



Forest and Population Characteristics of Vulnerable Relict Tsuga forrestii Downie in China


Abstract
1. Introduction
2. Materials and Methods
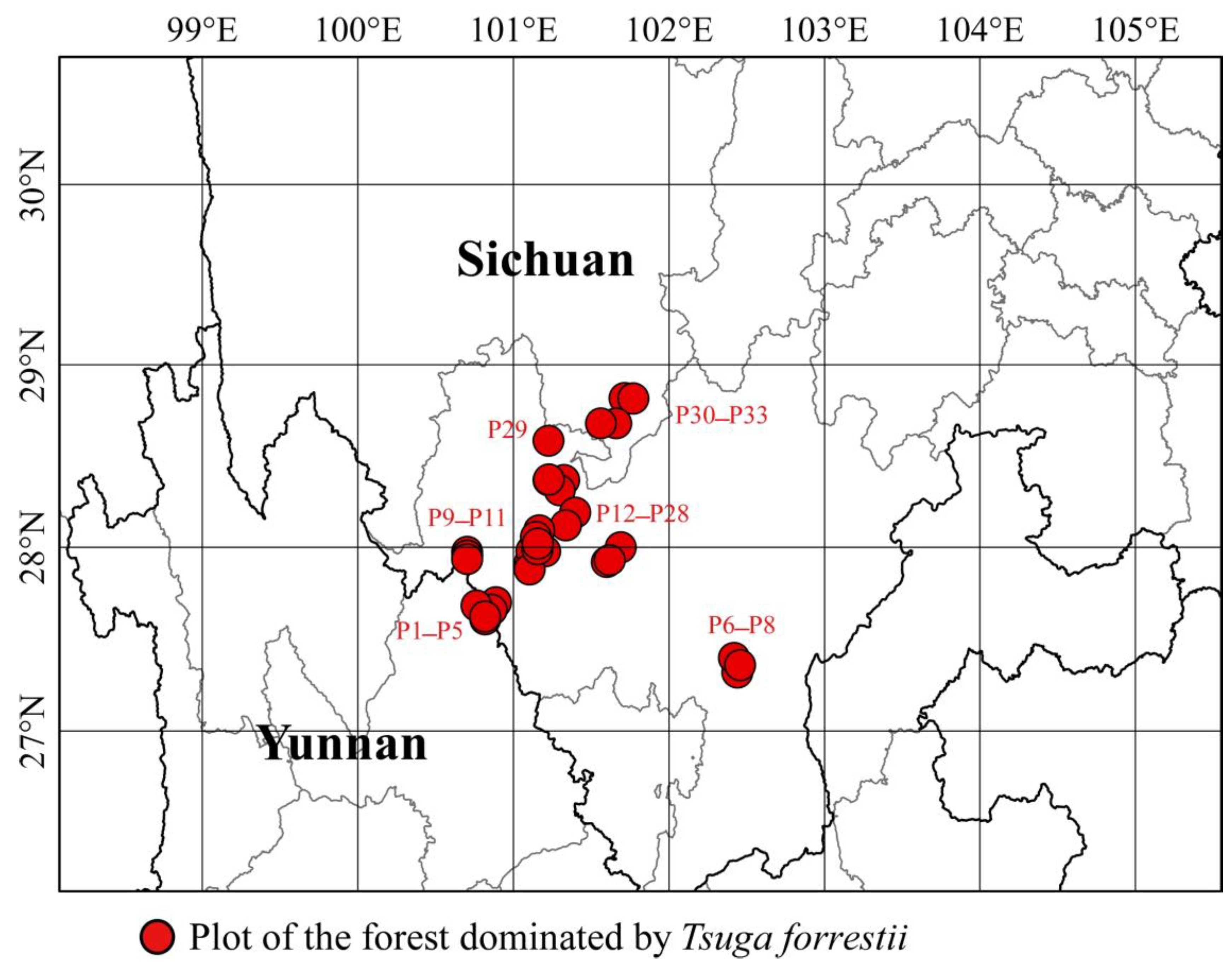
2.1. Study Area
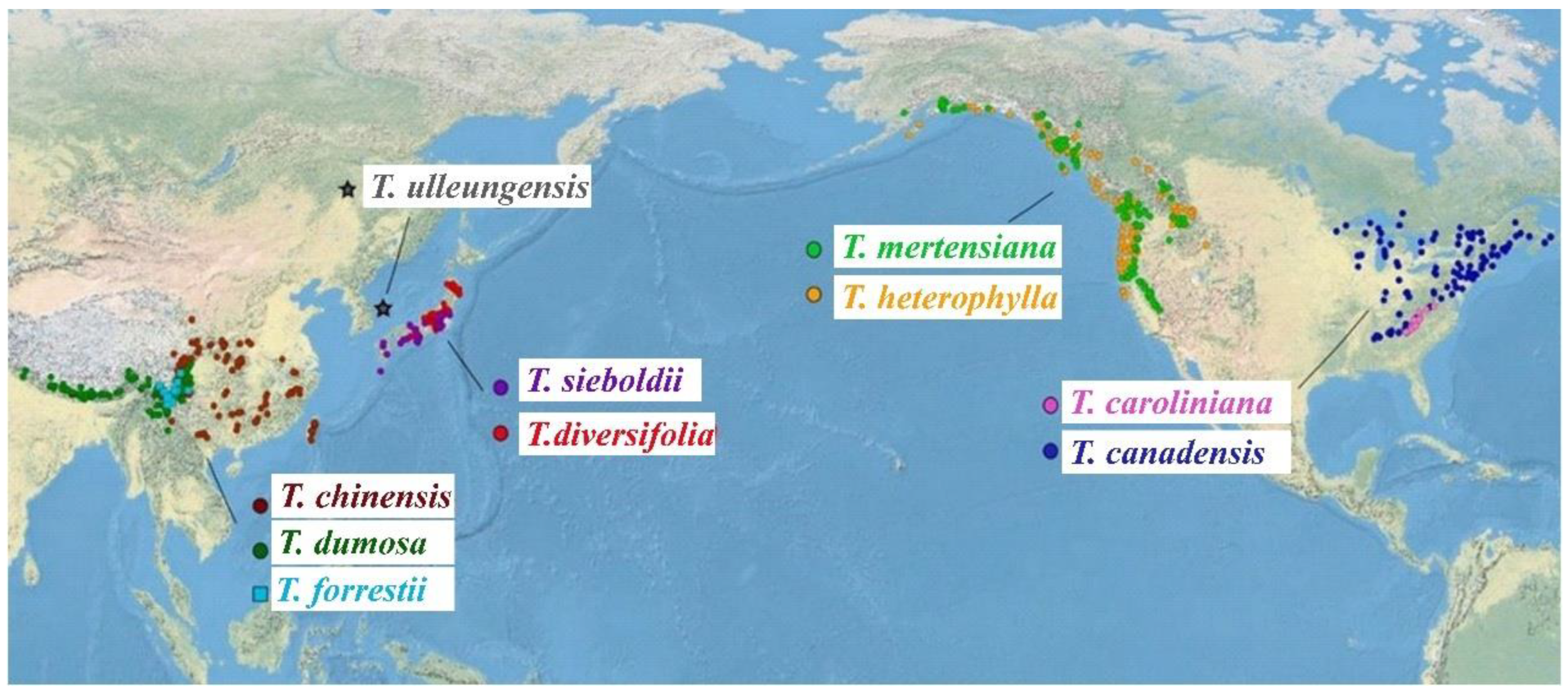
2.2. Study Species
2.3. Data Collection and Analysis
2.3.1. Forest Structure and Species Diversity
2.3.2. Tree Core Analysis of Tsuga forrestii
2.3.3. Phylogenetic Analysis and Phylogenetic Relatedness
3. Results
3.1. Forest Types and Structure
3.2. Forest Floristic Features and Species Diversity
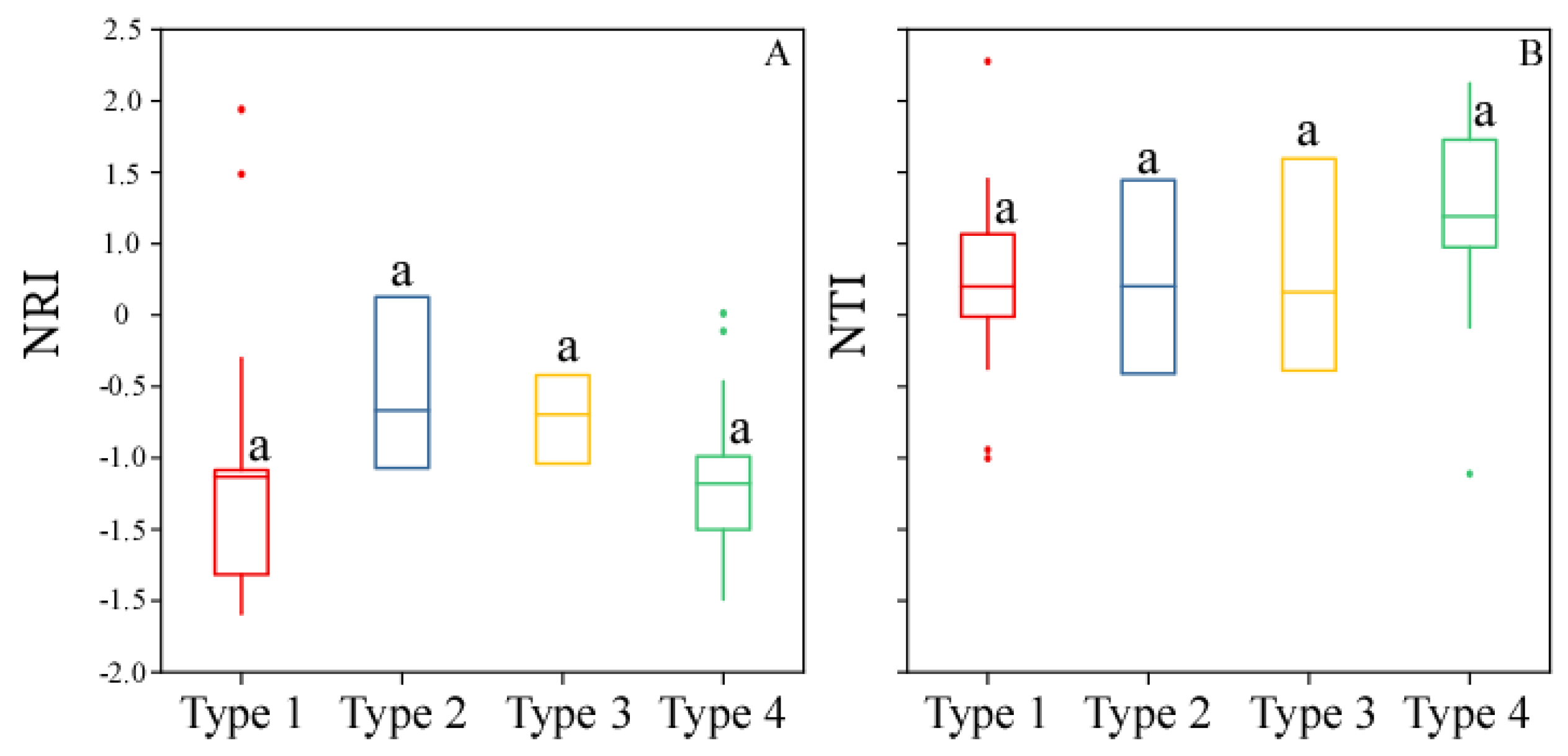
3.3. Phylogenetic Relatedness Among the Forest Types
3.4. Population Characteristics
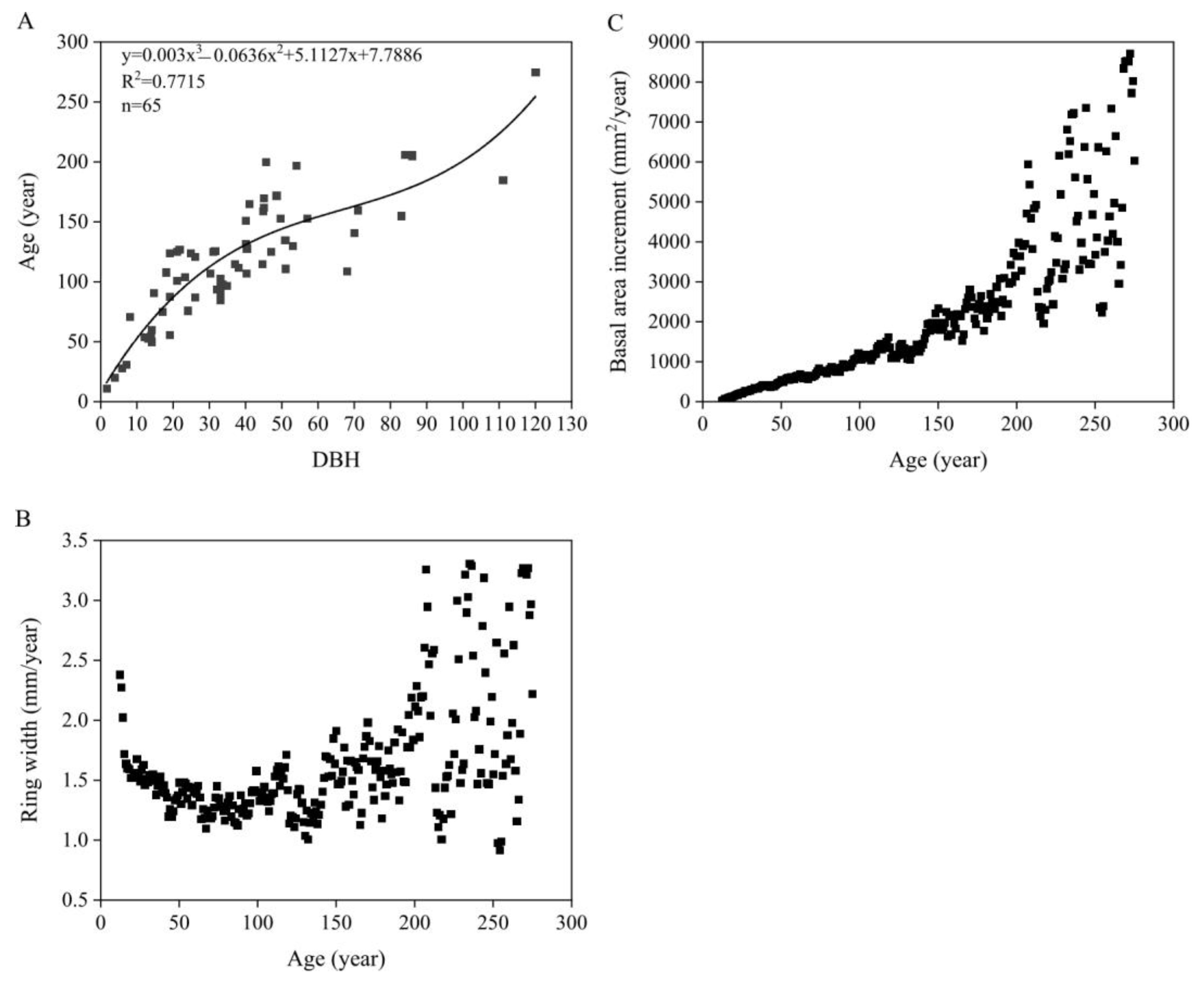
3.4.1. Growth Trends
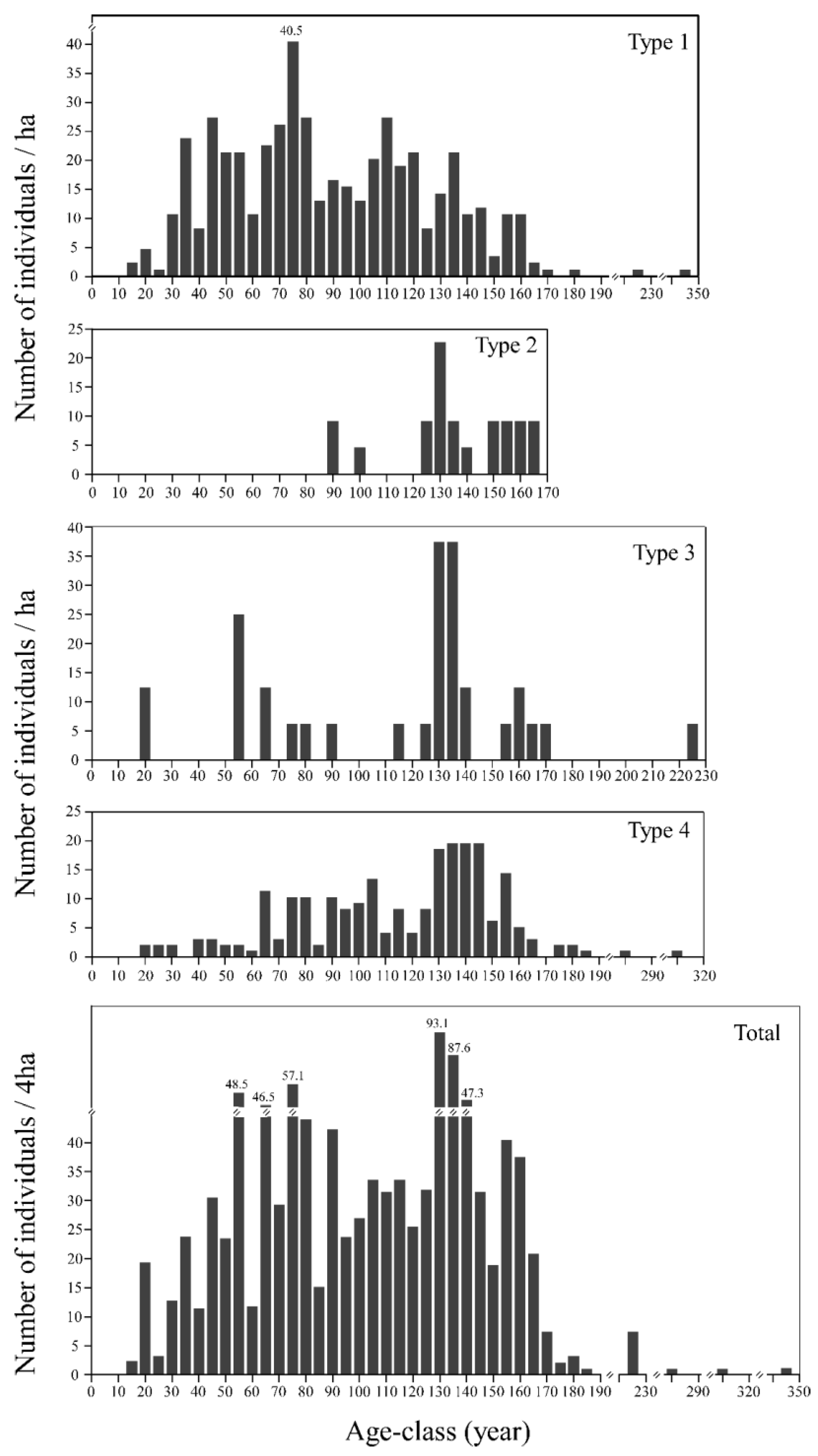
3.4.2. Age Structure and Regeneration
4. Discussion
4.1. Distribution, Forest Characteristics, and Habitats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Distribution Region | Major Habitats and Elevations | DBH (cm) | Height (m) | Age (year) | Major Associated Species | IUCN Threat Categories | Source |
|---|---|---|---|---|---|---|---|---|
