

Spatial Patterns and Functional Diversity of Nearshore and Offshore Coral-Reef Fish Communities in the South China Sea

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
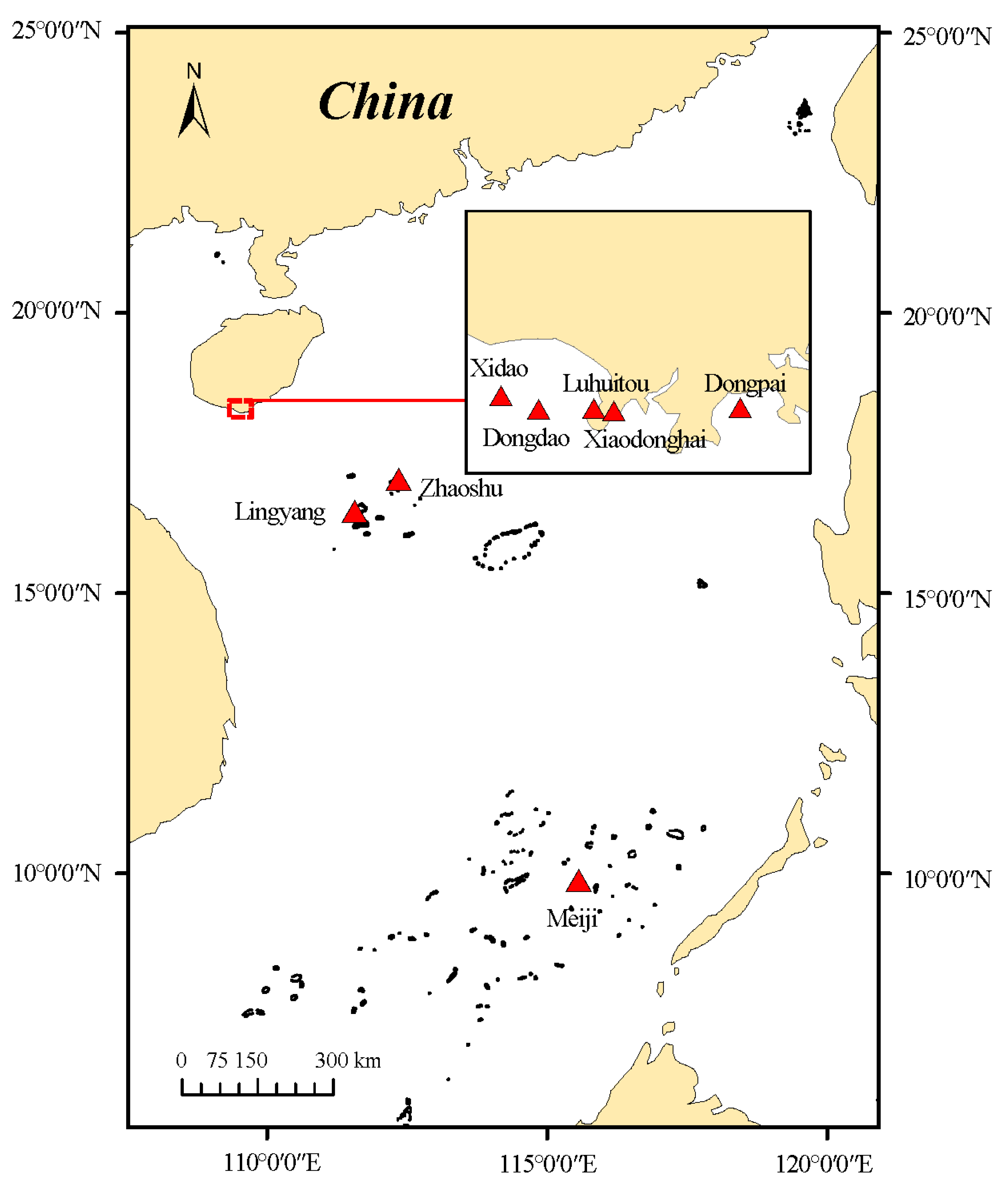
2.1. Study Area
2.2. Fish Community Survey and Trait Data Collection
2.3. Biodiversity and Functional Diversity Indices
2.4. Statistical Analyses
3. Results
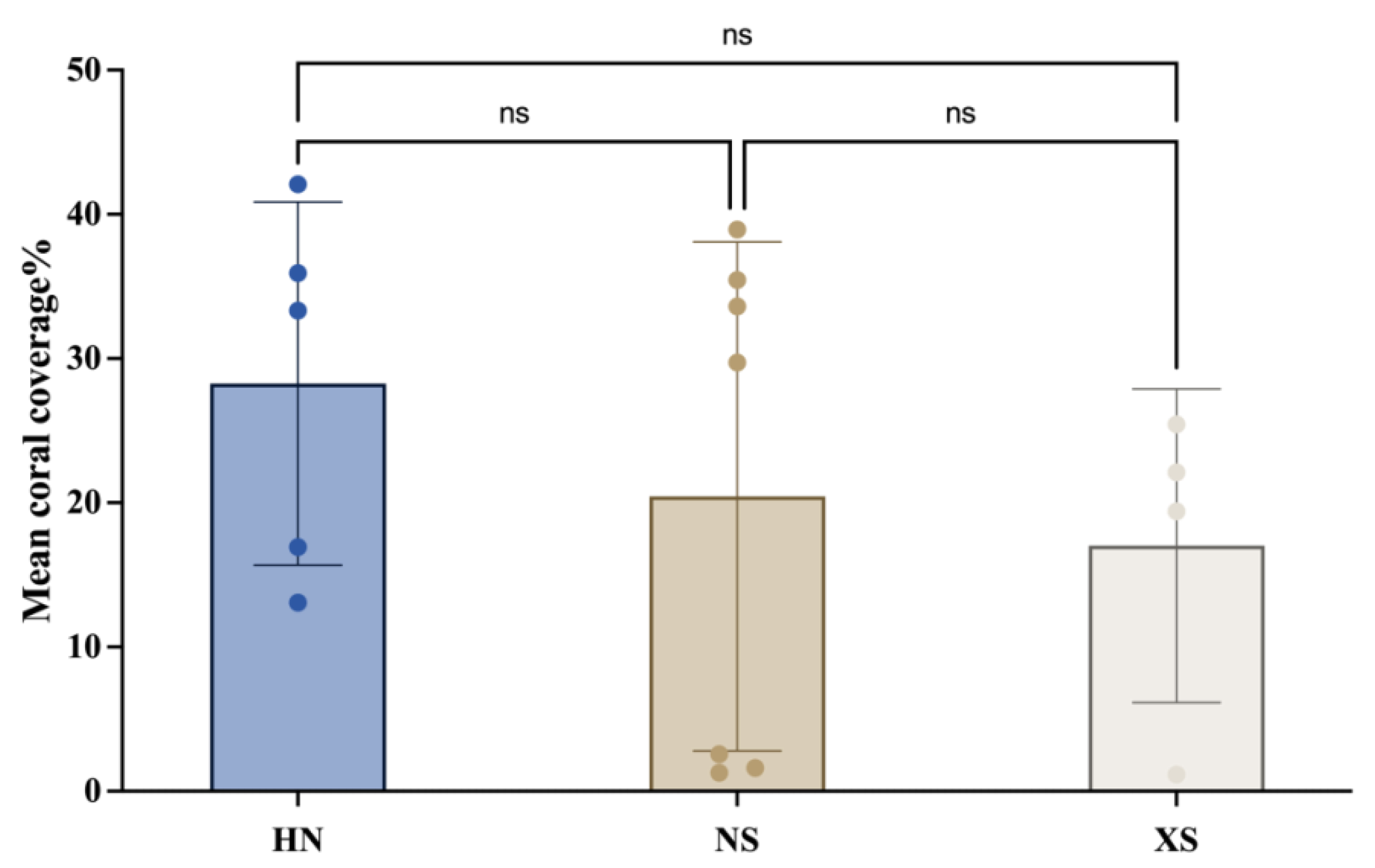
3.1. Sample and Coral Coverage
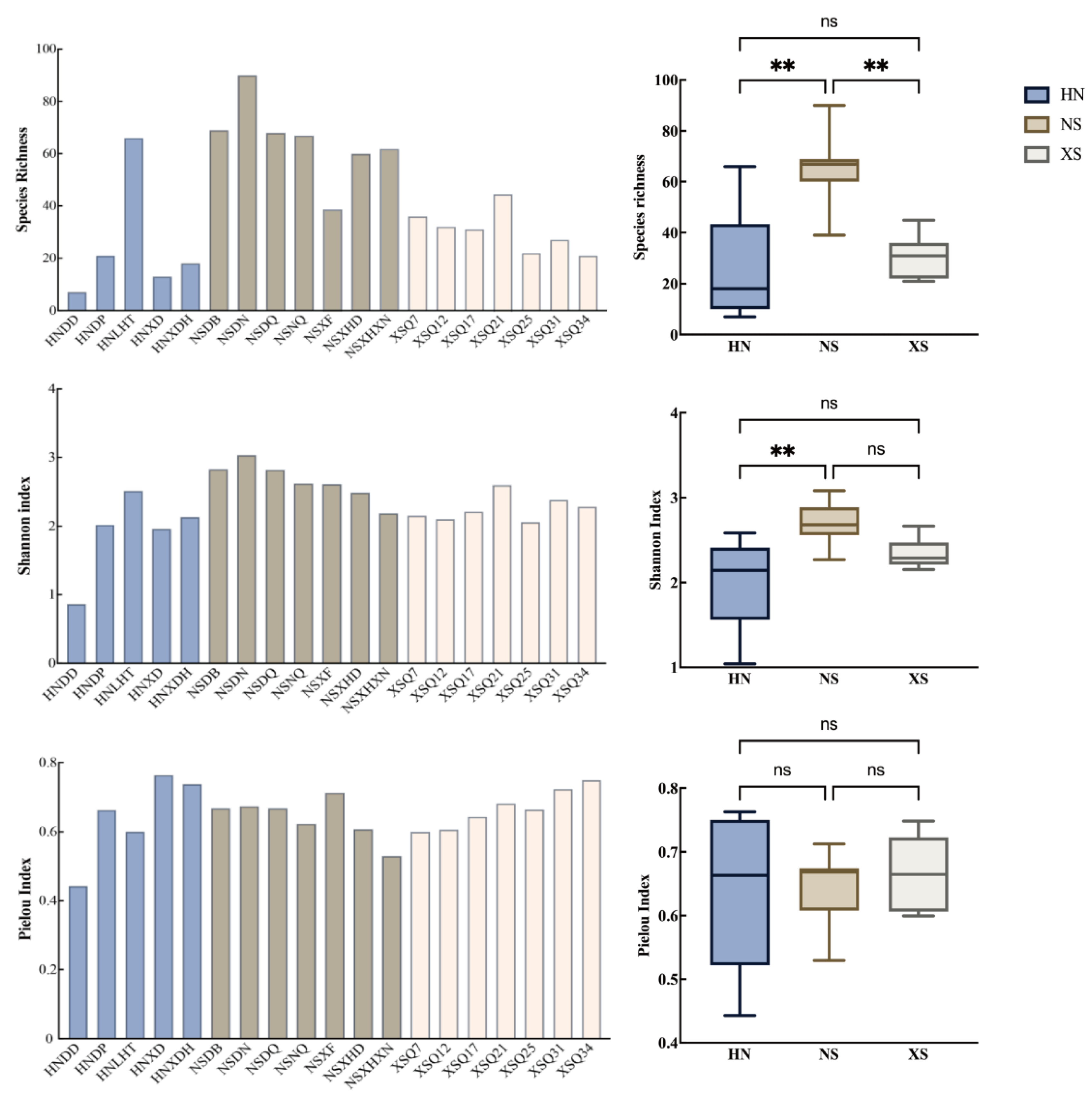
3.2. Taxonomic Diversity Patterns of Fish Communities
3.3. Fish Length Structure Analysis
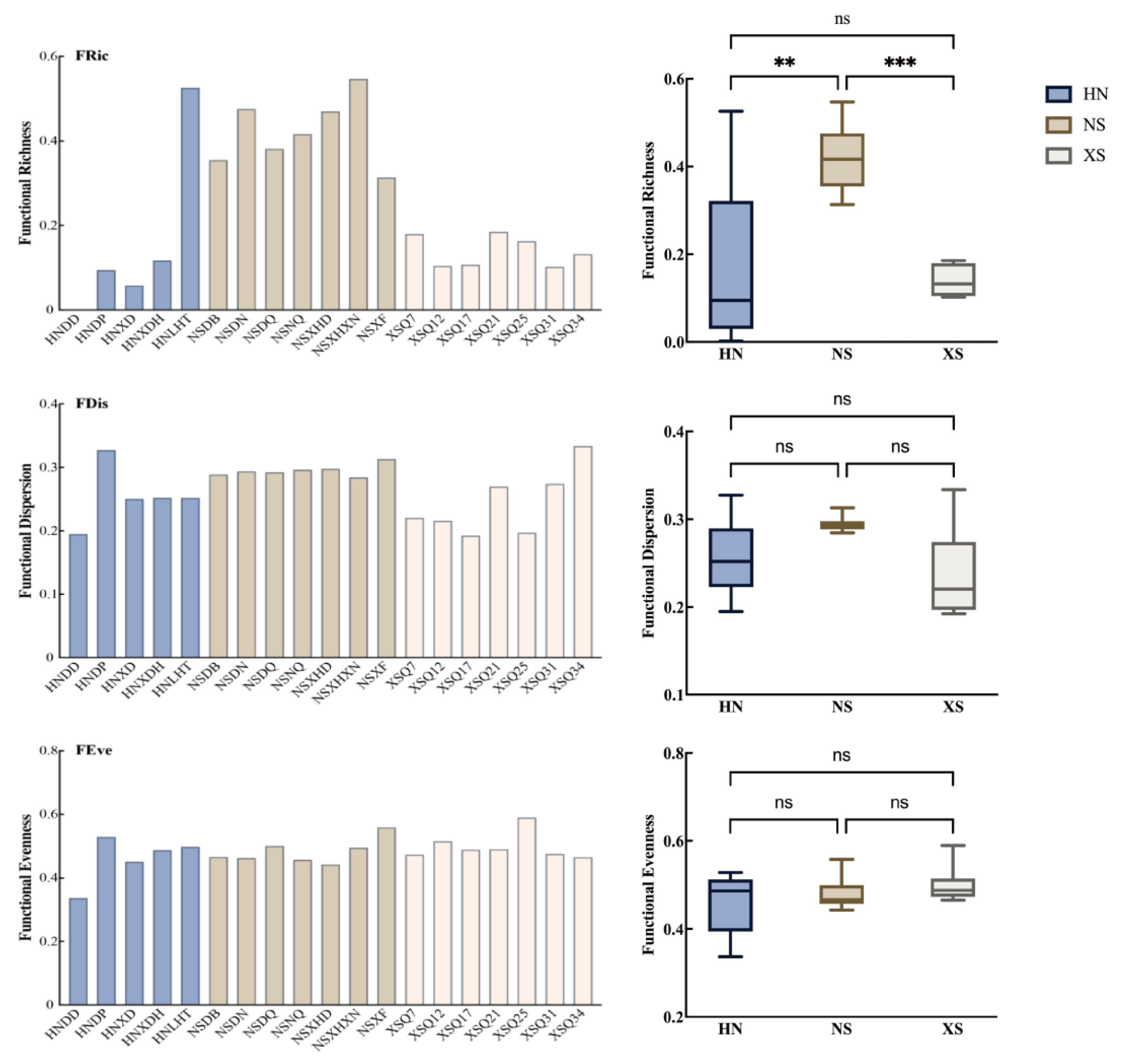
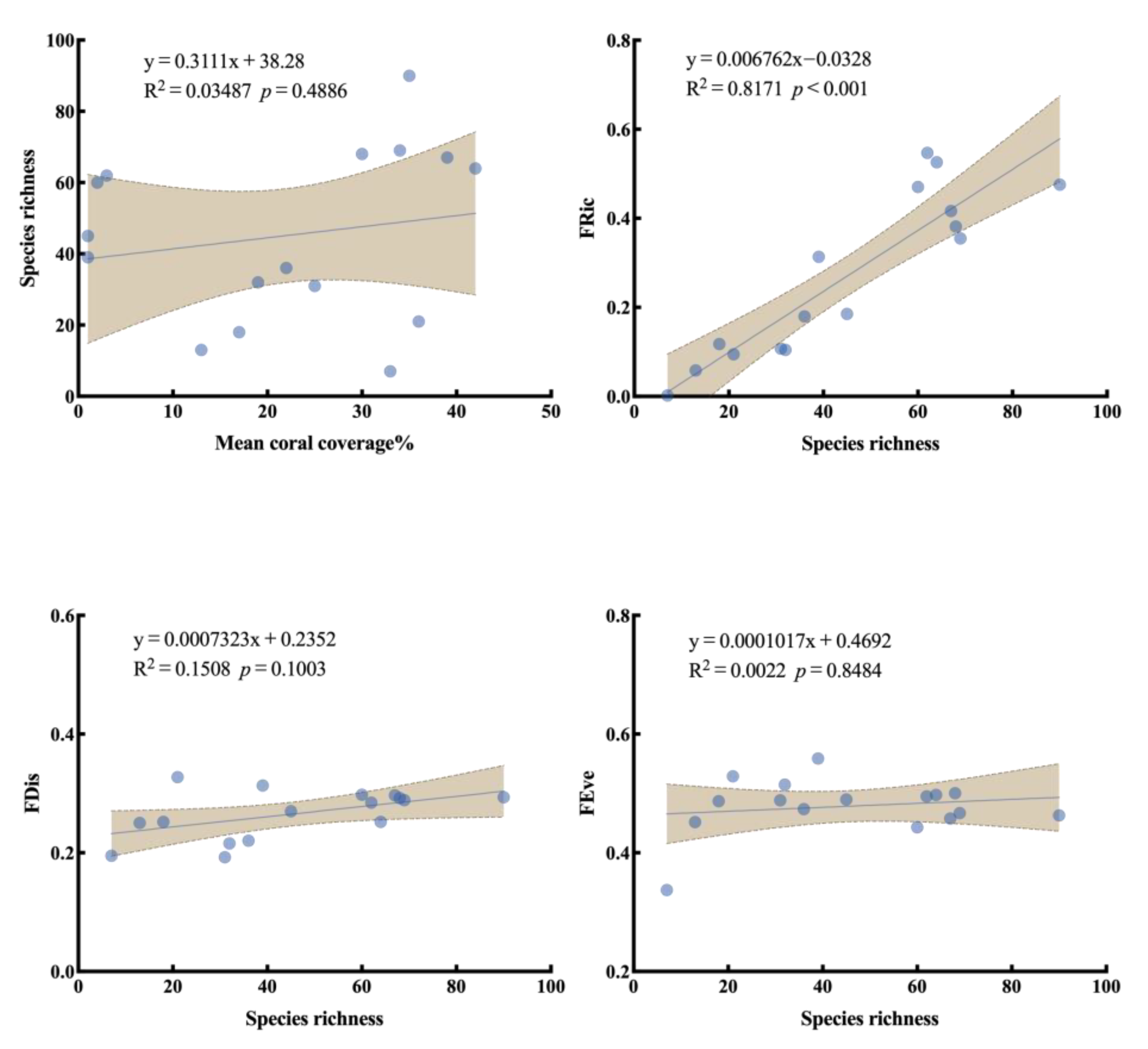
3.4. Functional Diversity and Trait Composition
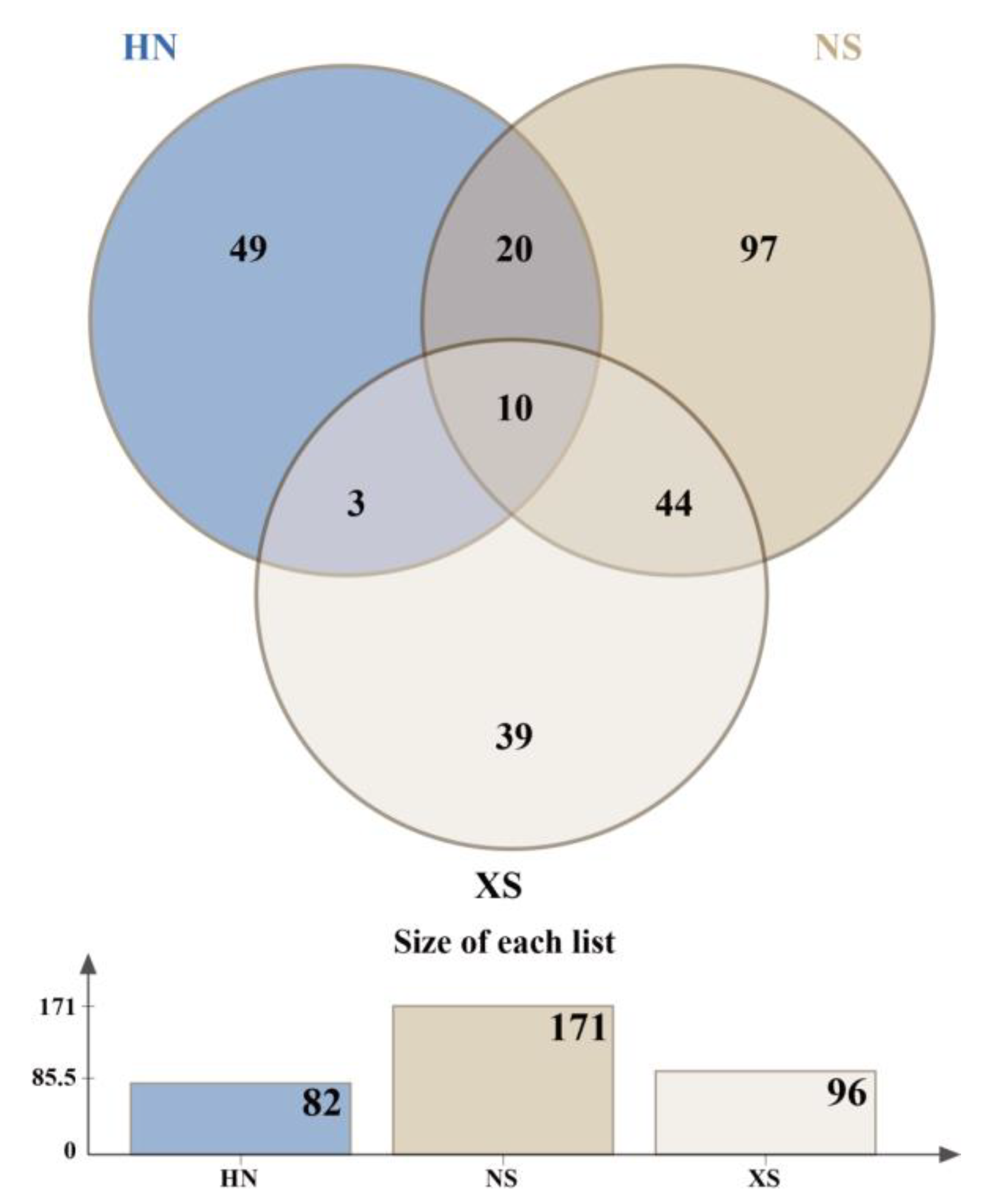
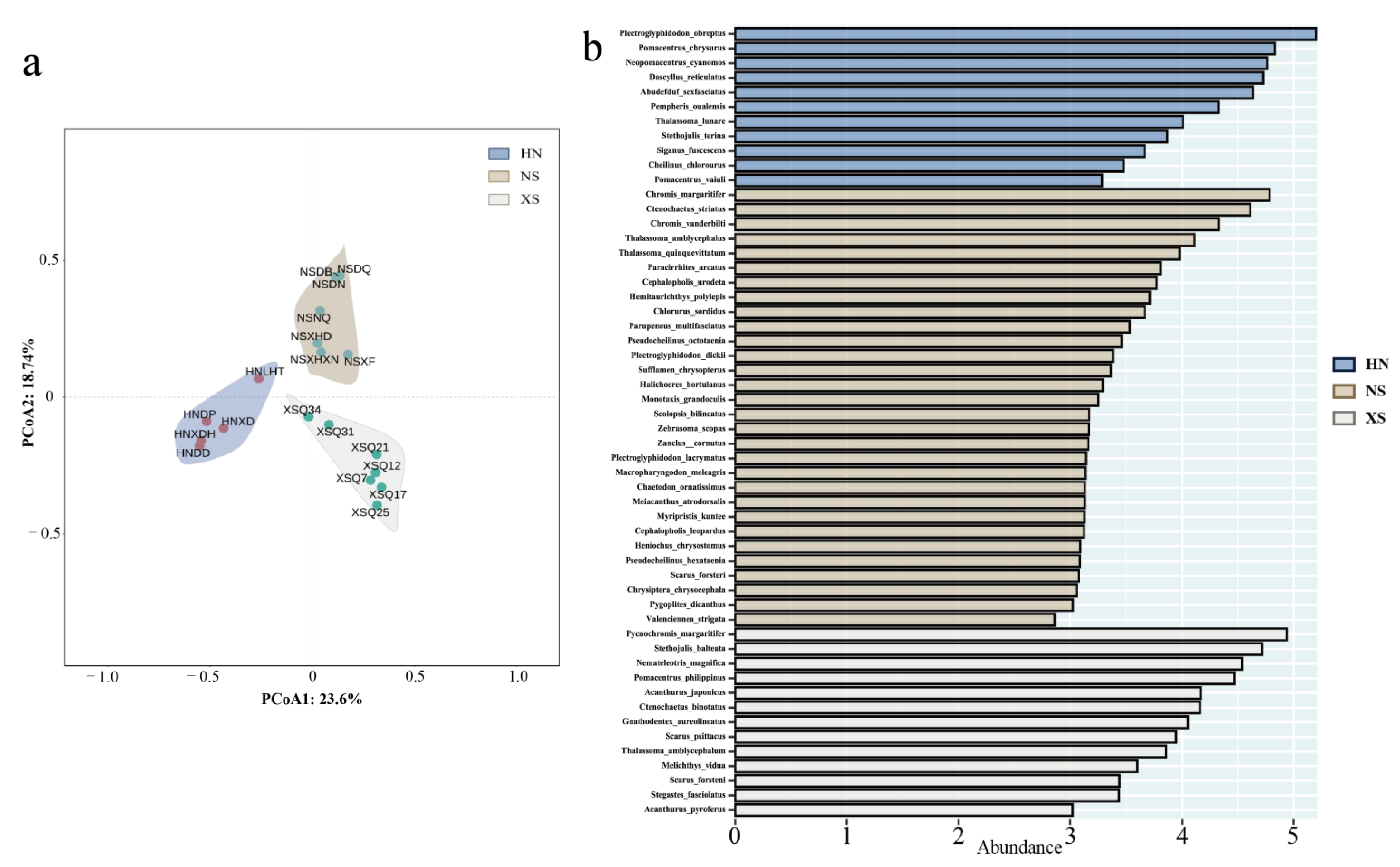
3.5. Community Structure and Indicator Species
4. Discussion
4.1. What Drives the Differences Between Nearshore and Offshore Coral-Reef Fish Communities?
4.2. Are Current Conservation Priorities Aligned with Functional Importance?
4.3. Where Should Conservation Efforts Be Spatially Focused?
4.4. How Can Conservation Strategies Be Refined to Enhance Reef Resilience?
4.5. Limitations and Future Research Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fisher, R.; O’Leary, R.A.; Low-Choy, S.; Mengersen, K.; Knowlton, N.; Brainard, R.E.; Caley, M.J. Species richness on coral reefs and the pursuit of convergent global