
Patterns of Species Richness and Its Endemism of Beetles in the Beijing–Tianjin–Hebei Region of China

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
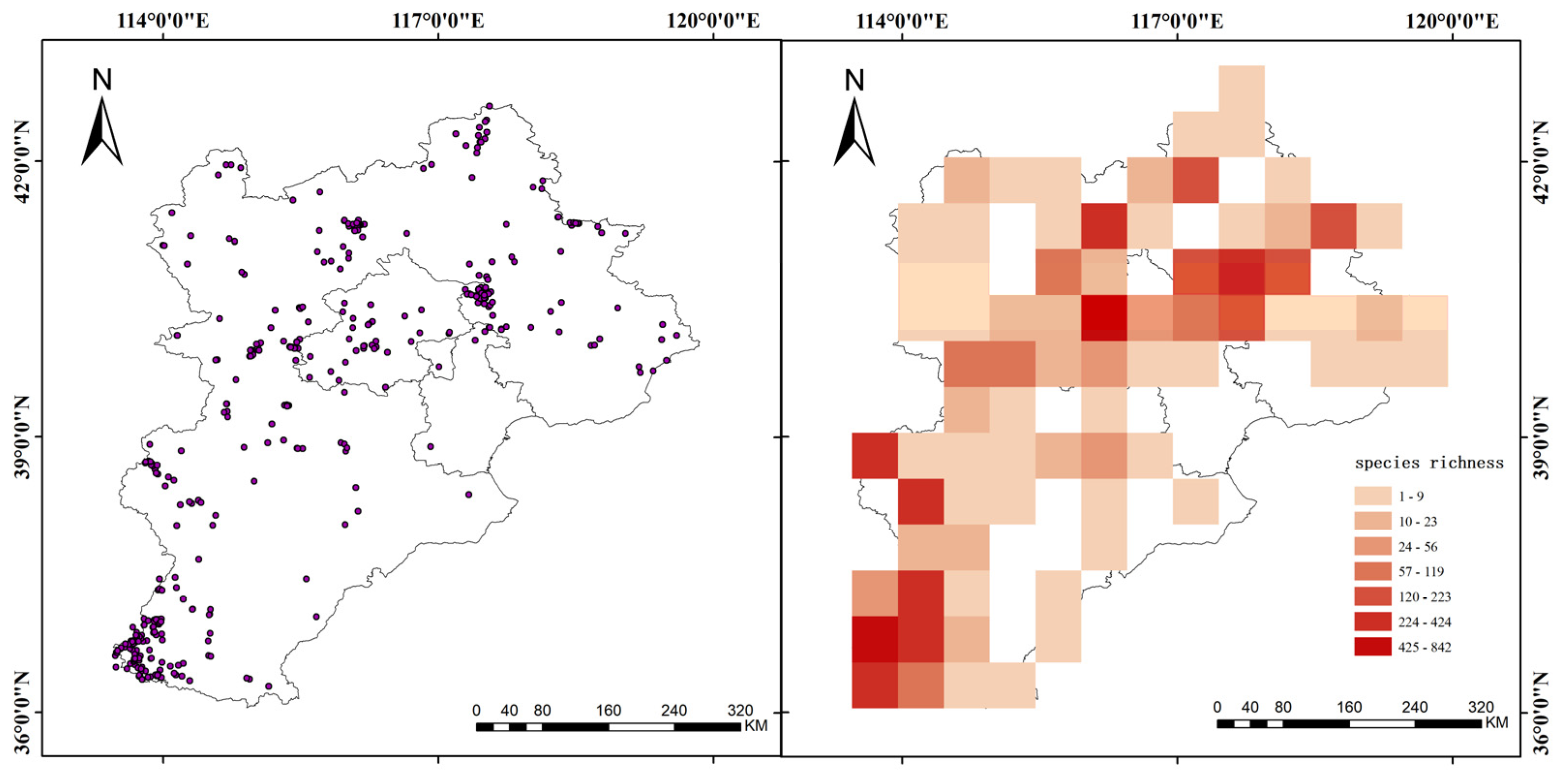
2.3. Mapping Species Richness and Assessing Sampling Bias
2.4. Identification of Areas of Endemism
3. Results
3.1. Assessment of Sampling Bias
3.2. Species Richness Model
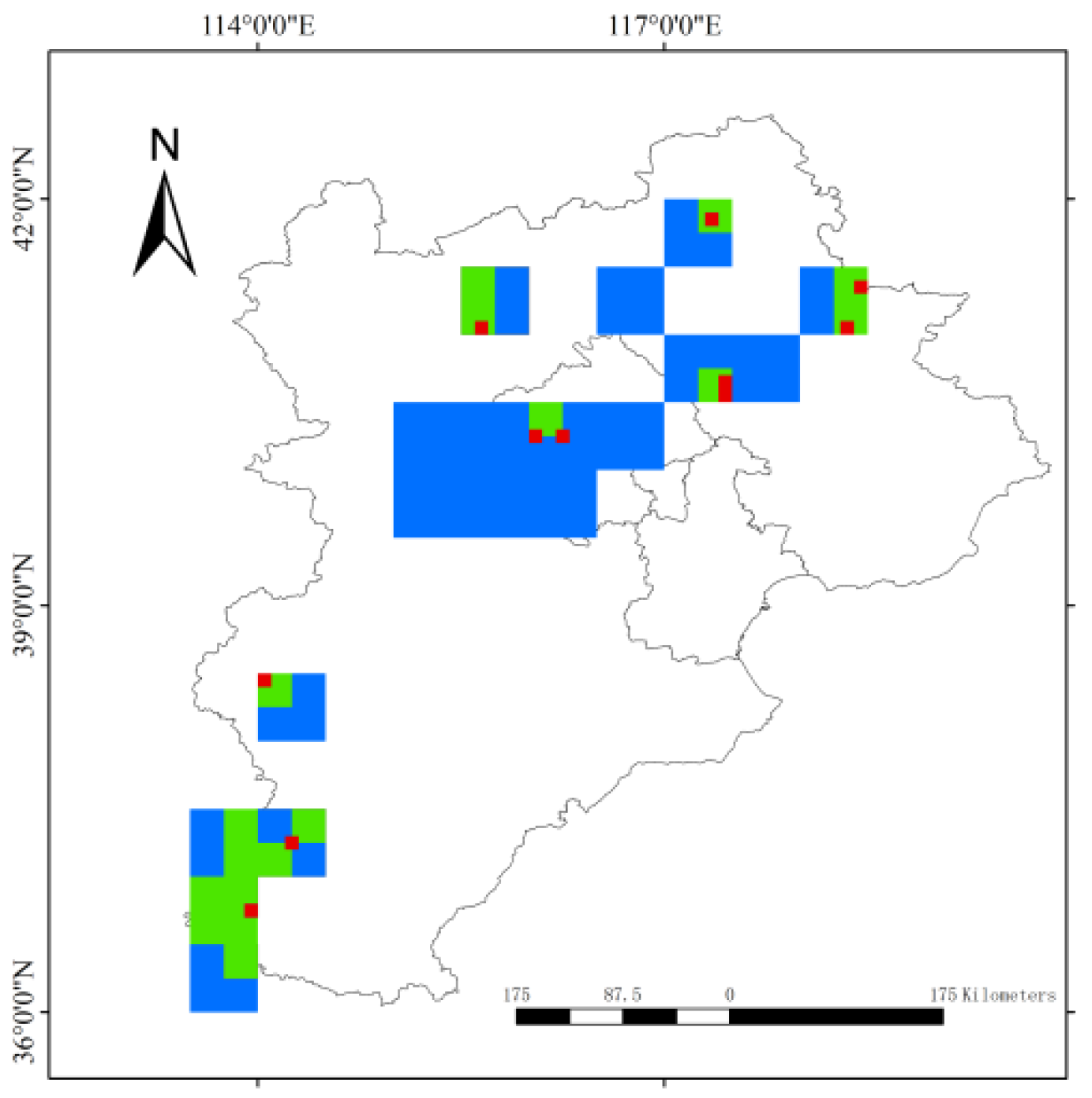
3.3. Parsimony Analysis of Endemicity
3.4. Endemicity Analysis
- (1)
- Southern Taihang Mountains (THM): This region was recognized by all three grid sizes, resulting in 22 consensus regions (Figure S2). Two consensus regions (8 and 10) were identified at the 0.1° grid scale, containing 48 and 17 species, respectively, with scores of 48 and 17. Under the 0.25° grid, consensus regions 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 were identified, with scores ranging from 4.499 to 60. Consensus regions 33, 34, 35, 36, 37, and 38 were identified under the 0.5° grid. The scores are 10.833, 14.5, 12.133, 18.104, 17, 19.271, 14.475, 12.05, 4.499, 13, 16.521, 22, 60, 48. A total of 343 species were contained in these consensus regions.
- (2)
- Xiaowutai Mountains National Nature Reserve (XWTM): This area was recognized by all three grid sizes, resulting in five consensus regions: 10, 12, 28, 30, and 37 (Figures S1–S3), scoring 17, 108, 118, 11.502, and 129.5. These consensus regions contained a total of 165 species.
- (3)
- Yanshan Mountains region (YM): This area was identified at all three grid sizes. Consensus regions 1, 4, 7, 9, 10, 24, 25, 26, 27, 31, 32, 34, and 35 (Figures S1–S3) were identified, with scores of 60, 18, 22, 17.993, 17, 17, 60, 48, 21.5, 23.226, 22.825, 60 and 22. These consensus regions contained a total of 217 species.
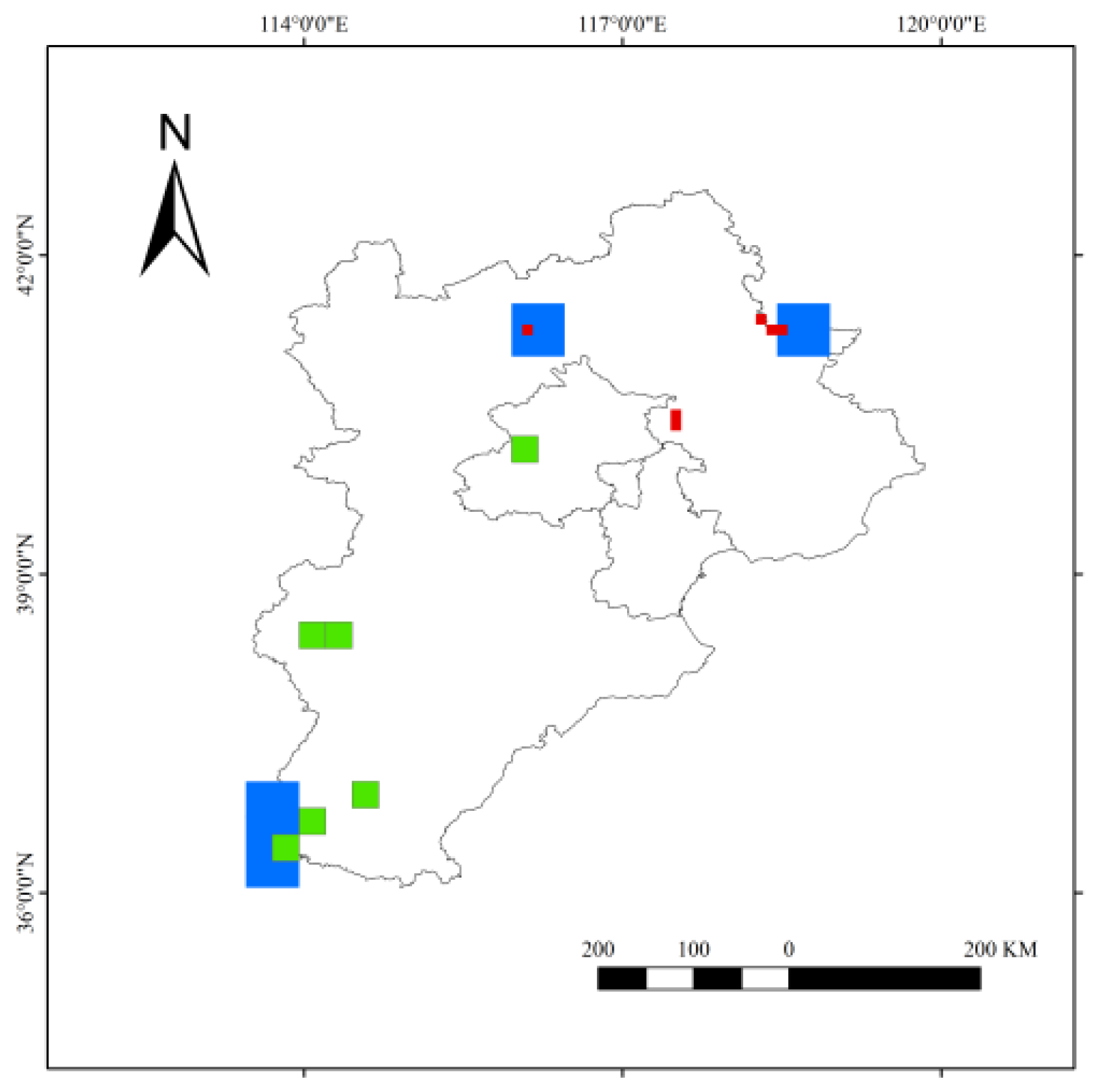
3.5. Jacard Similarity Index Analysis
4. Discussion
4.1. Consistency of Species Richness Patterns with Areas of Endemism
4.2. AOEs in the Beijing–Tianjin–Hebei Region
4.3. Similarities and Differences among the Three Regions
