The Diversity of Bitter Manioc (Manihot Esculenta Crantz) Cultivation in a Whitewater Amazonian Landscape

Abstract

1. Introduction
2. Materials and Methods
2.1. Quantitative Methods

2.2. Selection of Locations and Individuals
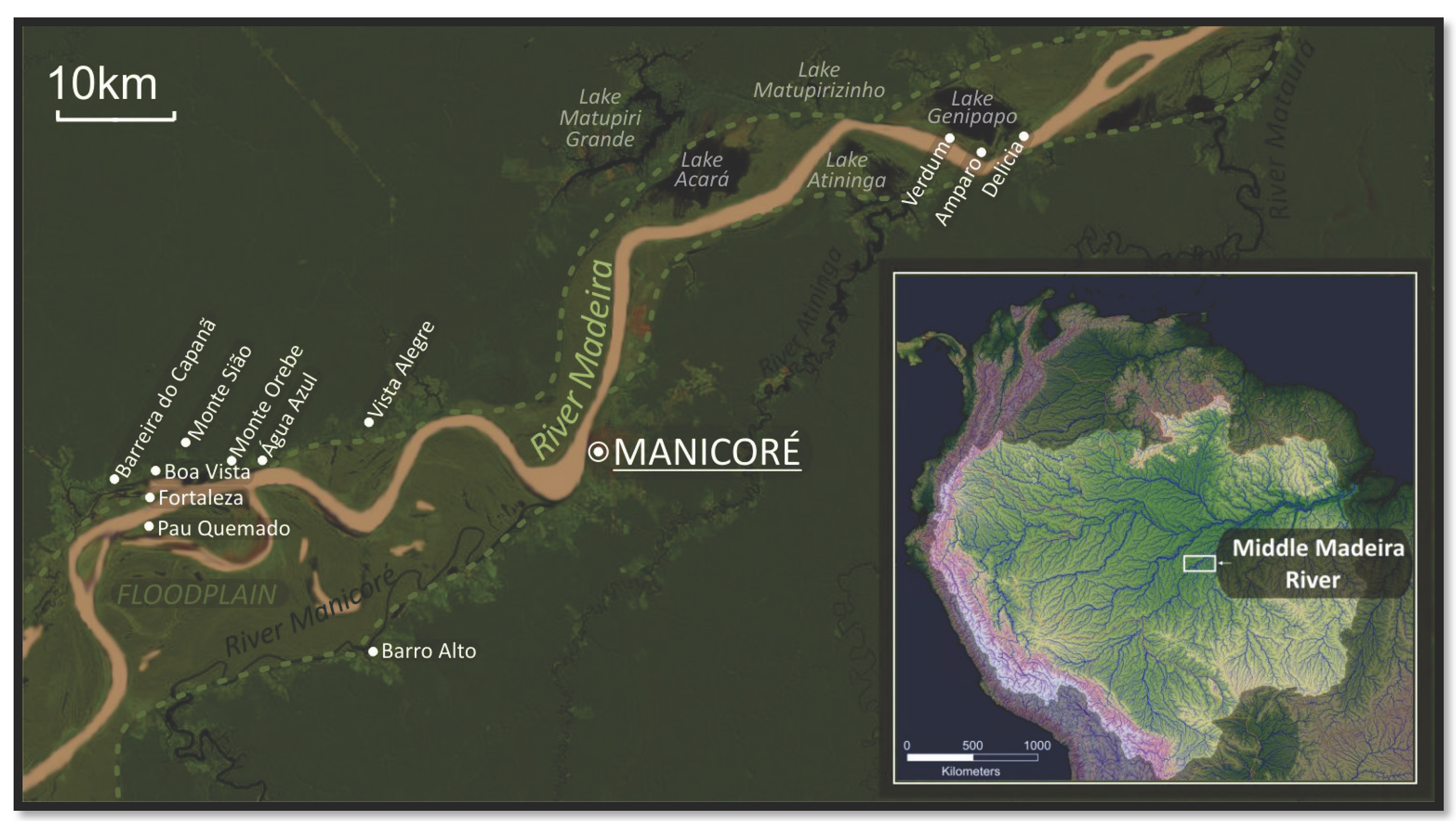

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Localities: | Strength Index | Ultisols | Oxisols | ADE | Floodplain |
|---|---|---|---|---|---|
| n= | Number of Fields | Number of Fields | Number of Fields | Number of Fields | |
| Barro Alto | 35 | 25 | 33 | 36 | |
| Barreira do Capanã | 24 | 12 | 11 | 17 | |
| Água Azul, Monte Orebe, Monte Sião | 9 | 8 | 8 | 8 | |
| Vista Alegre | 19 | 10 | 12 | 10 | |
| Água Azul floodplain | 22 | 24 | |||
| Lake Genipapo floodplain | 30 | 35 | |||
| Total | 139 | 55 | 64 | 71 | 59 |
3. Strong and Weak Bitter Manioc
| Characteristic | Weak Manioc
Low-Starch Fast-Maturing Low Starch Fast Yielding (LSFY) | Strong Manioc
High-Starch Slow-Maturing |
|---|---|---|
| Maturation | Faster | Slower |
| Starch production | Less (more water) | More (less water) |
| Resistance to rotting | Lower (6 months to 1.5 years) | Higher (up to 3 or 4 years in the ground) |
| Softening in water | Only in earlier stages | Softens even after years |
| Suitability to fallow age ( terra firme only) | Earlier successional stages | Later successional stages |
| Lengths of cropping-fallowing cycles | Shorter fallowing, shorter cropping | Longer fallowing, longer cropping |
