
Macrofaunal Communities Associated with Ecklonia radiata Holdfast Along the Southeast Coast of South Africa

Abstract
1. Introduction
2. Materials and Methods
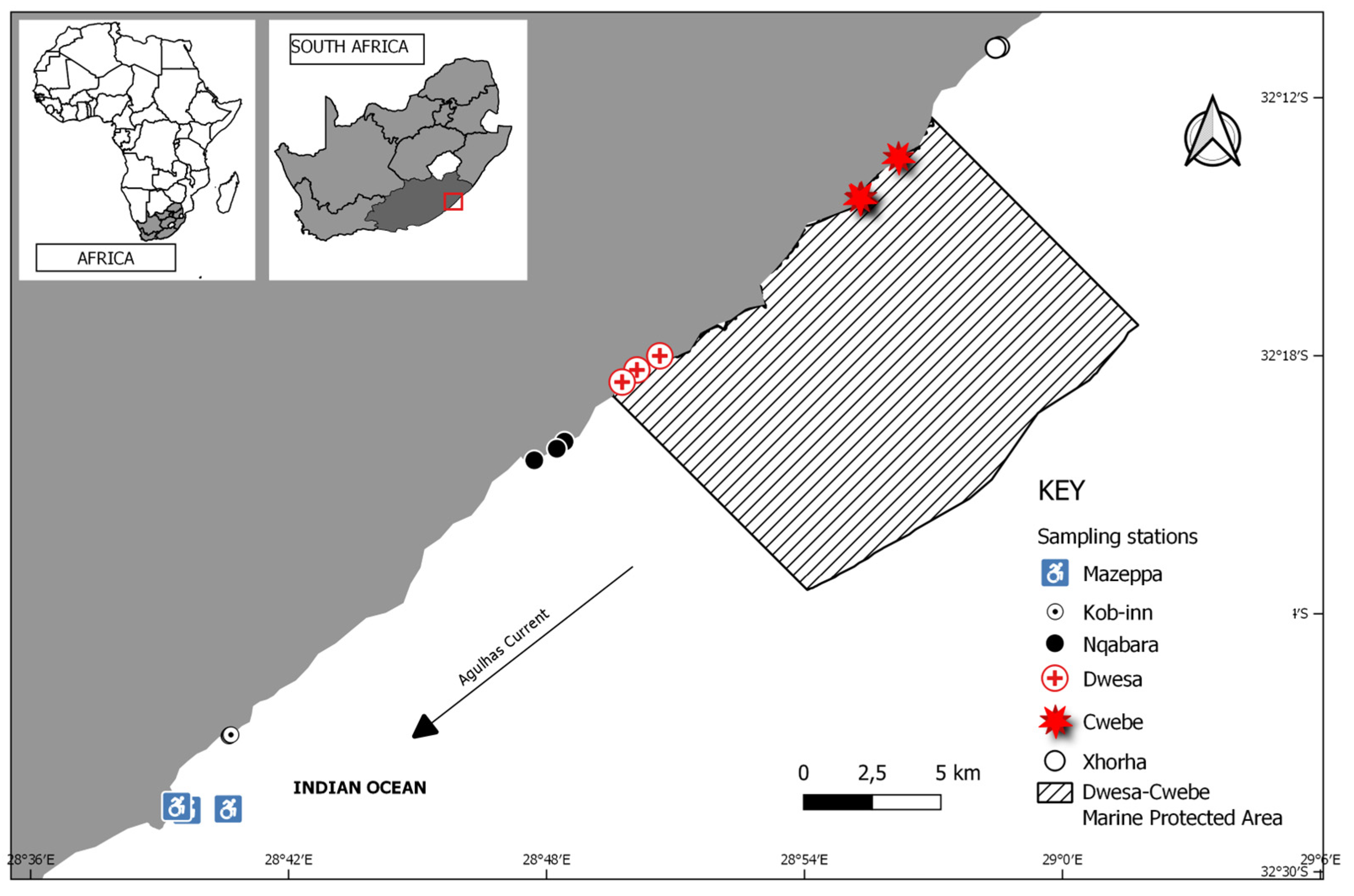
2.1. Study Area
Laboratory Processing
2.2. Data Analysis
2.2.1. Local Environmental Conditions
2.2.2. Statistical Analyses
3. Results
3.1. Univariate Community Variables
3.1.1. Univariate Patterns
3.1.2. Species Richness
3.1.3. Shannon’s Index
3.1.4. Abundance
3.1.5. Evenness
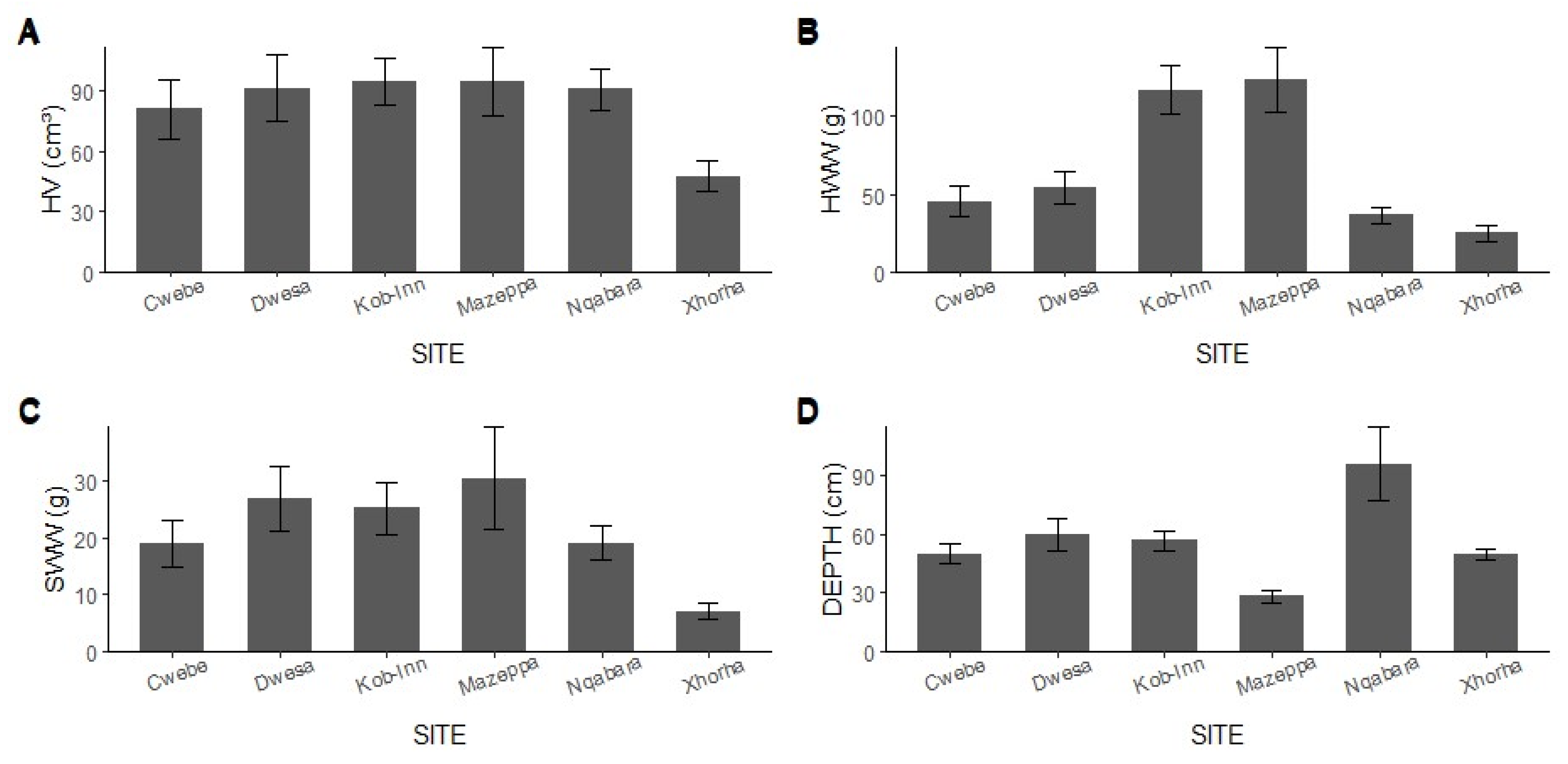
3.2. Local Environmental Conditions vs. Ecological Indices
3.3. Multivariate Patterns
3.3.1. Spatial Autocorrelation
3.3.2. Beta Diversity
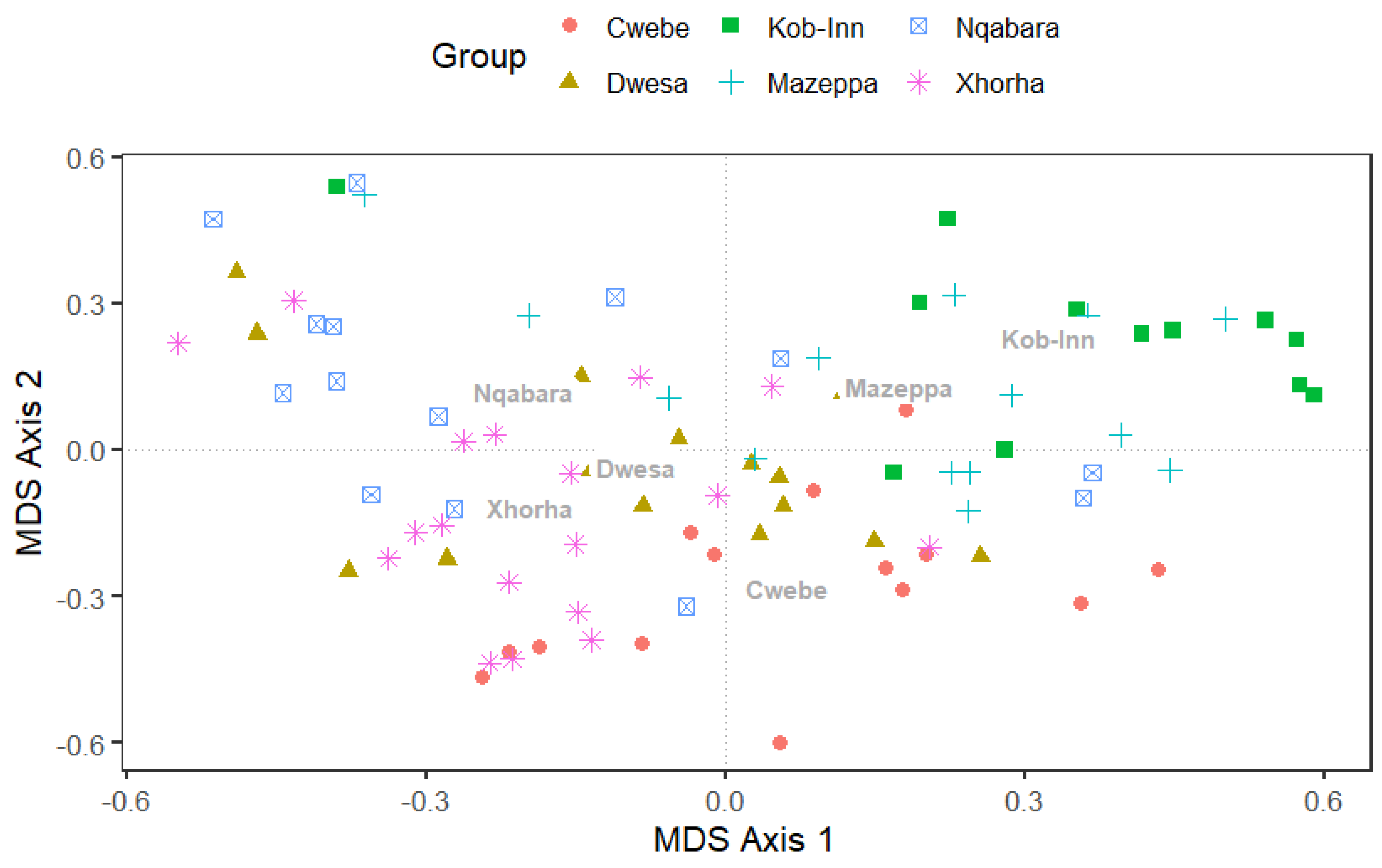
3.3.3. Multidimensional Scaling (MDS) and PERMANOVA
4. Discussion
4.1. Spatial Variation in Local Environmental Conditions Across the Kelp-Associated Coastal Systems
4.2. Alpha Diversity
4.3. Relationship Between Local Environmental Conditions and Biodiversity Patterns
4.4. Beta Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Darwin, C. The Origin of Species [On the Origin of Species]; PF Collier & Son: New York, NY, USA, 1909. [Google Scholar]
- Jones, D. Variation in the trophic structure and species composition of some invertebrate communities in polluted kelp forests in the North Sea. Mar. Biol. 1973, 20, 351–365. [Google Scholar] [CrossRef]
- Teagle, H.; Moore, P.J.; Jenkins, H.; Smale, D.A. Spatial variability in the diversity and structure of faunal assemblages associated with kelp holdfasts (Laminaria hyperborea) in the northeast Atlantic. PLoS ONE 2018, 13, e0200411. [Google Scholar] [CrossRef]
- Wernberg, T.; Coleman, M.A.; Babcock, R.C.; Bell, S.Y.; Bolton, J.J.; Connell, S.D.; Hurd, C.L.; Johnson, C.R.; Marzinelli, E.M.; Shears, N.T. Biology and Ecology of the Globally Significant Kelp Ecklonia Radiata; Oceanography and marine biology; Taylor & Francis: Abingdon, UK, 2019. [Google Scholar]
- Carbajal, P.; Gamarra Salazar, A.; Moore, P.J.; Pérez-Matus, A. Different kelp species support unique macroinvertebrate assemblages, suggesting the potential community-wide impacts of kelp harvesting along the Humboldt current system. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 14–27. [Google Scholar] [CrossRef]
- Teagle, H.; Hawkins, S.J.; Moore, P.J.; Smale, D.A. The role of kelp species as biogenic habitat formers in coastal marine ecosystems. J. Exp. Mar. Biol. Ecol. 2017, 492, 81–98. [Google Scholar] [CrossRef]
- Wernberg, T. Holdfast aggregation in relation to morphology, age, attachment and drag for the kelp Ecklonia radiata. Aquat. Bot. 2005, 82, 168–180. [Google Scholar] [CrossRef]