4.4. Data and Methodological Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, H. Distribution Patterns and Assessment of Vascular Plant Diversity in Yanshan Area. Master’s Thesis, Beijing Forestry University, Beijing, China, 2022. [Google Scholar] [CrossRef]
- Zhang, R.Z. Zoogeography of China, Revised Edition); Science Press: Beijing, China, 2011. [Google Scholar]
- Yan, X.Y.; Yang, Y.Y.; Wang, Y.S.; Yan, J.W. Trend and attribution analysis of habitat quality changes in Beijing-Tianjin-Hebei region in the last 40 years. Remote Sens. Technol. Appl. 2023, 38, 251–263. [Google Scholar]
- Zhang, R.Y. Species Diversity and Geographic Distribution Pattern of Beetles in Hebei Province. Master’s Thesis, Hebei University, Baoding, China, 2018. [Google Scholar]
- Nie, R.Z.; Bai, M.; Yang, X.K. Seventy years of beetle research in China. J. Appl. Entomol. 2019, 56, 884–906. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Lamoreux, J.F.; Morrison, J.C.; Ricketts, T.H.; Olson, D.M.; Dinerstein, E.; McKnight, M.W.; Shugart, H.H. Global tests of biodiversity concordance and the importance of endemism. Nature 2006, 440, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Qiao, G.X.; Lei, F.M. Use of parsimony analysis to identify areas of endemism of chinese birds: Implications for conservation and biogeography. Int. J. Mol. Sci. 2010, 11, 2097–2108. [Google Scholar] [CrossRef] [PubMed]
- Gomes-da-Silva, J.; Amorim, A.M.; Forzza, R.C. Distribution of the xeric clade species of pitcairnioideae (Bromeliaceae) in South America: A perspective based on areas of endemism. J. Biogeogr. 2017, 44, 1994–2006. [Google Scholar] [CrossRef]
- JI, F.F.; Liang, Y.; Yan, S.H.; Qin, T.L.; Shen, Z.J. Estimating aquatic plant diversity and distribution in rivers from Jingjinji region, China, using environmental DNA metabarcoding and a traditional survey method. Environ. Res. 2021, 199, 11348. [Google Scholar] [CrossRef]
