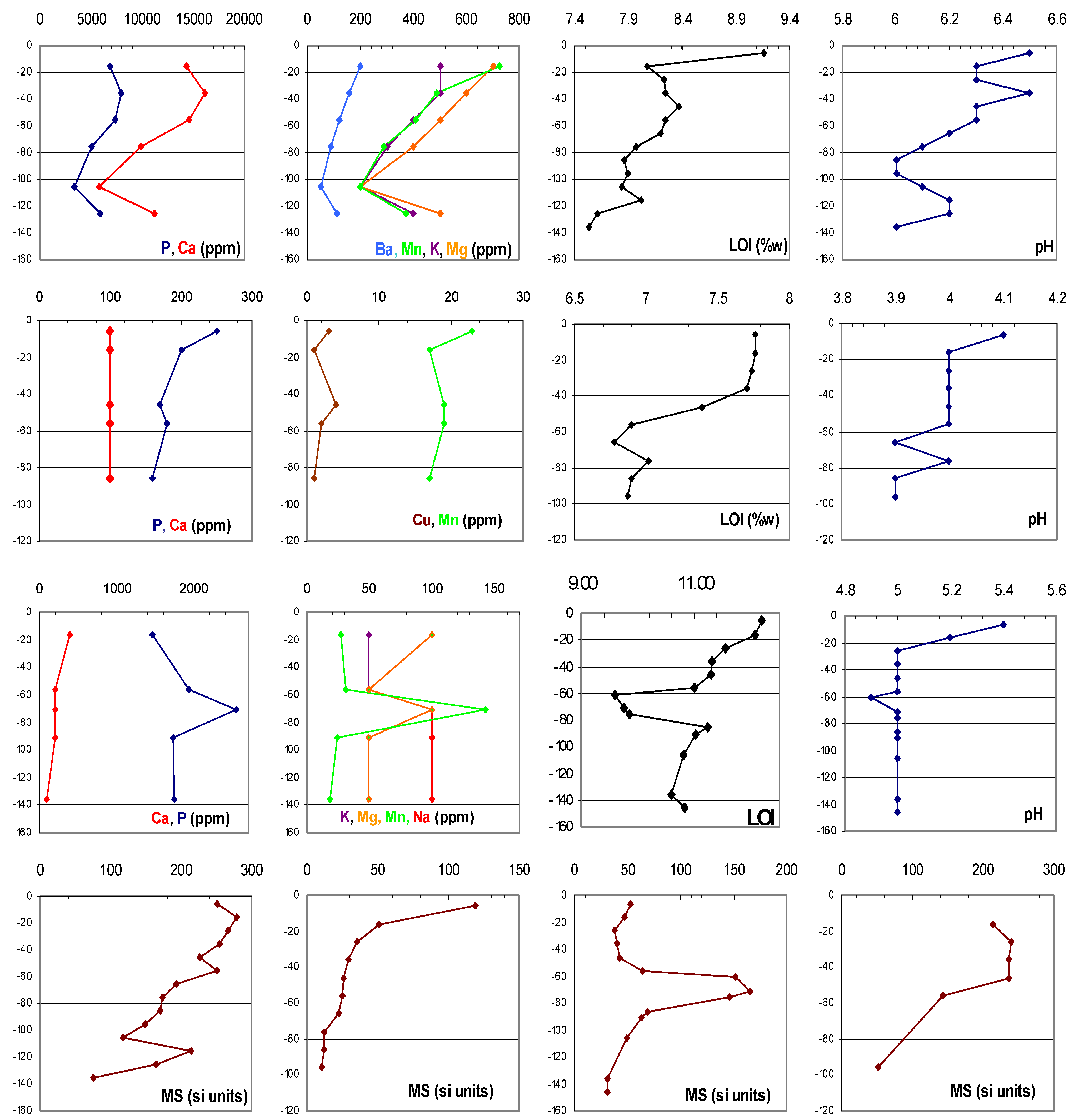
4. Anthropogenic Soils and Manioc

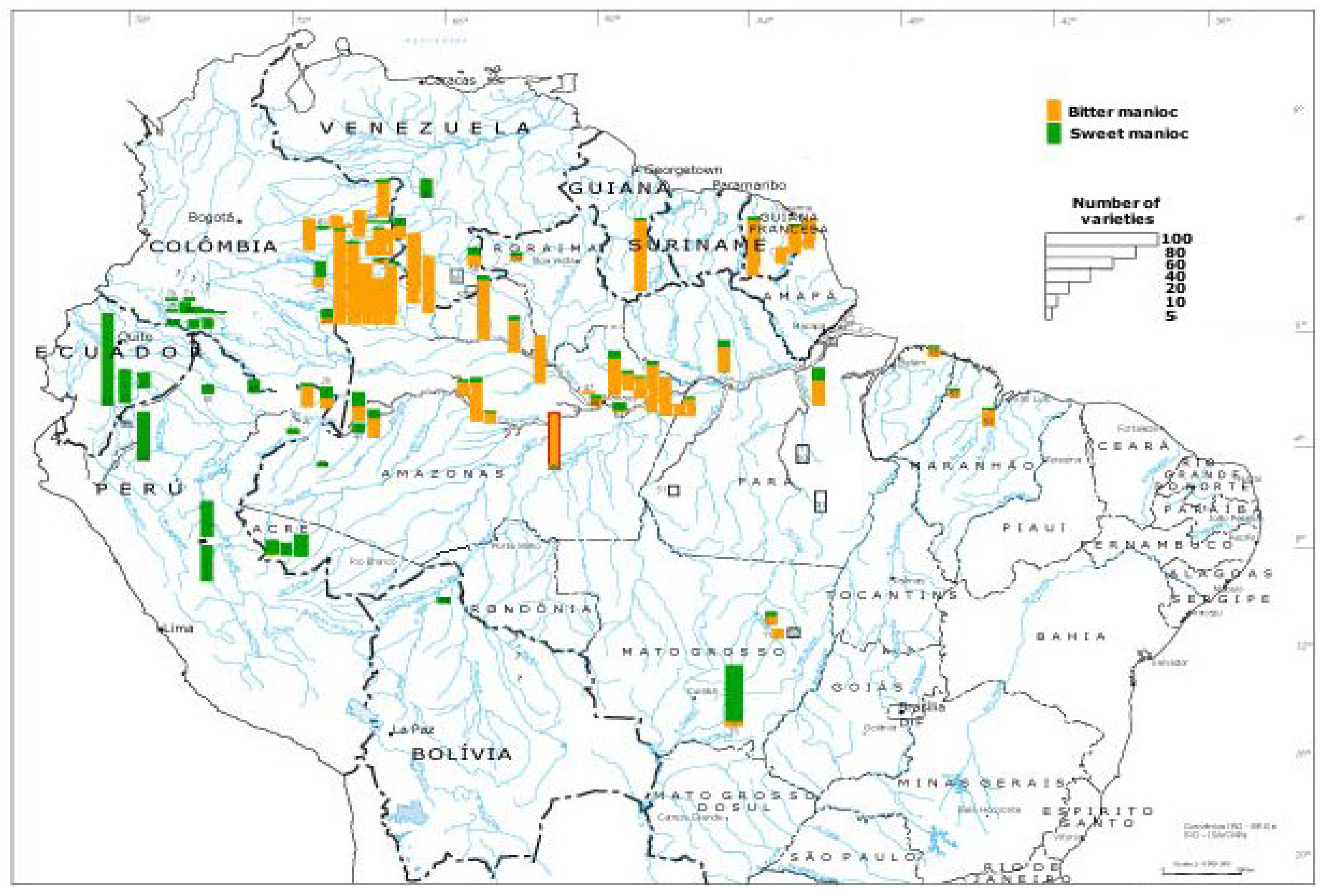
5. Crop Domestication and Intensification
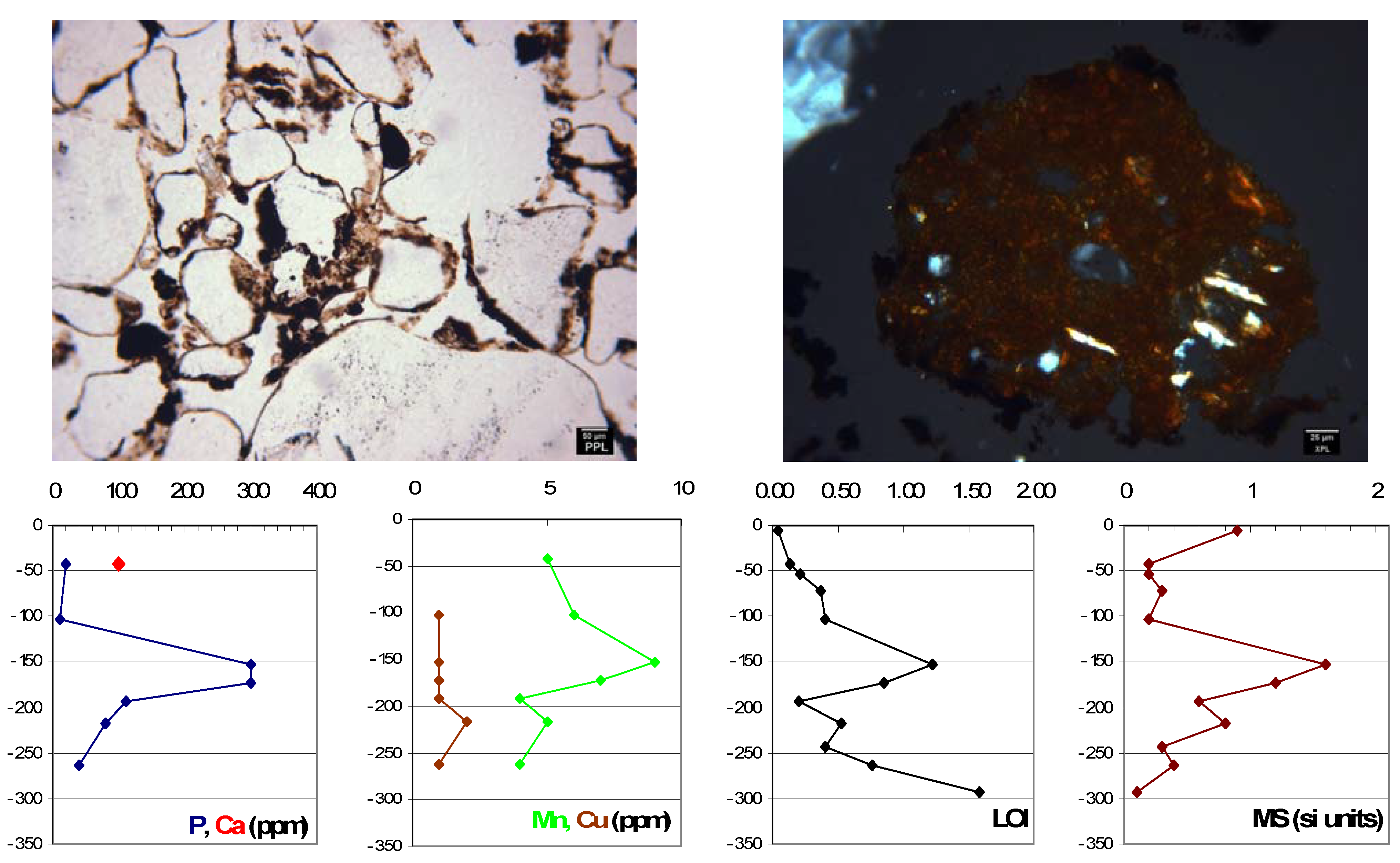
6. Discussion
7. Conclusion
- 1. One reviewer of this paper suggested that sequencing chloroplast non-coding areas or even specific nuclear genes (perhaps even the G3pdh already used in manioc) could allow - through coalescence analysis - estimates about when a specific lineage of the crop derived from another or from a wild ancestral population.
Acknowledgements
References
- Willey, G.R.; Phillips, P. Method and Theory in American Archaeology; University of Chicago Press: Chicago, IL, USA, 1965. [Google Scholar]
- Raymond, J.S. Beginnings of Sedentism in the Lowlands of Northwestern South America. In Recent Advances in the Archaeology of the Northen Andes: In Memory of Gerardo Reichel-Dolmatoff; Oyuela-Caycedo, A., Raymond, J.S., Eds.; Institute of Archaeology, University of California: Los Angeles, CA, USA, 1998; Volume 39, pp. 10–19. [Google Scholar]
- Marcos, J.G. A reassessment of the Ecuadorian Formative. In Archaeology of Formative Ecuador; Raymond, J.S., Burger, R.L., Eds.; Dumbarton Oaks Research Library and Collection: Washington, DC, USA, 2003; pp. 7–32. [Google Scholar]
- Mosley, M. The Maritime Foundations of Andean Civilization; Menlo Park: Cummings, 1975. [Google Scholar]
- Schaan, D.P. The Camutins chiefdom: Rise and development of social complexity on Marajó island, Brazilian Amazon. In PhD; University of Pittsburgh: Pittsburgh, PA, USA, 2004. [Google Scholar]
- Lathrap, D.W. The Upper Amazon; Thames & Hudson: Southampton, UK, 1970. [Google Scholar]
- Piperno, D.R.; Pearsall, D. The Origins of Agriculture in the Lowland Neotropics; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Oliver, J.R. On the road to agriculture in Amazonia: an overview. In Handbook of South American Archaeology; Silverman, H., Isbell, B., Eds.; Springer: New York, NY, USA, 2008; pp. 185–216. [Google Scholar]
- Brochado, J.; Lathrap, D. Amazonian Archaeology. 1982; unpublished manuscriptin possession of the author. [Google Scholar]
- Meggers, B.J. Prehistoria Sudamericana: Nuevas Perspectivas; Taraxacum: Washington, DC, USA, 1992; p. 381. [Google Scholar]
- Meggers, B.J. Judging the Future by the Past: The Impact of Environmental Instability on Prehistoric Amazonian Populations. In Indigenous Peoples and the Future of Amazonia: an Ecological Anthropology of an Endangered World; Sponsel, L.E., Ed.; The University of Arizona Press: Tucson, AZ, USA, 1995; pp. 15–43. [Google Scholar]
- Whitehead, N.L. Amazonian archaeology; searching for paradise? A review of recent literature and fieldwork. J. Archaeol. Res. 1996, 4, 241–264. [Google Scholar] [CrossRef]
- Clement, C.R. 1492 and the loss of Amazonian crop genetic resources. I The relation between domestication and human population decline. Econ. Bot. 1999, 53, 188–202. [Google Scholar] [CrossRef]
- Roosevelt, A.C. The maritime, highland, forest dynamic and origins of complex culture. In The Cambridge History of the Native Peoples of the Americas: South America; Salomon, F., Schwartz, S.B., Eds.; Cambridge University Press: Cambridge, MA, UK, 1999; Volume 2, pp. 264–349. [Google Scholar]
- Oliver, J.R. The archaeology of forest foraging and agricultural production in Amazonia. In Unknown Amazon. Culture in Nature in Ancient Brazil, 1st ed.; McEwan, C., Barreto, C., Neves, E.G., Eds.; The British Museum Press: London, UK, 2001; pp. 50–85. [Google Scholar]
- Heckenberger, M.J. Rethinking the Arawakan diaspora: Hierarchy, regionality and the Amazonian Formative. In Comparative Arawakan Histories: Rethinking Language Family and Culture Area in Amazonia; Hill, J.D., Santos-Granero, F., Eds.; University of Illinois Press: Chicago, IL, USA, 2002; pp. 99–122. [Google Scholar]
- Neves, E.G. Ecology, ceramic chronology and distribution, long-term history, and political change in the Amazonian floodplain. In Handbook of South American Archaeology; Silverman, H., Isbell, W., Eds.; Springer: New York, NY, USA, 2008; pp. 359–379. [Google Scholar]
- Heckenberger, M.J.; Neves, E.G. Amazonian Archaeology. Annu. Rev. Anthropol. 2009, 38, 251–266. [Google Scholar] [CrossRef]
- Viveiros de Castro, E. Images of nature and society in Amazonian ethnology. Annu. Rev. Anthropol. 1996, 25, 179–200. [Google Scholar] [CrossRef]
- Neves, E.G. Changing perspectives in Amazonian archaeology. In Archaeology of Latin America; Politis, G., Alberti, B., Eds.; Routledge: London, UK, 1999; pp. 217–243. [Google Scholar]
- Stahl, P.W. Paradigms in paradise: Revising standard Amazonian prehistory. Rev. Archaeol. 2002, 23, 39–51. [Google Scholar]
- Meggers, B.J. Amazonia: Man and Culture in a Counterfeit Paradise; Smithsonian Institution Press: Washington, DC, USA, 1971. [Google Scholar]
- Meggers, B.J. Vegetational fluctuation and prehistoric cultural adaptation in Amazonia: Some tentative correlations. World Archaeol. 1977, 8, 287–303. [Google Scholar] [CrossRef]
- Meggers, B.J. Climatic oscillation as a factor in the prehistory of Amazonia. Am. Antiq. 1979, 44, 252–266. [Google Scholar] [CrossRef]
- Meggers, B.J. Archaeological evidence for the impact of mega-nino events on Amazonia during the past 2 millennia. Clim. Change 1994, 28, 321–338. [Google Scholar] [CrossRef]
- Meggers, B.J. El contexto ecológico del Formativo. In Formativo Sudamericano, una Revaluación, 1st ed.; Ledergerber-Crespo, P., Ed.; Abya-Yala: Quito, Ecuador, 1999; pp. 383–393. [Google Scholar]
