
Trade-Off between Song Complexity and Colorfulness in Parid Birds


Abstract
1. Introduction
2. Materials and Methods
2.1. Song
2.2. Data Source and Selection
2.3. Analysis
2.4. Explanatory Variables
2.4.1. Colorfulness
2.4.2. Color Diversity
2.4.3. Elevational Distribution, Body Length and Mass
2.4.4. Climate Variables
2.5. Statistical Analyses
2.5.1. Bivariate Correlations and Phylogenetic Independent Contrasts
2.5.2. Phylogenetic Generalized Least Squares
3. Results
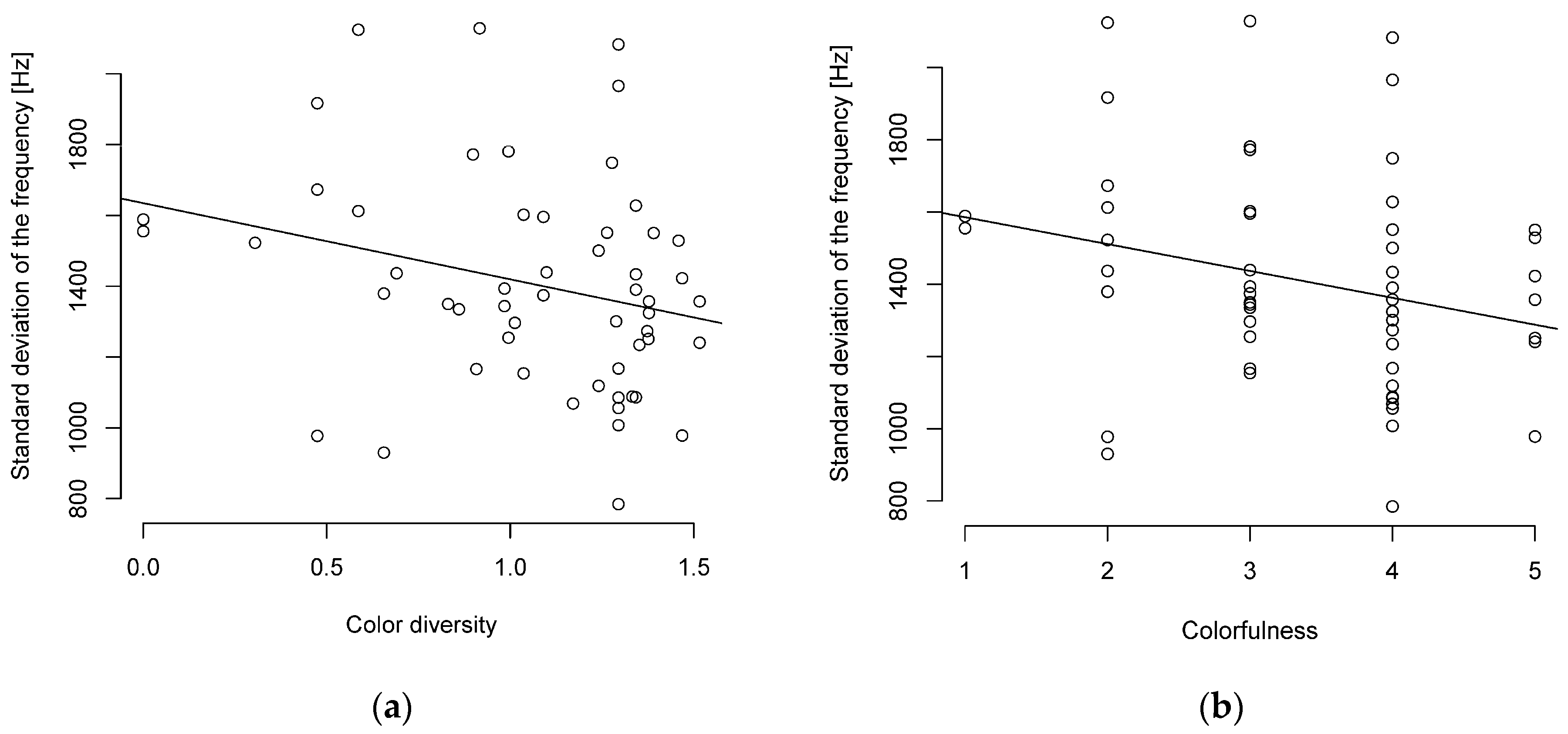
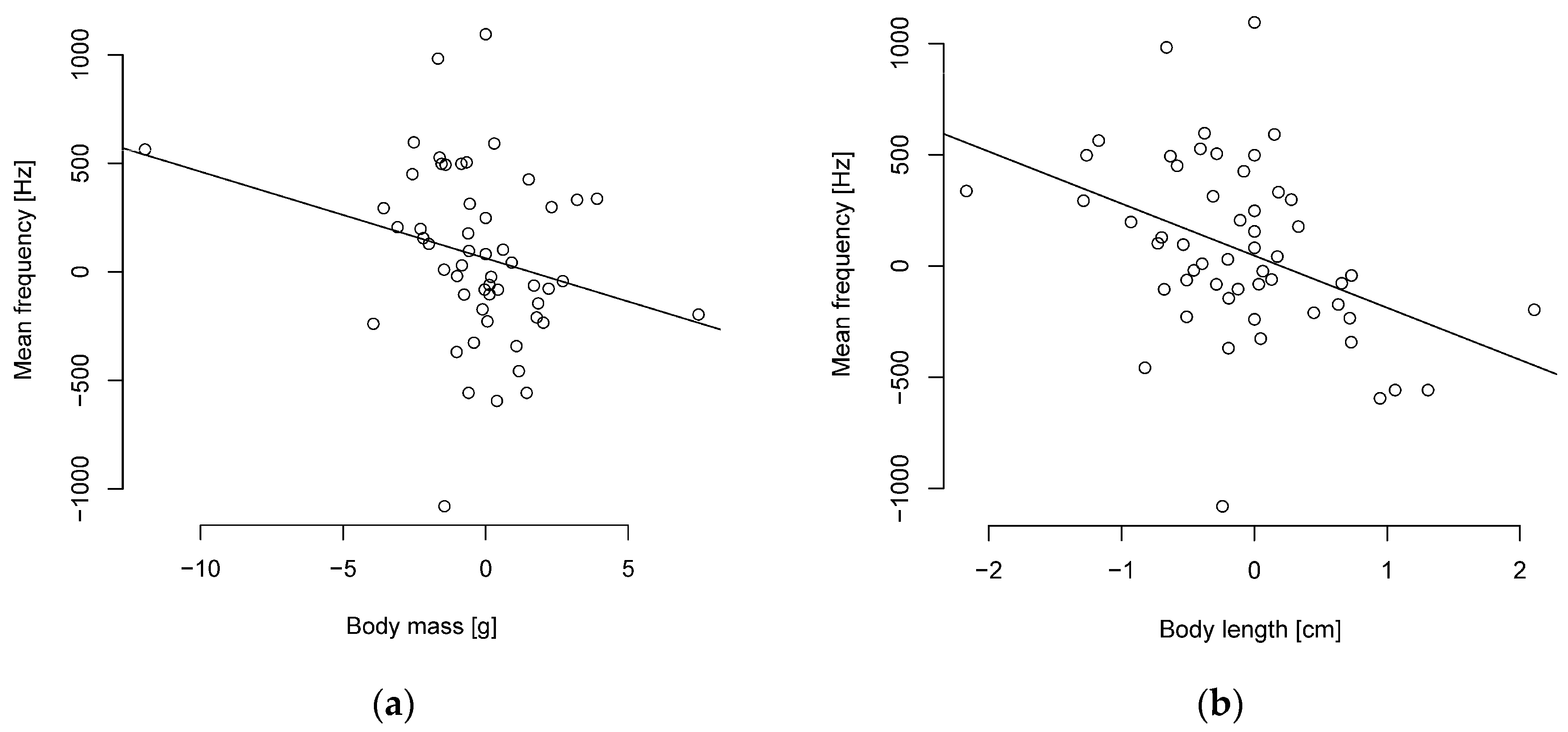
3.1. Relationship between Song Complexity and Colorfulness and Body Size
3.2. Other Correlations Found
3.2.1. Peak Frequency and Body Size
3.2.2. Song Variables and Minimum Temperature
3.2.3. Song Variables and Minimum Precipitation
3.2.4. Song Complexity and Elevation Distribution
3.2.5. Peak Frequency and Elevational Distribution
3.3. Phylogenetic Signal and Models of Evolution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kappeler, P. Verhaltensbiologie: Kapitel 7: Sexuelle Selektion: Evolutionäre Grundlagen; Kapitel 9.5.2 Besserer Nachwuchs Durch Gute Gene; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Lind, H.; Dabelsteen, T.; Gregor, P.M. Female great tits can identify mates by song. Anim. Behav. 1996, 52, 667–671. [Google Scholar] [CrossRef]
- Päckert, M. Song: The Learned Language of Three Major Bird Clades, In Bird Species; Tietze, D.T., Ed.; Springer: Cham, Switzerland, 2018; pp. 75–94. [Google Scholar] [CrossRef]
- Tietze, D.T.; Martens, J.; Fischer, B.S.; Sun, Y.-H.; Klussmann-Kolb, A.; Päckert, M. Evolution of leaf warbler songs (Aves: Phylloscopidae). Ecol. Evol. 2015, 5, 781–798. [Google Scholar] [CrossRef]
- Baker, M.; Bjerke, T.; Espmark, Y.; Lampe, H. Sexual response of female great tits to variation in size of males song repertoires. Am. Nat. 1986, 128, 491–498. [Google Scholar] [CrossRef]
- Garamszegi, L.Z.; Moreno, J.; Møller, A.P. Avian song complexity is associated with high field metabolic rate. Evol. Ecol. Res. 2006, 8, 75–90. [Google Scholar]