- Liu, T.; Liu, H.Y.; Wang, Y.N.; Xi, H.C.; Yang, Y.X. Assessing the diversity and distribution pattern of the speciose genus Lycocerus (Coleoptera: Cantharidae) by the global-scale data. Front. Ecol. Evol. 2022, 10, 794750. [Google Scholar] [CrossRef]
- Chen, Q.; Li, D.S.; An, M.T. Study on the pattern and impact of seed plant species richness in Renhuai City. J. Mt. Agric. Biol. 2024, 43, 18–27. [Google Scholar] [CrossRef]
- Sandel, B.; Arge, L.; Dalsgaard, B.; Davies, R.G.; Gaston, K.J.; Sutherland, W.J.; Svenning, J.C. The influence of late quaternary climate change velocity on species endemism. Science 2011, 334, 660–664. [Google Scholar] [CrossRef]
- Li, Y.L.; Wang, Y.J.; Zhang, H.; Li, S.Y.; Chai, Z.C. Exploring the species richness pattern and areas of endemism of Tenebrionidae (Coleoptera) in Xinjiang, China. Diversity 2022, 14, 558. [Google Scholar] [CrossRef]
- Wang, L.Y.; Hu, H.J.; Jiang, J.; Hu, Y.M. Species richness patterns of mammals and birds and their drivers in the Nanling Mountain Range. Biodiversity 2024, 32, 53–63. [Google Scholar] [CrossRef]
- Harold, A.S.; Mooi, R.D. Areas of endemism: Definition and recognition criteria. Syst. Biol. 1994, 43, 261–266. [Google Scholar] [CrossRef]
- Huang, J.H.; Chen, B.; Liu, C.R.; Lai, J.S.; Zhang, J.L.; Ma, K.P. Identifying hotspots of endemic woody seed plant diversity in China. Divers. Distrib. 2012, 18, 673–688. [Google Scholar] [CrossRef]
- Casazza, G.; Zappa, E.; Mariotti, M.G.; Médail, F.; Minuto, L. Ecological and hhistorical factors affecting distribution pattern and richness of endemic plant species: The Case of the Mariti-me and Ligurian Alps Hotspot. Divers. Distrib. 2008, 14, 47–58. [Google Scholar] [CrossRef]
- Huang, X.L.; Qiao, G.X.; Lei, F.M. Diversity and distribution of aphids in the Qinghai–Tibetan Plateau–Himalayas. Ecol. Entomol. 2006, 31, 608–615. [Google Scholar] [CrossRef]
- López-Pujol, J.; Zhang, F.M.; Sun, H.Q.; Ying, T.S.; Ge, S. Centres of plant endemism in China: Places for survival or for speciation. J. Biogeogr. 2011, 38, 1267–1280. [Google Scholar] [CrossRef]