- Lathrap, D.W. Our father the cayman, our mother the gourd: Spinden revisited or a unitary model for the emergence of agriculture in the New World. In Origins of Agriculture; Reed, C.E., Ed.; Mouton: The Hague, The Netherlands, 1977; pp. 713–751. [Google Scholar]
- Lathrap, D.W. Review of David Rindos’ “The Origins of Agriculture: an Evolutionary Perspective”. Econ. Geogr. 1984, 60, 339–344. [Google Scholar] [CrossRef]
- Simões, M.F. Coletores-pescadores ceramistas do litoral do Salgado (Pará). Boletim do Museu Paraense Emílio Goeldi, NS 1981, 78, 1–26. [Google Scholar]
- Roosevelt, A.C.; Housley, R.A.; Da Silveira, M.I.; Maranca, S.; Johnson, R. Eighth millennium pottery from a prehistoric shell midden in the Brazilian Amazon. Science 1991, 254, 1621–1624. [Google Scholar]
- Roosevelt, A.C. Early pottery in the Amazon: twenty years of scholarly obscurity. In The Emergence of Pottery: Technology and Innovation in Ancient Societies; Barnett, W.K., Hoopes, J.W., Eds.; Smithsonian Institution: Washington, DC, USA, 1995; pp. 115–131. [Google Scholar]
- Roosevelt, A.C.; Lima da Costa, M.; Lopes Machado, C.; Michab, M.; Mercier, N.; Valladas, H.; Feathers, J.; Barnett, W.; Imazio da Silveira, M.; Henderson, A.; Silva, J.; Chernoff, B.; Reese, D.S.; Holman, J.A.; Toth, N.; Schick, K. Paleoindian cave dwellers in the Amazon: The peopling of the Americas. Science 1996, 272, 373–384. [Google Scholar]
- Roosevelt, A.C. The lower Amazon: A dynamic human habitat. In Imperfect Balance: Landscape Transformations in the Precolumbian Americas, 1st ed.; Lentz, D.L., Ed.; Columbia University Press: New York, NY, USA, 2000; pp. 455–491. [Google Scholar]
- Miller, E.T. Pesquisas arqueológicas paleoindígenas no Brasil Ocidental. Estudios Atacameños 1987, 8, 37–61. [Google Scholar]
- Miller, E.T. Adaptação agrícola pré-histórica no alto rio Madeira. In Prehistoria Sudamericana: Nuevas Perspectivas; Meggers, B.J., Ed.; Taraxacum: Washington, DC, USA, 1992; pp. 219–231. [Google Scholar]
- Miller, E.T. Archaeology in the Hydrolectric projects of Eletronorte; Eletronorte: Brasília, Brazil, 1992. [Google Scholar]
- Miller, E.T. A limitação ambiental como barreira à transformação do período Formativo no Brasil. Tecnologia, produção de alimentos e formação de aldeias no sudoeste da Amazônia. In Formativo Sudamericano, una Revaluación, 1st ed.; Ledergerber-Crespo, P., Ed.; Abya-Yala: Quito, Ecuador, 1999; pp. 331–339. [Google Scholar]
- Mora, S. Cultivars, Anthropic Soils, and Stability: A Preliminary Report of Archaeological Research in Araracuara, Colombian Amazonia; University of Pittsburgh: Pittsburgh, PA, USA, 1991; p. 87. [Google Scholar]
- Evans, C.; Meggers, B.J. Archaeological Investigations on the Rio Napo, Eastern Ecuador; Smithsonian Institution Press: Washington, DC, USA, 1968; Volume 6. [Google Scholar]
- Hilbert, P.P. Archäologische Untersuchungen am mittleren Amazonas: Beiträge zur Vorgeschichte des südamerikanischen Tieflandes; Reimer: Berlin, 1968. [Google Scholar]
- Meggers, B.J. Reconstrução do comportamento locacional pré-histórico na Amazônia. Boletim do Museu Paraense Emílio Goeldi - Série Antropologia 1990, 6, 183–203. [Google Scholar]
- Meggers, B.J. The continuing quest for El Dorado: Part two. Lat. Am. Antiq. 2001, 12, 304–325. [Google Scholar] [CrossRef]
- Neves, E.G.; Petersen, J.; Bartone, R.; da Silva, C.A. Historical and socio-cultural origins of Amazonian dark earths. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers: Dordrecht, Dunch, The Netherlands, 2003; pp. 29–50. [Google Scholar]
- Neves, E.G.; Petersen, J.B.; Bartone, R.N.; Heckenberger, M.J. The timing of terra preta formation in the central Amazon: archaeological data from three sites. In Amazonian Dark Earths: Explorations in Space and Time; Glaser, B., Woods, W.I., Eds.; Springer: London, UK, 2004; pp. 125–134. [Google Scholar]
- Lima, H.P.; Neves, E.G.; Petersen, J.B. A fase Açutuba: Um novo complexo cerâmico na Amazônia central. Arqueologia Sul-Americana 2006, 2, 26–52. [Google Scholar]
- Herrera, L.F.; Cavelier, I.; Rodríguez, C.; Mora, S. The technical transformation of an agricultural system in the Colombian Amazon. World Archaeol. 1992, 24, 98–113. [Google Scholar]
- Roosevelt, A.C. The rise and fall of the Amazon chiefdoms. L’Homme 1993, 33, 255–283. [Google Scholar]
- Roosevelt, A.C. The development of prehistoric complex societies: Amazonia, a tropical forest. In Complex Polities in the Ancient Tropical World; Bacus, E.A., Lucero, L.J., Eds.; American Anthropological Association: Arlington, VA, USA, 1999; Volume 9, pp. 13–33. [Google Scholar]
- Heckenberger, M.J.; Petersen, J.B.; Neves, E.G. Village size and permanence in Amazonia: two archaeological examples from Brazil. Lat. Am. Antiq. 1999, 10, 353–376. [Google Scholar] [CrossRef]
- Heckenberger, M.J. The Ecology of Power: Culture, Place and Personhood in the Southern Amazon AD 1000–2000; Routledge: London, UK, 2005. [Google Scholar]
- Morcote-Ríos, G. Plantas y gentes antiguas en un igapó estacional del interfluvio Solimões-Iça (Amazonas-Putumayo). In Pueblos y paisajes antiguos de la selva amazónica; Morcote, G., Mora, S., Franky, C., Eds.; Universidad Nacional de Colombia/Taraxacum: Bogotá/Washington, DC, USA, 2006; pp. 253–259. [Google Scholar]
- Gomes, D.M.C. O uso social da cerâmica de Parauá, Santarém, baixo Amazonas: uma análise funcional. Arqueología Sul-Americana 2008, 4, 4–33. [Google Scholar]