| Tsuga forrestii | NW Yunnan and SW Sixhuan | Crest ridge, mountain slopes, steep inclines, and riversides at ca. 2330–3340 m a.s.l | 0.5–148 | 1.3–41 | 12–344 | Abies forrestii, Picea likiangensis, A. squamata, Lithocarpus variolosus, Castanopsis orthacantha, Quercus guyavifolia, Q. rehderiana, Polyspora speciosa, Sorbus meliosmifolia, Betula utilis, and Rhododendron rubiginosum | VU | This study |
| Tsuga dumosa | Sichuan, S Xizang, N and W Yunnan, Bhutan, N India, N Myanmar, Nepal, Sikkim, and N Vietnam | Mountain slopes and river basins at ca. 2300–3500 m a.s.l | 150–250 (–270) | 40–50 | (max age) 1007 | Abies, Picea, Larix, and Pinus | LC | [7,8,42] |
| Tsuga chinensis | Anhui, Fujian, S Gansu, Guangdong, Guangxi, N Guizhou, W Henan, W Hubei, Hunan, Jiangxi, S Shaanxi, Sichuan, Taiwan, Yunnan, and Zhejiang | Mountains, valleys, and river basins at ca. 550–(–1000)–2500(–3500) m a.s.l | 50–60 (–160) | 20–25 (–50) | 140 | Taxus wallichiana var. mairei, Quercus phillyraeoides, Photinia bodinieri, Engelhardia roxburghiana, Pinus armandi, Toxicodendron vernicifluum, Acer