- Yuan, S.; Huang, M.; Wang, X.S.; Ji, L.Q.; Zhang, Y.L. Centers of endemism and diversity patterns for typhlocybine leafhoppers (Hemiptera: Cicadellidae: Typhlocybinae) in China. Insect Sci. 2014, 21, 523–536. [Google Scholar] [CrossRef]
- Gao, C.; Chen, J.; Li, Y.; Jiang, L.Y.; Qiao, G.X. Congruent patterns between species richness and areas of endemism of the Greenideinae aphids (Hemiptera: Aphididae) revealed by global-scale data. Zool. J. Linn. Soc. 2018, 183, 791–807. [Google Scholar] [CrossRef]
- Zhao, Z.X.; Yang, L.; Long, J.K.; Chang, Z.M.; Zhou, Z.X.; Zhi, Y.; Yang, L.J.; Li, H.X.; Sui, Y.J.; Gong, N.; et al. Endemism patterns of planthoppers (Fulgoroidea) in China. Front. Ecol. Evol. 2021, 9, 683722. [Google Scholar] [CrossRef]
- Qian, H.; Mishler, D.B.; Zhang, J.; Qian, S.H. Global patterns and ecological drivers of taxonomic and phylogenetic endemism in angiosperm genera. Plant Divers. 2024, 46, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.X. Patterns of Diversity and Predictions of Suitable Areas for Important Agricultural and Forestry Pests of the Chinese Fulgoroidae (Cicadellidae). Ph.D. Thesis, Guizhou University, Guiyang, China, 2021. [Google Scholar]
- Valeria, B.; Claudio, B.; Adrián, G.; Facundo, S.; Ezequiel, M.; Ignacio, R.; Juan, J.R.; Pablo, T.; Jorge, D.W.; Agustín, M.A.; et al. Biodiversity of vertebrates in Argentina: Patterns of richness, endemism and conservation status. ZooKeys 2022, 1085, 101–127. [Google Scholar] [CrossRef]
- Zhang, Y. Species Diversity and Zonal Distribution of Beetles in Beijing-Tianjin-Hebei Region. Master’s Thesis, Hebei University, Baoding, China, 2014. [Google Scholar]
- Li, H.H.; Hao, S.L.; Hu, B.B.; Liu, G.Q.; Zhao, T.J. Baxian Mountain Forest Insects; Science Press: Beijing, China, 2020; pp. 79–156. [Google Scholar]
- Ren, G.D. Fauna Sinica, Insects, Volume 63, Coleoptera, Tenebrionidae (I); Science Press: Beijing, China, 2016. [Google Scholar]
- Yu, G.Y. Wangjiayuan Insects; Science Publishing House: Beijing, China, 2016; pp. 101–202. [Google Scholar]