- Bozarth, S.R.; Neves, E.G.; Woods, W.I.; Cascon, L.; Caromano, C. Phytoliths and terra preta: the Hatahara site example. In Terra Preta Nova: A Tribute to Wim Sombroek; Woods, W.I., Teixeira, W.G., Lehmann, J., Steiner, C., WinklerPrins, A., Rebellato, L., Eds.; Kluwer: Dordrecht, The Netherlands, 2008; pp. 85–99. [Google Scholar]
- Meggers, B.J. La cerámica temprana en América del Sur: ¿invención independiente o difusión? Revista de Arqueología Americana 1997, 13, 7–40. [Google Scholar]
- Arroyo-Kalin, M. Steps towards an ecology of landscape: a geoarchaeological approach to the study of anthropogenic dark earths in the Central Amazon region. PhD thesis, University of Cambridge, Cambridge, UK, 2008. [Google Scholar]
- Allen, W.L. A ceramic sequence from the alto pachitea, perú: some implications for the development of tropical forest culture in South America. PhD thesis, University of Illinois, Urbana, IL, USA, 1968. [Google Scholar]
- Lathrap, D.W. The tropical forest and the cultural context of Chavín. In Proceedings of the Dumbarton Oaks Conference on Chavin; Benson, E.P., Ed.; Trustees for Harvard University: Washington, DC, USA, 1971; pp. 73–100. [Google Scholar]
- Shady, R. Sociedades formativas del nororiente peruano. In Prehistoria Sudamericana: Nuevas Perspectivas; Meggers, B., Ed.; Taraxacum: Santiago, Chile, 1992; pp. 343–357. [Google Scholar]
- Morales, D. Chambira: alfareros tempranos de la Amazonia Peruana. In Estudios de Arqueología Peruana; Bonavia, D., Ed.; Fomciencias: Lima, 1992; pp. 149–157. [Google Scholar]
- DeBoer, W.R. Ceramic assemblage variability in the Formative of Ecuador and Peru. In Archaeology of Formative Ecuador; Raymond, J.S., Burger, R.L., Eds.; Dumbarton Oaks Research Library and Collection: Washington, DC, USA, 2003; pp. 289–336. [Google Scholar]
- Valdez, F.; Guffroy, J.; de Saulieu, G.; Hurtado, J.; Yepes, A. Découvert d'un site cérémoniel formatif sur le versant oriental des Andes. C. R. Palevol. 2005, 4, 369–374. [Google Scholar] [CrossRef]
- Piperno, D.R. Aboriginal agriculture and land usage in the Amazon basin, Ecuador. J. Archaeol. Sci. 1990, 17, 665–677. [Google Scholar] [CrossRef]
- Bush, M.B.; Colinvaux, P.A. A 7000-year pollen record from the Amazon lowlands, Ecuador. Vegetatio 1988, 76, 141–154. [Google Scholar]
- Bush, M.B.; Miller, M.C.; De Oliveira, P.E.; Colinvaux, P.A. Two histories of environmental change and human disturbance in eastern lowland Amazonia. Holocene 2000, 10, 543–553. [Google Scholar] [CrossRef]
- Bush, M.B.; Silman, M.R.; de Toledo, M.B.; Listopad, C.; Gosling, W.D.; Williams, C.; de Oliveira, P.E.; Krisel, C. Holocene fire and occupation in Amazonia: records from two lake districts. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 2007, 362, 209–218. [Google Scholar] [CrossRef]
- Bush, M.B.; Silman, M.R.; McMichael, C.; Saatchi, S. Fire, climate change and biodiversity in Amazonia: a late-Holocene perspective. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 2008, 363, 1795–1802. [Google Scholar] [CrossRef]
- Sombroek, W.G. Amazon Soils: a Reconnaissance of the Soils of the Brazilian Amazon Region; Centre for Agricultural Publications and Documentation: Wageningen, The Netherlands, 1966. [Google Scholar]
- Lehmann, J.; Kern, D.; Glaser, B.; Woods, W. Amazonian Dark Earths. Origins, Properties and Management; Kluwer Press: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Glaser, B.; Woods, W.I. Amazonian Dark Earths: Explorations in Space and Time; Springer: London, UK, 2004; Volume xiv, p. 216. [Google Scholar]
- Woods, W.; Teixeira, W.; Lehmann, J.; Steiner, C.; WinklerPrins, A.; Rebellato, L. Amazonian Dark Earths: Wim Sombroek's Vision; Kluwer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Hartt, C.F. Contribuções para a ethnologia do valle do Amazonas. Arquivos do Museu Nacional do Rio de Janeiro 1885, 6, 1–94. [Google Scholar]
- Nimuendajú, C. Os Tapajó. Boletim do Museu Paraense Emílio Goeldi 1949, 10, 93–106. [Google Scholar]
- Neves, E.G. The relevance of Curt Nimuendajú's archaeological work. In In Pursuit of a Past Amazon. Archaeological Researches in the Brazilian Guyana and in the Amazon region; Stenborg, P., Ed.; Värlskulturmuseet i Göteborg: Gothenburg, 2004; Volume 45, pp. 2–8. [Google Scholar]
- Smith, N.K.H. Anthrosols and human carrying capacity in Amazonia. Ann. Assoc. Am. Geogr. 1980, 70, 553–566. [Google Scholar] [CrossRef]
- Herrera, L.; Bray, W.; McEwan, C. Datos sobre la arqueología de Araracuara (comisaría del Amazonas, Colombia). Revista Colombiana de Antropología 1980-1981, 23, 183–251. [Google Scholar]
- Herrera, L. Relaciones entre ocupaciones prehispánicas y suelos negros en la cuenca del río Caquetá en Colombia. Revista CIAF 1981, 6, 225–242. [Google Scholar]
- Eden, M.J.; Bray, W.; Herrera, L.; McEwan, C. Terra preta soils and their archaeological context in the Caquetá basin of southeast Colombia. Am. Antiq. 1984, 49, 125–140. [Google Scholar] [CrossRef]
- Simões, M.F.; Corrêa, C.G. Pesquisas arqueológicas no baixo Uatumã-Jatapu (Amazonas). Revista de Arqueologia, Belém 1987, 4, 29–48. [Google Scholar]
- Simões, M.F.; Kalmann, A.L.M. Pesquisas arqueológicas no Médio Rio Negro (Amazonas). Revista de Arqueologia, Belém 1987, 4, 83–116. [Google Scholar]
- Neves, E.G. Levantamento Arqueológico da Área de Confluência dos rios Negro e Solimões,Estado do Amazonas: Continuidade das Escavações,Análise da Composição Química e Montagem de um Sistema de Informações Geográficas; FAPESP: São Paulo, Brazil, 2003. [Google Scholar]