| Soil associations | ADE / Floodplain | Oxisol / Ultisol |
4. The Diversity of Bitter Manioc Cultivation Systems on the Middle Madeira
4.1. Smaller Fields, Higher Yields
| Soil | n= | Mean Size (ha) | Std Dev | Std. Error | Median Size (ha) |
|---|---|---|---|---|---|
| Ultisol | 55 | 0.809 | 0.36 | 0.0485 | 1 |
| Oxisol | 64 | 1.012 | 0.455 | 0.0569 | 1 |
| ADE | 71 | 0.605 | 0.39 | 0.0462 | 0.5 |
| Floodplain | 59 | 0.631 | 0.326 | 0.0425 | 0.5 |
| Soil Type | n= | Farinha sacks per ha (Median) | Median Manioc Flour Production per ha | Mean sacks | SD sacks | SE sacks |
|---|---|---|---|---|---|---|
| Oxisol | 10 | 37.5 | 2,250 kg/ha | 81 | 7.2 | 22.8 |
| Ultisol | 6 | 35 | 2,100 kg/ha | 72 | 3.1 | 7.5 |
| ADE | 12 | 50 | 3,000 kg/ha | 107 | 9.8 | 34.1 |
| Floodplain | 6 | 50 | 3,000 kg/ha | 94 | 7.9 | 19.4 |

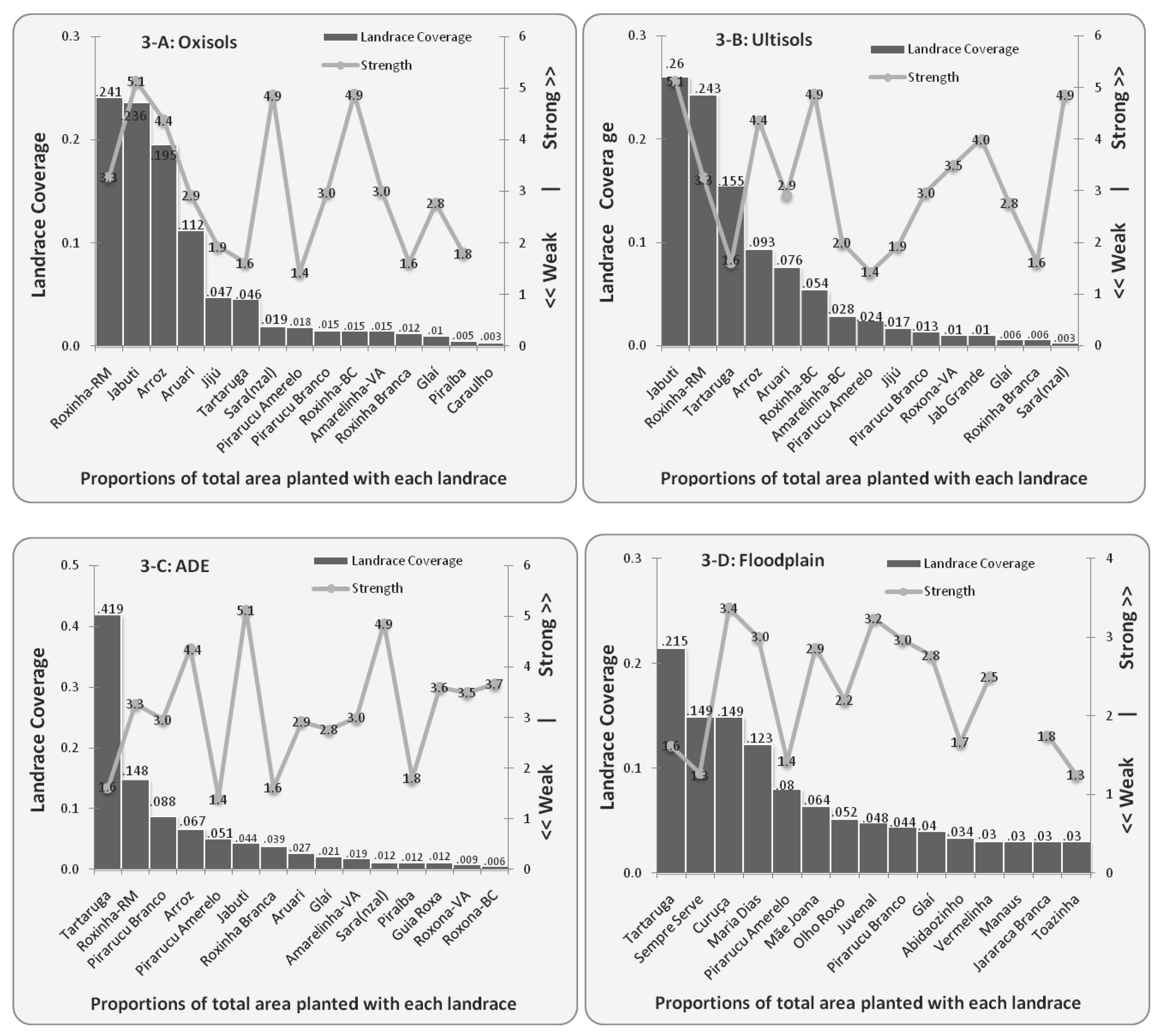
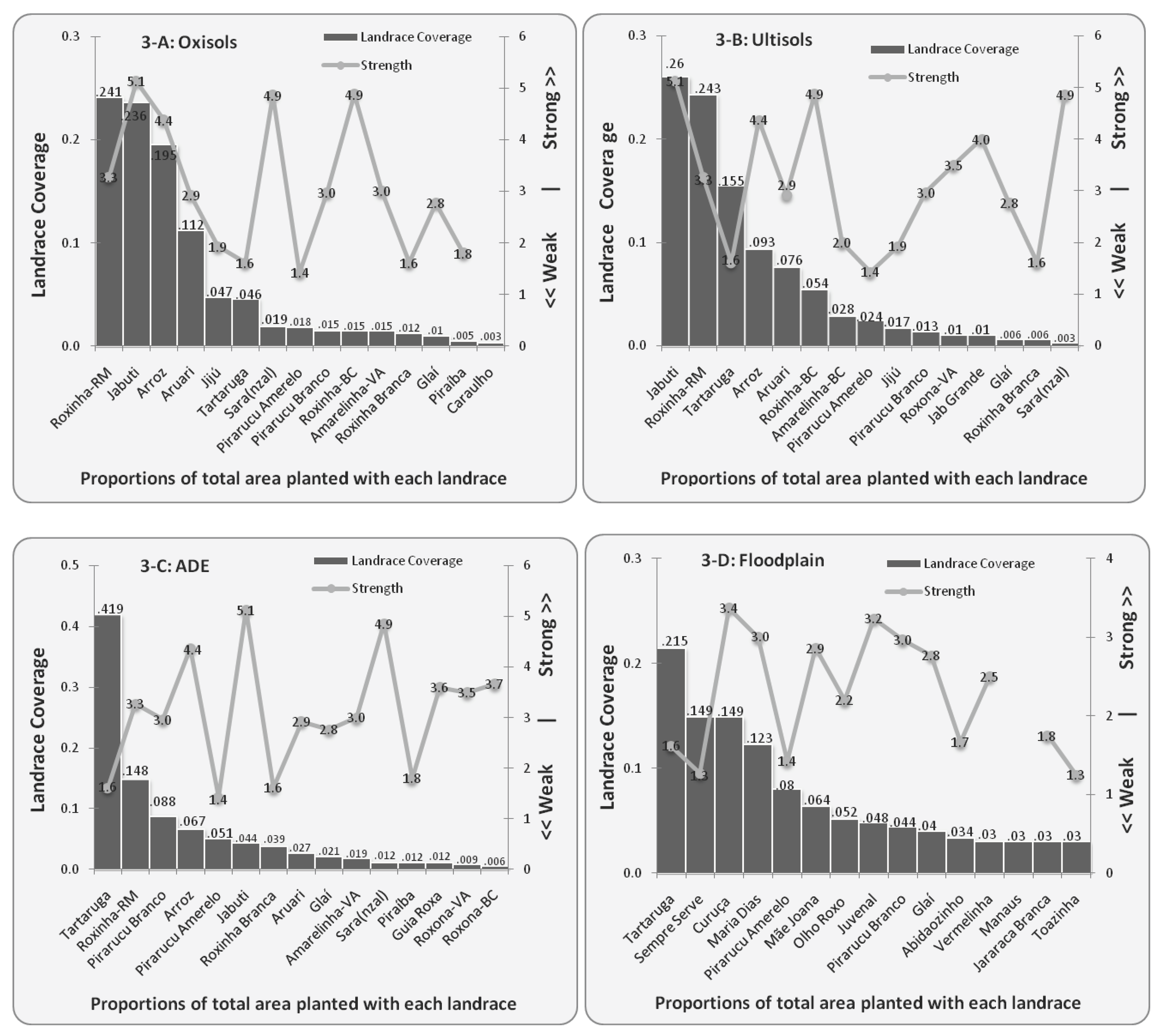
4.2. Diversity of Bitter Manioc Landraces in Different Ecological Contexts
| Strength | Landrace Coverage | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Landraces | Str | n | UL | n | OX | n | ADE | n | FP | n | Total | n |
| Tartaruga | 1.6 | 81 | 0.155 | 30 | 0.046 | 19 | 0.419 | 53 | 0.215 | 32 | 0.835 | 204 |
| Roxinha-RM | 3.2 | 29 | 0.243 | 28 | 0.241 | 35 | 0.148 | 18 | 0.632 | 149 | ||
| Jabuti | 5.1 | 47 | 0.260 | 24 | 0.236 | 34 | 0.044 | 15 | 0.540 | 159 | ||
| Arroz | 4.4 | 48 | 0.093 | 19 | 0.195 | 43 | 0.067 | 22 | 0.355 | 138 | ||
| Aruari | 2.9 | 27 | 0.076 | 20 | 0.112 | 27 | 0.027 | 9 | 0.215 | 70 | ||
