

Morphological and Molecular Characterization of Eggs and Paralarvae of Green Octopus, Octopus hubbsorum Berry 1953, from the Gulf of California


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Molecular Identification and Phylogenetic Analyses of Embryos and Paralarvae
2.3. Morphological Characterization of Egg Strings and Paralarvae
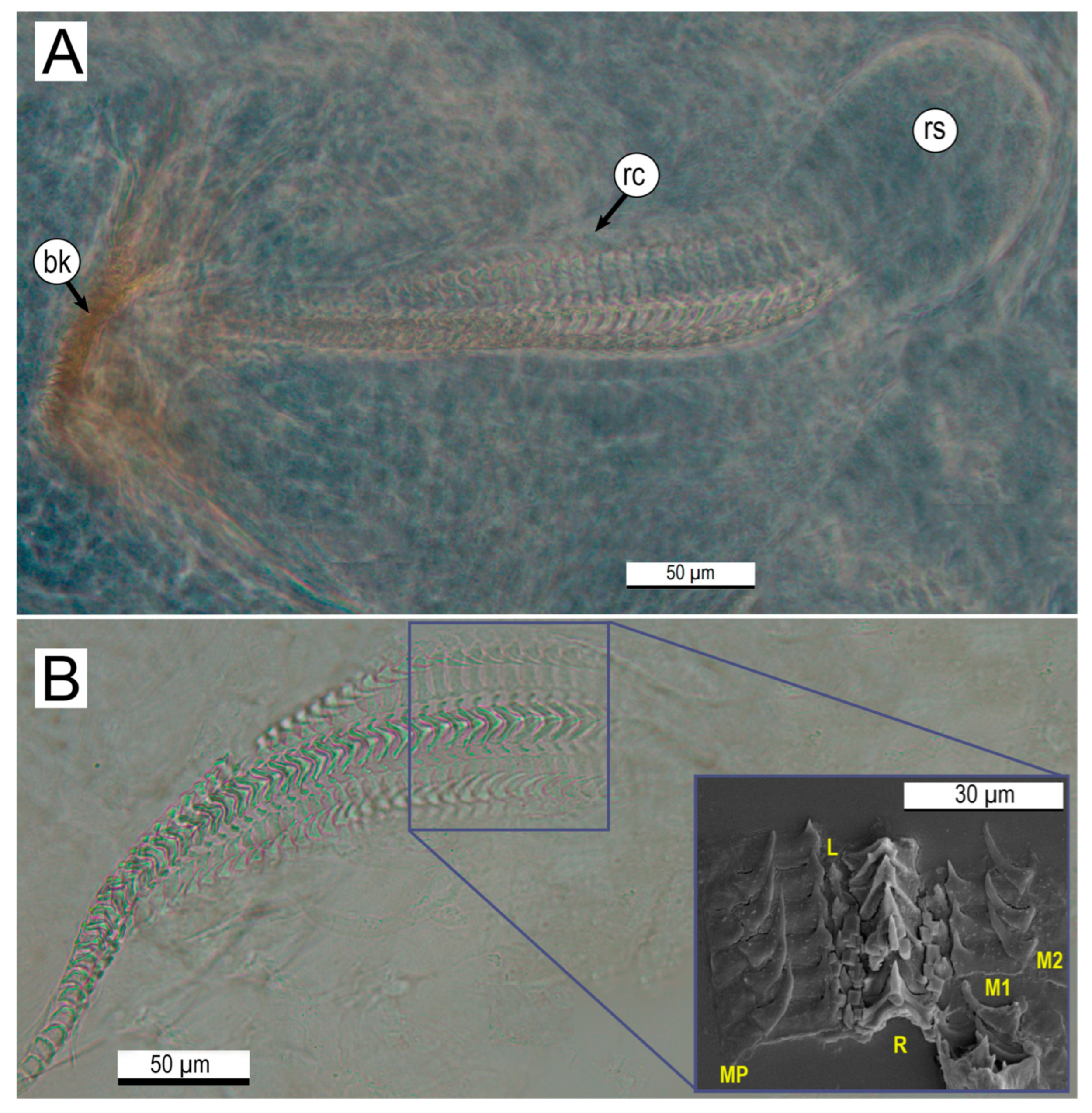
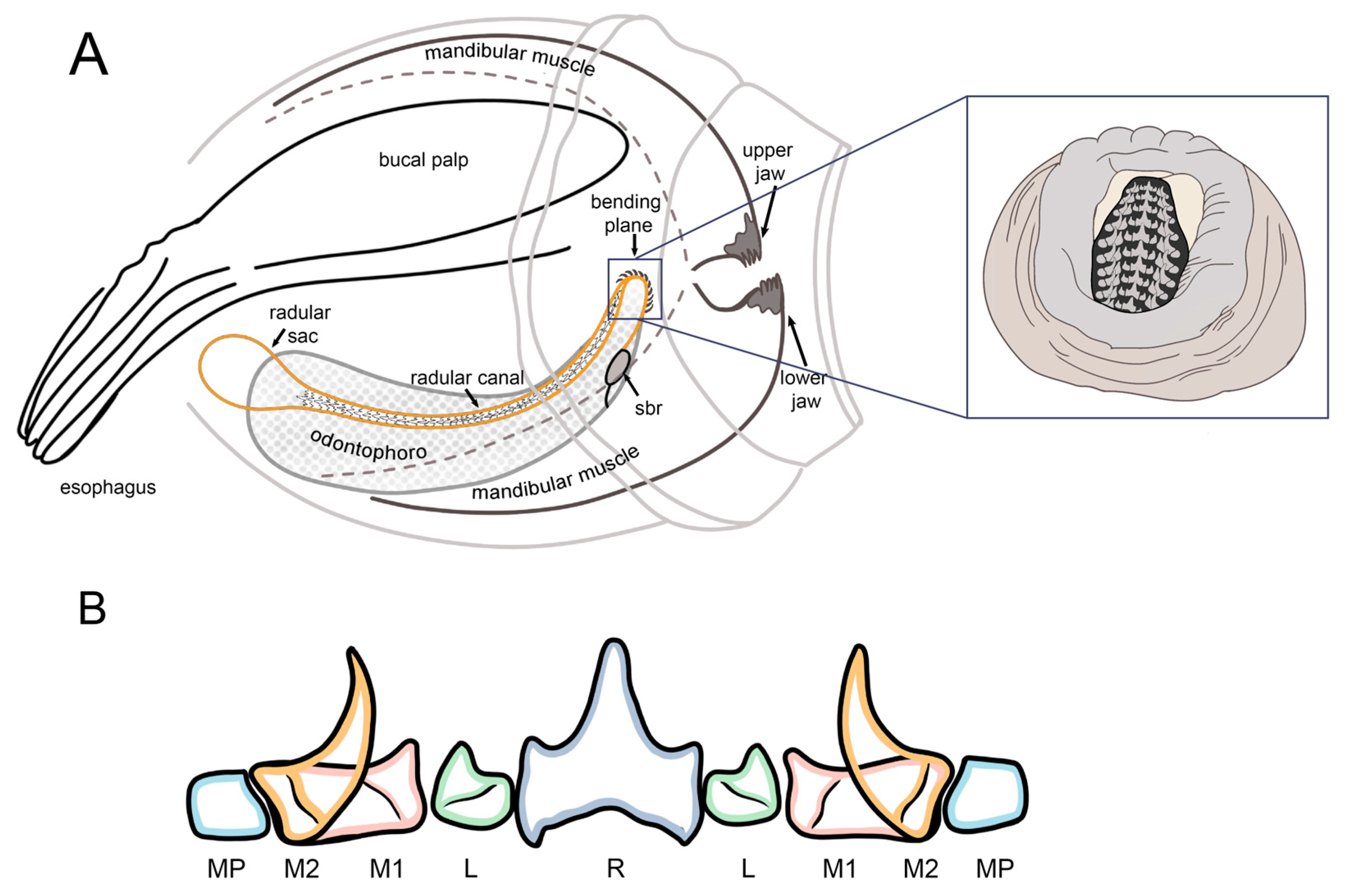
2.4. Microstructural Analyses of Suckers, Beaks, and Radulae of Paralarvae
2.5. Statistical Analyses
3. Results
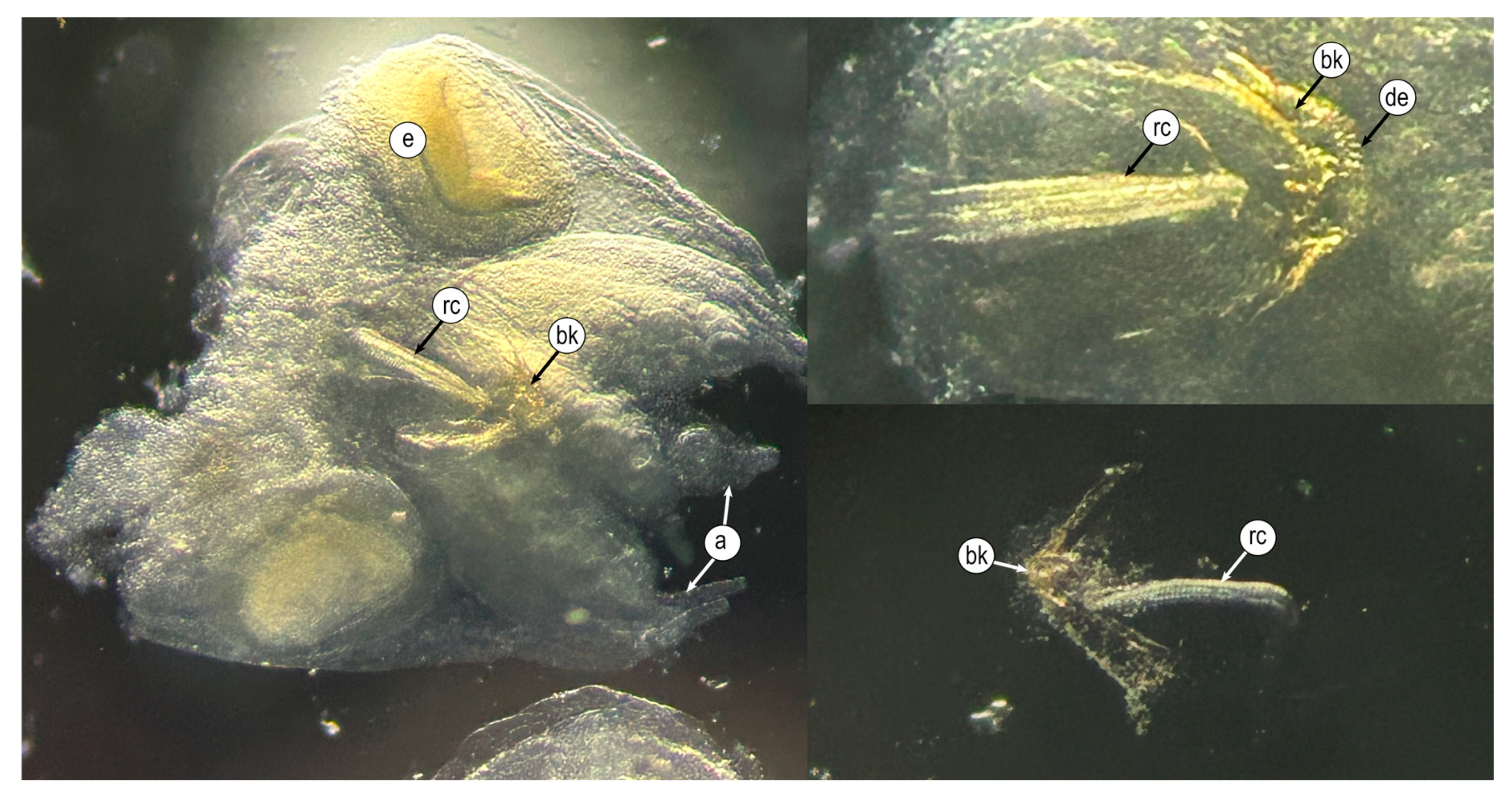
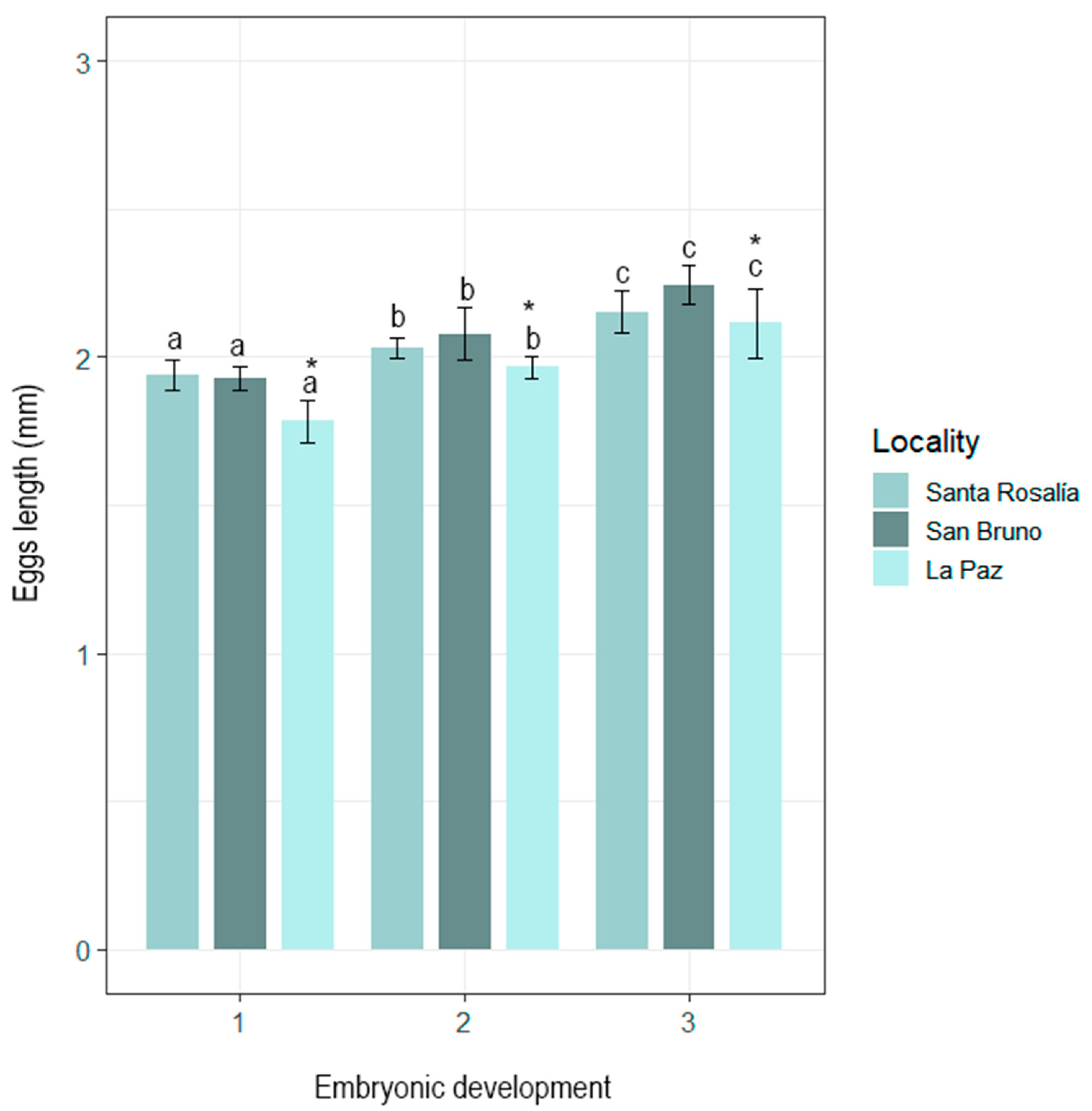
3.1. Morphological Characterization of Eggs and Paralarvae
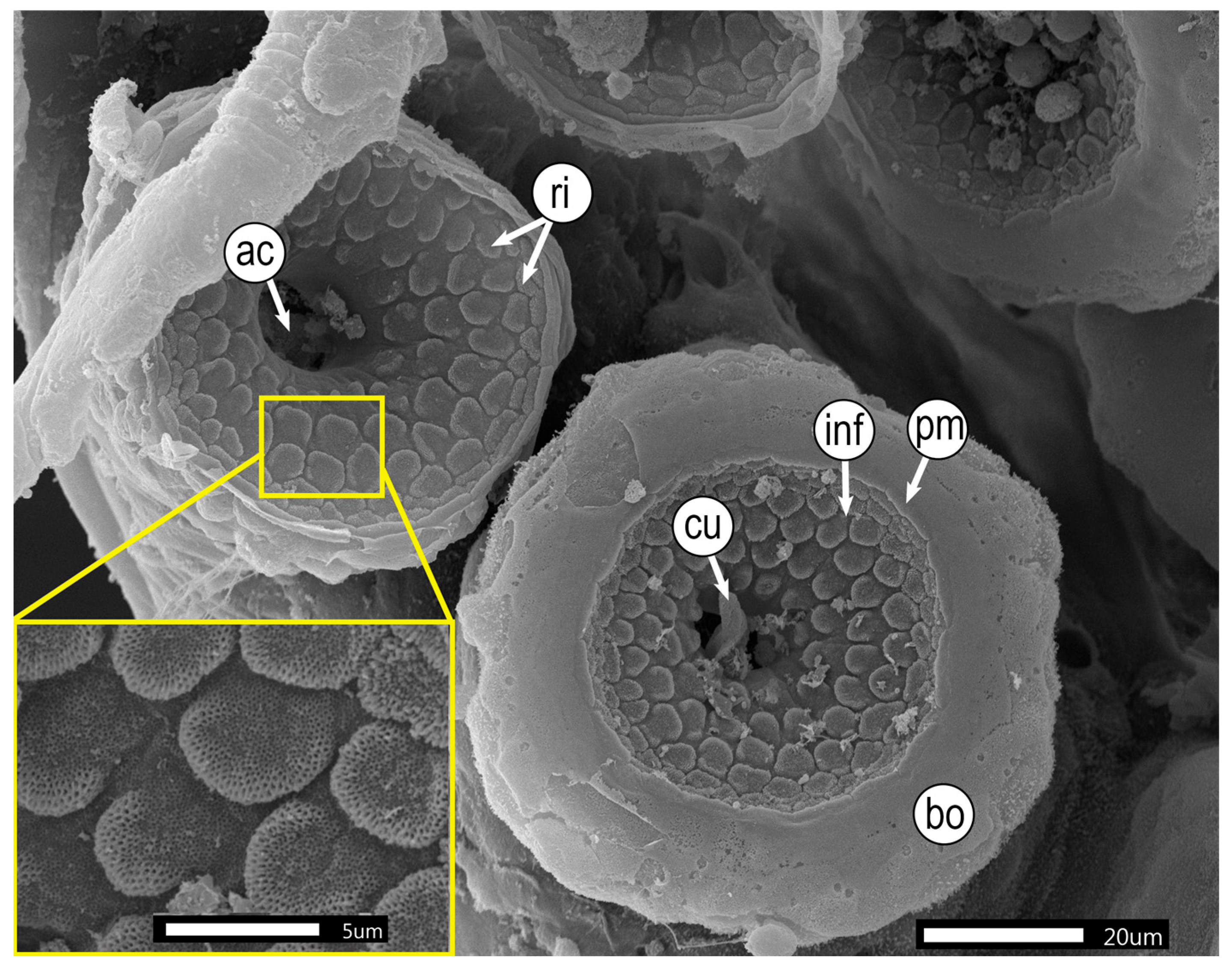
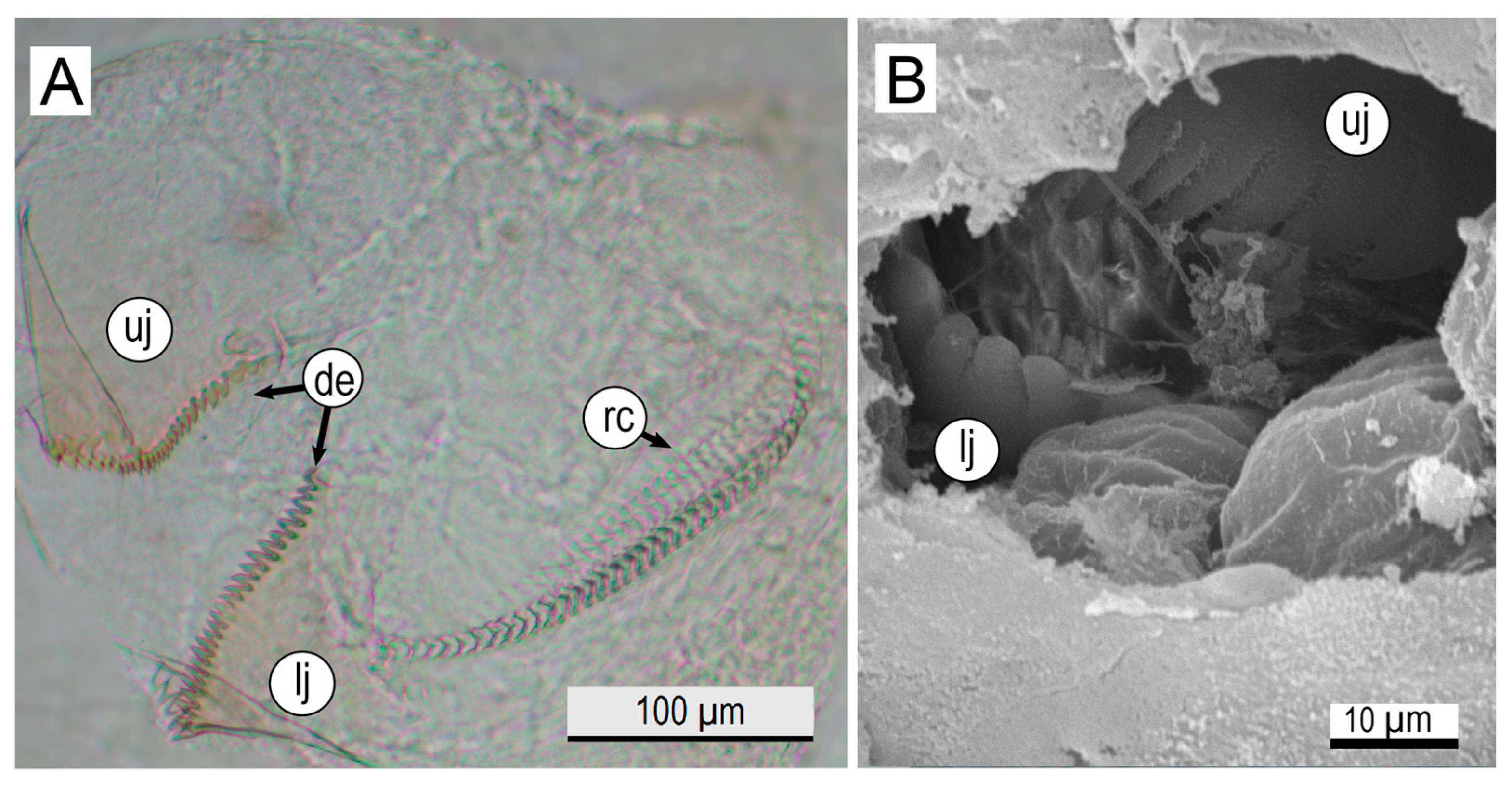
3.2. Microstructural Analyses of Suckers, Beaks, and Radulae of Paralarvae
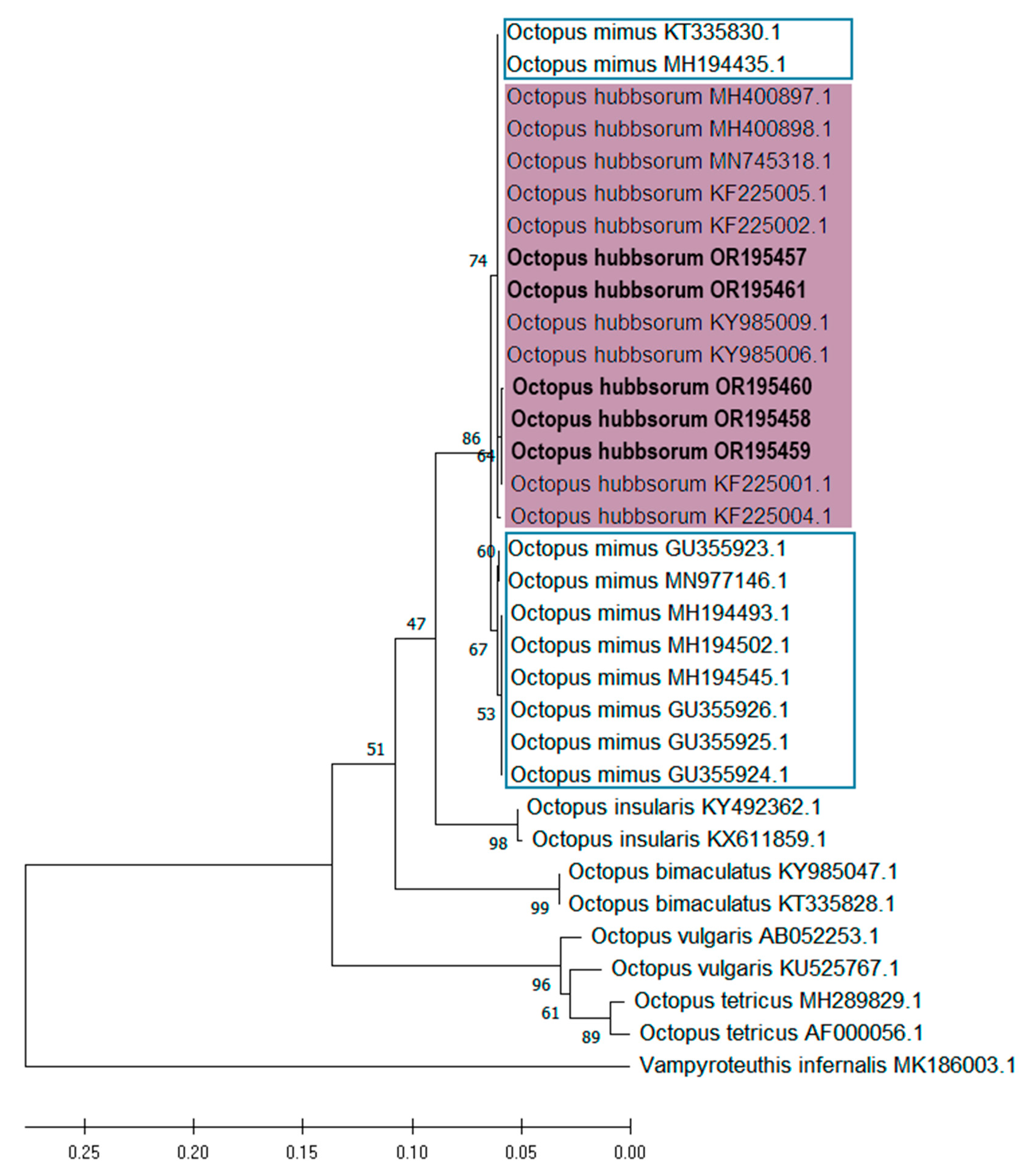
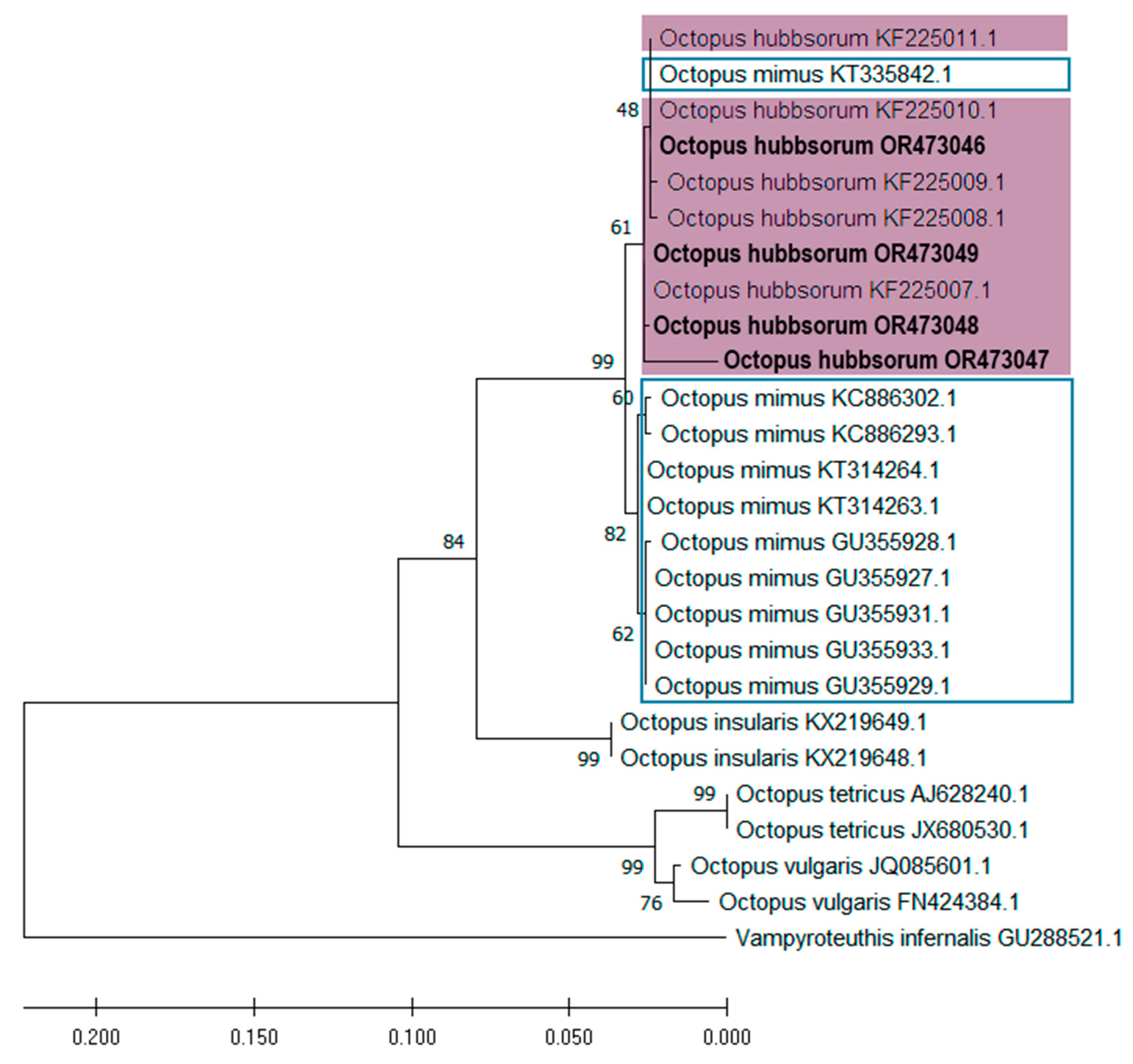
3.3. Molecular Identification and Phylogenetic Analyses of Embryos and Paralarvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Developmental Phase | Sampling Region | GenBank Accession Number | Reference | |
|---|---|---|---|---|---|
| COI | COIII | ||||
| Octopus | Paralarvae | San Bruno, BCS, Mex. | OR195457 | OR473046 | This study |
| hubbsorum | Embryo | San Bruno, BCS, Mex. | OR195458 | OR473047 | This study |
| Embryo | Santa Rosalía, BCS, Mex. | OR195459 | OR473048 | This study | |
| Embryo | Santa Rosalía, BCS, Mex. | OR195460 | OR473049 | This study | |
| Embryo | Ensenada de La Paz, BCS, Mex. | OR195461 | - | This study | |
| Adult | San Juanico, BCS, Mex. | MH400897 | - | [8] | |
| Adult | Acapulco, Gro, Mex. | MH400898 | - | [8] | |
| Adult | Bahía Magdalena, BCS, Mex | MN745318 | - | [73] | |
| Adult | Michoacán, Mex. | KF225001 | KF225007 | [21] | |
| Adult | Nayarit, Mex. | KF225002 | KF225008 | [21] | |
| Adult | Nayarit, Mex. | - | KF225009 | [21] | |
| Adult | Sonora, Mexico | KF225004 | KF225010 | [21] | |
| Adult | Malpelo Island, Colombia | KF225005 | KF225011 | [21] | |
| Adult | Santa Rosalía, BCS, Mex. | KY985006 | - | [74] | |
| Adult | Santa Rosalía, BCS, Mex. | KY985009 | - | [74] | |
| Octopus mimus | Adult | Salinas, Ecuador | KT335830 | KT335842 | [50] |
| Adult | - | - | GU355931 | [14] | |
| Adult | Coloso, Chile | GU355923 | GU355928 | [14] | |
| Adult | Callao, Perú | GU355924 | GU355927 | [14] | |
| Adult | Callao, Perú | GU355925 | GU355933 | [14] | |
| Adult | Coloso, Chile | GU355926 | GU355929 | [14] | |
| Adult | - | MN977146 | - | [75] | |
| Adult | Trujillo, Perú | MN194435 | - | [51] | |
| Adult | Miraflores, Perú | MH194493 | - | [51] | |
| Adult | Huarmey, Perú | MH194502 | - | [51] | |
| Adult | San Isidro, Perú | MH194545 | - | [51] | |
| Paralarvae | Caleta Punta Arenas, Chile | - | KT314263 | [76] | |
| Paralarvae | Caleta Punta Arenas, Chile | - | KT314264 | [76] | |
| Adult | Perú | - | KC886293 | [48] | |
| Adult | Chile | - | KC886302 | [48] | |
| Octopus | Adult | Veracruz, México | KY492362 | - | [77] |