estimates. Curr. Biol. 2015, 25, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Hoeksema, B.W.; Affendi, Y.A.; Ang, P.O.; Chen, C.A.; Huang, H.; Lane, D.J.W.; Licuanan, W.Y.; Vibol, O.; Vo, S.T.; et al. Conservation of reef corals in the South China Sea based on species and evolutionary diversity. Biodivers. Conserv. 2016, 25, 331–344. [Google Scholar] [CrossRef]
- Dai, X.; Li, Y.; Cai, Y.; Gong, Y.; Zhang, J.; Chen, Z. Variations in fish community structure at the lagoon of yongshu reef, south China Sea between 1999 and 2016–2019. Diversity 2019, 14, 763. [Google Scholar] [CrossRef]
- Zhang, K.; Guo, J.; Xu, Y.; Jiang, Y.E.; Fan, J.; Xu, S.; Chen, Z. Long-term variations in fish community structure under multiple stressors in a semi-closed marine ecosystem in the South China Sea. Sci. Total Environ. 2020, 745, 140892. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, K.; Jiang, Y.E.; Li, Y.J.; Fan, J.T.; Yu, W.M.; Chen, Z.Z. Diversity and structure of demersal fish community over the northern slope in the South China Sea. Front. Mar. Sci. 2022, 9, 809636. [Google Scholar] [CrossRef]
- Huang, H.; Chen, Z.; Huang, L. Status of Coral Reefs in China (2010—2019); Ocean Press: Beijing, China, 2021. [Google Scholar]
- Duprey, N.N.; Yasuhara, M.; Baker, D.M. Reefs of tomorrow: Eutrophication reduces coral biodiversity in an urbanized seascape. Glob. Change Biol. 2016, 22, 3550–3565. [Google Scholar] [CrossRef]