- Moraes, C.d.P. Arqueologia na Amazônia Central Vista de uma Perspectiva da Região do Lago do Limão. Masters, Universidade de São Paulo, São Paulo, Brazil, 2006. [Google Scholar]
- Andrade, Á. Investigación arqueológica de los antrosolos de Araracuara (Amazonas). Arqueología Colombiana 1986, 31, 1–101. [Google Scholar]
- Woods, W.I.; McCann, J.M.; Meyer, D.W. Amazonian dark earth analysis: state of knowledge and directions for future research. Papers and Proceedings of the Applied Geography Conferences 2000, 23, 114–121. [Google Scholar]
- Glaser, B.; Balashov, E.; Haumaier, L.; Guggenberger, G.; Zech, W. Black carbon in density fractions of anthropogenic soils of the Brazilian Amazon region. Org. Geochem. 2000, 21, 669–678. [Google Scholar]
- Glaser, B.; Guggenberger, G.; Zech, W.; Ruivo, M.d.L. Soil organic mater stability in Amazonian dark earths. In Amazonian Dark Earths: Origins, Properties and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer Press: Dordrecht, The Netherlands, 2003; pp. 141–158. [Google Scholar]
- Lima, H.N.; Schaefer, C.E.R.; Mello, J.W.V.; Gilkes, R.J.; Ker, J.C. Pedogenesis and pre-Colombian land use of "Terra Preta Anthrosols" ("Indian black earth") of western Amazonia. Geoderma 2002, 110, 1–17. [Google Scholar] [CrossRef]
- Kern, D.C.; Kämpf, N. Antigos assentamentos indígenas na formação de solos com terra preta arqueológica na região de Oriximinã, Para. Revista Brasileira de Ciência do Solo 1989, 13, 219–225. [Google Scholar]
- Kern, D.C. Geoquímica e pedogeoquímica em sítios arqueológicos com terra preta na floresta nacional de Caxiuanã (Portel-PA). PhD, Universidade Federal do Pará, Belém, Brazil, 1996. [Google Scholar]
- Erickson, C.L. Historical ecology and future explorations. In Amazonian Dark Earths: Origins, Properties and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer Press: Dordrecht, The Netherlands, 2003; pp. 455–500. [Google Scholar]
- Rebellato, L. Interpretando a variabilidade cerâmica e as assinaturas químicas e físicas do solo no sítio arqueológico Hatahara -AM. Masters, Universidade de São Paulo, Brazil, 2007. [Google Scholar]
- Zeidler, J.A. La etnoarqueología de una vivienda Achuar y sus implicaciones arqueológicas. Miscelánea Antropológica Ecuatoriana 1983, 3, 155–193. [Google Scholar]
- Hecht, S.B. Indigenous soil management and the creation of Amazonian dark earths: implications of Kayapó practices. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 355–372. [Google Scholar]
- Silva, F.A. Cultural behaviors of indigenous populations and the formation of the archaeological record in Amazonian dark earths: the Asurini do Xingú case study. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W.I., Eds.; Kluwer Press: Dordrecht, The Netherlands, 2003; pp. 373–385. [Google Scholar]
- Silva, F.A.; Rebellato, L. Use of space and formation of Terra Preta: the Asuriní do Xingu case study. In Amazonian Dark Earths: Explorations in Space and Time; Glaser, B., Woods, W.I., Eds.; Springer: Berlin, Germany, 2004; pp. 159–167. [Google Scholar]
- Schmidt, M.; Heckenberger, M.J. Amazonian dark earth formation in the Upper Xingú of southeastern Amazonia, Mato Grosso, Brazil. In 71st Annual meeting of the Society of American Archaeology; San Juan, Puerto Rico, 2006. [Google Scholar]
- Simões, M.F. Contribução á arqueologia dos arredores do baixo rio Negro, Amazonas. Programa Nacional de Pesquisa Arqueológicas. Resultados preliminares do quinto ano 1969–1970. Publicações avulsas 1974, 5, 165–188. [Google Scholar]
- Simões, M.F.; Machado, A.L. Pesquisas arqueológicas no lago de Silves (Amazonas). Revista de Arqueologia, Belém 1987, 4, 49–82. [Google Scholar]
- Machado, J.S. A Formação de Montículos Artificiais: um Estudo de Caso no Sítio Hatahara, Amazonas. Masters, Universidade de São Paulo, São Paulo, Brazil, 2005. [Google Scholar]
- Woods, W.I. Comments on the black earths of Amazonia. Papers and Proceedings of the Applied Geography Conferences 1995, 18, 159–165. [Google Scholar]
- Arroyo-Kalin, M.; Neves, E.G.; Woods, B.; Bartone, R.; Lima, E.P.; Rebellato, L.; Moraes, C.P.; Silva, C.A.d.; Daniel, A.R.-P. Mix and grow? Assessing the geoarchaeological significance of Terras Pretas in the Central Amazon region. In Developing International Geoarchaeology 2007; Cambridge, 2007. [Google Scholar]
- Arroyo-Kalin, M. Steps towards an ecology of landscape: the pedo-stratigraphy of anthropogenic dark earths. In Amazonian Dark Earths: Wim Sombroek’s Vision; Woods, W., Teixeira, W., Lehmann, J., Steiner, C., WinklerPrins, A., Rebellato, L., Eds.; Kluwer: Dordrecht, The Netherlands, 2008; pp. 33–83. [Google Scholar]
- Arroyo-Kalin, M. Towards a historical ecology of pre-Columbian central Amazonia. In 71st annual meeting of the Society of American Archaeology; San Juan, Puerto Rico, 2006. [Google Scholar]
- Neves, E.G.; Petersen, J. The political economy of pre-Columbian Amerindians: landscape transformations in central Amazonia. In Time and Complexity in Historical Ecology: Studies in the Neotropical Lowlands; Balée, W., Erickson, C.L., Eds.; Columbia University Press: New York, NY, USA, 2006; pp. 279–310. [Google Scholar]
- Arroyo-Kalin, M.; Neves, E.G.; Woods, W.I. Anthropogenic dark earths of the Central Amazon region: remarks on their evolution and polygenetic composition. In Amazonian Dark Earths: Wim Sombroek's Vision; Woods, W., Teixeira, W., Lehmann, J., Steiner, C., WinklerPrins, A., Rebellato, L., Eds.; Kluwer: Dordrecht, The Netherlands, 2008; pp. 99–125. [Google Scholar]