- Yu, G.Y. Ecological Atlas of Beijing Forestry Insects; Science Publishing House: Beijing, China, 2018; pp. 298–300. [Google Scholar]
- Guo, X.L. Insect Resource Diversity in the Fountain Green Forest Area of Hebei Province. Master’s Thesis, Hebei University, Baoding, China, 2017. [Google Scholar]
- Tang, S.Y. Insect Diversity and Phylogenetic Composition of Wuling Mountains. Master’s Thesis, Hebei University, Baoding, China, 2017. [Google Scholar]
- Guan, H.H. Insect Species Diversity and Resource Status in Liaoheyuan Nature Reserve. Master’s Thesis, Hebei University, Baoding, China, 2018. [Google Scholar]
- Zhang, J. A Preliminary Examination of Insect Resources and Analysis of Beetle Species Diversity in Wuyuezhai National Forest Park. Master’s Thesis, Hebei University, Baoding, China, 2019. [Google Scholar]
- Iwan, D.; Lobl, I. Catalogue of Palaearctic Coleoptera: Tenebrionoidea; Koninklijke Brill NV Press: Leiden, The Netherlands, 2020; Volume 5, pp. 27–641. [Google Scholar]
- Liu, L. Species Diversity and Phylogenetic Analysis of Insects in Laoya Mountain. Master’s Thesis, Hebei University, Baoding, China, 2021. [Google Scholar]
- Di, J.M.; Ren, G.D. Ecological Atlas of Insects of Hebei; Science Press: Beijing, China, 2021; pp. 138–344. [Google Scholar]
- Zhou, H.Z. Fauna Sinica, Insecta, Volume 75, Coleoptera, Histeroidea; Science Press: Beijing, China, 2022; pp. 60–608. [Google Scholar]
- Zu, G.H.; Liu, F.; Liu, X.J. Ecological Atlas of Tianjin Insects; Nankai University Press: Tianjin, China, 2022; pp. 177–284. [Google Scholar]
- Yan, Y. Insect Species Diversity and Resource Status in Heilongshan National Forest Park, Hebei Province. Master’s Thesis, Hebei University, Baoding, China, 2017. [Google Scholar]
- Colwell, R.K.; Elsensohn, J.E. EstimateS Turns 20: Statistical estimation of species richness and shared species from samples, with non-parametric extrapolation. Ecography 2014, 37, 609–613. [Google Scholar] [CrossRef]