| Pirarucu Amarelo | 1.4 | 21 | 0.024 | 6 | 0.018 | 7 | 0.051 | 10 | 0.08 | 10 | 0.173 | 49 |
| Pirarucu Branco | 3.0 | 33 | 0.013 | 7 | 0.015 | 8 | 0.088 | 14 | 0.044 | 9 | 0.160 | 38 |
| Sempre Serve | 1.3 | 26 | 0.149 | 26 | 0.149 | 26 | ||||||
| Curuça | 3.4 | 27 | 0.149 | 27 | 0.149 | 27 | ||||||
| Maria Dias | 3.0 | 3 | 0.123 | 4 | 0.123 | 12 | ||||||
| Glaí | 2.1 | 13 | 0.006 | 3 | 0.01 | 4 | 0.021 | 8 | 0.04 | 6 | 0.077 | 33 |
| Roxinha-BC | 4.9 | 8 | 0.054 | 9 | 0.015 | 6 | 0.003 | 2 | 0.072 | 39 | ||
| Jijú | 1.9 | 14 | 0.017 | 5 | 0.047 | 11 | 0.003 | 3 | 0.067 | 19 | ||
| Mãe Joana | 2.9 | 15 | 0.064 | 12 | 0.064 | 22 | ||||||
| Roxinha Branca | 1.6 | 13 | 0.006 | 5 | 0.012 | 5 | 0.039 | 7 | 0.057 | 17 | ||
| Olho Roxo | 2.2 | 5 | 0.052 | 9 | 0.052 | 9 | ||||||
| Juvenal | 3.2 | 13 | 0.048 | 12 | 0.048 | 20 | ||||||
| Amarelinha-VA | 3.0 | 2 | 0.002 | 2 | 0.015 | 4 | 0.019 | 2 | 0.036 | 20 | ||
| Sara(nzal) | 4.9 | 7 | 0.003 | 3 | 0.019 | 6 | 0.012 | 8 | 0.034 | 17 | ||
| Abidãozinho | 1.7 | 9 | 0.034 | 9 | 0.034 | 9 | ||||||
| Vermelinha | 2.5 | 2 | 0.03 | 3 | 0.030 | 3 | ||||||
| Manaus | -- | 0 | 0.03 | 1 | 0.030 | 1 | ||||||
| Jararaca Branca | 1.8 | 4 | 0.03 | 1 | 0.030 | 1 | ||||||
| Toazinha | 1.3 | 2 | 0.03 | 2 | 0.030 | 4 | ||||||
| Amarelinha-BC | 2.0 | 2 | 0.028 | 7 | 0.001 | 1 | 0 | 0 | 0.029 | 8 | ||
| Direitinha | 1.4 | 10 | 0.026 | 9 | 0.026 | 9 | ||||||
| Beleza | 3.5 | 4 | 0.023 | 5 | 0.023 | 5 | ||||||
| Japim | 2.0 | 1 | 0.02 | 1 | 0.020 | 1 | ||||||