- Guest, J.R.; Tun, K.; Low, J.; Vergés, A.; Marzinelli, E.M.; Campbell, A.H.; Bauman, A.G.; Feary, D.A.; Chou, L.M.; Steinberg, P.D. 27 years of benthic and coral community dynamics on turbid, highly urbanised reefs off Singapore. Sci. Rep. 2016, 6, 36260. [Google Scholar] [CrossRef]
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.H.; Saksena-Taylor, P.; et al. Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Mar. Pollut. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef]
- Luo, Y.; Huang, L.; Lei, X.; Yu, X.; Liu, C.; Jiang, L.; Huang, H. Light availability regulated by particulate organic matter affects coral assemblages on a turbid fringing reef. Mar. Environ. Res. 2022, 177, 105613. [Google Scholar] [CrossRef]
- Shi, Y.; Li, Y.; Lu, W.; Sun, D. Taxonomic diversity of fish species in coral reef area from Dongsha Islands. South China Fish. Sci. 2009, 5, 10–16. [Google Scholar]
- Li, Y.; Zhang, J.; Chen, Z.; Jiang, Y.; Gong, Y.; Cai, Y.; Yang, Y. Study on taxonomic diversity of fish in Zhubi Reef of Nansha Islands. South China Fish. Sci. 2020, 16, 36–41. [Google Scholar]
- Xie, H.; Liu, Y.; Zhao, J.; Li, C.; Shi, J.; Xiao, Y.; Wang, T. Species composition and succession of coral reef fish in Yuzhuo Reef, Xisha Islands. South China Fish. Sci. 2024, 20, 46–55. [Google Scholar]
