
A New Species of Eocene Whitefly—Gregorites michalskii sp. nov. (Hemiptera, Sternorrhyncha, Aleyrodidae) from Baltic Amber


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Geological Setting
2.2. Morphology and Documentation
3. Results
Systematic Palaeontology
- Order: Hemiptera Linnaeus, 1758 [24]
- Suborder: Sternorrhyncha Amyot et Audinet-Serville, 1843 [25]
- Infraorder: Aleyrodomorpha Chou, 1963 [26]
- Family: Aleyrodidae Westwood, 1840 [6]
- Subfamily: Aleurodicinae Quaintance et Baker, 1913 [8]
- Genus: Gregorites Drohojowska et Szwedo, 2024 [12]
- Type species: Gregorites aegiri Drohojowska et Szwedo, 2024; by original designation
- Composition: Gregorites aegiri Drohojowska et Szwedo, 2024; G. bergelmiri Drohojowska et Szwedo, 2024; G. skadii Drohojowska et Szwedo, 2024; G. bestlae Drohojowska et Szwedo, 2024; G. thrymi Drohojowska et Szwedo, 2024; G. halogii Drohojowska et Szwedo, 2024; G. ymiri Drohojowska et Szwedo, 2024 [12]; and G. michalskii sp. nov.
- Gregorites michalskii sp. nov., Drohojowska et Szwedo