| Roxona-VA | 3.5 | 2 | 0.010 | 2 | 0 | 0 | 0.009 | 3 | 0.019 | 11 | ||
| Piraiba | 1.8 | 5 | 0 | 0 | 0.005 | 3 | 0.012 | 3 | 0.017 | 8 | ||
| Guia Roxa | 3.6 | 5 | 0 | 0 | 0.001 | 1 | 0.012 | 2 | 0.013 | 3 | ||
| Jararaca Branca | 1.0 | 1 | 0.008 | 1 | 0.008 | 1 | ||||||
| Roxona-BC | 3.7 | 3 | 0 | 1 | 0 | 0 | 0.006 | 4 | 0.006 | 5 | ||
| Julio da Varzea | -- | 0 | 0 | 0 | 0 | 0 | 0.006 | 1 | 0.006 | 1 | ||
| Manaus | 3.0 | 1 | 0.002 | 2 | 0 | 0 | 0.003 | 3 | 0.005 | 7 | ||
| Caraulho | -- | 0 | 0 | 0 | 0.003 | 1 | 0.002 | 1 | 0.005 | 2 | ||
| Vermelão | 3.0 | 1 | 0.004 | 1 | 0.004 | 1 | ||||||
| Galhadinha | 4.0 | 1 | 0.003 | 1 | 0.003 | 1 | ||||||
| Amerelona | 2.00 | 1 | 0 | 0 | 0 | 0 | 0.003 | 1 | 0.003 | 1 | ||
| Expomani | -- | 0 | 0.002 | 1 | 0 | 0 | 0 | 0 | 0.002 | 1 | ||
| Manauense | 1.0 | 1 | 0.002 | 1 | 0.002 | 3 | ||||||
| Azolão | 8.0 | 1 | 0 | 0 | 0 | 1 | 0.001 | 1 | 0.001 | 2 | ||
| Pacu | 4.0 | 1 | 0 | 0 | 0 | 0 | 0.001 | 1 | 0.001 | 1 | ||
| Bonitinha-BA | -- | 0 | 0 | 0 | 0 | 0 | 0.001 | 2 | 0.001 | 4 | ||
| Bonitinha-BC | 3.0 | 3 | 0 | 0 | 0 | 1 | 0 | 1 | 0.001 | 4 | ||

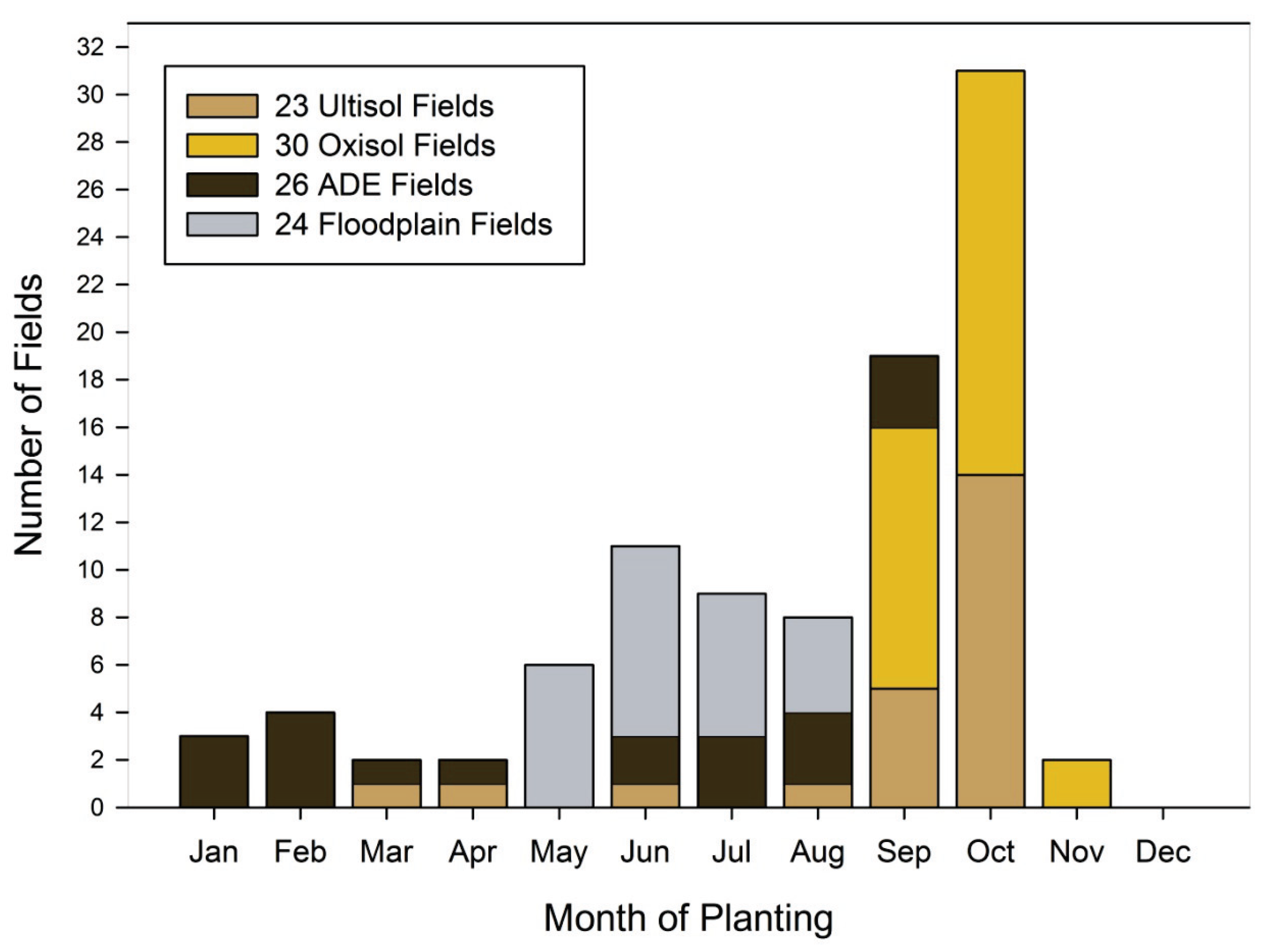
4.3. Fallow Periods and Seasonality in Different Ecological Contexts
| Soil | n= | Mean | Std. Dev | Std. Error | Median |
|---|---|---|---|---|---|
| Ultisol | 55 | 13.164 | 8.082 | 1.09 | 12 |
| Oxisol | 64 | 21.344 | 15.533 | 1.942 | 15 |
| ADE | 71 | 6.458 | 6.839 | 0.812 | 4 |
| Floodplain | 59 | 1.593 | 4.039 | 0.526 | 0 |