- Schaefer, C.; Lima, H.N.; Gilkes, R.J.; Mello, J.W.V. Micromorphology and electron microprobe analysis of phosphorus and potassium forms of an Indian Black Earth (IBE) Anthrosol from Western Amazonia. Aust. J. Soil Res. 2004, 42, 401–409. [Google Scholar] [CrossRef]
- Sergio, C.S.; Santana, G.P.; da Costa, G.m.; Horbe, A.M.C. Identification and characterization of maghemite in ceramic artifacts and archaeological black earth of Amazon region. Soil Sci. 2006, 171, 59–64. [Google Scholar] [CrossRef]
- Topoliantz, S.; Ponge, J.F. Charcoal consumption and casting activity by Pontoscolex corethrurus (Glossoscolecidae). Appl. Soil Ecol. 2005, 28, 217–224. [Google Scholar] [CrossRef]
- Donatti, P.B. A Ocupação Pré-Colonial da Area do Lago Grande, Iranduba, AM. Masters, Universidade de São Paulo, São Paulo, Brazil, 2003. [Google Scholar]
- Andrade, Á. Desarrollo de los sistemas agrícolas tradicionales en la Amazonía. Boletín del Museo del Oro 1988, 21, 38–59. [Google Scholar]
- Mullins, C.E. Magnetic-susceptibility of soil and its significance in soil science: review. J. Soil Sci. 1977, 28, 223–246. [Google Scholar] [CrossRef]
- Steiner, C.; Teixeira, W.; Zech, W. Slash and char—an alternative to slash and burn practiced in the Amazon basin. In Amazonian Dark Earths: Explorations in Space and Time; Glaser, B., Woods, W.I., Eds.; Springer-Verlag: Heidelberg, Germany, 2004; pp. 182–193. [Google Scholar]
- Denevan, W.M. Semi-intensive pre-European cultivation and the origins of anthropogenic dark earths in Amazonia. In Amazonian Dark Earths: Explorations in Space and Time; Glaser, B., Woods, W.I., Eds.; Springer: Berlin, Germany, 2004; pp. 135–143. [Google Scholar]
- Balée, W. The culture of Amazonian forests. In Resource Management in Amazonia: Indigenous and Folk Strategies; Posey, D., Balée, W., Eds.; New York Botanical Garden: Bronx, NY, USA, 1989; Volume 7, pp. 1–21. [Google Scholar]
- Clement, C.R.; McCann, J.M.; Smith, N.J. Agrobiodiversity in Amazonia and its relationship with dark earths. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 159–178. [Google Scholar]
- Junqueira, A.B.; Shepard, G.H., Jr.; Clement, C.R. Secondary forests on anthropogenic soils in Brazilian Amazonia conserve agrobiodiversity. Biodiversity Conserv. 2010. [Google Scholar] [CrossRef]
- Woods, W.I.; McCann, J.M. The anthropogenic origin and persistence of Amazonian dark earths. Yearb. Conf. Lat. Am. Geogr. 1999, 25, 7–14. [Google Scholar]
- German, L.A. Historical contingencies in the coevolution of environment and livelihood: contributions to the debate on Amazonian Black Earth. Geoderma 2003, 111, 307–331. [Google Scholar] [CrossRef]
- Hiraoka, M.; Yamamoto, S.; Matsumoto, E.; Nakamura, S.; Falesi, I.C.; Ronaldo, A.; Baena, C. Contemporary use and management of Amazonian dark earths. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 387–406. [Google Scholar]
- Fraser, J.A.; Clement, C.R. Dark earths and manioc cultivation in central Amazonia: a window on pre-Columbian agricultural systems? Boletim do Museu Paraense Emílio Goeldi 2008, 3, 175–194. [Google Scholar] [CrossRef]
- Fraser, J.A. The diversity of bitter manioc (Manihot esculenta Crantz) cultivation in one of the richest landscapes of Amazonia. Diversity 2010. submitted. [Google Scholar]
- Balée, W. People of the fallow: an historical ecology of foraging in lowland South America. In Conservation of Neotropical Forests: Working from Traditional Resource Use; Redford, K.H., Padoch, C., Eds.; Columbia University Press: New York, NY, USA, 1992; pp. 35–46. [Google Scholar]
- Politis, G. Moving to produce: Nukak mobility and settlement patterns in Amazonia. World Archaeol. 1996, 27, 492–511. [Google Scholar] [CrossRef]
- Politis, G. Plant exploitation among the Nukak Hunter-gatherers of Amazonia: between ecology and ideology. In Prehistory of Food: Appetites for Change; Gosden, C., Hather, J., Eds.; Routledge: London, UK, 1999; pp. 99–126. [Google Scholar]
- Balée, W. The research program of Historical Ecology. Annu. Rev. Anthropol. 2006, 35, 75–98. [Google Scholar] [CrossRef]
- Clement, C.R. Domesticação de paisagens e plantas amazônicas—a interação de etnobotânica, genética molecular e arqueologia. In Pueblos, Plantas y Paisajes Antiguos en la Selva Tropical Amazónica.; Morcote, G., Mora, S., Franky, C., Eds.; Editorial Universidad Nacional: Bogotá, Colombia, 2006; pp. 97–112. [Google Scholar]
- Clement, C.R. Fruit trees and the transition to food production in Amazonia. In Time and Complexity in Historical Ecology: Studies in the Neotropical Lowlands; Balée, W., Erickson, C.L., Eds.; Columbia University Press: New York, NY, USA, 2006; pp. 165–185. [Google Scholar]
- Gnecco, C.; Mora, S. Late Pleistocene-early Holocene tropical forest occupations at San Isidro and Peña Roja, Colombia. Antiquity 1997, 71, 683–690. [Google Scholar]
- Mora, S.; Gnecco, C. Archaeological hunter-gatherers in tropical forests: a view from Colombia. In Under the Canopy: The Archaeology of Tropical Rainforests; Mercader, J., Ed.; Rutgers University Press: London, UK, 2003; pp. 271–290. [Google Scholar]
- Cardale Schrimpff, M. The earliest inhabitants. In Calima and Malagana: Art and Archaeology in Southwestern Columbia; Cardale Schrimpff, M., Ed.; Pro Calima Foundation: Bogotá, Colombia, 2005; pp. 26–35. [Google Scholar]
- Aceituno, F.J.; Castillo, N. Mobility strategies in Colombia's middle mountain range between the early and middle Holocene. Before Farming 2005, 2, 1–17. [Google Scholar]