- LSID: urn:lsid:zoobank.org:act:CB63DC80-5ACC-4990-B65F-212456458D6D
- Etymology. The specific epithet was given in honour to Mr. Artur Michalski (Wrocław, Poland), an amber collector and an excellent photographer of amber inclusions, who kindly donated the specimen for research.
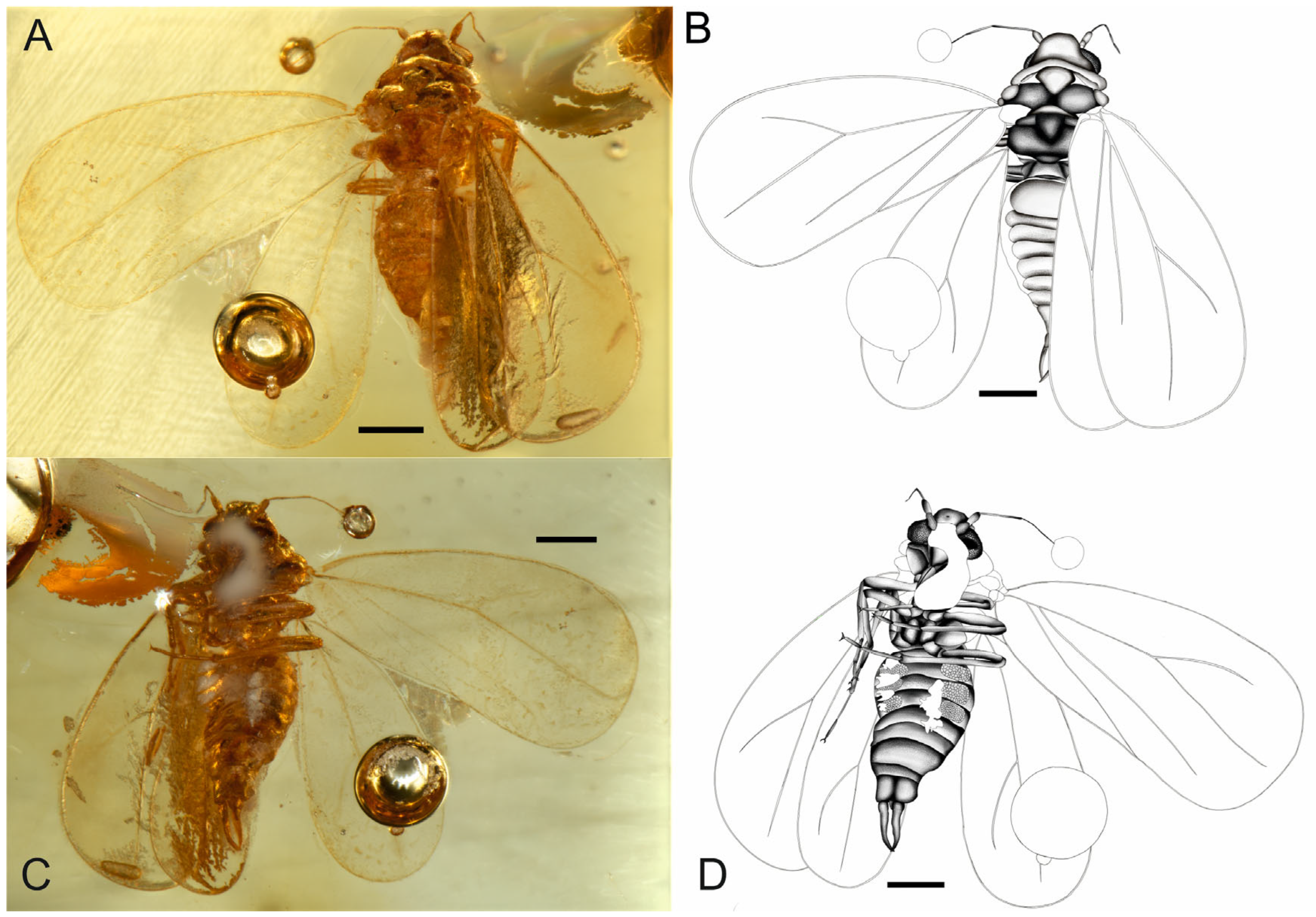
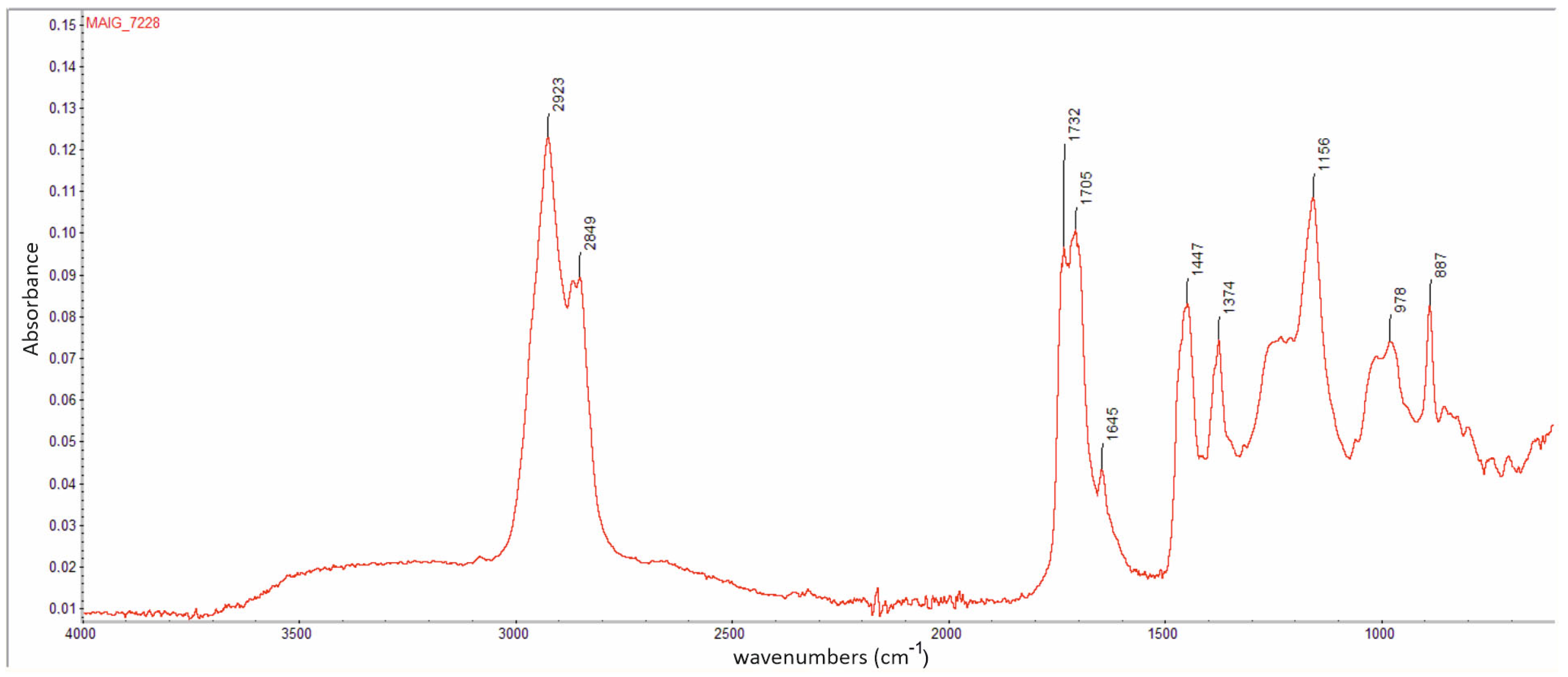
- Holotype. Male. It was part of the Artur Michalski collection and was deposited in the collection of the Museum of Amber Inclusions of the University of Gdańsk, MAIG 7228. It is a small piece of amber measuring 21 mm × 14 mm × 3 mm, and is approximately oval in shape. The specimen is preserved and in good condition, and the dorsal and ventral sides of the inclusion are generally well preserved; the anteroventral side is partly covered by a milky veil (milky coating emulsion, ‘Verlumung’). Gas bubbles are partly obstructing the hind wing and tip of the antenna. Biosyninclusions: Acari (larva), stellate hairs; other inclusions: gas bubbles and mineral intrusions between layers of resin.