| insularis | Adult | Ceará, Brazil | KX611859 | - | [78] |
| Adult | Veracruz, México | - | KX219649 | [77] | |
| Adult | Veracruz, México | - | KX219648 | [77] | |
| Octopus s | Adult | Guerrero Negro, BCS, Mexico | KY985047 | - | [72] |
| bimaculatu | Adult | Golfo de California, Mexico | KT335828 | - | [77] |
| Octopus | Adult | Merimbula, NSW, Australia | MH289829 | - | [79] |
| tetricus | Adult | - | AF000056 | - | [80] |
| Adult | Australia | - | AJ628240 | [28] | |
| Adult | Australia: Bendalong | - | JX680530 | [81] | |
| Octopus s | Adult | - | AB052253 | - | [82] |
| vulgari | Adult | Chile: Juan Fernandez Island | KU525767 | - | [7] |
| Adult | French Southern and Antarctic Lands: Saint Paul | - | FN424384 | [83] | |
| Adult | Mediterranean Sea | - | JQ085601 | [84] | |
| Vampyroteuthis | Adult | New Zealand | MK186003 | - | [85] |
| infernalis | Adult | - | - | GU288521 | [86] |
References
- De Queiroz, K. Ernst Mayr and the Modern Concept of Species. Proc. Natl. Acad. Sci. USA 2005, 102, 6600–6607. [Google Scholar] [CrossRef] [PubMed]
- Lima, F.D.; Strugnell, J.M.; Leite, T.S.; Lima, S.M.Q. A Biogeographic Framework of Octopod Species Diversification: The Role of the Isthmus of Panama. PeerJ 2020, 8, e8691. [Google Scholar] [CrossRef] [PubMed]
- Warnke, K.; Soeller, R.; Blohm, D.; Saint-Paul, U. Assessment of the Phylogenetic Relationship between Octopus vulgaris Cuvier, 1797 and O. mimus Gould 1852, Using Mitochondrial 16S r DNA in Combination with Morphological Characters. Abh. Geol. Bundesanst. 2002, 57, 117. [Google Scholar]
- Norman, M.D.; Hochberg, F.G. The Current State of Octopus Taxonomy. Phuket Mar. Biol. Cent. Res. Bull. 2005, 66, 127–154. [Google Scholar]
- Ortiz, N.; Ré, M.E.; Márquez, F. First Description of Eggs, Hatchlings and Hatchling Behaviour of Enteroctopus megalocyathus (Cephalopoda: Octopodidae). J. Plankton Res. 2006, 28, 881–890. [Google Scholar] [CrossRef]
- Amor, M.D.; Laptikhovsky, V.; Norman, M.D.; Strugnell, J.M. Genetic Evidence Extends the Known Distribution of Octopus insularis to the Mid-Atlantic Islands Ascension and St Helena. J. Mar. Biol. Assoc. U. K. 2015, 97, 753–758. [Google Scholar] [CrossRef]
- Amor, M.D.; Norman, M.D.; Roura, A.; Leite, T.S.; Gleadall, I.G.; Reid, A.; Perales-Raya, C.; Lu, C.C.; Silvey, C.J.; Vidal, E.A.G.; et al. Morphological Assessment of the Octopus vulgaris Species Complex Evaluated in Light of Molecular-Based Phylogenetic Inferences. Zool. Scr. 2017, 46, 275–288. [Google Scholar] [CrossRef]
- Díaz-Santana-Iturrios, M.; Salinas-Zavala, C.A.; García-Rodríguez, F.J.; Granados-Amores, J. Taxonomic Assessment of Species of the Genus Octopus from the Northeastern Pacific via Morphological, Molecular and Morphometric Analyses. PeerJ 2019, 7, e8118. [Google Scholar] [CrossRef]
- Sweeney, M.J.; Roper, C.F.E.; Mangold, K.M.; Clark, M.R.; Boletzky, S.V. “Larval” and Juvenile Cephalopods: A Manual for Their Identification; Smithsonian Institution Press: Washington, DC, USA, 1992; p. 279. [Google Scholar]