davidii, Rhododendron ovatum, Schima argentea, Camellia cuspidate, R. latoucheae, Q. multinervis, Betula luminifera, Prunus dielsiana, Schima superba, R. simiarum, and Castanopsis eyrei | LC | [7,8,43,44,45,46,48] |
| Tsuga sieboldii | Central Honshu, Shikoku, and Kyushu, Japan | Mountains (steep ridges) and hills at ca. 400–1500 m a.s.l | (150)–250 | 25–30 | 500–800 | Abies firma, Pseudotsuga japonica, Chamaecyparis obtusa, Cryptomeria japonica, Pinus densiflora, P. parviflora, and Sciadopitys verticillata | NT | [1,7,47] |
| Tsuga diversifolia | Central Honshu, particularly in Nagano, Tochigi, Gunma, and Yamanashi prefectures, Japan | Mountains, 700–2000 m | 50–60 | 20–25 | Picea jezoensis, Abies homolepis, A. veitchii, A. mariesii, Larix kaempferi, Pinus parviflora, Thuja standishii, Thujopis dola brata var. hondae, Betula ermanii, B. corylifolia, Sorbus japonica, Alnus hirsuta var. sibirica, Quercus mongolica var. grosseserrata, and Rhododendron spp. | LC | [1,7] | |
| Tsuga ulleungensis | Ulleungdo in South Korea | Mountain slopes, 310–500 m | Acer, Camelia, and Fagus | CR | [4,12] | |||