- Diagnosis. The fore wing vein CuP (claval vein) reaches half of the fore wing length (longer than half of the fore wing in G. aegiri, shorter in G. ymiri, G. hallogi and G. skadii). The metatibia lacks a row of setae forming a comb (the metatibial comb is present in other species of the genus, except G. bestlae). There are long claspers that are longer than the pygofer, distinctly hooked at the apex and crossed apically. The total length of the body is the longest among the known species of the genus.
- Description. The total length is ca. 1.48 mm, including the claspers. The head includes compound eyes and is narrower than the pronotum, with a vertex trapezoid, convex disc, a slightly arcuate anterior margin, an acutely concave posterior margin and lateral margins that diverge posteriad. The compound eyes are easily visible in the dorsal view, with ommatidia of similar size that are arranged in rows. The lateral ocelli are easily visible in the dorsal view, and the anteriad is half of the vertex length and is placed above the compound eyes. The vertex smoothly shifts to the frons, and the frons has a median concavity but lacks a median ocellus. The postclypeus is concave, with an enlarged and protruding anteclypeus and a triangular and small clypellus. Narrow loral plates and narrow genae are present. The compound eyes are not divided, and the ommatidia are of a similar size and are arranged in regular rows. The antennal fovea margins are slightly elevated. Antenna with seven antennomeres, a cylindrical scapus (first antennomere) that is about twice as long as it is wide, an elongated and barrel-shaped pedicel (second antennomere) and a slightly widening apicad are present. The third antennomere is the longest, and the fourth and seventh antennomeres are cylindrical and of a similar length. The apical one is a tapering apicad (Figure 3A,B). The base of the rostrum is thick and easily visible, with two segments of a visible labium that is relatively short and an apex that reaches the tips of the procoxae.

4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drohojowska, J.; Szwedo, J.; Żyła, D.; Huang, D.-Y.; Müller, P. Fossils reshape the Sternorrhyncha evolutionary tree (Insecta, Hemiptera). Sci. Rep. 2020, 10, 11390. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, D.A. (Ed.) The Complete Insect: Anatomy, Physiology, Evolution, and Ecology; Princeton University Press: Princeton, MA, USA, 2023; 368p. [Google Scholar] [CrossRef]
- Szwedo, J. The unity, diversity and conformity of bugs (Hemiptera) through time. Earth Environ. Sci. Trans. R. Soc. Edinb. 2018, 107, 109–128. [Google Scholar] [CrossRef]
- McGavin, G.; Davranoglou, L.-R. Essential Entomology, 2nd ed.; Oxford University Press: Oxford, UK, 2023. [Google Scholar]
- Drohojowska, J.; Wegierek, P.; Evans, G.A.; Huang, D.Y. Are contemporary whiteflies living fossils? Morphology and systematic status of the oldest representatives of the Middle-late Jurassic Aleyrodomorpha (Sternorrhyncha, Hemiptera) from Daohugou. Palaeoentomology 2019, 2, 171–182. [Google Scholar] [CrossRef]
- Westwood, J.O. An introduction to the Modern Classification of Insects Founded on the Natural Habits and Corresponding Organization of Different Families; Longman, Orme, Brown and Green: London, UK, 1840; Volume 2, pp. i–xi+1–158. [Google Scholar]
- Shcherbakov, D.E. The most primitive whiteflies (Hemiptera; Aleyrodidae; Bernaeinae subfam. nov.) from the Mesozoic of Asia and Burmese amber, with an overview of Burmese amber hemipterans. Bull. Br. Mus. Nat. Hist. (Geol.) 2000, 56, 29–37. [Google Scholar]
- Quaintance, A.L.; Baker, A. Classification of the Aleyrodidae. Part 1; US Government Printing Office: Washington, DC, USA, 1913; Volume 27, pp. 1–93.