- Messenger, J.B.; Young, J.Z. The Radular Apparatus of Cephalopods. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 161–182. [Google Scholar] [CrossRef]
- Young, R.E.; Harman, R.F.; Hochberg, F.G. Octopodid Paralarvae from Hawaiian Waters. Veliger 1989, 32, 152–165. [Google Scholar]
- Zaragoza, N.; Quetglas, A.; Moreno, A. Identification Guide for Cephalopod Paralarvae from the Mediterranean Sea; ICES Cooperative Research Reports (CRR), No. 324; ICES: Copenhagen, Denmark, 2015; p. 91. [Google Scholar]
- Warnke, K.; Söller, R.; Blohm, D.; Saint-Paul, U. Rapid Differentiation between Octopus vulgaris Cuvier (1797) and Octopus Mimus Gould (1852), Using Randomly Amplified Polymorphic DNA. J. Zool. Syst. Evol. Res. 2000, 38, 119–122. [Google Scholar] [CrossRef]
- Acosta-Jofré, M.S.; Sahade, R.; Laudien, J.; Chiappero, M.B. A Contribution to the Understanding of Phylogenetic Relationships among Species of the Genus Octopus (Octopodidae: Cephalopoda). Sci. Mar. 2012, 76, 311–318. [Google Scholar] [CrossRef]
- Santana-Cisneros, M.L.; Rodríguez-Canul, R.; Zamora-Briseño, J.A.; Améndola-Pimenta, M.; De Silva-Dávila, R.; Ordóñez-López, U.; Velázquez-Abunader, I.; Ardisson, P.L. Morphological and Molecular Identification of Octopoda (Mollusca: Cephalopoda) Paralarvae from the Southern Gulf of Mexico. Bull. Mar. Sci. 2021, 97, 281–304. [Google Scholar] [CrossRef]
- Jesus, M.D.; Sales, J.B.d.L.; Martins, R.S.; Ready, J.S.; Costa, T.A.S.; Ablett, J.D.; Schiavetti, A. Traditional Knowledge Aids Description When Resolving the Taxonomic Status of Unsettled Species Using Classical and Molecular Taxonomy: The Case of the Shallow-Water Octopus Callistoctopus furvus (Gould, 1852) From the Western Atlantic Ocean. Front. Mar. Sci. 2021, 7, 595244. [Google Scholar] [CrossRef]
- De Luna Sales, J.B.; Haimovici, M.; Ready, J.S.; Souza, R.F.; Ferreira, Y.; de Cassia Silva Pinon, J.; Costa, L.F.C.; Asp, N.E.; Sampaio, I.; Schneider, H. Surveying Cephalopod Diversity of the Amazon Reef System Using Samples from Red Snapper Stomachs and Description of a New Genus and Species of Octopus. Sci. Rep. 2019, 9, 5956. [Google Scholar] [CrossRef]
- Jereb, P.; Roper, C.F.E.; Norman, M.; Finn, J. Cephalopods of the World. An Annotated and Illustrated Catalogue of Cephalopod Species Known to Date. Octopods and Vampire Squids. In FAO Species Catalogue for Fishery Purposes; FAO: Rome, Italy, 2016; Volume 3, p. 398. [Google Scholar]
- Gotshall, D. Marine Animals of Baja California: A Guide to the Common Fish and Invertebrates; Sea Challengers: Monterey, CA, USA, 1987. [Google Scholar]
- Domínguez-Contreras, J.F.; Ceballos-Vázquez, B.P.; Hochberg, F.G.; Arellano-Martínez, M. A New Record in a Well-Established Population of Octopus hubbsorum (Cephalopoda: Octopodidae) Expands Its Known Geographic Distribution Range and Maximum Size. Am. Malacol. Bull. 2013, 31, 95–99. [Google Scholar] [CrossRef]