- Doutrelant, C.; Blondel, J.; Perret, P.; Lambrechts, M.M. Blue tit song repertoire size, male quality and interspecific competition. J. Avian Biol. 2000, 31, 360–366. [Google Scholar] [CrossRef]
- Nowicki, S.; Hasselquist, D.; Bensch, S.; Peters, S. Nestling growth and song repertoire size in great reed warblers: Evidence for song learning as an indicator mechanism in mate choice. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 2419–2424. [Google Scholar] [CrossRef] [PubMed]
- Bostwick, K. (Ed.) Handbook of Bird Biology: Chapter 4: Feathers and Plumages, 3rd ed.; Wiley: New York, USA, 2016. [Google Scholar]
- Hill, G.E. Plumage coloration is a sexually selected indicator of male quality. Nature 1991, 350, 337–339. [Google Scholar] [CrossRef]
- Senar, J.C.; Figuerola, J.; Domènech, J. Plumage coloration and nutritional condition in the great tit Parus major: The roles of carotenoids and melanins differ. Die Naturwissenschaften 2003, 90, 234–237. [Google Scholar] [CrossRef]
- Slagsvold, T.; Lifjeld, J.T. Variation in plumage colour of the great tit Parus major in relation to habitat, season and food. J. Zool. 1985, 206, 321–328. [Google Scholar] [CrossRef]
- Orteu, A.; Jiggins, C.D. The genomics of coloration provides insights into adaptive evolution. Nat. Rev. Genet. 2020, 21, 461–475. [Google Scholar] [CrossRef]
- Dufva, R.; Allander, K. Intraspecific variation in plumage coloration reflects immune response in great tit (Parus major) males. Funct. Ecol. 1995, 9, 785. [Google Scholar] [CrossRef]
- Völker, O. Die Isolierung eines gelben und eines roten Lipochroms aus Vogelfedern. J. Für Ornithol. 1951, 93, 20–26. [Google Scholar] [CrossRef]
- Weatherhead, P.J.; Robertson, R.J. Offspring quality and the polygyny threshold: ‘The sexy son hypothesis’. Am. Nat. 1979, 113, 201–208. [Google Scholar] [CrossRef]
- Petrie, M. Improved growth and survival of offspring of peacocks with more elaborate trains. Nature 1994, 371, 598–599. [Google Scholar] [CrossRef]