5. Conclusion: Are Ecological Limitations on Bitter Manioc Cultivation Real or Imagined?
Acknowledgements
References
- Heckenberger, M.J. Manioc agriculture and sedentism in Amazonia: the Upper Xingu example. Antiquity 1998, 72, 633–648. [Google Scholar]
- Oliver, J.R. The archaeology of forest foraging and agricultural production in Amazonia. In Unknown Amazon. Culture in Nature in Ancient Brazil, 1st ed.; McEwan, C., Barreto, C., Neves, E.G., Eds.; The British Museum Press: London, UK, 2001; pp. 50–85. [Google Scholar]
- Lathrap, D.W. The Upper Amazon; Thames & Hudson: Southampton, UK, 1970. [Google Scholar]
- Clement, C.R.; Cristo-Araújo, M.; d’Eeckenbrugge, G.C.; Pereira, A.A.; Picanço-Rodrigues, D. Origin and Domestication of Native Amazonian Crops. Diversity 2010, 2, 72–106. [Google Scholar] [CrossRef]
- Arroyo-Kalin, M. The Amazonian Formative: Crop Domestication and Anthropogenic Soils. Diversity 2010, 2, 473–504. [Google Scholar] [CrossRef]
- Meggers, B.J. Amazonia. Man and Culture in a Counterfeit Paradise. Revised Edition; Smithsonian Institution Press: Washington, DC, USA, 1996. [Google Scholar]
- McKey, D.; Beckerman, S. Chemical ecology, plant evolution, and the evolution of traditional manioc cultivation systems. In Tropical Forests, People and Food. Biocultural Interactions and Applications to Development; Hladik, C.M., Hladik, A., Linares, O.F., Pagezy, H., Semple, A., Hadley, M., Eds.; UNESCO, Parthenon: Paris, France, 1993; pp. 83–112. [Google Scholar]
- McKey, D.; Cavagnaro, T.R.; Cliff, J.; Gleadow, R. Chemical ecology in coupled human and natural systems: people, manioc, multitrophic interactions and global change. Chemoecology 2010, in press. [Google Scholar]
- Adams, C.; Murrieta, R.; Siqueira, A.; Neves, W.; Sanches, R. Bread of the Land: The Invisibility of Manioc in the Amazon. In Amazon Peasant Societies in a Changing Environment: Political Ecology, Invisibility and Modernity in the Rainforest; Adams, A., Murrieta, R., Neves, W., Harris, M., Eds.; Springer: Berlin, Germany, 2009; pp. 281–306. [Google Scholar]
- Emperaire, L. Elementos de discussão sobre a conservação da agrobiodiversidade: o exemplo da mandioca (Manihot esculenta Crantz) na Amazônia brasileira. In Biodiversidade da Amazônia; Capobianco, J.P., Ed.; Estação Liberdade-ISA: São Paulo, Brazil, 2001; pp. 225–234. [Google Scholar]
- Olsen, K.M. Population history of Manihot esculenta (Euphorbiaceae) inferred from nuclear DNA sequences. Molecular Ecology 2002, 11, 901–911. [Google Scholar]
- Olsen, K.M.; Schaal, B.A. DNA Sequence data and inferences on cassava’s origin of domestication. In Documenting Domestication: New Genetic and Archaeological Paradigms; Zeder, M.A., Bradley, D.G., Emshwiller, E., Smith, B.D., Eds.; University of California Press: Berkeley, CA, USA, 2006; pp. 123–132. [Google Scholar]
- Steward, J.H. Handbook of South American Indians. Volume 3: The Tropical Forest Tribes; Bulletin 143, Bureau of American Ethnology, Smithsonian Institution: Washington, DC, USA, 1948. [Google Scholar]
- Steward, J.H.; Faron, L.C. Native Peoples of South America; McGraw-Hill: New York, NY, USA, 1959. [Google Scholar]
- Hames, R.B.; Vickers, W.T. Introduction. In Adaptive Responses of Native Amazonians; Hames, R.B., Vickers, W.T., Eds.; Academic Press: New York, NY, USA, 1983; pp. 1–26. [Google Scholar] [Green Version]
- Meggers, B.J. Environmental Limitation on the Development of Culture. Amer. Anthropol. 1954, 56, 801–824. [Google Scholar]
- Meggers, B.J.; Evans, E. Archaeological Investigations at the Mouth of the Amazon; Smithsonian Institution, Bureau of American Ethnology, Bulletin 167: Washington, DC, USA, 1957. [Google Scholar]
- Carneiro, R.L. The cultivation of manioc among the Kuikuru of the Upper Xingu. In Adaptive Responses of Native Amazonians; Hames, R.B., Vickers, W.T., Eds.; Academic Press: New York, NY, USA, 1983; pp. 65–112. [Google Scholar]
- Heckenberger, M.J. Biocultural Diversity in the Southern Amazon. Diversity 2010, 2, 1–16. [Google Scholar] [CrossRef]
- Heckenberger, M.J. The Ecology of Power: Culture, Place and Personhood in the Southern Amazon AD 1000–2000; Routledge: London, UK, 2005. [Google Scholar]
- Heckenberger, M.J.; Kuikuro, A.; Kuikuro, U.T.; Russell, J.C.; Schmidt, M.; Fausto, C.; Franchetto, B. Amazonia 1492: Pristine forest or cultural parkland? Science 2003, 301, 1710–1714. [Google Scholar] [CrossRef]
- Heckenberger, M.J.; Russell, J.C.; Fausto, C.; Toney, J.R.; Schmidt, M.J.; Pereira, E.; Franchetto, B.; Kuikuro, A. Pre-Columbian urbanism, anthropogenic landscapes, and the future of the Amazon. Science 2008, 321, 1214–1217. [Google Scholar] [CrossRef]
- Heckenberger, M.J.; Russell, J.C.; Toney, J.R.; Schmidt, M.J. The legacy of cultural landscapes in the Brazilian Amazon: implications for biodiversity. Phil. Trans. Roy. Soc. B-Biol. Sci. 2007, 362, 197–208. [Google Scholar] [CrossRef]