- Anderson, M.J.; Connell, S.D.; Gillanders, B.M.; Diebel, C.E.; Blom, W.M.; Saunders, J.E.; Landers, T.J. Relationships between taxonomic resolution and spatial scales of multivariate variation. J. Anim. Ecol. 2005, 74, 636–646. [Google Scholar] [CrossRef]
- Anderson, M.J.; Diebel, C.E.; Blom, W.M.; Landers, T.J. Consistency and variation in kelp holdfast assemblages: Spatial patterns of biodiversity for the major phyla at different taxonomic resolutions. J. Exp. Mar. Biol. Ecol. 2005, 320, 35–56. [Google Scholar] [CrossRef]
- Smith, S.D.; Simpson, R.D.; Cairns, S.C. The macrofaunal community of Ecklonia radiata holdfasts: Description of the faunal assemblage and variation associated with differences in holdfast volume. Aust. J. Ecol. 1996, 21, 81–95. [Google Scholar] [CrossRef]
- Blight, A.; Thompson, R. Epibiont species richness varies between holdfasts of a northern and a southerly distributed kelp species. J. Mar. Biol. Assoc. U. K. 2008, 88, 469–475. [Google Scholar] [CrossRef]
- Fernández, C.; Pineiro-Corbeira, C.; Barrientos, S.; Barreiro, R. Could the annual Saccorhiza polyschides replace a sympatric perennial kelp (Laminaria ochroleuca) when it comes to supporting the holdfast-associated fauna? Mar. Environ. Res. 2022, 182, 105772. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.D. Evaluating stress in rocky shore and shallow reef habitats using the macrofauna of kelp holdfasts. J. Aquat. Ecosyst. Stress Recovery 2000, 7, 259–272. [Google Scholar] [CrossRef]
- Smith, S.D.; Simpson, R.D. Effects of pollution on holdfast macrofauna of the kelp Ecklonia radiata: Discrimination at different taxonomic levels. Mar. Ecol. Prog. Ser. Oldendorf 1993, 96, 199–208. [Google Scholar] [CrossRef]
- Teagle, H.; Smale, D.A. Climate-driven substitution of habitat-forming species leads to reduced biodiversity within a temperate marine community. Divers. Distrib. 2018, 24, 1367–1380. [Google Scholar] [CrossRef]
- Thompson, R.; Crowe, T.; Hawkins, S. Rocky intertidal communities: Past environmental changes, present status and predictions for the next 25 years. Environ. Conserv. 2002, 29, 168–191. [Google Scholar] [CrossRef]
- Zahn, L.A.; Claisse, J.T.; Williams, J.P.; Williams, C.M.; Pondella, D.J. The biogeography and community structure of kelp forest macroinvertebrates. Mar. Ecol. 2016, 37, 770–785. [Google Scholar] [CrossRef]
- Shelamoff, V.; Layton, C.; Tatsumi, M.; Cameron, M.J.; Wright, J.T.; Johnson, C.R. Patch size and density of canopy-forming kelp modify influences of ecosystem engineering on understorey algal and sessile invertebrate assemblages. Mar. Ecol. Prog. Ser. 2019, 632, 59–79. [Google Scholar] [CrossRef]