- Lovette, I.J. (Ed.) Handbook of Bird Biology: Chapter 2: Avian Diversity and Classification, Chapter 3: How Birds Evolve, 3rd ed.; Wiley: Chichester, UK, 2016. [Google Scholar]
- Badyaev, A.V.; Hill, G.E.; Weckworth, B.V. Species divergence in sexually selected traits: Increase in song elaboration is related to decrease in plumage ornamentation in finches. Evolution 2002, 56, 412–419. [Google Scholar] [CrossRef]
- Zahavi, A. Mate selection: A selection for a handicap. J. Theor. Biol. 1975, 53, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Shutler, D. Sexual selection: When to expect trade-offs. Biol. Lett. 2011, 7, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Mason, N.A.; Shultz, A.J.; Burns, K.J. Elaborate visual and acoustic signals evolve independently in a large, phenotypically diverse radiation of songbirds. Proc. Biol. Sci. 2014, 281, 20140967. [Google Scholar] [CrossRef] [PubMed]
- Ornelas, J.F.; González, C.; de los Monteros, A.E. Uncorrelated evolution between vocal and plumage coloration traits in the trogons: A comparative study. J. Evol. Biol. 2009, 22, 471–484. [Google Scholar] [CrossRef]
- Hailman, J. The organization of major vocalizations in the Paridae. Wilson Bull. 1989, 101, 305–343. [Google Scholar]
- Fjeldså, J.; Christidis, L.; Ericson, P.G.P. (Eds.) Largest Avian Radiation; Lynx Edicions: Barcelona, Spain, 2020; 445p. [Google Scholar]
- Gosler, A.; Clement, P. Family Paridae (tits and chickadees). In Handbook of the Birds of the World; 12; Elliot, A., Christie, D., Eds.; Lynx Edicions: Barcelona, Spain, 2007; 815p. [Google Scholar]
- Johansson, U.S.; Nylinder, S.; Ohlson, J.I.; Tietze, D.T. Reconstruction of the late Miocene biogeographical history of tits and chickadees (Aves: Passeriformes: Paridae): A comparison between discrete area analyses and probabilistic diffusion approach. J. Biogeogr. 2018, 45, 14–25. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2019. Available online: https://www.R-project.org/ (accessed on 7 July 2020).