| Tsuga mertensiana | Alaska and Canada | Coast and mountains, 1200–3350 m | 100–150 | 30–40 (–45) | Abies lasiocarpa, A. amabilis, Picea glauca, P. sitchensis, P. engelmannii, Pinus spp., Tsuga heterophylla, Xanthocyparis nootkatensis, Juniperus occidentalis, and Betula papyrifera | LC | [1,7] | |
| Tsuga heteropylla | NW of North America | Coast and mountains, 600–1800 | 150–250 | 60–70 | LC | [1,7] | ||
| Tsuga caroliniana | States of Georgia, North Carolina, South Carolina, Tennessee, and Virginia | Mountains and valleys, 600–1500 m | 50–60 | 20–25 | Broad-leaved trees and shrubs | NT | [1,7] | |
| Tsuga canadensis | Eastern half of North America | Coast and mountains, 600–1500 | 150–200 | 30–40 (–48) | Pinus strobus, Picea rubens, Abies balsamea, and Betula or Populus | NT | [1,7] |
4.2. Growth Trends
4.3. Regeneration Challenges and Conservation Recommendations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Farjon, A.; Filer, D. An Atlas of the World’s Conifers: An Analysis of Their Distribution, Biogeography, Diversity and Conservation Status; Brill: Leiden, The Netherlands, 2013. [Google Scholar]