- Pliego-Cárdenas, R.; Hochberg, F.G.; De León, F.J.G.; Barriga-Sosa, I.D.L.A. Close Genetic Relationships between Two American Octopuses: Octopus hubbsorum Berry, 1953, and Octopus mimus Gould, 1852. J. Shellfish. Res. 2014, 33, 293–303. [Google Scholar] [CrossRef]
- Cardoso, F.; Villegas, P.; Estrella, C. Observaciones Sobre La Biología de Octopus mimus (Cephalopoda: Octopoda) En La Costa Peruana. Rev. Peru. Biol. 2004, 11, 45–50. [Google Scholar] [CrossRef]
- Sauer, W.H.H.; Gleadall, I.G.; Downey-Breedt, N.; Doubleday, Z.; Gillespie, G.; Haimovici, M.; Ibáñez, C.M.; Katugin, O.N.; Leporati, S.; Lipinski, M.R.; et al. World Octopus Fisheries. Rev. Fish. Sci. Aquac. 2021, 29, 279–429. [Google Scholar] [CrossRef]
- Ibáñez, C.M.; Carrasco, S.A.; Díaz-Santana-Iturrios, M.; Cisneros, R.; Pardo-Gandarillas, M.C. Octopus mimus, the Changos’ Octopus. In Octopus Biology and Ecology; Elsevier: Amsterdam, The Netherlands, 2024; pp. 119–131. ISBN 9780128206393. [Google Scholar]
- Flores-Valle, A. Descripción Morfológica y Determinación De Identidades Genéticas De Pulpos De Las Costas Mexicanas. Master’s Thesis, Universidad Autónoma Metropolitana—Unidad Iztapalapa, Mexico City, Mexico, 2010; p. 119. [Google Scholar]
- Aljanabi, S.M.; Martinez, I. Universal and Rapid Salt-Extraction of High Quality Genomic DNA for PCR-Based Techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Guzik, M.T.; Norman, M.D.; Crozier, R.H. Molecular Phylogeny of the Benthic Shallow-Water Octopuses (Cephalopoda: Octopodinae). Mol. Phylogenet. Evol. 2005, 37, 235–248. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, Weighting, and Phylogenetic Utility of Mitochondrial Gene Sequences and a Compilation of Conserved Polymerase Chain Reaction Primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Hall, T.A. BIOEDIT: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/ NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Akaike, H. Information Theory and an Extension of the Maximum Likelihood Principle BT—Selected Papers of Hirotugu Akaike. In Selected Papers of Hirotugu Akaike; Springer: New York, NY, USA, 1998; pp. 199–213. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research-an Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Hochberg, F.G.; Nixon, M.; Toll, R.B. Order Octopoda Leach, 1818. Smithson. Contrib. Zool. 1992, 513, 213–280. [Google Scholar]
- Geiger, D.L.; Marshall, B.A.; Ponder, W.F.; Sasaki, T.; Warén, A. Techniques for Collecting, Handling, Preparing, Storing and Examining Small Molluscan Specimens. Molluscan. Res. 2007, 27, 1–50. [Google Scholar] [CrossRef]
- Kruta, I.; Landman, N.H.; Tanabe, K. Ammonoid Radula. In Ammonoid Paleobiology: From Anatomy to Ecology; Springer: Dordrecht, The Netherlands, 2015; pp. 485–505. [Google Scholar]
- Posit Team. RStudio: Integrated Development Environment for R; Posit Software: Boston, MA, USA, 2023. [Google Scholar]
- Alejo-Plata, M.D.C.; Herrera Alejo, S.N. First Description of Eggs and Paralarvae of Green Octopus Octopus hubbsorum (Cephalopoda: Octopodidae) under Laboratory Conditions. Am. Malacol. Bull. 2014, 32, 132–139. [Google Scholar] [CrossRef]