- Castillo, N.; Aceituno, F. El bosque domesticado, el bosque cultivado: Un proceso milenario en el valle medio del río Porce en el noroccidente colombiano. Lat. Am. Antiq. 2006, 17, 561–578. [Google Scholar] [CrossRef]
- Morcote-Ríos, G.; Bernal, R. Remains of palms (palmae) at archaeological sites in the New World: A review. Botanical Rev. 2001, 67, 309–350. [Google Scholar] [CrossRef]
- Mora, S. Early Inhabitants of the Amazonian Tropical Rain Forest: A Study of Humans and Environmental Dynamics; University of Pittsburg: Pittsburg, PA, USA, 2003; Volume 3. [Google Scholar]
- Roosevelt, A.C. Review of Santiago Mora’s ‘Early Inhabitants of the Amazonian Tropical Rain Forest: A Study of Humans and Environmental Dynamics’. Am. Anthropol. 2006, 108, 250–251. [Google Scholar] [CrossRef]
- French, C.A.I. Geoarchaeology in Action: Studies in Soil Micromorphology and Landscape Evolution; Routledge: London, UK, 2003. [Google Scholar]
- Woods, W.I. Development of anthrosol research. In Amazonian Dark Earths: Origins, Properties and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer Press: Dordrecht, The Netherlands, 2003; pp. 3–14. [Google Scholar]
- Davidson, D.; Simpson, I. The time dimension in landscape ecology: cultural soils and spatial pattern in early landscapes. In Issues and Perspectives in Landscape Ecology; Wiends, J.A., Moss, M.R., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 152–158. [Google Scholar]
- Cavelier, I.; Rodríguez, C.; Herrera, L.F.; Morcote, G.; Mora, S. No sólo de caza vive el hombre: ocupación del bosque amazónico, Holoceno temprano. In Ámbito y Ocupaciones Tempranas de la América Tropical; Cavelier, I., Mora, S., Eds.; Fundación Erigaie, Instituto Colombiano de Antropología: Santafé de Bogotá, Colombia, 1995; pp. 27–44. [Google Scholar]
- Costa, F.W.d.S. Análise das Indústrias Líticas da Area de Confluência dos rios Negro e Solimões. Masters, Universidade de São Paulo, São Paulo, Brazil, 2002. [Google Scholar]
- Anderson, E. Plants, Man and Life; Little, Brown and co.: Boston, MA, USA, 1952. [Google Scholar]
- Descola, P. From scattered to nucleated settlement: a process of socioeconomic change among the Achuar. In Cultural Transformations and Ethnicity in Modern Ecuador; Whitten, N.E., Jr., Ed.; University of Chicago Press: Chicago, IL, USA, 1981; pp. 614–646. [Google Scholar]
- Descola, P. Homeostasis as a cultural system: the Jívaro case. In Amazonian Indians from Prehistory to the Present: Anthropological Perspectives; Roosevelt, A.C., Ed.; University of Arizona Press: Tucson, AZ, USA, 1994; pp. 203–224. [Google Scholar]
- Salick, J.; Cellinese, N.; Knapp, S. Indigenous diversity of cassava: generation, maintenance, use and loss among the Amuesha, Peruvian Upper Amazon. Econ. Bot. 1997, 51, 6–19. [Google Scholar] [CrossRef]
- Emperaire, L. Elementos de discussão sobre a conservação da agrobiodiversidade: o exemplo da mandioca (Manihot esculenta Crantz) na Amazônia brasileira. In Biodiversidade da Amazônia brasileira, avaliação e ações prioritárias para a conservação, uso sustentável e repartição dos benefícios; Capobiando, J.-P., Ed.; ISA, Estação Liberdade: São Paulo, Brazil, 2001; pp. 225–234. [Google Scholar]
- Porro, A. História indígena do alto e médio Amazonas: séculos XVI a XVIII. In História dos Índios no Brasil; Carneiro da Cunha, M., Ed.; Compahia das Letras-FAPESP-SMC (São Paulo)-Editora Schwarcz: São Paulo, Brazil, 1992; pp. 175–196. [Google Scholar]
- Porro, A. Social organisation and political power in the Amazon Floodplain: the ethnohistorical sources. In Amazonian Indians: From Prehistory to the Present. Anthropological Perspectives; Roosevelt, A.C., Ed.; University of Arizona Press: Tuson, AZ, USA, 1994; pp. 79–94. [Google Scholar]
- Evans, C.; Meggers, B.J. Archeological Investigations in British Guiana; Smithsonian Institution Press: Washington, DC, USA, 1960. [Google Scholar]
- Harris, D.R. The ecology of swidden cultivation in the Upper Orinoco Rain Forest, Venezuela. Geogr. Rev. 1971, 61, 475–495. [Google Scholar]
- Hugh-Jones, C. From the Milk River, Spatial and Temporal Processes in Northwest Amazonia; Cambridge University Press: Cambridge, UK, 1979. [Google Scholar]
- Dufour, D.L. The composition of some foods used in northwest Amazonia. Interciencia 1988, 13, 83–86. [Google Scholar]
- Dufour, D. Diet and nutritional status of Amazonian peoples. In Amazonian Indians, from Prehistory to the Present. Anthropological Perspectives; Roosevelt, A.C., Ed.; The University of Arizona Press: Tucson, AZ, USA, 1994; pp. 151–175. [Google Scholar]
- Carneiro, R.L. The cultivation of manioc among the Kuikuru of the upper Xingú. In Adaptive Responses of Native Amazonians; Hames, R.B., Vickers, W.T., Eds.; Academic Press: New York, NY, USA, 1983; pp. 65–112. [Google Scholar]
- Wright, R. Indian slavery in the northwest Amazon. Boletim do Museu Paraense Emílio Goeldi—Série Antropologia 1991, 7, 149–179. [Google Scholar]
- Silverwood-Cope, P.L. A contribution to the ethnography of the Colombian Makú. PhD thesis, University of Cambridge, Cambridge, UK, 1972. [Google Scholar]
- Reid, H.A. Some aspects of movement, growth and change among the Hupdu Makú Indians of Brazil. PhD thesis, University of Cambridge, Cambridge, UK, 1979. [Google Scholar]
- Koch-Grünberg, T. Dos Años entre los Indios; Editorial Universitaria Nacional: Bogotá, Colombia, 1995 (1909); vol 2. [Google Scholar]