- Blamey, L.K. Ecosystem Effects of a Rock-Lobster ‘Invasion’: Comparitive and Modelling Approaches. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2010. [Google Scholar]
- Steyn, C.; Shannon, L.J.; Blamey, L.K. Changes in food-web structure and energy flow in kelp forest ecosystems on the south-west coast of South Africa following the invasion of Jasus lalandii. Food Webs 2021, 28, e00200. [Google Scholar] [CrossRef]
- Bué, M.; Smale, D.A.; Natanni, G.; Marshall, H.; Moore, P.J. Multiple-scale interactions structure macroinvertebrate assemblages associated with kelp understory algae. Divers. Distrib. 2020, 26, 1551–1565. [Google Scholar] [CrossRef]
- Bustamante, R.; Branch, G.; Eekhout, S. The influences of physical factors on the distribution and zonation patterns of South African rocky-shore communities. Afr. J. Mar. Sci. 1997, 18, 119–136. [Google Scholar] [CrossRef]
- Mcquaid, C.D.; Dower, K.M. Enhancement of habitat heterogeneity and species richness on rocky shores inundated by sand. Oecologia 1990, 84, 142–144. [Google Scholar] [CrossRef]
- Ríos, C.; Arntz, W.E.; Gerdes, D.; Mutschke, E.; Montiel, A. Spatial and temporal variability of the benthic assemblages associated to the holdfasts of the kelp Macrocystis pyrifera in the Straits of Magellan, Chile. Polar Biol. 2007, 31, 89–100. [Google Scholar] [CrossRef]
- Bolton, J.J.; Anderson, R.J.; Smit, A.J.; Rothman, M.D. South African kelp moving eastwards: The discovery of Ecklonia maxima (Osbeck) Papenfuss at De Hoop Nature Reserve on the south coast of South Africa. Afr. J. Mar. Sci. 2012, 34, 147–151. [Google Scholar] [CrossRef]
- Dunga, L.; Lück-Vogel, M.; Blamey, L.K.; Bolton, J.; Rothman, M.; Desmet, P.; Sink, K. Mapping South Africa’s canopy-forming kelp forests using low-cost, high-resolution Sentinel-2 imagery. Estuar. Coast. Shelf Sci. 2024, 310, 108989. [Google Scholar] [CrossRef]
- Anderson, R.; Carrick, P.; Levitt, G.; Share, A. Holdfasts of adult kelp Ecklonia maxima provide refuges from grazing for recruitment of juvenile kelps. Mar. Ecol. Prog. Ser. 1997, 159, 265–273. [Google Scholar] [CrossRef]
- Beviss-Challinor, M.; Field, J. Analysis of a benthic community food web using isotopically labeled potential food. Mar. Ecol. Prog. Ser. 1982, 9, 223–230. [Google Scholar] [CrossRef]
- Katharoyan, C.; Peer, N.; Landschoff, J.; Griffiths, C.L.; Samaai, T.; Beeslaar, D. Kelp holdfasts in the Great African Seaforest provide habitat for diverse assemblages of macroinvertebrates. Aquat. Biol. 2024, 33, 33–45. [Google Scholar] [CrossRef]