- Montero-Ruíz, R.U.; De Silva-Dávila, R.; Amezcua-Gómez, C.A.; Valdez-Cibrián, A.; Kozak, E.R. Description of the Spawning, Embryonic Development, and Paralarvae of the Green Octopus Octopus hubbsorum Berry 1953 (Cephalopoda: Octopodidae) under Laboratory Conditions. Mar. Biol. 2023, 170, 116. [Google Scholar] [CrossRef]
- Guerra, A.; Cortez, T.; Rocha, F. Redescripción Del Pulpo de Los Changos, Octopus mimus Gould, 1852, Del Litoral Chileno-Peruano (Mollusca, Cephalopoda). Iberus 1999, 17, 37–57. [Google Scholar]
- Warnke, K. Observations on the Embryonic Development of Octopus mimus (Mollusca: Cephalopoda) from Northern Chile. Veliger 1999, 42, 211–217. [Google Scholar]
- Castro-Fuentes, H.; Olivares-Paz, A.; Quintana-Fellay, A.; Zuñiga-Romero, O. Descripción Del Desarrollo Embrionario y Paralarva de Octopus mimus (Gould, 1852) (Mollusca: Cephalopoda) En Cautiverio. Estud. Oceanol. 2002, 21, 13–25. [Google Scholar]
- Valdez-Cibrián, A.; Díaz-Santana-iturrios, M.; Landa-Jaime, V.; Michel-Morfín, J.E. First Detection of an Ocellate Octopus in the Revillagigedos Ecoregion, a Biodiversity Hotspot Located in the Tropical East Pacific Province. Zookeys 2020, 2020, 81. [Google Scholar] [CrossRef]
- Pardo-Gandarillas, M.C.; Ibáñez, C.M.; Yamashiro, C.; Méndez, M.A.; Poulin, E. Demographic Inference and Genetic Diversity of Octopus mimus (Cephalopoda: Octopodidae) throughout the Humboldt Current System. Hydrobiologia 2018, 808, 124–135. [Google Scholar] [CrossRef]
- Magallón-Gayón, E.; del Río-Portilla, M.Á.; de los Angeles Barriga-Sosa, I. The Complete Mitochondrial Genomes of Two Octopods of the Eastern Pacific Ocean: Octopus mimus and ‘Octopus’ fitchi (Cephalopoda: Octopodidae) and Their Phylogenetic Position within Octopoda. Mol. Biol. Rep. 2020, 47, 943–952. [Google Scholar] [CrossRef]
- Pliego-Cárdenas, R.; Flores, L.; Markaida, U.; Barriga-Sosa, I.D.L.Á.; Mora, E.; Arias, E. Genetic Evidence of the Presence of Octopus mimus in the Artisanal Fisheries of Octopus in Santa Elena Peninsula, Ecuador. Am. Malacol. Bull. 2016, 34, 51–55. [Google Scholar] [CrossRef]
- Marín, A.; Serna, J.; Robles, C.; Ramírez, B.; Reyes-Flores, L.E.; Zelada-Mázmela, E.; Sotil, G.; Alfaro, R. A Glimpse into the Genetic Diversity of the Peruvian Seafood Sector: Unveiling Species Substitution, Mislabeling and Trade of Threatened Species. PLoS ONE 2018, 13, e0206596. [Google Scholar] [CrossRef] [PubMed]
- Leite, T.S.; Haimovici, M.; Molina, W.; Warnke, K. Morphological and Genetic Description of Octopus insularis, a New Cryptic Species in the Octopus vulgaris Complex (Cephalopoda: Octopodidae) from the Tropical Southwestern Atlantic. J. Molluscan Stud. 2008, 74, 63–74. [Google Scholar] [CrossRef]
- Undheim, E.A.B.; Norman, J.A.; Thoen, H.H.; Fry, B.G. Genetic Identification of Southern Ocean Octopod Samples Using MtCOI. C. R. Biol. 2010, 333, 395–404. [Google Scholar] [CrossRef]