- Wang, T.; Li, C.; Wang, G.; Zhao, J.; Shi, J.; Xie, H.; Liu, Y. Species composition and succession of coral reef fishes on Qilianyu Island, Xisha Islands. Biodivers. Sci. 2024, 32, 23481. [Google Scholar] [CrossRef]
- Li, Y.Z.; Shi, Y.R.; Ai, H.; Dong, L.N.; Li, N.N.; Li, X.; Gao, T.X. Large scale distribution patterns of taxonomic diversity of fish in coral reef waters, South China Sea. J. Fish. Sci. China 2011, 18, 619–628. [Google Scholar] [CrossRef]
- Lin, X.; Hu, S.; Liu, Y.; Zhang, L.; Huang, H.; Liu, S. Disturbance-mediated changes in coral reef habitat provoke a positive feeding response in a major coral reef detritivore, Ctenochaetus striatus. Front. Mar. Sci. 2021, 8, 682697. [Google Scholar] [CrossRef]
- Lin, X.; Hu, S.; Zhou, Y.; Huang, H.; Zhang, L.; Liu, S. A multiple-methods approach to investigate dietary differences among nominally herbivorous fishes. Mar. Biol. 2023, 170, 134. [Google Scholar] [CrossRef]
- Halpern, B.S.; Floeter, S.R. Functional diversity responses to changing species richness in reef fish communities. Mar. Ecol. Prog. Ser. 2008, 364, 147–156. [Google Scholar] [CrossRef]
- Diamond, J.; Roy, D. Patterns of functional diversity along latitudinal gradients of species richness in eleven fish families. Glob. Ecol. Biogeogr. 2023, 32, 450–465. [Google Scholar] [CrossRef]
- Mouillot, D.; Lepretre, A. A comparison of species diversity estimators. Res. Popul. Ecol. 1999, 41, 203–215. [Google Scholar] [CrossRef]