- Nkohla, N.; Dlaza, T. Site-and habitat-dependent variations in the diversity of polychaetes associated with golden kelp Ecklonia radiata holdfasts along the southeast coast of South Africa. Afr. J. Mar. Sci. 2024, 46, 41–54. [Google Scholar] [CrossRef]
- Branch, G. Living Shores; Penguin Random House South Africa: New York, NY, USA, 2018. [Google Scholar]
- Branch, G.; Branch, M.; Bannister, A. The Living Shores of Southern Africa; Penguine Random House South Africa: New York, NY, USA, 1981. [Google Scholar]
- Branch, G.; Branch, M.; Griffiths, C.; Beckley, L. Two Oceans: A Guide to The Marine Life of Southern Africa; Penguin Random House South Africa: New York, NY, USA, 2022. [Google Scholar]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa. Part 1. Errantia; BM(NH): London, UK, 1967. [Google Scholar]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa. Part 2. Sedentaria; BM(NH): London, UK, 1967. [Google Scholar]
- Day, J.H. A Guide to Marine Life on South African Shores; University of Cape Town: Cape Town, South Africa, 1969. [Google Scholar]
- Griffiths, C. Crustacean systematics in South Africa—Status and historical overview. Trans. R. Soc. S. Afr. 1999, 54, 43–52. [Google Scholar] [CrossRef]
- Griffiths, C.L. The Gammaridean and Caprellid Amphipoda of Southern Africa. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 1974. [Google Scholar]
- Kensley, B.F. Isopoda. Guides to the Freshwater Invertebrates of Southen Africa. Volume 4: Crustacea III, Bathynellacea, Amphipoda, Isopoda, Spelaeogriphacea, Tanaidacea and Decapoda. Afr. J. Aquat. Sci. 2001, 27, 95. [Google Scholar]
- Olbers, J.M. Taxonomy, Biodiversity and Biogeography of the Brittle Stars (Echinodermata: Ophiuroidea) of South Africa. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2016. [Google Scholar]
- Simon, C.; Kara, J.; Clarke, D.; Sedick, S. Revisiting ‘A monograph on the Polychaeta of southern Africa’: Establishing taxonomic research priorities in southern Africa. Afr. J. Mar. Sci. 2022, 44, 83–100. [Google Scholar] [CrossRef]
- Zuur, A.; Ieno, E.; Elphick, C. Zuur AF, Ieno EN, Elphick CS. A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1: 3–14. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Clarke, K.; Gorley, R. PRIMER Version 7: User Manual/Tutorial; PRIMER-E: Auckland, New Zealand, 2015; Volume 192. [Google Scholar]
- Baldrich, Á.M.; Rodríguez-Villegas, C.; Buschmann, A.H. The giant kelp holdfasts macroinvertebrate assemblages: Towards benthic management and conservation using alpha and beta diversity in Northern Patagonia. Reg. Stud. Mar. Sci. 2024, 80, 103896. [Google Scholar]