- Ortiz, N.; Ré, M.E. The Eggs and Hatchlings of the Octopus Robsonella fontaniana (Cephalopoda: Octopodidae). J. Mar. Biol. Assoc. U. K. 2011, 91, 705–713. [Google Scholar] [CrossRef]
- Carrasco, S.A. The Early Life History of Two Sympatric New Zealand Octopuses: Eggs and Paralarvae of Octopus huttoni and Pinnoctopus cordiformis. N. Z. J. Zool. 2014, 41, 32–45. [Google Scholar] [CrossRef]
- Lango-Reynoso, F.; Chávez-Villalba, J.; Cochard, J.C.; Le Pennec, M. Oocyte Size, a Means to Evaluate the Gametogenic Development of the Pacific Oyster, Crassostrea gigas (Thunberg 1793). Aquaculture 2000, 190, 183–199. [Google Scholar] [CrossRef]
- McEdward, L.; Carson, S. Variation in Egg Organic Content and Its Relationship with Egg Size in the Starfish Solaster stimpsoni. Mar. Ecol. Prog. Ser. 1987, 37, 159–169. [Google Scholar] [CrossRef]
- Moran, A.L.; McAlister, J.S. Egg Size as a Life History Character of Marine Invertebrates: Is It All It’s Cracked up to Be? Biol. Bull. 2009, 216, 226–242. [Google Scholar] [CrossRef]
- Diekmann, R.; Piatkowski, U.; Schneider, M. Early Life and Juvenile Cephalopods Around Seamounts of the Subtropical Eastern North Atlantic: Illustrations and a Key for Their Identification; Institut für Meereskunde: Kiel, Germany, 2002; p. 42. [Google Scholar]
- Norman, M.D.; Kubodera, T. Taxonomy and Biogeography of an Australian Subtropical Octopus with Japanese Affinities. In Proceedings of the 7th and 8th Symposia on Collection Building and Natural History Studies in Asia and the Pacific Rim; National Museum of Nature and Science: Tsukuba, Japan, 2006; pp. 171–189. [Google Scholar]
- Kubodera, T. Distribution and Abundance of the Early Life Stages of Octopus, Octopus dofleini Wülker, 1910 in the North Pacific. Bull. Mar. Sci. 1991, 49, 235–244. [Google Scholar]
- Schmidtberg, H. The Structure of Suckers of Newly Hatched Sepia officinalis, Loligo vulgaris, and Octopus vulgaris. Vie Milieu 1997, 47, 155–159. [Google Scholar]
- Forsythe, J.W.; Hanlon, R.T. Aspects of Egg Development, Post-Hatching Behavior, Growth and Reproductive Biology of Octopus burryi Voss, 1950 (Mollusca: Cephalopoda). Vie Milieu 1985, 35, 273–282. [Google Scholar]
- Nixon, M.; Mangold, K. The Early Life of Octopus vulgaris (Cephalopoda: Octopodidae) in the Plankton and at Settlement: A Change in Lifestyle. J. Zool. 1996, 239, 301–327. [Google Scholar] [CrossRef]
- Villanueva, R.; Norman, M.D. Biology of the Planktonic Stages of Benthic Octopuses. Oceanogr. Mar. Biol. 2008, 46, 111–208. [Google Scholar] [CrossRef]
- Nixon, M. The Radulae of Cephalopoda. Smithson. Contrib. Zool. 1998, 586, 39–53. [Google Scholar]
- Roper, C.F.E.; Voss, G.L. Guidelines for Taxonomic Descriptions of Cephalopod Species. Mem. Mus. Vic. 1983, 44, 48–63. [Google Scholar] [CrossRef]
- Samuel, D.V.; Patterson, J. A Comparative Study on the Radula of Three Coleoid Cephalopods. S. Pac. Study 2003, 24, 33–38. [Google Scholar]
- Boletzky, S.V. The Larvae of Cephalopoda: A Review. Thalass. Jugosl. 1974, 10, 45–76. [Google Scholar]