- Oliveira, U.; Brescovit, A.D.; Santos, A.J. Sampling effort and species richness assessment: A case study on Brazilian spiders. Biodivers. Conserv. 2017, 26, 1481–1493. [Google Scholar] [CrossRef]
- Rosen, B.R. From fossils to earth history: Applied historical biogeography. In Analytical Biogeography; Myers, A.A., Giller, P.S., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 437–481. [Google Scholar] [CrossRef]
- Nihei, S.S. Misconceptions about parsimony analysis of endemicity. J. Biogeogr. 2006, 33, 2099–2106. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Zheng, W. Algorithm Improvement and Optimization for Constructing Phylogenetic Trees Based on Maximum Parsimony Principle. Master’s Thesis, Shenzhen Institutes of Advanced Technology, Chinese Academy of Sciences, Shenazhen, China, 2013. [Google Scholar]
- Huang, X.L.; Lei, F.M.; Qiao, G.X. Areas of endemism and patterns of diversity for Aphids of the Qinghai-Tibetan Plateau and the Himalayas. J. Biogeogr. 2008, 35, 230–240. [Google Scholar] [CrossRef]
- Santos, C.M.D.; Fuhlendorf, M. GeX: An automated tool for generating XYD files for anal-ysis of endemicity using VNDM. Cladistics 2019, 35, 125–129. [Google Scholar] [CrossRef]
- Szumik, C.A.; Cuezzo, F.; Goloboff, P.A.; Chalup, A.E. An optimality criterion to determine areas of endemism. Syst. Biol. 2002, 51, 806–816. [Google Scholar] [CrossRef]
- Prado, J.R.; Brennand, P.G.G.; Godoy, L.P.; Libardi, G.S.; Abreu-Júnior, E.F.; Roth, P.R.O.; Chiquito, E.A.; Percequillo, A.R. Species richness and areas of endemism of Oryzomyine rodents (Cricetidae, Sigmodontinae) in South America: An NDM/VNDM approach. J. Biogeogr. 2015, 42, 540–551. [Google Scholar] [CrossRef]
- Du, C.; Chen, J.; Jiang, L.; Qiao, G. High correlation of species diversity patterns between specialist herbivorous insects and their specific hosts. J. Biogeogr. 2020, 47, 1232–1245. [Google Scholar] [CrossRef]
- Jetz, W.; Rahbek, C.; Colwell, R.K. The Coincidence of rarity and richness and the potential signature of history in centres of endemism. Ecol. Lett. 2004, 7, 1180–1191. [Google Scholar] [CrossRef]