- Bergmann, H.-H.; Helb, H.-W.; Baumann, S. Die Stimmen der Vögel Europas; Aula: Wiebelsheim, Germany, 2008. [Google Scholar]
- Tang, S.; Ke, D.; Yu, T.; Wang, C.; Zhao, Q.; Fan, H.; Zhang, G.; Wang, C.; Lu, X. Social organization, demography and genetic mating system of a Tibetan cooperative breeder. Ibis 2017, 159, 687–692. [Google Scholar] [CrossRef]
- Gebauer, A.; Jacob, J.; Kaiser, M.; Eck, S. Chemistry of the uropygial gland secretion of Hume’s ground jay Pseudopodoces humilis and its taxonomic implications. J. Für Ornithol. 2004, 145, 352–355. [Google Scholar] [CrossRef]
- Araya-Salas, M.; Smith-Vidaurre, G. Warbler: An R Package to Streamline Analysis of Animal Acoustic Signals. 2017. Available online: https://CRAN.R-project.org/package=warbleR (accessed on 7 July 2020).
- Marchal, J.; Fabianek, F.; Scott, C.; Corben, C.; Riggs, D.; Wilson, P. Bioacoustics: Analyse Audio Recordings and Automatically Extract Animal Vocalizations; R Package: 2019. Available online: https://CRAN.R-project.org/package=bioacoustics (accessed on 7 July 2020).
- Sueur, J.; Aubin, T.; Simonis, C. Seewave: A Free Modular Tool for Sound Analysis and Synthesis. 2008. Available online: https://CRAN.R-project.org/package=seewave (accessed on 7 July 2020).
- Giannakopoulos, T. pyAudioAnalysis: An open-source python library for audio signal analysis. PLoS ONE 2015, 10, e0144610. [Google Scholar] [CrossRef] [PubMed]
- Van Rossum, G.; Guido, J.; Fred, L.; Drake, J. Python Tutorial; Python Software Foundation: Wilmington, DE, USA, 1995. [Google Scholar]
- Center for Conservation Bioacoustics. Raven Pro: Interactive Sound Analysis Software, Version 1.5. 2014. Available online: http://ravensoundsoftware.com/ (accessed on 7 July 2020).
- Pearse, W.D.; Morales-Castilla, I.; James, L.S.; Farrell, M.; Boivin, F.; Davies, T.J. Global macroevolution and macroecology of passerine song. Evolution 2018, 72, 944–960. [Google Scholar] [CrossRef] [PubMed]
- del Hoyo, J.; Pascual, C.; Sargatal, J. Illustrated Checklist of the Birds of the World; Lynx Edition: Barcelona, Spain, 2016. [Google Scholar]
- Tietze, D.T.; Borthakur, U. Historical biogeography of tits (Aves: Paridae, Remizidae). Org. Divers. Evol. 2012, 12, 433–444. [Google Scholar] [CrossRef]
- BirdLife International, & NaturServe. Bird Species Distribution Maps of the World; BirdLife International and NatureServe: Cambridge, UK, 2011; Available online: https://datazone.birdlife.org/species/requestdis (accessed on 7 July 2020).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling, R Package Version 3.0-12; 2020. Available online: https://CRAN.R-project.org/package=raster (accessed on 7 July 2020).