- Wu, Z.-Y.; Raven, P.H.; Hong, D.-Y. (Eds.) Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1998. [Google Scholar]
- Lepage, B.A. A new species of Tsuga (Pinaceae) from the middle Eocene of Axel Heiberg Island, Canada, and an assessment of the evolution and biogeographical history of the genus. Bot. J. Linn. Soc. 2003, 141, 257–296. [Google Scholar] [CrossRef]
- Holman, G.; Del Tredici, P.; Havill, N.; Lee, N.S.; Cronn, R.; Cushman, K.; Campbell, C.S. A new species and introgression in eastern Asian hemlocks (Pinaceae: Tsuga). Syst. Bot. 2017, 42, 733–746. [Google Scholar] [CrossRef]
- Feng, Y.Y.; Shen, T.T.; Shao, C.C.; Du, H.; Ran, J.H.; Wang, X.Q. Phylotranscriptomics reveals the complex evolutionary and biogeographic history of the genus Tsuga with an East Asian-North American disjunct distribution. Mol. Phylogenet. Evol. 2021, 157, 107066. [Google Scholar] [CrossRef]
- Chen, Z.; Shang, X.; Xie, Z.; Zhang, X. Phylogenetic relationship and evolutionary patterns of Tsuga pollen morphology: A cluster analysis-based study. J. Nanjing For. Univ. 2024, 48, 37. [Google Scholar]