- Moline, P.M.; Linder, H.P. Input data, analytical methods and biogeography of elegia (restionaceae). J. Biogeogr. 2006, 33, 47–62. [Google Scholar] [CrossRef]
- Linder, H.P. On Areas of endemism, with an example from the African restionaceae. Syst. Biol. 2001, 50, 892–912. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, H.L.; Yago, M.; Settele, J.; Li, X.S.; Ueshima, R.; Grishin, N.V.; Wang, M. Species richness of eurasian zephyrus Hairstreaks (Lepidoptera: Lycaenidae: Theclini) with implications on historical biogeography: An NDM/VNDM approach. PLoS ONE 2018, 13, e0191049. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Mao, L.F.; Sandel, B.; Swenson, N.G.; Svenning, J.C. High plant endemism in China is partially linked to reduced Glacial-Interglacial climate change. J. Biogeogr. 2016, 43, 145–154. [Google Scholar] [CrossRef]
- Shipley, R.B.; McGuire, L.J. The environmental conditions of endemism hotspots shape the functional traits of mammalian assemblages. Biol. Sci. 2024, 291, 20232773. [Google Scholar] [CrossRef] [PubMed]
- Jing, T.T. Species Diversity Composition and Distribution Patterns of Insects in Yanshan-Taihang Mountains. Master’s Thesis, Hebei University, Baoding, China, 2023. [Google Scholar]
- Zhang, N. Species Diversity and Zonation of Insects in the South Section of Taihang Mountains. Master’s Thesis, Hebei University, Baoding, China, 2022. [Google Scholar]
- Yang, L.K. Composition and Distribution Pattern of Coleoptera in Xiaowutai Mountain. Master’s Thesis, Hebei University, Baoding, China, 2013. [Google Scholar]
- Iveth, S.E.; Adolfo, S.E.; Javier, C.G.; Ana Rosa, L.F. Richness, distribution, and endemism of neotropical subtribe Ponerinae (Orchidaceae, Epidendreae). Braz. J. Bot. 2024, 47, 501–517. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Ren, G.D. Species diversity, zonation composition and vertical distribution of t-he family Anacardiaceae in the major forest parks of Hebei Province. Hebei For. Sci. Technol. 2017, 4, 1–7. [Google Scholar] [CrossRef]