- Goldman, I. The Cubeo: Indians of the Northwest Amazon; University of Illinois Press: Champaign, IL, USA, 1963. [Google Scholar]
- Chernela, J.M. Managing rivers of hunger: the Tukano of Brazil. In Resource Management in Amazonia: Indigenous and Folk Strategies; Posey, D., Balée, W., Eds.; New York Botanical Garden: Bronx, NY, USA, 1989; Volume 7, pp. 238–248. [Google Scholar]
- Chernela, J.M. Social meaning and material transaction: the Wanano-Tukano of Brazil and Colombia. J. Anthropol. Archaeol. 1992, 11, 111–124. [Google Scholar] [CrossRef]
- Piperno, D.R. Identifying crop plants with phytoliths (and starch grains) in Central and South America:A review and an update of the evidence. Quat. Int. 2009, 193, 146–159. [Google Scholar] [CrossRef]
- Chandler-Ezell, K.; Pearsall, D.M.; Zeidler, J.A. Root and tuber phytoliths and starch grains document manioc (Manihot esculenta), arrowroot (Maranta arundinacea), and Llerén (Calathea sp.) at the Real alto site, Ecuador. Econ. Bot. 2006, 60, 103–120. [Google Scholar]
- Olsen, K.M.; Schaal, B.A. Evidence on the origin of cassava—Phylogeography of Manihot esculenta. Proc. Natl. Acad. Sci. USA 1999, 96, 5586–5591. [Google Scholar] [CrossRef]
- Ugent, D. Chemosystematics in Archaeology: A Preliminary Study of the Use of Chromatography and Spectrophotometry in the Identification of Four Prehistoric Root Crop Species from the Desert Coast of Peru; Wac 2, Barquisimeto; Venezuela, Sep, 1990; Routledge: Barquisimeto; Venezuela, 1990; pp. 215–226. [Google Scholar]
- Ugent, D.; Pozorski, S.; Pozorski, T. Archaeological manioc (Manihot) from coastal Peru. Econ. Bot. 1986, 40, 78–102. [Google Scholar] [CrossRef]
- Rossen, J.; Dillehay, T.D.; Ugent, D. Ancient cultigens or modern intrusions?: Evaluating plant remains in an Andean case study. J. Archaeol. Sci. 1996, 23, 391–407. [Google Scholar] [CrossRef]
- Perry, L. Starch analyses reveal multiple functions of quartz “Manioc” grater flakes from the Orinoco Basin, Venezuela. Interciencia 2002, 27, 635–639. [Google Scholar]
- Piperno, D.R.; Holst, I. The presence of starch grains on prehistoric stone tools from the humid neotropics: indications of early tuber use and agriculture in Panama. J. Archaeol. Sci. 1998, 25, 765–776. [Google Scholar] [CrossRef]
- Piperno, D.R.; Ranere, A.J.; Holst, I.; Hansell, P. Starch grains reveal early root crop horticulture in the Panamanian tropical forest. Nature 2000, 407, 894–897. [Google Scholar] [CrossRef]
- Gnecco, C.; Aceituno, J. Poblamiento temprano y espacios antropogénicos en el norte de Suramérica. Complutum 2004, 15, 151–164. [Google Scholar]
- Behling, H.; Hooghiemstra, H. Late Quaternary palaeoecology and palaeoclimatology from pollen records of the savannas of the Llanos Orientales in Colombia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 139, 251–267. [Google Scholar] [CrossRef]
- Léotard, G.; Duputié, A.; Kjellberg, F.; Douzery, E.J.P.; Debain, C.; Granville, J.-J.d.; McKey, D. Phylogeography and the origin of cassava: New insights from the northern rim of the Amazonian basin. Mol. Phylogenet. Evol. 2009, 53, 329–334. [Google Scholar] [CrossRef]
- Olsen, K.M. SNPs, SSRs and inferences on cassava’s origin. Plant Mol. Biol. 2004, 56, 517–526. [Google Scholar] [CrossRef]
- Perry, L. Starch granule size and the domestication of manioc (Manihot esculenta) and sweet potato (Ipomoea batatas). Econ. Bot. 2002, 56, 335–349. [Google Scholar] [CrossRef]
- Freitas, H.A.d.; Pessenda, L.C.R.; Aravena, R.; Gouveia, S.E.M.; Ribeiro, A.D.; Boulet, R. Late Quaternary vegetation dynamics in the southern Amazon Basin inferred from carbon isotopes in soil organic matter. Quaternary Res. 2001, 55, 39–46. [Google Scholar] [CrossRef]
- Burbridge, R.E.; Mayle, F.E.; Killeen, T.J. Fifty-thousand-year vegetation and climate history of Noel Kempff Mercado National Park, Bolivian Amazon. Quaternary Res. 2004, 61, 215–230. [Google Scholar] [CrossRef]
- Mayle, F.E.; Langstroth, R.P.; Fisher, R.A.; Meir, P. Long-term forest-savannah dynamics in the Bolivian Amazon: implications for conservation. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 2007, 362, 291–307. [Google Scholar] [CrossRef]
- Pickersgill, B. Domestication of plants in the Americas: insights from mendelian and molecular genetics. Ann. Bot. 2007, 100, 925–940. [Google Scholar] [CrossRef]
- Clement, C.R.; Cristo-Araújo, M.d.; d’Eeckenbrugge, G.C.; Pereira, A.A.; Picanço-Rodrigues, D. Origin and Domestication of Native Amazonian Crops. Diversity 2010, 2, 72–106. [Google Scholar] [CrossRef]
- McKey, D.; Beckerman, S. Chemical ecology, plant evolution, and the evolution of traditional manioc cultivation systems. In Tropical Forests, People and Food.Biocultural Interactions and Applications to Development; Hladik, C.M., Hladik, A., Linares, O.F., Pagezy, H., Semple, A., Hadley, M., Eds.; UNESCo, Parthenon: Paris, France, 1933; pp. 83–112. [Google Scholar]
- Pujol, B.; David, P.; McKey, D. Germination ecology of cassava (Manihot esculenta Crantz, Euphorbiaceae) in traditional agroecosystems: seed and seedling biology of a vegetatively propagated domesticated plant. Econ. Bot. 2002, 56, 366–379. [Google Scholar] [CrossRef]
- Mühlen, G.S.; Martins, P.S.; Ando, A. Variabilidade genética de etnovariedades de mandioca, avaliada por marcadores de DNA. Sci. Agric. 2000, 57, 319–328. [Google Scholar]
- Elias, M.; Mühlen, G.S.; McKey, D.; Roa, A.C.; Tohme, J. Genetic diversity of traditional South American landraces of Cassava (Manihot esculenta Crantz): an analysis using microsatellites. Econ. Bot. 2004, 58, 242–256. [Google Scholar] [CrossRef]