- Farjon, A. A Handbook of the World’s Conifers: Revised and Updated Edition; Brill: Leiden, The Netherlands, 2010; pp. 1007–1019. [Google Scholar]
- Fu, L.; Li, N.; Mill, R.R. Tsuga. Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1999; Volume 4, pp. 39–41. [Google Scholar]
- Ying, T.S.; Chew, M.L.; Chang, H.C. Atlas of the Gymnosperms of China; China Science and Technology Press: Beijing, China, 2003. [Google Scholar]
- Havill, N.P.; Campbell, C.S.; Vining, T.F.; LePage, B.; Bayer, R.J.; Donoghue, M.J. Phylogeny and biogeography of Tsuga (Pinaceae) inferred from nuclear ribosomal ITS and chloroplast DNA sequence data. Syst. Bot. 2008, 33, 478–489. [Google Scholar] [CrossRef]
- Chen, L.; Liu, X.; Wang, Z.; Wu, X.; Hong, K.; Xie, C. Comparative Chloroplast Genome Analyses of Six Hemlock Trees in East Asia: Insights into Their Genomic Characterization and Phylogenetic Relationship. Forests 2023, 14, 2136. [Google Scholar] [CrossRef]
- IUCN (The IUCN Red List of Threatened Species), 2010. Available online: https://www.iucnredlist.org/ (accessed on 19 November 2024).