- Vallina, S.M.; Cermeno, P.; Dutkiewicz, S.; Loreau, M.; Montoya, J.M. Phytoplankton functional diversity increases ecosystem productivity and stability. Ecol. Model. 2017, 361, 184–196. [Google Scholar] [CrossRef]
- Hallett, L.M.; Stein, C.; Suding, K.N. Functional diversity increases ecological stability in a grazed grassland. Oecologia 2017, 183, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, P.; Li, G.; Zhou, D. Relationships between functional diversity and ecosystem functioning: A review. Acta Ecol. Sin. 2014, 34, 85–91. [Google Scholar] [CrossRef]
- Wen, Z.; Zheng, H.; Smith, J.R.; Zhao, H.; Liu, L.; Ouyang, Z. Functional diversity overrides community-weighted mean traits in linking land-use intensity to hydrological ecosystem services. Sci. Total Environ. 2019, 682, 583–590. [Google Scholar] [CrossRef]
- Yeager, L.A.; Deith, M.C.; McPherson, J.M.; Williams, I.D.; Baum, J.K. Scale dependence of environmental controls on the functional diversity of coral reef fish communities. Glob. Ecol. Biogeogr. 2017, 26, 1177–1189. [Google Scholar] [CrossRef]
- Ruiz-Benito, P.; Ratcliffe, S.; Jump, A.S.; Gómez-Aparicio, L.; Madrigal-González, J.; Wirth, C.; Kändler, G.; Lehtonen, A.; Dahlgren, J.; Kattge, J.; et al. Functional diversity underlies demographic responses to environmental variation in European forests. Glob. Ecol. Biogeogr. 2017, 26, 128–141. [Google Scholar] [CrossRef]