- Wilson, W.M.; Dufour, D. Why bitter cassave? Productivity of ‘bitter’ and ‘sweet’ cassava in a Tukanoan Indian settlement in the northwest Amazon. Econ. Bot. 2002, 56, 49–57. [Google Scholar] [CrossRef]
- Gragson, T.L. The use of underground plant organs and its relation to habitat selection among the Pume Indians of Venezuela. Econ. Bot. 1997, 51, 377–384. [Google Scholar] [CrossRef]
- Duputié, A.; David, P.; Debain, C.; McKey, D. Natural hybridization between a clonally propagated crop, cassava (Manihot esculenta Crantz) and a wild relative in French Guiana. Mol. Ecol. 2007, 16, 3025–3038. [Google Scholar] [CrossRef]
- Olsen, K.M.; Schaal, B. Insights on the evolution of a vegetatively propagated crop species. Mol. Ecol. 2007, 16, 2838–2840. [Google Scholar] [CrossRef]
- Arroyo-Kalin, M.; Clement, C.R.; Fraser, J.A. Extensive to intensive, or first fruits and then roots? Bitter/sweet thoughts on agricultural intensification in pre-Columbian Amazonia. In Shifting Cultivation session, World Archaeological Congress 6; Dublin, Ireland, 2008. [Google Scholar]
- Beckerman, S. Swidden in Amazonia and the Amazon rim. In Comparative Farming Systems; Turner, B.L., Brush, S.B., Eds.; Guilford Press: New York, NY, USA, 1987; pp. 55–94. [Google Scholar]
- Lima, H.P. História das Caretas: A Tradição Borda Incisa na Amazônia Central. PhD thesis, Universidade de São Paulo, São Paulo, Brazil, 2008. [Google Scholar] [Green Version]
- Myers, T.P. Dark earth in the upper Amazon. In Amazonian Dark Earths: Explorations in Space and Time; Glaser, B., Woods W., I., Eds.; Springer: Berlin, Germany, 2004; pp. 67–94. [Google Scholar] [Green Version]
- DeBoer, W.R. The archaeological evidence for manioc cultivation: a cautionary note. Am. Antiq. 1975, 40, 419–433. [Google Scholar] [CrossRef]
- Perry, L. Starch analyses reveal the relationship between tool type and function: an example from the Orinoco valley of Venezuela. J. Archaeol. Sci. 2004, 31, 1069–1081. [Google Scholar] [CrossRef]
- Perry, L. Reassessing the traditional interpretation of “manioc” artifacts in the Orinoco valley of Venezuela. Lat. Am. Antiq. 2005, 16, 1–18. [Google Scholar] [CrossRef]
- Reichel-Dolmatoff, G. Monsú: un Sitio Arqueológico; Banco Popular: Bogotá, Colombia, 1985. [Google Scholar] [Green Version]
- Reichel-Dolmatoff, G. Arqueología de Colombia: un texto introductorio; Fundación Segunda Expedición Botánica: Bogotá, Colombia, 1997 (1986); p. 281. [Google Scholar] [Green Version]
- Sanoja, M. From Foraging to Food Production in Northeastern Venezuela and the Caribbean. In Foraging and Farming: The Evolution of Plant Exploitation; Harris, D.R., Hillman, G.C., Eds.; Unwin Hyman: London, UK, 1989; pp. 523–537. [Google Scholar] [Green Version]
- Angulo Valdés, C. Modos de vida en la prehistoria de la llanura atlántica de Colombia. In Prehistoria Sudamericana: Nuevas Perspectivas; Meggers, B.J., Ed.; Taraxacum: Washington, DC, USA, 1992; pp. 253–270. [Google Scholar] [Green Version]
- Williams, D. Prehistoric Guiana; Ian Randle Publishers: Kingston, Canada, 2003. [Google Scholar] [Green Version]
- Zucchi, A. Datos recientes para un nuevo modelo sobre la expansión de los Grupos Maipures del Norte. América Negra 1993, 6, 131–148. [Google Scholar]
- Zucchi, A. A new model of the northern Arawakan expansion. In Comparative Arawakan Histories: Rethinking Language Family and Culture Area in Amazonia; Hill, J.D., Santos-Granero, F., Eds.; University of Illinois Press: Chicago, IL, USA, 2002; pp. 199–225. [Google Scholar] [Green Version]
- Hornborg, A. Ethnogenesis, regional integration, and ecology in prehistoric Amazonia: toward a system perspective. Curr. Anthropol. 2005, 46, 589–620. [Google Scholar] [CrossRef]
- Morales, D. Chambira: una cultura de sabana árida en la Amazonia Peruana. Investigaciones Sociales 1995, 149–157. [Google Scholar] [Green Version]
- Roosevelt, A.C. Parmana. Prehistoric Maize and Manioc Susbistence along the Amazon and Orinoco; Academic Press: New York, NY, USA, 1980. [Google Scholar] [Green Version]
- Roosevelt, A.C. The excavations at Corozal, Venezuela: stratigraphy and ceramic seriation. Yale University Publications in Anthropology 1997, 83, 1–393. [Google Scholar]
- Roosevelt, A.C. Resource management in Amazonia before the conquest: beyond ethnographic projection. In Resource Management in Amazonia: Indigenous and Folk Strategies; Posey, D., Balée, W., Eds.; New York Botanical Garden: Bronx, NY, USA, 1989; Volume 7, pp. 3–62. [Google Scholar]
- Raymond, J.S. Ceremonialism in the early Formative of Ecuador. In El Mundo Ceremonial Andino; Millones, L., Onuki, Y., Eds.; National Museum of Ethnology: Osaka, Japan, 1993; Volume 37, pp. 25–43. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Arroyo-Kalin, M. The Amazonian Formative: Crop Domestication and Anthropogenic Soils. Diversity 2010, 2, 473-504. https://doi.org/10.3390/d2040473
Arroyo-Kalin M. The Amazonian Formative: Crop Domestication and Anthropogenic Soils. Diversity. 2010; 2(4):473-504. https://doi.org/10.3390/d2040473
Chicago/Turabian StyleArroyo-Kalin, Manuel. 2010. "The Amazonian Formative: Crop Domestication and Anthropogenic Soils" Diversity 2, no. 4: 473-504. https://doi.org/10.3390/d2040473
APA StyleArroyo-Kalin, M. (2010). The Amazonian Formative: Crop Domestication and Anthropogenic Soils. Diversity, 2(4), 473-504. https://doi.org/10.3390/d2040473