- Ronowicz, M.; Kukliński, P.; Włodarska-Kowalczuk, M. Diversity of kelp holdfast-associated fauna in an Arctic fjord-inconsistent responses to glacial mineral sedimentation across different taxa. Estuar. Coast. Shelf Sci. 2018, 205, 100–109. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.C. metan: An R package for multi-environment trial analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Dray, S.; Pélissier, R.; Couteron, P.; Fortin, M.-J.; Legendre, P.; Peres-Neto, P.R.; Bellier, E.; Bivand, R.; Blanchet, F.G.; De Cáceres, M.; et al. Community ecology in the age of multivariate multiscale spatial analysis. Ecol. Monogr. 2012, 82, 257–275. [Google Scholar] [CrossRef]
- Landeiro, V.L.; Franz, B.; Heino, J.; Siqueira, T.; Bini, L.M. Species-poor and low-lying sites are more ecologically unique in a hyperdiverse Amazon region: Evidence from multiple taxonomic groups. Divers. Distrib. 2018, 24, 966–977. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; KINDT, R.; Legendre, P.; O’hara, R.G.; Simpson, G.; Solymos, P.; Stevens, H.; Wagner, H. Multivariate Analysis of Ecological Communities in R: Vegan Tutorial; R Package Version 1.7; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Quensen, J.; Simpson, G.; Oksanen, J. ggordiplots: Make Ggplot Versions of Vegans Ordiplots; R Package Version; R Core Team: Vienna, Austria, 2018; Volume 8. [Google Scholar]
- Awad, A.A.; Griffiths, C.L.; Turpie, J.K. Distribution of South African marine benthic invertebrates applied to the selection of priority conservation areas. Divers. Distrib. 2002, 8, 129–145. [Google Scholar] [CrossRef]
- Guilhaumon, F.; Basset, A.; Barbone, E.; Mouillot, D. Species–area relationships as a tool for the conservation of benthic invertebrates in Italian coastal lagoons. Estuar. Coast. Shelf Sci. 2012, 114, 50–58. [Google Scholar] [CrossRef]
- Gundersen, H.; Rinde, E.; Bekkby, T.; Hancke, K.; Gitmark, J.K.; Christie, H. Variation in population structure and standing stocks of kelp along multiple environmental gradients and implications for ecosystem services. Front. Mar. Sci. 2021, 8, 578629. [Google Scholar] [CrossRef]
- Wampler, P. Rivers and Streams—Water and Sediment in Motion. Nat. Educ. Knowl. 2012, 3, 18. [Google Scholar]
- Anderson, R.; Stegenga, H.; Bolton, J. Seaweeds of the South African South Coast; World Wide Web Electronic Publication, University of Cape Town: Cape Town, South Africa, 2016; Available online: http://southafrseaweeds.uct.ac.za (accessed on 20 December 2024).
- Fowles, A.E.; Edgar, G.J.; Stuart-Smith, R.D.; Kirkpatrick, J.B.; Hill, N.; Thomson, R.J.; Strain, E.M. Effects of pollution from anthropogenic point sources on the recruitment of sessile estuarine reef biota. Front. Mar. Sci. 2018, 5, 417. [Google Scholar] [CrossRef]