- Brock, V.E. A preliminary report on a method of estimating reef fish populations. J. Wildl. Manag. 1954, 18, 297–308. [Google Scholar] [CrossRef]
- Bell, J.D.; Craik, G.J.S.; Pollard, D.A.; Russell, B.C. Estimating length frequency distributions of large reef fish underwater. Coral Reefs 1985, 4, 41–44. [Google Scholar] [CrossRef]
- Bohnsack, J.A.; Bannerot, S.P. A Stationary Visual Census Technique for Quantitatively Assessing Community Structure of Coral Reef Fishes; NOAA Technical Report NFWS 41; NOAA: Silver Spring, MD, USA, 1986; pp. 1–15.
- English, S.; Wilkinson, C.; Baker, V. Survey Manual for Tropical Marine Resources; Australian Institute of Marine Science: Townsville, Australia, 1997.
- Mouillot, D.; Villéger, S.; Parravicini, V.; Kulbicki, M.; Arias-González, J.E.; Bender, M.; Chabanet, P.; Floeter, S.R.; Friedlander, A.; Vigliola, L.; et al. Functional over-redundancy and high functional vulnerability in global fish faunas on tropical reefs. Proc. Natl. Acad. Sci. USA 2014, 111, 13757–13762. [Google Scholar] [CrossRef]
- Mouillot, D.; Graham, N.A.; Villéger, S.; Mason, N.W.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. (Eds.) FishBase; World Wide Web Electronic Publication: Boston, MA, USA, 2025; Version (02/2025); Available online: www.fishbase.org (accessed on 28 March 2025).
- Shao, K.T. Taiwan Fish Database. WWW Web Electronic Publication. 2025. Available online: http://fishdb.sinica.edu.tw (accessed on 28 March 2025).