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the ‘Geospatial’ Data Abstraction Library, R Package Version 1.4-8; 2019. Available online: https://rdrr.io/cran/rgdal/ (accessed on 7 July 2020).
- Pebesma, E.J.; Bivand, R.S. S Classes and Methods for Spatial Data: The sp Package. R News. 2005. Available online: https://www.researchgate.net/publication/228765679_S_Classes_and_Methods_for_Spatial_Data_the_sp_Package (accessed on 7 July 2020).
- Bivand, R.; Pebesma, E.J.; Gómez-Rubio, V. Applied Spatial Data Analysis with R, 2nd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Elle, O. Einführung in die multivariate Statistik für Feldornithologen: Hauptkomponentenanalyse, Diskriminanzanalyse und Clusteranalyse. Vogelwarte 2005, 43, 19–38. [Google Scholar]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; Northwestern University: Evanston, IL, USA, 2020. [Google Scholar]
- Garamszegi, L.Z. Modern Phylogenetic Comparative Methods and Their Application in Evolutionary Biology; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Paradis, E.; Schliep, K. Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R, R package version Version 5.3. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Orme, D.; Freckleton, R.; Thomas, G.; Petzoldt, T.; Fritz, S.; Isaac, N. Caper: Comparative Analyses of Phylogenetics and Evolution in R, R Package Version 1.0.1. 2018. Available online: https://CRAN.R-project.org/package=caper (accessed on 7 July 2020).
- Brock, C.D.; Challenger, W.; Glor, R.E.; Harmon, L.J.; Weir, J.T. Geiger: Investigating evolutionary radiations, R package version 2.0.6.1. Bioinformatics 2008, 24, 129–131. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Blomberg, S.P.; Cornwell, W.K.; Cowan, P.D.; Helmus, M.R.; Kembeland, S.W.; Morlon, H.; Webb, C.O. Picante: Tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Shutler, D.; Weatherhead, P.J. Targets of sexual selection: Song and plumage of wood warblers. Evolution 1990, 44, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J.; Tuschhoff, E. Songs versus colours versus horns: What explains the diversity of sexually selected traits? Biol. Rev. Camb. Philos. Soc. 2020, 95, 847–864. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, G.C. Loudness of birdsong is related to the body size, syntax and phonology of passerine species. J. Evol. Biol. 2009, 23, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.; Örnborg, J.; Andersson, M. Ultraviolet sexual dimorphism and assortative mating in blue tits. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1998, 265, 445–450. [Google Scholar] [CrossRef]
- Miller, E.T.; Leighton, G.M.; Freeman, B.G.; Lees, A.C.; Ligon, R.A. Ecological and geographical overlap drive plumage evolution and mimicry in woodpeckers. Nat. Commun. 2019, 10, 1602. [Google Scholar] [CrossRef] [PubMed]
- McWilliams, S.; Adkins-Regan, E.; Vleck, C. (Eds.) Handbook of Bird Biology: Chapter 7: Bird Physiology, 3rd ed.; Wiley: Chichester, UK, 2016. [Google Scholar]
- Badyaev, A.V.; Leaf, E.S. Habitat associations of song characteristics in Phylloscopus and Hippolais warblers. Auk 1997, 114, 40–46. [Google Scholar] [CrossRef]
- Bertelli, S.; Tubaro, P.L. Body mass and habitat correlates of song structure in a primitive group of birds. Biol. J. Linn. Soc. 2002, 77, 423–430. [Google Scholar] [CrossRef]
- Derryberry, E.P.; Seddon, N.; Derryberry, G.E.; Claramunt, S.; Seeholzer, G.F.; Brumfield, R.T.; Tobias, J.A. Ecological drivers of song evolution in birds: Disentangling the effects of habitat and morphology. Ecol. Evol. 2018, 8, 1890–1905. [Google Scholar] [CrossRef] [PubMed]
- Greig, E.I.; Price, J.J.; Pruett-Jones, S. Song evolution in Maluridae: Influences of natural and sexual selection on acoustic structure. Emu Austral Ornithol. 2013, 113, 270–281. [Google Scholar] [CrossRef]