- Junk, W.J.; Furch, K. The physical and chemical properties of Amazonian waters and their relationships with the biota. In Key Environments: Amazonia; Prance, G.T., Lovejoy, T.E., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 3–17. [Google Scholar]
- Sioli, H. The Amazon and its main affluents: hydrography, morphology of the river courses, and river types. In The Amazon: Limnology and Landscape Ecology of a Mighty Tropical River and Its Basin; Sioli, H., Ed.; Junk Publishers: Dordrecht, Germany, 1984; pp. 127–165. [Google Scholar]
- Neves, E.G. Ecology, ceramic chronology and distribution, long-term history, and political change in the Amazonian floodplain. In Handbook of South American Archaeology; Silverman, H., Isbell, W.H., Eds.; Springer: New York, NY, USA, 2008; pp. 359–403. [Google Scholar]
- Roosevelt, A.C. The development of prehistoric complex societies: Amazonia, a tropical forest. In Complex Polities in the Ancient Tropical World; Bacus, E.A., Lucero, L.J., Eds.; American Anthropological Association: Arlington, VA, USA, 1999; pp. 13–33. [Google Scholar]
- Roosevelt, A.C. The lower Amazon: a dynamic human habitat. In Imperfect Balance: Landscape Transformations in the Precolumbian Americas, 1st ed.; Lentz, D.L., Ed.; Columbia University Press: New York, NY, USA, 2000; pp. 455–491. [Google Scholar]
- Heckenberger, M.J.; Petersen, J.B.; Neves, E.G. Village size and permanence in Amazonia: two archaeological examples from Brazil. Latin American Antiquity 1999, 10, 353–376. [Google Scholar] [CrossRef]
- Neves, E.G.; Arroyo-Kalin, M.; Bartone, R.N.; Castro, M.W.M.; Costa, F.W.S.; Lima, H.P.; Lima, M.N.; Moraes, C.P.; Py-Daniel, A.R.; Rebellato, L.; Silva, C.A.; Tamanaha, E.K.; Woods, W.I. Site reoccupation and population growth in the late first millenium AD in the Central Amazon. Unpublished Manuscript.
- Neves, E.G.; Petersen, J.B. The political economy of pre-Columbian Amerindians: Landscape transformations in Central Amazonia. In Time and Complexity in Historical Ecology: Studies in the Neotropical Lowlands; Balée, W., Erickson, C.L., Eds.; Columbia University Press: New York, NY, USA, 2006; pp. 279–309. [Google Scholar]
- Petersen, J.B.; Neves, E.G.; Heckenberger, M.J. Gift from the past: terra preta and prehistoric occupation in Amazonia. In Unknown Amazon. Culture in Nature in Ancient Brazil, 1st ed.; McEwan, C., Barreto, C., Neves, E.G., Eds.; The British Museum Press: London, UK, 2001; pp. 86–107. [Google Scholar]
- Rebellato, L.; Woods, W.I.; Neves, E.G. Pre-Columbian settlement dynamics in the Central Amazon. In Amazonian Dark Earths: Wim Sombroek's Vision; Woods, W.I., Teixeira, W.G., Lehmann, J., Steiner, C., WinklerPrins, A.M.G.A., Rebellato, L., Eds.; Springer: Berlin, Germany, 2009; pp. 15–31. [Google Scholar]
- Roosevelt, A.C. Parmana: Prehistoric Maize and Manioc Subsistence along the Amazon and Orinoco; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Ohly, J.; Junk, W. Multiple uses of central Amazon floodplains: reconciling ecological conditions, requirements for environmental protection, and socioeconomic needs. In Varzea: Diversity, development and conservation of Amazonias's whitewater floodplains; Padoch, C., Ayres, J.M., Pinedo-Vasquez, M., Henderson, A., Eds.; New York Botanical Garden: New York, NY, USA, 1999; pp. 283–299. [Google Scholar]
- Moran, E. Models of Native and Folk Adaptation in the Amazon. In Advances in Economic Botany; Posey, D., Balée, W., Eds.; New York Botanical Garden: Bronx, New York, NY, USA, 1989; pp. 22–29. [Google Scholar]
- Boster, J.S. Classification, cultivation and selection of Aguaruna cultivars of Manihot esculenta (Euphorbiaceae). Advan. Econ. Bot. 1984, 1, 34–47. [Google Scholar]
- Chernela, J.M. Os cultivares de mandioca na área do Uaupés (Tukano). In Suma Etnológica Brasileira, 2nd ed.; Ribeiro, D., Ed.; Vozes: Petrópolis, Brazil, 1987; pp. 151–158. [Google Scholar]
- Elias, M.; Rival, L.; McKey, D. Perception and management of cassava (Manihot esculenta Crantz) diversity among Makushi Amerindians of Guyana (South America). J. Ethnobiology 2000, 20, 239–265. [Google Scholar]
- Wilson, W.M.; Dufour, D.L. Why “bitter” cassava? Productivity of “bitter” and “sweet” cassava in a Tukanoan Indian settlement in the northwest Amazon. Econo. Bot. 2002, 56, 49–57. [Google Scholar] [CrossRef]
- Carneiro, R.L. Slash-and-burn cultivation among the Kuikuru and its implications for Cultural Development in the Amazon basin. In The Evolution of Horticultural Systems in Native South America, Causes and Consequences. Anthropológica Supplement Publication No. 2; Wilbert, J., Ed.; Editorial Sucre: Caracas, Venezuela, 1961; pp. 122–132. [Google Scholar]