- Waide, R.B.; Willig, M.R.; Steiner, C.F.; Mittelbach, G.; Gough, L.; Dodson, S.I.; Juday, G.P.; Parmenter, R. The relationship between productivity and species richness. Annu. Rev. Ecol. Syst. 1999, 30, 257–300. [Google Scholar] [CrossRef]
- Keylock, C. Simpson diversity and the Shannon–Wiener index as special cases of a generalized entropy. Oikos 2005, 109, 203–207. [Google Scholar] [CrossRef]
- Jost, L. The relation between evenness and diversity. Diversity 2010, 2, 207–232. [Google Scholar] [CrossRef]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D. Functional diversity. In Encyclopedia of Biodiversity, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2001; pp. 587–596. [Google Scholar]
- Mammola, S.; Carmona, C.P.; Guillerme, T.; Cardoso, P. Concepts and applications in functional diversity. Funct. Ecol. 2021, 35, 1869–1885. [Google Scholar] [CrossRef]
- Laureto, L.M.O.; Cianciaruso, M.V.; Samia, D.S.M. Functional diversity: An overview of its history and applicability. Nat. Conserv. 2015, 13, 112–116. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field–methodology matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Li, S.; Yu, K.; Chen, T.; Shi, Q.; Zhang, H. Assessment of coral bleaching using symbiotic zooxanthellae density and satellite remote sensing data in the Nansha Islands, South China Sea. Chin. Sci. Bull. 2011, 56, 1031–1037. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Z.; Liang, J.; Chen, S.Q.; Zhao, J.M. Analysis on the outbreak period and cause of Acanthaster planci in Xisha Islands in recent 15 years. Chin. Sci. Bull. 2019, 64, 3478–3484. [Google Scholar]
- Reimer, J.D.; Kise, H.; Wee, H.B.; Lee, C.L.; Soong, K. Crown-of-thorns starfish outbreak at oceanic Dongsha Atoll in the northern South China Sea. Mar. Biodivers. 2019, 49, 2495–2497. [Google Scholar] [CrossRef]
- Heng, W.K.; Ho, M.J.; Kuo, C.Y.; Huang, Y.Y.; Ko, C.Y.; Jeng, M.S.; Chen, C.A. Crown-of-thorns starfish outbreak at Taiping island (Itu aba), spratlys, south China Sea. Bull. Mar. Sci. 2022, 98, 101–102. [Google Scholar] [CrossRef]
- Hughes, T.P.; Huang, H.U.I.; Young, M.A. The wicked problem of China’s disappearing coral reefs. Conserv. Biol. 2013, 27, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Huang, L.; McCook, L.J.; Huang, H. Joint protection of a crucial reef ecosystem. Science 2022, 377, 1163. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.S. Post-settlement diet shift of Chlorurus sordidus and Scarus schlegeli (Pisces: Scardiae). Zool. Stud.-Taipei- 2002, 41, 47–58. [Google Scholar]
- Nakai, T.; Sano, M.; Kurokura, H. Feeding habits of the darkfin hind Cephalopholis urodeta (Serranidae) at Iriomote Island, southern Japan. Fish. Sci. 2001, 67, 640–643. [Google Scholar] [CrossRef]
- Yaakub, S.M.; Bellwood, D.R.; van Herwerden, L.; Walsh, F.M. Hybridization in coral reef fishes: Introgression and bi-directional gene exchange in Thalassoma (family Labridae). Mol. Phylogenetics Evol. 2006, 40, 84–100. [Google Scholar] [CrossRef]
- Arai, T.; Amalina, R.; Bachok, Z. Similarity in the feeding ecology of parrotfish (Scaridae) in coral reef habitats of the Malaysian South China Sea, as revealed by fatty acid signatures. Biochem. Syst. Ecol. 2015, 59, 85–90. [Google Scholar] [CrossRef]
- Ho, C.T.; Fu, Y.C.; Sun, C.L.; Kao, S.J.; Jan, R.Q. Plasticity of feeding habits of two Plectroglyphidodon damselfishes on coral reefs in southern Taiwan: Evidence from stomach content and stable isotope analyses. Zool. Stud. 2009, 48, 649–656. [Google Scholar]
- Hata, H.; Ceccarelli, D.M. Farming behaviour of territorial damselfishes. In Biology of Damselfishes; CRC Press: Boca Raton, FL, USA, 2016; pp. 122–152. [Google Scholar]
- Frédérich, B.; Fabri, G.; Lepoint, G.; Vandewalle, P.; Parmentier, E. Trophic niches of thirteen damselfishes (Pomacentridae) at the Grand Récif of Toliara, Madagascar. Ichthyol. Res. 2009, 56, 10–17. [Google Scholar] [CrossRef]
- Clements, K.D.; Choat, J.H. Nutritional ecology of parrotfishes (Scarinae, Labridae). In Biology of Parrotfishes; CRC Press: Boca Raton, FL, USA, 2018; pp. 42–68. [Google Scholar]
- El Rahimi, S.A.; Hendra, E.; Isdianto, A.; Luthfi, O.M. Feeding preference of herbivorous fish: Family Scaridae. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 869, p. 012004. [Google Scholar]
- Wilson, S.K.; Fisher, R.; Pratchett, M.S.; Graham, N.A.J.; Dulvy, N.K.; Turner, R.A.; Cakacaka, A.; Polunin, N.V. Habitat degradation and fishing effects on the size structure of coral reef fish communities. Ecol. Appl. 2010, 20, 442–451. [Google Scholar] [CrossRef]