- Shi, G.R. Multivariate data analysis in palaeoecology and palaeobiogeography—A review. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1993, 105, 199–234. [Google Scholar] [CrossRef]
- Hausdorf, B.; Hennig, C. Biotic element analysis in biogeography. Syst. Biol. 2003, 52, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Szumik, C.A.; Goloboff, P.A. Areas of endemism: An improved optimality criterion. Syst. Biol. 2004, 53, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Carine, M.A.; Humphries, C.J.; Guma, I.R.; Reyes-Betancort, J.A.; Guerra, A.S. Areas and algorithms: Evaluating numerical Approaches for the delimitation of areas of endemism in the canary islands archipelago. J. Biogeogr. 2009, 36, 593–611. [Google Scholar] [CrossRef]
- Morrone, J.J. Parsimony analysis of endemicity (PAE) revisited. J. Biogeogr. 2014, 41, 842–854. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Family | Genus | Species | |||
|---|---|---|---|---|---|---|
| Co-Families | Similarity (Cj) | Co-Genera | Similarity (Cj) | Co-Species | Similarity (Cj) | |
| A and B | 56 | 0.87 | 443 | 0.57 | 708 | 0.46 |
| A and C | 57 | 0.87 | 556 | 0.69 | 966 | 0.63 |
| B and C | 55 | 0.90 | 441 | 0.66 | 699 | 0.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Y.; Ren, G. Patterns of Species Richness and Its Endemism of Beetles in the Beijing–Tianjin–Hebei Region of China. Diversity 2024, 16, 496. https://doi.org/10.3390/d16080496
Niu Y, Ren G. Patterns of Species Richness and Its Endemism of Beetles in the Beijing–Tianjin–Hebei Region of China. Diversity. 2024; 16(8):496. https://doi.org/10.3390/d16080496
Chicago/Turabian StyleNiu, Yuxian, and Guodong Ren. 2024. "Patterns of Species Richness and Its Endemism of Beetles in the Beijing–Tianjin–Hebei Region of China" Diversity 16, no. 8: 496. https://doi.org/10.3390/d16080496
APA StyleNiu, Y., & Ren, G. (2024). Patterns of Species Richness and Its Endemism of Beetles in the Beijing–Tianjin–Hebei Region of China. Diversity, 16(8), 496. https://doi.org/10.3390/d16080496