- Enderlein, G. Udamoselis, eine neue Aleurodiden-Gattung. Zool. Anz. 1909, 34, 230–233. [Google Scholar]
- Drohojowska, J.; Szwedo, J. Whiteflies (Hemiptera: Sternorrhyncha: Aleyrodidae) from the Lowermost Eocene Oise amber. Zootaxa 2013, 3636, 319–347. [Google Scholar] [CrossRef] [PubMed]
- Breithaupt, A. Kurze Charakteristik des Mineral-Systems; Selbstverlag [Self Published]: Freiberg, Germany, 1820; pp. i–xxiv+1–78. [Google Scholar]
- Drohojowska, J.; Śladowska, J.; Szwedo, J. First whiteflies from the Eocene amber of Denmark. Acta Palaeontol. Pol. 2024, 69, 649–676. [Google Scholar] [CrossRef]
- Drohojowska, J.; Zmarzły, M.; Szwedo, J. The discovery of a fossil whitefly from Lower Lusatia (Germany) presents a challenge to current ideas about Baltic amber. Sci. Rep. 2024, 14, 23102. [Google Scholar] [CrossRef] [PubMed]
- Drohojowska, J.; Bogusiak, W.; Kurkina, S.; Szwedo, J. A third aleurodicine from the Eocene Baltic amber—Eogroehnia carsteni nov. gen., nov. sp. (Hemiptera, Sternorrhyncha, Aleyrodidae). Geobios 2025, 88–89, 71–76. [Google Scholar] [CrossRef]
- Wolfe, A.P.; Tappert, R.; Muehlenbachs, K.; Boudreau, M.; McKellar, R.C.; Basinger, J.F.; Garrett, A. A new proposal concerning the botanical origin of Baltic amber. Proc. R. Soc. B Biol. Sci. 2009, 276, 3403–3412. [Google Scholar] [CrossRef] [PubMed]
- Weitschat, W.; Wichard, W. Chapter 6. Baltic amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 80–115. [Google Scholar]
- Kosmowska-Ceranowicz, B. Bursztyn w Polsce i na świecie. Amber in Poland and in the World. Wydanie 2, 2nd ed.; Wydawnictwo Uniwersytetu Warszawskiego: Warsaw, Poland, 2017; 311p. [Google Scholar] [CrossRef]
- Małka, A.; Jegliński, W.; Kramarska, R. The formation of amber accumulations in the form of fossil beaches on the southern shore of the Bay of Gdańsk in the Holocene. In The Baltic Sea–A Mediterranean of North Europe: In the Light of Archaeological, Historical and Natural Science Research from Ancient to Early Medieval Times; Felczak, O., Ed.; Gdańsk Archaeological Museum: Gdańsk, Poland, 2015; pp. 19–22. [Google Scholar]
- Kramarska, R.; Kasiński, J.R.; Słodkowska, B. Bursztyn, sukcynit (amber, succinite). In Bilans Perspektywicznych Zasobów Kopalin Polski wg Stanu na 31.12.2018 r; Szamałek, K., Szuflicki, M., Mizerski, W., Eds.; PIG-PIB: Warszawa, Poland, 2020; pp. 371–377. (In Polish) [Google Scholar]
- Kasiński, J.R.; Kramarska, R.; Słodkowska, B.; Sivkov, V.V.; Piwocki, M. Paleocene and Eocene deposits on the eastern margin of the Gulf of Gdańsk (Yantarny P-1 borehole, Kaliningrad region, Russia). Geol. Q. 2020, 64, 29–53. [Google Scholar] [CrossRef]
- Ross, A.J.; Bojarski, B.; Szwedo, J. A critical review of the age of Baltic amber. In Book of Abstracts. 40th Anniversary of the Palaeoentomological Section of the Polish Entomological Society International Palaeoentomological Conference Fossil Insects Bridging Past & Present, 22–25 May 2025, Kraków, Poland; Kopeć, K., Sontag, E., Soszyńska, A., Szwedo, J., Eds.; Institute of Systematics and Evolution of Animals, Polish Academy of Sciences: Kraków, Poland, 2025; pp. 69–70. [Google Scholar]