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.-F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.; Yang, Y.; Ohsawa, M.; Luu, H.T.; et al. Identifying long-term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q.; Ohsawa, M. Tertiary relic deciduous forests on a humid subtropical mountain, Mt. Emei, Sichuan, China. Folia Geobot. 2002, 37, 93–106. [Google Scholar] [CrossRef]
- Tang, C.Q.; Yang, Y.; Ohsawa, M.; Momohara, A.; Hara, M.; Cheng, S.; Fan, S. Population structure of relict Metasequoia glyptostroboides and its habitat fragmentation and degradation in south-central China. Biol. Conserv. 2011, 144, 279–289. [Google Scholar] [CrossRef]
- Tang, C.Q.; Yang, Y.; Ohsawa, M.; Yi, S.R.; Momohara, A.; Su, W.H.; Wu, Z.L.; Zhang, Z.Y.; Peng, M.C.; Wu, Z.L. Evidence for the persistence of wild Ginkgo biloba (Ginkgoaceae) populations in the Dalou Mountains, southwestern China. Am. J. Bot. 2012, 99, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q. The Subtropical Vegetation of Southwestern China: Plant Distribution, Diversity and Ecology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 11. [Google Scholar]
- He, L.Y.; Tang, C.Q.; Wu, Z.L.; Wang, H.C.; Ohsawa, M.; Yan, K. Forest structure and regeneration of the Tertiary relict Taiwania cryptomerioides in the Gaoligong Mountains, Yunnan, southwestern China. Phytocoenologia 2015, 45, 135–156. [Google Scholar] [CrossRef]
- Qian, S.; Yang, Y.; Tang, C.Q.; Momohara, A.; Yi, S.; Ohsawa, M. Effective conservation measures are needed for wild Cathaya argyrophylla populations in China: Insights from the population structure and regeneration characteristics. For. Ecol. Manag. 2016, 361, 358–367. [Google Scholar] [CrossRef]
- Tang, C.Q.; He, L.Y. Population status of the Tertiary relict Tetracentron sinense in the subtropical Ailao Mountains, Yunnan, SW China, and proposed conservation efforts. Annu. Rep. Pro Nat. Fund 2013, 21, 141–150. [Google Scholar]
- Tang, C.Q.; Yang, Y.; Momohara, A.; Wang, H.-C.; Luu, H.T.; Li, S.; Song, K.; Qian, S.; LePage, B.; Dong, Y.-F.; et al. Forest characteristics and population structure of Glyptostrobus pensilis, a globally endangered relict species of southeastern China. Plant Divers. 2019, 41, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Ai, X.; Yao, L.; Huang, J.; Xu, Y.; Lu, X.; Zang, R. Do climate and human disturbance determine the sizes of endangered Metasequoia glyptostroboides trees in their native range? Glob. Ecol. Conserv. 2020, 21, e00850. [Google Scholar] [CrossRef]
- Milne, R.I.; Abbott, R.J. The origin and evolution of Tertiary relict floras. Adv. Bot. Res. 2002, 38, 281–314. [Google Scholar]
- Tang, C.Q.; Yao, S.-Q.; Han, P.-B.; Wen, J.-R.; Li, S.; Peng, M.-C.; Wang, C.-Y.; Matsui, T.; Li, Y.-P.; Lu, S.; et al. Forest characteristics, population structure and growth trends of threatened relict Pseudotsuga forrestii in China. Plant Divers. 2023, 45, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Nualart, N.; Herrando-Moraira, S.; Dong, Y.-F.; Grote, P.J.; Van Ngoc, N.; Van Sam, H.; Li, S.; et al. Identifying long-term stable refugia for dominant Castanopsis species of evergreen broad-leaved forests in East Asia: A tool for ensuring their conservation. Biol. Conserv. 2022, 273, 109663. [Google Scholar] [CrossRef]
- Tang, C.Q.; Lu, X.; Du, M.-R.; Xiao, S.-L.; Li, S.; Han, P.-B.; Zeng, J.-L.; Wen, J.-R.; Yao, S.-Q.; Shi, Y.-C.; et al. Forest characteristics and population structure of a threatened palm tree Caryota obtusa in the karst forest ecosystem of Yunnan, China. J. Plant Ecol. 2022, 15, 829–843. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD: Multivariate Analysis of Ecological Data, Version 698 7.0 for Windows; Wild Blueberry Media: Corvallis, OR, USA, 2016. [Google Scholar]
- Lande, R. Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos 1996, 76, 5–13. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley: New York, NY, USA, 1969. [Google Scholar]
- Wu, Z.Y. The areal-types of Chinese genera of seed plants. Plant Divers. 1991, 13, 1–139. (In Chinese) [Google Scholar]
- Wu, Z.Y. The areal-types of the world families of seed plants. Revised version. Acta Bot. Yunnanica 2003, 5, 535–538. (In Chinese) [Google Scholar]
- Rubino, D.L.; McCarthy, B.C. Dendroclimatological analysis of white oak (Quercus alba L., Fagaceae) from an old-growth forest of southeastern Ohio, USA. J. Torrey Bot. Soc. 2000, 127, 240–250. [Google Scholar] [CrossRef]
- Miller, E.T.; Farine, D.R.; Trisos, C.H. Phylogenetic community structure metrics and null models: A review with new methods and software. Ecography 2017, 40, 461–477. [Google Scholar] [CrossRef]
- Webb, C.O. Exploring the phylogenetic structure of ecological communities: An example for rain forest trees. Am. Nat. 2000, 156, 145–155. [Google Scholar] [CrossRef]
- Helmus, M.R.; Bland, T.J.; Williams, C.K.; Ives, A.R. Phylogenetic measures of biodiversity. Am. Nat. 2007, 169, E68–E83. [Google Scholar] [CrossRef]
- Du, W.B. Patterns of Plant Diversity and Formation Mechanism in the Kunlun Mountains; Lanzhou University: Lanzhou, China, 2021. [Google Scholar]
- Jin, Y.; Qian, H.V. PhyloMaker: An R package that can generate very large phylogenies for vascular plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. Available online: https://www.R-project.org/ (accessed on 23 September 2024).