- Jones, D. Ecological studies on macroinvertebrate populations associated with polluted kelp forests in the North Sea. Helgol. Mar. Res. 1971, 22, 417–441. [Google Scholar] [CrossRef]
- Wilsey, B.J.; Potvin, C. Biodiversity and ecosystem functioning: Importance of species evenness in an old field. Ecology 2000, 81, 887–892. [Google Scholar] [CrossRef]
- Coleman, M.A.; Vytopil, E.; Goodsell, P.J.; Gillanders, B.M.; Connell, S.D. Diversity and depth-related patterns of mobile invertebrates associated with kelp forests. Mar. Freshw. Res. 2007, 58, 589–595. [Google Scholar] [CrossRef]
- Hamylton, S.; Barnes, R. The effect of sampling effort on spatial autocorrelation in macrobenthic intertidal invertebrates. Hydrobiologia 2018, 811, 239–250. [Google Scholar] [CrossRef]
- Sor, R.; Legendre, P.; Lek, S. Uniqueness of sampling site contributions to the total variance of macroinvertebrate communities in the Lower Mekong Basin. Ecol. Indic. 2018, 84, 425–432. [Google Scholar] [CrossRef]
- Lasiak, T. The shellfish-gathering practices of indigenous coastal people in Transkei: Patterns, preferences and perceptions. S. Afr. J. Ethnol. 1993, 16, 115–120. [Google Scholar]
- Mills, C.G. Shellfish Utilization and Its Effect on Rocky Shore Biota in Transkei. Master’s Thesis, Faculty of Science, Department of Environmental and Geographical Science, University of Cape Town, Cape Town, South Africa, 1985. [Google Scholar]
- Baliwe, N.G.; Pfaff, M.C.; Branch, G.M. Assessing the effects of no-take zones in a marine protected area spanning two ecoregions and rock substrate types. Front. Mar. Sci. 2022, 9, 893260. [Google Scholar] [CrossRef]
- Kirkman, S.; Mann, B.; Sink, K.; Adams, R.; Livingstone, T.; Mann-Lang, J.; Pfaff, M.; Samaai, T.; Van Der Bank, M.; Williams, L. Evaluating the evidence for ecological effectiveness of South Africa’s marine protected areas. Afr. J. Mar. Sci. 2021, 43, 389–412. [Google Scholar] [CrossRef]
- Ruhí, A.; Datry, T.; Sabo, J.L. Interpreting beta-diversity components over time to conserve metacommunities in highly dynamic ecosystems. Conserv. Biol. 2017, 31, 1459–1468. [Google Scholar] [CrossRef]
- Dubois, R.; Proulx, R.; Pellerin, S. Ecological uniqueness of plant communities as a conservation criterion in lake-edge wetlands. Biol. Conserv. 2020, 243, 108491. [Google Scholar]
- Esquete, P.; Bamber, R.; Moreira, J.; Troncoso, J. Redescription and postmarsupial development of Apseudopsis latreillii (Crustacea: Tanaidacea). J. Mar. Biol. Assoc. U. K. 2012, 92, 1023–1041. [Google Scholar] [CrossRef]