- Carneiro, R.L. Slash-and-burn agriculture: a closer look at its implications for settlement patterns. In Men and Cultures; Wallace, A.F.C., Ed.; University of Pennsylvania Press: Philadelphia, PA, USA, 1961; pp. 229–234. [Google Scholar]
- Heckler, S.; Zent, S. Piaroa Manioc Varietals: Hyperdiversity or Social Currency? Human Ecology 2008, 36, 679–697. [Google Scholar] [CrossRef]
- Wilson, W.M.; Dufour, D.L. Ethnobotanical evidence for cultivar selection among the Tukanoans: Manioc (Manihot esculenta Crantz) in the Northwest Amazon. Culture Agr. 2006, 28, 122–130. [Google Scholar] [CrossRef]
- Balée, W. People of the fallow: a historical ecology of foraging in lowland South America. In Conservation of Neotropical Forests: Working from Traditional Resource Use; Redford, K.H., Padoch, C., Eds.; Columbia University Press: New York, NY, USA, 1992; pp. 35–46. [Google Scholar]
- Rival, L. Trekking Through History: The Huaorani of Amazonian Ecuador; Columbia University Press: New York, NY, USA, 2002. [Google Scholar]
- Fraser, J.A. Amazonian Dark Earths and Caboclo Subsistence on the Middle Madeira River, Brazil. Unpublished Dphil Thesis, University of Sussex, Brighton, UK, 2009. [Google Scholar]
- Fraser, J.A.; Cardoso, T.; Junqueira, A.B.; Falcão, N.; Clement, C.R. Historical ecology and dark earths in whitewater and blackwater landscapes: comparing the Middle Madeira and Lower Negro rivers. In Amazonian Dark Earths: Wim Sombroek's Vision; Woods, W.I., Teixeira, W.G., Lehmann, J., Steiner, C., WinklerPrins, A.M.G.A., Rebellato, L., Eds.; Springer: Berlin, Germany, 2009; pp. 229–264. [Google Scholar]
- Amazonian Dark Earths. Origins, Properties and Management; Lehmann, J.; Kern, D.C.; Glaser, B.; Woods, W. (Eds.) Kluwer Academic Publishers: Dordrecht, Germany, 2003.
- Amazonian Dark Earths: Explorations in Space and Time; Glaser, B.; Woods, W.I. (Eds.) Springer: Berlin, Germany, 2004.
- Amazonian Dark Earths: Wim Sombroek's Vision; Woods, W.I.; Teixeira, W.G.; Lehmann, J.; Steiner, C.; WinklerPrins, A.M.G.A.; Rebellato, L. (Eds.) Springer: Berlin, Germany, 2009.
- Neves, E.G.; Petersen, J.; Bartone, R.; da Silva, C.A. Historical and socio-cultural origins of Amazonian Dark Earths. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers: Dordrecht, Germany, 2003; pp. 29–50. [Google Scholar]
- Neves, E.G. El Formativo que nunca terminó: La larga historia de estabilidad en las ocupaciones humanas de la Amazonía central. Boletín de Arqueología PUCP 2007, 11, 117–142. [Google Scholar]
- Hecht, S.B. Indigenous soil management and the creation of Amazonian Dark Earths: implications of Kayapó practices. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers: Dordrecht, 2003; pp. 355–372. [Google Scholar]
- Schmidt, M.J. Historical Landscapes in the Neotropics: A Model for Pre-Historical Anthrosol (Terra Preta) Formation in the Upper Xingu. In Conference paper at International Encounter of Amazonian Archaeology (EIAA), Belem, Brazil; 2008. [Google Scholar]
- WinklerPrins, A.M.G.A. Sweep and Char and the Creation of Amazonian Dark Earths in Homegardens. In Amazonian Dark Earths: Wim Sombroek's Vision; Woods, W.I., Teixeira, W.G., Lehmann, J., Steiner, C., WinklerPrins, A.M.G.A., Rebellato, L., Eds.; Springer: Dordrecht, Germany, 2009; pp. 205–210. [Google Scholar]
- Steiner, C.; Teixeira, W.G.; Woods, W.I.; Zech, W. Indigenous Knowledge about Terra Preta Formation. In Amazonian Dark Earths: Wim Sombroek's Vision; Woods, W.I., Teixeira, W.G., Lehmann, J., Steiner, C., WinklerPrins, A.M.G.A., Rebellato, L., Eds.; Springer: Berlin, Germany, 2009; pp. 193–204. [Google Scholar]
- Schmidt, M.J. Reconstructing Tropical Nature: Prehistoric and Modern Anthrosols (Terra Preta) in the Amazon Rainforest, Upper Xingu River, Brazil. Unpublished PhD Thesis, University of Florida, 2010. [Google Scholar]
- Lehmann, J.; Joseph, S. Biochar for Environmental Management: Science and Technology; Earthscan: London, UK, 2009. [Google Scholar]
- Glaser, B.; Guggenberger, G.; Zech, W.; Ruivo, M.d.L. Soil Organic Mater Stability in Amazonian Dark Earths. In Amazonian Dark Earths. Origins, Properties and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers: Dordrecht, 2003; pp. 141–158. [Google Scholar]
- Lehmann, J.; da Silva, J.P.; Steiner, C.; Nehls, T.; Zech, W.; Glaser, B. Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: fertilizer, manure and charcoal amendments. Plant Soil 2003, 343–357. [Google Scholar]
- Lehmann, J.; Kern, D.C.; German, L.; McCann, J.; Martins, G.C.; Moreira, A. Soil fertility and production potential. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W.I., Eds.; Kluwer: Dortrecht, 2003; pp. 105–124. [Google Scholar]