- Szwedo, J.; Stroiński, A. Who’s that girl? The singular Tropiduchidae planthopper from the Eocene Baltic amber (Hemiptera: Fulgoromorpha). Palaeontol. Electron. 2017, 20, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Drohojowska, J.; Szwedo, J. New Aleyrodidae (Hemiptera: Sternorrhyncha: Aleyrodomorpha) from the Eocene Baltic amber. Pol. J. Entomol. 2011, 80, 659–677. [Google Scholar] [CrossRef]
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordinus, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis; Editio decima, reformata; Tomus Primus, Impensis Direct; Laurentii Salvii: Stockholm, Sweden, 1758; 824p. [Google Scholar]
- Amyot, C.J.-B.; Audinet-Serville, J.G. Deuxième Partie. Homoptères. Homoptera Latr. Histoire Naturelle des Insects. Hemiptères; Librairie encyclopedique de Roret: Paris, France, 1843; pp. 1–676. [Google Scholar]
- Chou, I. Some viewpoints about insect taxonomy. Acta Entomol. Sinica 1963, 12, 586–596. [Google Scholar]
- Nieć, M.; Kramarska, R.; Sałaciński, R. Perspektywy poszukiwań złóż bursztynu w Polsce. Zesz. Nauk. IGSMiE PAN 2010, 79, 345–362. (In Polish) [Google Scholar]
- Rappsilber, I. Bitterfelder Bernstein: Geschichte–Vielfalt–Entstehung. Bitterfeld Amber: History–Diversity–Origin; Ampyx-Verlag: Halle, Germany, 2022; 324p. [Google Scholar]
- Perkovsky, E.; Zosimovich, V.Y.; Vlaskin, A.P. Chapter 7. Rovno amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 116–136. [Google Scholar]
- Gazda, L. (Ed.) Lubelski bursztyn. Znaleziska, geologia, złoża, perspektywy; Państwowa Wyższa Szkoła Zawodowa w Chełmie: Kraków, Poland, 2016; 226p. (In Polish) [Google Scholar]
- Martirosyan, O.V. Iskopaemye smoly arkticheskikh territoriï Danii, Kanady, SShA i Norvegii: K istorii izucheniya [Fossil resins of the Arctic territories of Denmark, Canada, the USA and Norway: To the history of their study]. Zesz. Nauk. IGSMiE PAN 2024, 153, 141–155. (In Russian) [Google Scholar]
- Gerling, D. Whiteflies: Their Bionomics, Pest Status and Management; Intercept Ltd.: Andover, The Netherlands, 1990; 348p. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drohojowska, J.; Gorzelańczyk, A.; Szwedo, J. A New Species of Eocene Whitefly—Gregorites michalskii sp. nov. (Hemiptera, Sternorrhyncha, Aleyrodidae) from Baltic Amber. Diversity 2025, 17, 487. https://doi.org/10.3390/d17070487
Drohojowska J, Gorzelańczyk A, Szwedo J. A New Species of Eocene Whitefly—Gregorites michalskii sp. nov. (Hemiptera, Sternorrhyncha, Aleyrodidae) from Baltic Amber. Diversity. 2025; 17(7):487. https://doi.org/10.3390/d17070487
Chicago/Turabian StyleDrohojowska, Jowita, Anita Gorzelańczyk, and Jacek Szwedo. 2025. "A New Species of Eocene Whitefly—Gregorites michalskii sp. nov. (Hemiptera, Sternorrhyncha, Aleyrodidae) from Baltic Amber" Diversity 17, no. 7: 487. https://doi.org/10.3390/d17070487
APA StyleDrohojowska, J., Gorzelańczyk, A., & Szwedo, J. (2025). A New Species of Eocene Whitefly—Gregorites michalskii sp. nov. (Hemiptera, Sternorrhyncha, Aleyrodidae) from Baltic Amber. Diversity, 17(7), 487. https://doi.org/10.3390/d17070487