- Fraser, C.I.; Nikula, R.; Waters, J.M. Oceanic rafting by a coastal community. Proc. R. Soc. B Biol. Sci. 2011, 278, 649–655. [Google Scholar]
- Kensley, B.F. Guide to the Marine Isopods of Southern Africa; The Rustica Press: Cape Town, South Africa, 1978. [Google Scholar]
- Bruce, N.L. Revision of the isopod crustacean genus Mothocya Costa, in Hope, 1851 (Cymothoidae: Flabellifera), parasitic on marine fishes. J. Nat. Hist. 1986, 20, 1089–1192. [Google Scholar] [CrossRef]
- Deheyn, D.; Jangoux, M. Colour varieties as sibling species in the polychromatic ophiuroid Amphipholis squamata (Echinodermata): Evidence from inheritance of body colour and luminescence characters. J. Exp. Mar. Biol. Ecol. 1999, 234, 219–234. [Google Scholar] [CrossRef]
- Jones, M.; Smaldon, G. Aspects of the biology of a population of the cosmopolitan brittlestar Amphipholis squamata (Echinodermata) from the Firth of Forth, Scotland. J. Nat. Hist. 1989, 23, 613–625. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GLMM | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fixed Effects | Estimate | Standard Error | z Value | p Value | |||||
| Species richness | |||||||||
| Site (Mazeppa) | 0.63 | 0.18 | 3.63 | 0.001 | |||||
| AIC: 521.8 BIC:541.7 | |||||||||
| Species abundance | |||||||||
| Site (Dwesa) | 0.81 | 0.30 | 2.71 | 0.007 | |||||
| Site (Mazeppa) | 0.94 | 0.30 | 3.30 | 0.001 | |||||
| AIC: 765.6 BIC:785.4 | |||||||||
| Multiple Comparison of Means: Tukey Contrasts | |||||||||
| Pairs | Estimate | Standard Error | z Value | p Value | Indices | ||||
| Mazeppa vs. Cwebe | 0.63 | 0.17 | 3.63 | 0.004 | Species Richness | ||||
| Mazeppa vs. Kob-Inn | 0.81 | 0.18 | 4.47 | 0.001 | |||||
| Xhorha vs. Mazeppa | −0.50 | 0.17 | −2.92 | 0.041 | |||||
| Mazeppa vs. Cwebe | 0.94 | 0.30 | 3.30 | 0.001 | Species Abundance | ||||
| Kob-Inn vs. Dwesa | −1.07 | 0.30 | −3.54 | 0.001 | |||||
| Mazeppa vs. Kob-Inn | 1.21 | 0.29 | 4.11 | 0.001 | |||||
| Nqabara vs. Mazeppa | −0.93 | 0.31 | −3.04 | 0.029 | |||||
| Xhora vs. Mazeppa | −0.86 | 0.29 | −2.93 | 0.040 | |||||
| PERMANOVA | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| PERMANOVA Factors | Df | SS | Pseudo-F | p Value | |||||
| Site | 5 | 67,224 | 4.27 | 0.001 | |||||
| Residuals | 85 | 2.68 | |||||||
| Total | 90 | 3.34 | |||||||
| PERMDISP | |||||||||
| Df | SS | Pseudo-F | pvalue | ||||||
| Sites | 5 | 0.07 | 1.45 | 0.40 | |||||
| Residuals | 85 | 0.80 | |||||||
| PERMANOVA Pairwise Test | |||||||||
| Pairs | t-value | p-value | Unique Permutations | ||||||
| Cwebe vs. Dwesa | 2.16 | 0.001 | 997 | ||||||