- Martin, J.P.; Doucet, S.M.; Knox, R.C.; Mennill, D.J. Body size correlates negatively with the frequency of distress calls and songs of Neotropical birds. J. Field Ornithol. 2011, 82, 259–268. [Google Scholar] [CrossRef]
- Mason, N.A.; Burns, K.J. The effect of habitat and body size on the evolution of vocal displays in Thraupidae (tanagers), the largest family of songbirds. Biol. J. Linn. Soc. 2015, 114, 538–551. [Google Scholar] [CrossRef]
- Seddon, N. Ecological adaptation and species recognition drives vocal evolution in Neotropical Suboscine birds. Evolution 2005, 59, 200. [Google Scholar] [CrossRef]
- Tubaro, P.L.; Mahler, B. Acoustic frequencies and body mass in New World doves. Condor 1998, 100, 54–61. [Google Scholar] [CrossRef]
- Wallschläger, D. Correlation of song frequency and body weight in passerine birds. Experientia 1980, 36, 412. [Google Scholar] [CrossRef]
- Podos, J. Correlated evolution of morphology and vocal signal structure in Darwin’s finches. Nature 2001, 409, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Podos, J.; Nowicki, S. Beaks, adaptation, and vocal evolution in Darwin’s finches. Evolution 2004, 54, 501. [Google Scholar] [CrossRef]
- Wiley, R.H. Associations of song properties with habitats for territorial Oscine birds of eastern North America. Am. Nat. 1991, 138, 973–993. [Google Scholar] [CrossRef]
- Caro, L.M.; Caycedo-Rosales, P.C.; Bowie, R.C.K.; Slabbekoorn, H.; Cadena, C.D. Ecological speciation along an elevational gradient in a tropical passerine bird? J. Evol. Biol. 2013, 26, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Boncoraglio, G.; Saino, N. Habitat structure and the evolution of bird song: A meta-analysis of the evidence for the acoustic adaptation hypothesis. Funct. Ecol. 2007, 21, 134–142. [Google Scholar] [CrossRef]
- Harmon, L.J. Phylogenetic Comparative Methods; CreateSpace Independent Publishing Platform: Scotts Valley, CA, USA, 2019; 286p. [Google Scholar]
- Hill, G.E.; Montgomerie, R. Plumage colour signals nutritional condition in the house finch. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1994, 258, 47–52. [Google Scholar] [CrossRef]
- D’Alba, L.; van Hemert, C.; Handel, C.M.; Shawkey, M.D. A natural experiment on the condition-dependence of achromatic plumage reflectance in black-capped chickadees. PLoS ONE 2011, 6, e25877. [Google Scholar] [CrossRef]
- Sheldon, B.C.; Andersson, S.; Griffith, S.C.; Örnborg, J.; Sendecka, J. Ultraviolet colour variation influences blue tit sex ratios. Nature 1999, 402, 874–877. [Google Scholar] [CrossRef]
- Örnborg, J.; Andersson, S.; Griffith, S.C.; Sheldon, B.C.E.N. Seasonal changes in a ultraviolet structural colour signal in blue tits, Parus caeruleus. Biol. J. Linn. Soc. 2002, 76, 237–245. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explanatory Variables | Estimate | Standard Error | t-Value | Pr (>|t|) | R² Adj. | AIC |
|---|---|---|---|---|---|---|
| Intercept | 5385.79 | 935.10 | 5.76 | <0.001 | 0.22 | 884.10 |
| Color diversity | −929.20 | 282.72 | −3.29 | 0.002 | ||
| Body length | −211.87 | 59.77 | −3.55 | 0.001 | ||
| Intercept | 3611.63 | 488.29 | 7.40 | <0.001 | 0.22 | 884.08 |
| Color diversity | −953.98 | 285.01 | −3.35 | 0.002 | ||
| Body mass | −63.56 | 17.91 | −3.55 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tietze, D.T.; Hahn, A. Trade-Off between Song Complexity and Colorfulness in Parid Birds. Diversity 2024, 16, 332. https://doi.org/10.3390/d16060332
Tietze DT, Hahn A. Trade-Off between Song Complexity and Colorfulness in Parid Birds. Diversity. 2024; 16(6):332. https://doi.org/10.3390/d16060332
Chicago/Turabian StyleTietze, Dieter Thomas, and Antje Hahn. 2024. "Trade-Off between Song Complexity and Colorfulness in Parid Birds" Diversity 16, no. 6: 332. https://doi.org/10.3390/d16060332
APA StyleTietze, D. T., & Hahn, A. (2024). Trade-Off between Song Complexity and Colorfulness in Parid Birds. Diversity, 16(6), 332. https://doi.org/10.3390/d16060332