- Wells, M.J. Octopus: Physiology and Behaviour of an Advanced Invertebrate, 1st ed.; Springer: Dordrecht, The Netherlands, 1978. [Google Scholar]
- López-Uriarte, E.; Ríos-Jara, E.; Pérez-Peña, M. Range Extension for Octopus hubbsorum (Mollusca: Octopodidae) in the Mexican Pacific. Bull. Mar. Sci. 2005, 77, 171–176. [Google Scholar]
- Defeo, O.; Carlos Castilla, J. Harvesting and Economic Patterns in the Artisanal Octopus mimus (Cephalopoda) Fishery in a Northern Chile Cove. Fish. Res. 1998, 38, 121–130. [Google Scholar] [CrossRef]
- Dueñas-Romero, J.d.J.; Granados-Amores, J.; Palacios-Salgado, D.S.; Domínguez-Contreras, J.F.; Flores-Ortega, J.R.; García-Rodríguez, F.J. Diversity and Population Genetic Structure of Octopus hubbsorum in the Mexican Pacific Inferred from Mitochondrial DNA Sequences. Mar. Freshw. Res. 2020, 72, 35–43. [Google Scholar] [CrossRef]
- Domínguez-Contreras, J.F.; Munguia-Vega, A.; Ceballos-Vázquez, B.P.; Arellano-Martínez, M.; García-Rodríguez, F.J.; Culver, M.; Reyes-Bonilla, H. Life Histories Predict Genetic Diversity and Population Structure within Three Species of Octopus Targeted by Small-Scale Fisheries in Northwest Mexico. PeerJ 2018, 6, e4295. [Google Scholar] [CrossRef] [PubMed]
- Velasco, A.; Ramilo-Fernández, G.; Sotelo, C.G. A Real-Time PCR Method for the Authentication of Common Cuttlefish (Sepia officinalis) in Food Products. Foods 2020, 9, 286. [Google Scholar] [CrossRef]
- Pardo-Gandarillas, M.C.; Ibáñez, C.M.; Ruiz, J.F.; Bustos, C.A.; Peña, F.A.; Landaeta, M.F. Paralarvae of Cephalopods in Channels and Fjords of the Southern Tip of Chile (46–53°S). Fish. Res. 2016, 173, 175–182. [Google Scholar] [CrossRef]
- Flores-Valle, A.; Pliego-Cárdenas, R.; De Lourdes Jimenéz-Badillo, M.; Arredondo-Figueroa, J.L.; De Los Ángeles Barriga-Sosa, I. First Record of Octopus insularis Leite and Haimovici, 2008 in the Octopus Fishery of a Marine Protected Area in the Gulf of Mexico. J. Shellfish. Res. 2018, 37, 221–227. [Google Scholar] [CrossRef]
- Lima, F.D.; Berbel-Filho, W.M.; Leite, T.S.; Rosas, C.; Lima, S.M.Q. Occurrence of Octopus insularis Leite and Haimovici, 2008 in the Tropical Northwestern Atlantic and Implications of Species Misidentification to Octopus Fisheries Management. Mar. Biodivers. 2017, 47, 723–734. [Google Scholar] [CrossRef]
- Ramos, J.E.; Pecl, G.T.; Moltschaniwskyj, N.A.; Semmens, J.M.; Souza, C.A.; Strugnell, J.M. Population Genetic Signatures of a Climate Change Driven Marine Range Extension. Sci. Rep. 2018, 8, 9558. [Google Scholar] [CrossRef]
- Carlini, D.B.; Graves, J.E. Phylogenetic Analysis of Cytochrome c Oxidase I Sequences to Determine Higher-Level Relationships within the Coleoid Cephalopods. Bull. Mar. Sci. 1999, 64, 57–76. [Google Scholar]
- Reid, A.; Wilson, N. Reid y Wilson, Octopuses of the Kermadec Islands_ Discovery and Description of New Member of the Octopus. Bull. Auckl. Mus. 2015, 20. [Google Scholar]
- Minakata, H.; Iwakoshi, E.; Takuwa, K. Octopus vulgaris MRNA for Cytochrome c Oxidase Subunit I. 2016, Unpublished work.