- Kämpf, N.; Woods, W.I.; Sombroek, W.G.; Kern, D.C.; Cunha, T.J.F. Classification of Amazonian Dark Earths and other ancient anthropic soils. In Amazonian Dark Earths. Origins, Properties and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers Press: Dordrecht, 2003; pp. 77–102. [Google Scholar]
- Woods, W.I.; McCann, J.M. The anthropogenic origin and persistence of Amazonian dark earths. Yearbook, Conference of Latin Americanist Geographers 1999, 25, 7–14. [Google Scholar]
- Denevan, W.M. Cultivated landscapes of Native Amazonia and the Andes; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Denevan, W.M. A bluff model of riverine settlement in prehistoric Amazonia. Ann. Assn. Amer. Geogr. 1996, 86, 654–681. [Google Scholar] [CrossRef]
- Denevan, W.M. Semi-intensive pre-European cultivation and the origins of Anthropogenic Dark Earths in Amazonia. In Amazonian Dark Earths: Explorations in Space and Time; Glaser, B., Woods, W.I., Eds.; Springer: Berlin, Germany, 2004; pp. 135–143. [Google Scholar]
- Denevan, W.M. Pre-European forest cultivation in Amazonia. In Time and Complexity in Historical Ecology: Studies in the Neotropical Lowlands; Balée, W., Erickson, C., Eds.; Columbia University Press: New York, NY, USA, 2006; pp. 153–163. [Google Scholar]
- Stahl, P.W. Paradigms in Paradise: revising standard Amazonian Prehistory. Rev. Archaeolo. 2002, 23, 39–51. [Google Scholar]
- Schmidt, M.J. Farming and Patterns of Agrobiodiversity on the Amazon Floodplain in the Vicinity of Mamiraua, Amazonas, Brazil. Unpublished Master’s Thesis, University of Florida, FL, USA, 2003. [Google Scholar]
- Pereira, K.J.C. Agricultura tradicional e manejo da agrobiodiversidade na Amazônia Central: Um estudo de caso nos roçados de mandioca das Reservas de Desenvolvimento Sustentável Amanã e Mamirauá, Amazonas. Unpublished PhD Thesis, University of São Paulo, São Paulo, Brazil, 2008. [Google Scholar]
- German, L.A. The Dynamics of Terra Preta: An Integrated Study of Human-Environmental Interaction in a Nutrient-Poor Amazonian Ecosystem. Unpublished PhD Thesis, University of Georgia, Athens, GA, USA, 2001. [Google Scholar]
- German, L.A. Ethnoscientific understandings of Amazonian Dark Earths. In Amazonian Dark Earths: Origin, Properties and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W.I., Eds.; Kluwer Academic Publishers: Dordrecht, 2003; pp. 179–201. [Google Scholar]
- German, L.A. Historical contingencies in the coevolution of environment and livelihood: contributions to the debate on Amazonian Black Earth. Geoderma 2003, 111, 307–331. [Google Scholar] [CrossRef]
- German, L.A. Ecological Praxis and Blackwater Ecosystems: A Case Study from the Brazilian Amazon. Human Ecology 2004, 32, 653–683. [Google Scholar] [CrossRef]
- Hiraoka, M.; Yamamoto, S.; Matsumoto, E.; Nakamura, S.; Falesi, I.; Baena, A. Contemporary use and management of Amazonian Dark Earths. In Amazonian Dark Earths: Origin, Properties, and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer Academic Publishers: Dordrecht, 2003; pp. 387–406. [Google Scholar]
- McCann, J.M. Subsidy from culture: anthropogenic soils and vegetation in Tapajônia, Brazilian Amazonia. Unpublished PhD Thesis, University of Wisconsin-Madison, WI, USA, 2004. [Google Scholar]
- Sillitoe, P. Ethnobiology and applied anthropology: rapprochement of the academic with the practical. J. Roy. Anthropol. Inst. 2006, N.S., 119–142. [Google Scholar] [CrossRef]
- Adams, C.; Murrieta, R.; Neves, W.; Harris, M. Amazon Peasant Societies in a Changing Environment: Political Ecology, Invisibility and Modernity in the Rainforest; Springer: New York, NY, USA, 2009; p. 360. [Google Scholar]
- Smith, N.J.H. The Amazon River Forest: A Natural History of Plants, Animals, and People; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Fraser, J.A.; Clement, C.R. Dark Earths and Manioc Cultivation in Central Amazonia: A Window on Pre-Colombian Agricultural Systems? Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas 2008, 3, 175–194. [Google Scholar] [CrossRef]
© 2010 by the author; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fraser, J.A. The Diversity of Bitter Manioc (Manihot Esculenta Crantz) Cultivation in a Whitewater Amazonian Landscape. Diversity 2010, 2, 586-609. https://doi.org/10.3390/d2040586
Fraser JA. The Diversity of Bitter Manioc (Manihot Esculenta Crantz) Cultivation in a Whitewater Amazonian Landscape. Diversity. 2010; 2(4):586-609. https://doi.org/10.3390/d2040586
Chicago/Turabian StyleFraser, James A. 2010. "The Diversity of Bitter Manioc (Manihot Esculenta Crantz) Cultivation in a Whitewater Amazonian Landscape" Diversity 2, no. 4: 586-609. https://doi.org/10.3390/d2040586
APA StyleFraser, J. A. (2010). The Diversity of Bitter Manioc (Manihot Esculenta Crantz) Cultivation in a Whitewater Amazonian Landscape. Diversity, 2(4), 586-609. https://doi.org/10.3390/d2040586