- Smith, S.A.; Brown, J.W. Constructing a broadly inclusive seed plant phylogeny. Am. J. Bot. 2018, 105, 302–314. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Swenson, N.G. Phylogenetic resolution and quantifying the phylogenetic diversity and dispersion of communities. PLoS ONE 2009, 4, e4390. [Google Scholar] [CrossRef] [PubMed]
- Su, W.P.; Yang, Y.M.; Du, F. A Study on Seed Plant Floristic Characteristics of Tsuga dumosa Community in Lancangjiang Nature Reserve. J. West China For. Sci. 2007, 2, 80–85. (In Chinese) [Google Scholar]
- Wang, D.X. Growth law of Tsuga chinensis in Huoditang Forest Region. J. Northwest A F Univ. (Nat. Sci. Ed.) 2008, 4, 83–88. (In Chinese) [Google Scholar]
- Feng, X.L.; Hu, G.; Liu, Z.H. Research on the Community Characteristics and Population Dynamics of Tsuga chinesis var. tchekiangensis on Gaopo in Guiyang City. Guizhou For. Sci. Technol. 2011, 39, 26–29. (In Chinese) [Google Scholar]
- Li, J.; Liu, P.; Dong, G.; He, X.; Kang, B. Community Structure and Species Composition of Tsuga chinensis Natural Forests in Micangshan Nature Reserve of Shaanxi. Shaanxi For. Sci. Technol. 2014, 3, 35–38. (In Chinese) [Google Scholar]
- Chen, B.; Zhao, J.T.; Guan, Q.W. Community composition and structure of a mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest in Jiangxi Wuyishan, China. Acta Ecol. Sin. 2018, 38, 7359–7372. (In Chinese) [Google Scholar]
- Suzuki, E.; Tsukahara, J. Age structure and regeneration of old growth Cryptomeria japonica forests on Yakushima Island. Bot. Mag. 1987, 100, 223–241. [Google Scholar] [CrossRef]
- Luo, J.W. Growth and biomass of Tsuga chinensis var. tchekiangensis in a natural forest in Guangze, Fujian, China. J. Fujian Coll. For. 2011, 31, 156–160. (In Chinese) [Google Scholar]
- Rossi, S.; DesLauriers, A.; Griçar, J.; Seo, J.-W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Hood, S.; Sala, A. Ponderosa pine resin defenses and growth: Metrics matter. Tree Physiol. 2015, 35, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Puettmann, K.J.; Wilson, S.M.; Baker, S.C.; Donoso, P.J.; Drössler, L.; Amente, G.; Harvey, B.D.; Knoke, T.; Lu, Y.; Nocentini, S.; et al. Silvicultural alternatives to conventional even-aged forest management-what limits global adoption? For. Ecosyst. 2015, 2, 8. [Google Scholar] [CrossRef]
- Gautier-Hion, A.; Duplantier, J.M.; Quris, R.; Feer, F.; Sourd, C.; Decoux, J.P.; Dubost, G.; Emmons, L.; Erard, C.; Hecketsweiler, P.; et al. Fruit characters as a basis of fruit choice and seed dispersal in a tropical forest vertebrate community. Oecologia 1985, 65, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Peres, C.A. Synergistic effects of subsistence hunting and habitat fragmentation on Amazonian forest vertebrates. Conserv. Biol. 2001, 15, 1490–1505. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, P.-B.; Lu, S.-G.; Tang, C.Q. Forest and Population Characteristics of Vulnerable Relict Tsuga forrestii Downie in China. Diversity 2025, 17, 148. https://doi.org/10.3390/d17030148
Han P-B, Lu S-G, Tang CQ. Forest and Population Characteristics of Vulnerable Relict Tsuga forrestii Downie in China. Diversity. 2025; 17(3):148. https://doi.org/10.3390/d17030148
Chicago/Turabian StyleHan, Peng-Bin, Shu-Gang Lu, and Cindy Q. Tang. 2025. "Forest and Population Characteristics of Vulnerable Relict Tsuga forrestii Downie in China" Diversity 17, no. 3: 148. https://doi.org/10.3390/d17030148
APA StyleHan, P.-B., Lu, S.-G., & Tang, C. Q. (2025). Forest and Population Characteristics of Vulnerable Relict Tsuga forrestii Downie in China. Diversity, 17(3), 148. https://doi.org/10.3390/d17030148