| Cwebe vs. Kob-Inn | 2.47 | 0.001 | 998 | ||||||
| Cwebe vs. Mazeppa | 216 | 0.001 | 998 | ||||||
| Cwebe vs. Nqabara | 2.28 | 0.001 | 998 | ||||||
| Cwebe vs. Xhorha | 2.01 | 0.001 | 999 | ||||||
| Dwesa vs. Kob-Inn | 2.59 | 0.001 | 999 | ||||||
| Dwesa vs. Mazeppa | 2.18 | 0.001 | 999 | ||||||
| Dwesa vs. Nqabara | 1.64 | 0.001 | 997 | ||||||
| Dwesa vs. Xhorha | 1.62 | 0.003 | 997 | ||||||
| Kob-Inn vs. Mazeppa | 1.49 | 0.006 | 998 | ||||||
| Kob-Inn vs. Nqabara | 2.25 | 0.001 | 998 | ||||||
| Kob-Inn vs. Xhorha | 2.52 | 0.001 | 998 | ||||||
| Mazeppa vs. Nqabara | 1.78 | 0.001 | 997 | ||||||
| Mazeppa vs. Xhorha | 2.00 | 0.001 | 998 | ||||||
| Nqabara vs. Xhorha | 1.64 | 0.002 | 999 | ||||||
| Species | Average | Standard Deviation | Ava | Avb | Ratio | Cumulative Contribution (%) |
|---|---|---|---|---|---|---|
| Zeuxoides helleri | 0.13789 | 0.13450 | 7.31700 | 5.53600 | 1.02520 | 15 |
| Amphipolis squamata | 0.06122 | 0.11534 | 2.12700 | 4.28600 | 0.53080 | 21.6 |
| Phascolosoma sp | 0.03676 | 0.06994 | 1.36500 | 0.39300 | 0.52560 | 25.6 |
| Gunnarea gaimardi | 0.03630 | 0.09143 | 1.65100 | 0.25000 | 0.39700 | 29.6 |
| Amphibalanus venustus | 0.03424 | 0.06127 | 1.58700 | 0.42900 | 0.55890 | 33.3 |
| Lepidonotus semitectus | 0.03207 | 0.04542 | 1.33300 | 0.07100 | 0.70610 | 36.8 |
| Limaria tuberculata | 0.03102 | 0.05949 | 2.65100 | 0.60700 | 0.52150 | 40.2 |
| Cirolana venusticauda | 0.02855 | 0.04533 | 1.38100 | 1.96400 | 0.62980 | 43.3 |
| Lepidonotus durbanensis | 0.02691 | 0.05010 | 0.14300 | 0.96400 | 0.53720 | 46.2 |
| Limnoria quadripunctata | 0.02221 | 0.06438 | 3.42900 | 0.46400 | 0.34500 | 48.6 |
| Arabella iricolor | 0.02197 | 0.03398 | 0.55600 | 0.82100 | 0.64650 | 51.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nkohla, N.; Dlaza, T.S. Macrofaunal Communities Associated with Ecklonia radiata Holdfast Along the Southeast Coast of South Africa. Diversity 2025, 17, 203. https://doi.org/10.3390/d17030203
Nkohla N, Dlaza TS. Macrofaunal Communities Associated with Ecklonia radiata Holdfast Along the Southeast Coast of South Africa. Diversity. 2025; 17(3):203. https://doi.org/10.3390/d17030203
Chicago/Turabian StyleNkohla, Naledi, and Thembinkosi Steven Dlaza. 2025. "Macrofaunal Communities Associated with Ecklonia radiata Holdfast Along the Southeast Coast of South Africa" Diversity 17, no. 3: 203. https://doi.org/10.3390/d17030203
APA StyleNkohla, N., & Dlaza, T. S. (2025). Macrofaunal Communities Associated with Ecklonia radiata Holdfast Along the Southeast Coast of South Africa. Diversity, 17(3), 203. https://doi.org/10.3390/d17030203