- Guerra, Á.; Roura, Á.; González, Á.F.; Pascual, S.; Cherel, Y.; Pérez-Losada, M. Morphological and Genetic Evidence That Octopus vulgaris Cuvier, 1797 Inhabits Amsterdam and Saint Paul Islands (Southern Indian Ocean). ICES J. Mar. Sci. 2010, 67, 1401–1407. [Google Scholar] [CrossRef]
- Fadhlaoui-Zid, K.; Knittweis, L.; Aurelle, D.; Nafkha, C.; Ezzeddine, S.; Fiorentino, F.; Ghmati, H.; Ceriola, L.; Jarboui, O.; Maltagliati, F. Genetic Structure of Octopus vulgaris (Cephalopoda, Octopodidae) in the Central Mediterranean Sea Inferred from the Mitochondrial COIII Gene. C. R. Biol. 2012, 335, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Braid, H.E.; Bolstad, K.S.R. Cephalopod Biodiversity of the Kermadec Islands: Implications for Conservation and Some Future Taxonomic Priorities. Invertebr. Syst. 2019, 33, 402–425. [Google Scholar] [CrossRef]
- Strugnell, J.; Allcock, A.L. Co-Estimation of Phylogeny and Divergence Times of Argonautoidea Using Relaxed Phylogenetics. Mol. Phylogenet. Evol. 2010, 54, 701–708. [Google Scholar] [CrossRef] [PubMed]



| Octopus hubbsorum | Octopus mimus | ||||||
|---|---|---|---|---|---|---|---|
| Alejo-Plata et al. [42] | Montero-Ruiz et al. [43] | This Study | Guerra et al. [44] | Warnke [45] | Castro-Fuentes et al. [46] | Ibañez et al. [24] | |
| Eggs | |||||||
| Total length (TL) | 1.66 ± 0.74 | - | 2.04 ± 0.12 | - | 2.03 ± 0.25 | 2.13 ± 0.33 | 1.67 ± 0.09 |
| - | - | (1.6–2.3) | (2.6–3.2) | - | (2.22–2.51) | - | |
| Width (W) | 0.12 ± 0.06 | - | 0.78 ± 0.09 | - | 0.9 ± 0.08 | 0.79 ± 0.14 | 0.89 ± 0.04 |
| - | - | (0.7–1) | - | - | (0.76–1.12) | - | |
| Paralarva | |||||||
| Total length (TL) | 1.6 ± 0.13 | 2.1 | 1.64 ± 0.01 | - | 2.34 ± 0.19 | 3.1 ± 0.09 | 2.97 ± 0.19 |
| 2.17 ± 0.08 | |||||||
| Mantle length (ML) | 1.22 ± 0.16 | 1.55 | 1.26 ± 0.08 | - | 1.85 ± 0.08 | 1.53 ± 0.08 | 1.61 ± 0.09 |
| 1.68 ± 0.19 | |||||||
| Mantle width (MW) | 0.75 ± 0.01 | - | 0.85 ± 0.01 | - | 0.84 ± 0.09 | 1.41 ± 0.09 | - |
| 0.92 ± 0.02 | |||||||
| Head width (HW) | 0.6 ± 0.07 | 0.87 | 0.75 ± 0.03 | - | - | - | 1.03 ± 0.08 |
| 0.75 ± 0.01 | |||||||
| Eye diameter (ED) | 0.18 ± 0.03 | 0.18 | 0.26 | - | - | 0.46 ± 0.02 | 0.32 ± 0.02 |
| 0.35 | |||||||
| Funnel length (FL) | - | - | 0.32 ± 0.01 | - | - | 0.67 ± 0.09 | 0.72 ± 0.09 |
| 0.45 ± 0.03 | |||||||
| Arm length (AL) | 0.30 ± 0.06 | 0.5 | 0.34 ± 0.04 | - | 0.9–1 | 0.65 ± 0.1 | 0.82 ± 0.11 |
| 0.72 ± 0.09 | |||||||
| No. of suckers per arm | 3 | 3 | 3 | - | 3 | 3 | 3 |
| 3 | |||||||
| Sucker diameter (SD) | - | - | 0.077 ± 0.002 | - | - | 0.14 ± 0.01 | 0.10 ± 0.01 |
| 0.087 ± 0.003 | |||||||
| Kölliker’s organs | Present | Present | Present | - | - | Present | Present |
| Octopus hubbsorum | Octopus mimus | |||||
|---|---|---|---|---|---|---|
| Alejo-Plata et al. [42] | Montero-Ruiz et al. [43] | This Study | Warnke [45] | Castro-Fuentes et al. [46] | Ibañez et al. [24] | |
| Dorsal | ||||||
| Posterior cup (PC) | 14–16 | 3–5 | 4 | - | 3–7 | 2–4 |
| Visceral (V) | 5–6 | 5–8 | 6–7 | 6–8 | 6–8 | 6–11 |
| 9–10 | ||||||
| Mantle (DM) | - | 4–5 | - | 3–4 | 3–4 | 3–4 |
| Head (DH) | 14 [2 + 4 + 8] | 10 [2 +4 + 4] | 10 [2 + 4 + 4] | 10 [2+ 4 + 4] | 10 [2 + 4 + 4] | 10 |
| 12 [4 + 4 + 4] | ||||||
| Eyes (DE) | - | 1 | 1 | 1 | 3–4 | 1–3 |
| 2 | ||||||
| Arm (A) | 4 | 3–4 | - | 2–4 | 3–4 | 3–4 |
| 1 | ||||||
| Ventral | ||||||
| Mantle (VM) | 38–43 | 28–34 | 29–32 | 21–24 | 24–31 | 18–39 |
| 32–39 | ||||||
| Head (VH) | 2 | 2 | 2 | 2 | 2 | 2 |
| 2 | ||||||
| Eyes (VE) | 1 | 2 | - | - | - | - |
| Funnel (F) | 6 [4 + 2] | 6 [4 +2] | 6 [4 + 2] | 5(6) [3(4) + 2] | 6 [4 + 2] | 5–8 |
| 6 [4 + 2] | ||||||
| Arms base (AB) | 1 | - | 1 | - | - | - |
| 1 | ||||||
| Arms (A) | 4 | 3–4 | 3 | 2–4 | 3–4 | 3–4 |
| 3 | ||||||
| Total | 85–93 | - | 74–77 | 75–89 | - | - |
| 84–96 | ||||||
| Species | O. mimus_1 | O. hubbsorum_1 | O. hubbsorum_2 | O. mimus_2 |
|---|---|---|---|---|
| O. mimus_1 | 0.000 | |||
| O. hubbsorum_1 | 0.010 | 0.000 | ||
| O. hubbsorum_2 | 0.008 | 0.002 | 0.000 | |
| O. mimus_2 | 0.008 | 0.002 | 0.000 | 0.000 |
| Species | O. hubbsorum_1 | O. hubbsorum_2 | O. mimus |
|---|---|---|---|
| O. hubbsorum_1 | 0.000 | ||
| O. hubbsorum_2 | 0.002 | 0.000 | |
| O. mimus | 0.011 | 0.010 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Flores, M.; Morelos-Castro, R.M.; Arellano-Martínez, M. Morphological and Molecular Characterization of Eggs and Paralarvae of Green Octopus, Octopus hubbsorum Berry 1953, from the Gulf of California. Diversity 2025, 17, 470. https://doi.org/10.3390/d17070470
García-Flores M, Morelos-Castro RM, Arellano-Martínez M. Morphological and Molecular Characterization of Eggs and Paralarvae of Green Octopus, Octopus hubbsorum Berry 1953, from the Gulf of California. Diversity. 2025; 17(7):470. https://doi.org/10.3390/d17070470
Chicago/Turabian StyleGarcía-Flores, Maritza, Rosa María Morelos-Castro, and Marcial Arellano-Martínez. 2025. "Morphological and Molecular Characterization of Eggs and Paralarvae of Green Octopus, Octopus hubbsorum Berry 1953, from the Gulf of California" Diversity 17, no. 7: 470. https://doi.org/10.3390/d17070470
APA StyleGarcía-Flores, M., Morelos-Castro, R. M., & Arellano-Martínez, M. (2025). Morphological and Molecular Characterization of Eggs and Paralarvae of Green Octopus, Octopus hubbsorum Berry 1953, from the Gulf of California. Diversity, 17(7), 470. https://doi.org/10.3390/d17070470