- Oliver, T.H.; Isaac, N.J.; August, T.A.; Woodcock, B.A.; Roy, D.B.; Bullock, J.M. Declining resilience of ecosystem functions under biodiversity loss. Nat. Commun. 2015, 6, 10122. [Google Scholar] [CrossRef] [PubMed]
- Pratchett, M.S.; Munday, P.L.; Wilson, S.K.; Graham, N.A.; Cinner, J.E.; Bellwood, D.R.; Jones, G.P.; Polunin, N.V.C.; Mcclanahan, T.R. Effects of climate-induced coral bleaching on coral-reef fishes—Ecological and economic consequences. In Oceanography and Marine Biology; CRC Press: Boca Raton, FL, USA, 2008; pp. 257–302. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Sites | Code | Lon/E | Lat/N | Depth | Coral Coverage |
|---|---|---|---|---|---|---|
| Sanya, Hainan (HN) | Dongdao Island | DD | 109.42 | 18.22 | 3 m~6 m | 33% |
| Dongpai | DP | 109.3 | 18.15 | 3 m~9 m | 36% | |
| Luhuitou | LHT | 109.48 | 18.2 | 3 m~6 m | 42% | |
| Xidao Island | XD | 109.37 | 18.23 | 3 m~9 m | 13% | |
| Xiaodonghai | XDH | 109.5 | 18.2 | 3 m~9 m | 17% | |
| Nansha Islands (NS) | Meiji Island | DB | 115.56 | 9.86 | 3 m~15 m | 34% |
| DN | 3 m~15 m | 35% | ||||
| DQ | 3 m~15 m | 30% | ||||
| NQ | 3 m~15 m | 39% | ||||
| XF | 3 m~15 m | 1% | ||||
| XHD | 3 m~15 m | 2% | ||||
| XHXN | 3 m~15 m | 3% | ||||
| Xisha Island (XS) | Zhaoshu Island | Q07 | 112.27 | 16.97 | 3 m~15 m | 22% |
| Q12 | 3 m~15 m | 19% | ||||
| Q17 | 3 m~15 m | 25% | ||||
| Lingyang Island | Q21 | 111.6 | 16.47 | 3 m~15 m | 1% | |
| Q25 | 3 m~15 m | - | ||||
| Q31 | 3 m~15 m | - | ||||
| Q34 | 3 m~15 m | - |
| Sites | Total Individuals (n) | F1 | F2 | Sample Coverage |
|---|---|---|---|---|
| HNDD | 107 | 4 | 0 | 0.96 |
| HNDP | 156 | 7 | 4 | 0.96 |
| HNLHT | 1642 | 18 | 7 | 0.99 |
| HNXD | 63 | 6 | 3 | 0.91 |
| HNXDH | 215 | 6 | 2 | 0.97 |
| NSDB | 992 | 25 | 10 | 0.97 |
| NSDN | 1813 | 26 | 12 | 0.99 |
| NSDQ | 966 | 22 | 14 | 0.98 |
| NSNQ | 1992 | 15 | 16 | 0.99 |
| NSXF | 531 | 10 | 6 | 0.98 |
| NSXHD | 1360 | 19 | 11 | 0.99 |
| NSXHXN | 1343 | 23 | 6 | 0.98 |
| XSQ07 | 596 | 12 | 4 | 0.98 |
| XSQ12 | 428 | 12 | 6 | 0.97 |
| XSQ17 | 751 | 14 | 5 | 0.98 |
| XSQ21 | 520 | 15 | 10 | 0.97 |
| XSQ25 | 420 | 4 | 5 | 0.99 |
| XSQ31 | 261 | 7 | 8 | 0.97 |
| XSQ34 | 605 | 4 | 2 | 0.99 |
| Trait | Slope | Intercept | R2 | p_Value |
|---|---|---|---|---|
| size_0–7 cm | 0.001 | −0.0253 | 0.6063 | *** |
| mob_mobile within a reef | 0.0051 | −0.0363 | 0.3945 | ** |
| gre_solitary | −0.0045 | 0.4514 | 0.3739 | ** |
| diet_sessile invertebrates | 0.0001 | −0.0013 | 0.311 | * |
| diet_herbivorous-detritivorous | −0.0023 | 0.2098 | 0.2485 | * |
| mob_sedentary | −0.004 | 0.2639 | 0.2481 | * |
| act_diurnal-noctural | 0.0001 | −0.0009 | 0.1694 | ns |
| gre_large group | 0.0027 | 0.0379 | 0.0934 | ns |
| size_15.1–30 cm | 0.0016 | 0.1842 | 0.0774 | ns |
| size_7.1–15 cm | −0.0021 | 0.7483 | 0.0707 | ns |
| lev_pelagic | 0.0001 | −0.0012 | 0.0683 | ns |
| size_50.1–80 cm | 0.0002 | 0.0031 | 0.0627 | ns |
| diet_planktivorous | 0.0026 | 0.2453 | 0.0626 | ns |
| diet_piscivorous | 0.0008 | 0.0417 | 0.0622 | ns |
| size_30.1–50 cm | −0.0007 | 0.0886 | 0.0595 | ns |
| gre_medium group | 0.0007 | 0.0256 | 0.0525 | ns |
| lev_bentho-pelagic | −0.0004 | 0.9904 | 0.0323 | ns |
| act_nocturnal | −0.0005 | 0.0675 | 0.032 | ns |
| diet_omnivorous | −0.0012 | 0.4076 | 0.0257 | ns |
| act_diurnal | 0.0004 | 0.9334 | 0.0231 | ns |
| lev_benthic | 0.0003 | 0.0108 | 0.0204 | ns |
| gre_small group | 0.0011 | 0.4399 | 0.0181 | ns |
| mob_mobile among reefs | −0.0011 | 0.7724 | 0.0108 | ns |
| diet_macroalgal herbivorous | 0 | 0.0043 | 0.0094 | ns |
| gre_pairing | 0.0001 | 0.0452 | 0.001 | ns |
| diet_mobile invertebrates | 0 | 0.0924 | 0.0003 | ns |
| size_ > 80 cm | 0 | 0.0011 | 0.0002 | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Hu, S.; Huang, L.; Lin, X.; Huang, H.; Liu, S. Spatial Patterns and Functional Diversity of Nearshore and Offshore Coral-Reef Fish Communities in the South China Sea. Diversity 2025, 17, 432. https://doi.org/10.3390/d17060432
Zhang C, Hu S, Huang L, Lin X, Huang H, Liu S. Spatial Patterns and Functional Diversity of Nearshore and Offshore Coral-Reef Fish Communities in the South China Sea. Diversity. 2025; 17(6):432. https://doi.org/10.3390/d17060432
Chicago/Turabian StyleZhang, Chen, Simin Hu, Lintao Huang, Xianzhi Lin, Hui Huang, and Sheng Liu. 2025. "Spatial Patterns and Functional Diversity of Nearshore and Offshore Coral-Reef Fish Communities in the South China Sea" Diversity 17, no. 6: 432. https://doi.org/10.3390/d17060432
APA StyleZhang, C., Hu, S., Huang, L., Lin, X., Huang, H., & Liu, S. (2025). Spatial Patterns and Functional Diversity of Nearshore and Offshore Coral-Reef Fish Communities in the South China Sea. Diversity, 17(6), 432. https://doi.org/10.3390/d17060432