

Species Delimitation of Argonauta Paralarvae Reveals an Extensive Morphological Variability in the Northern Humboldt Current System


Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Morphological Description
2.3. DNA Extraction, Amplification and Sequencing
2.4. Species Delimitation
3. Results
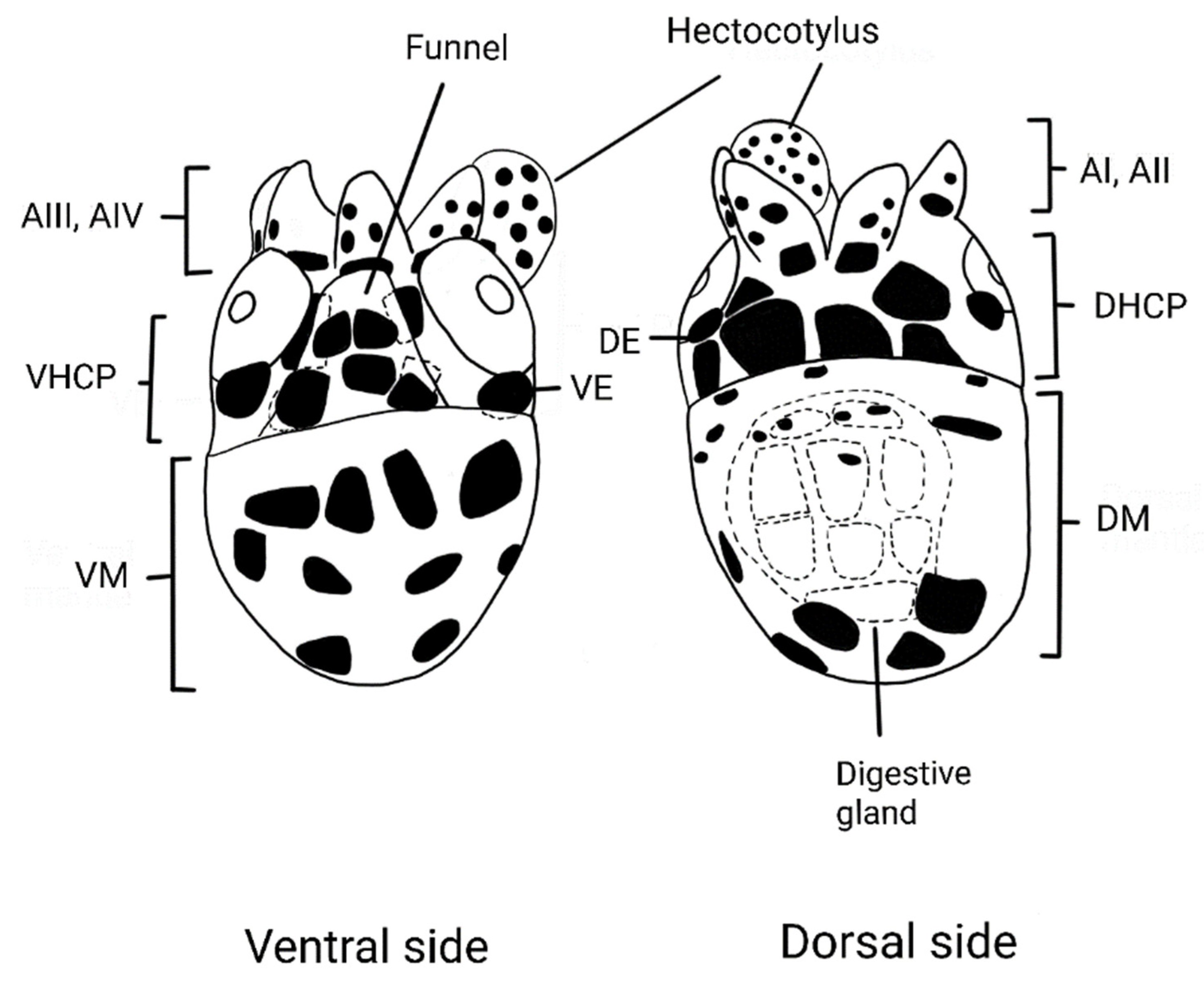
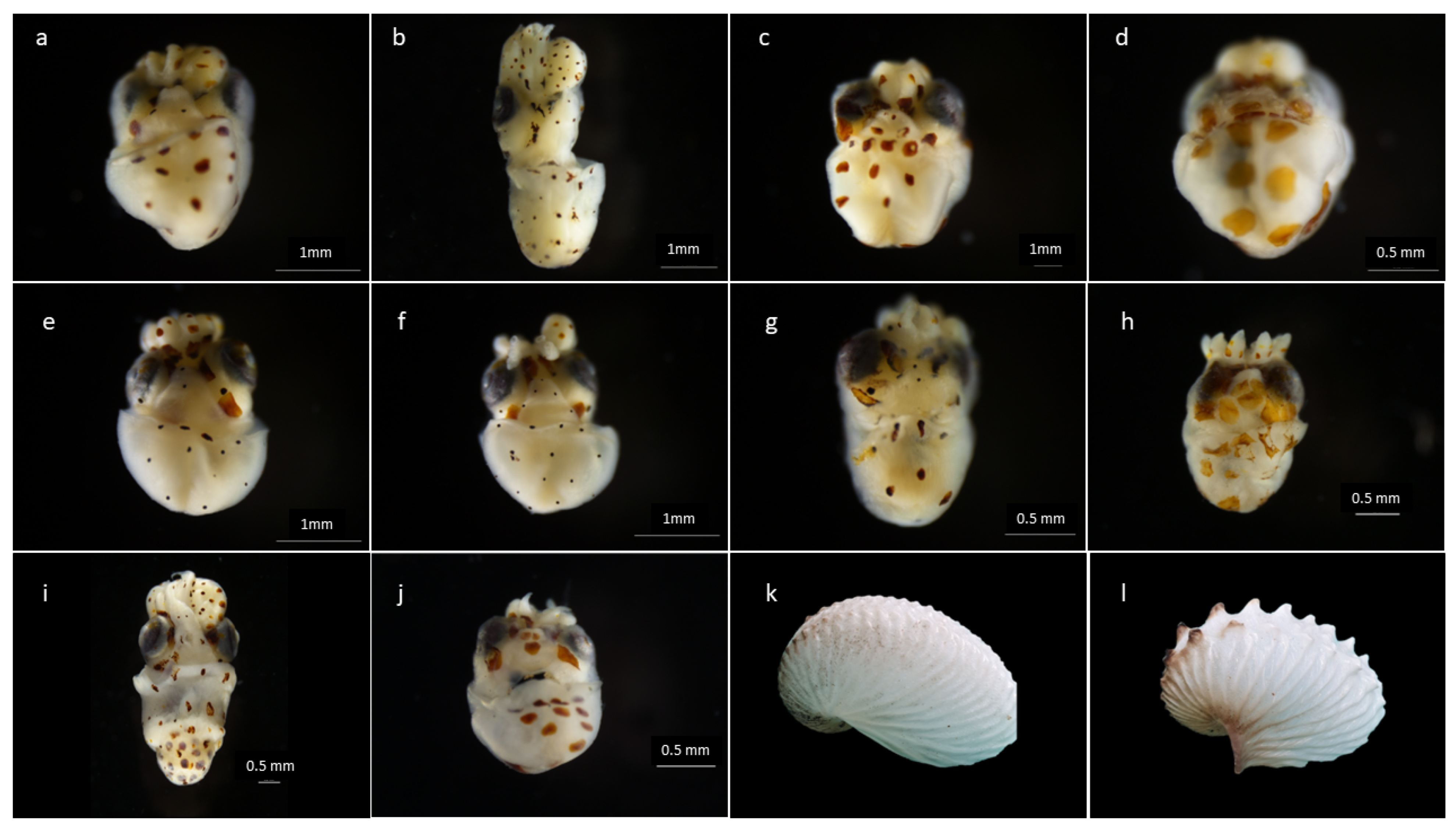
3.1. Morphological Determination
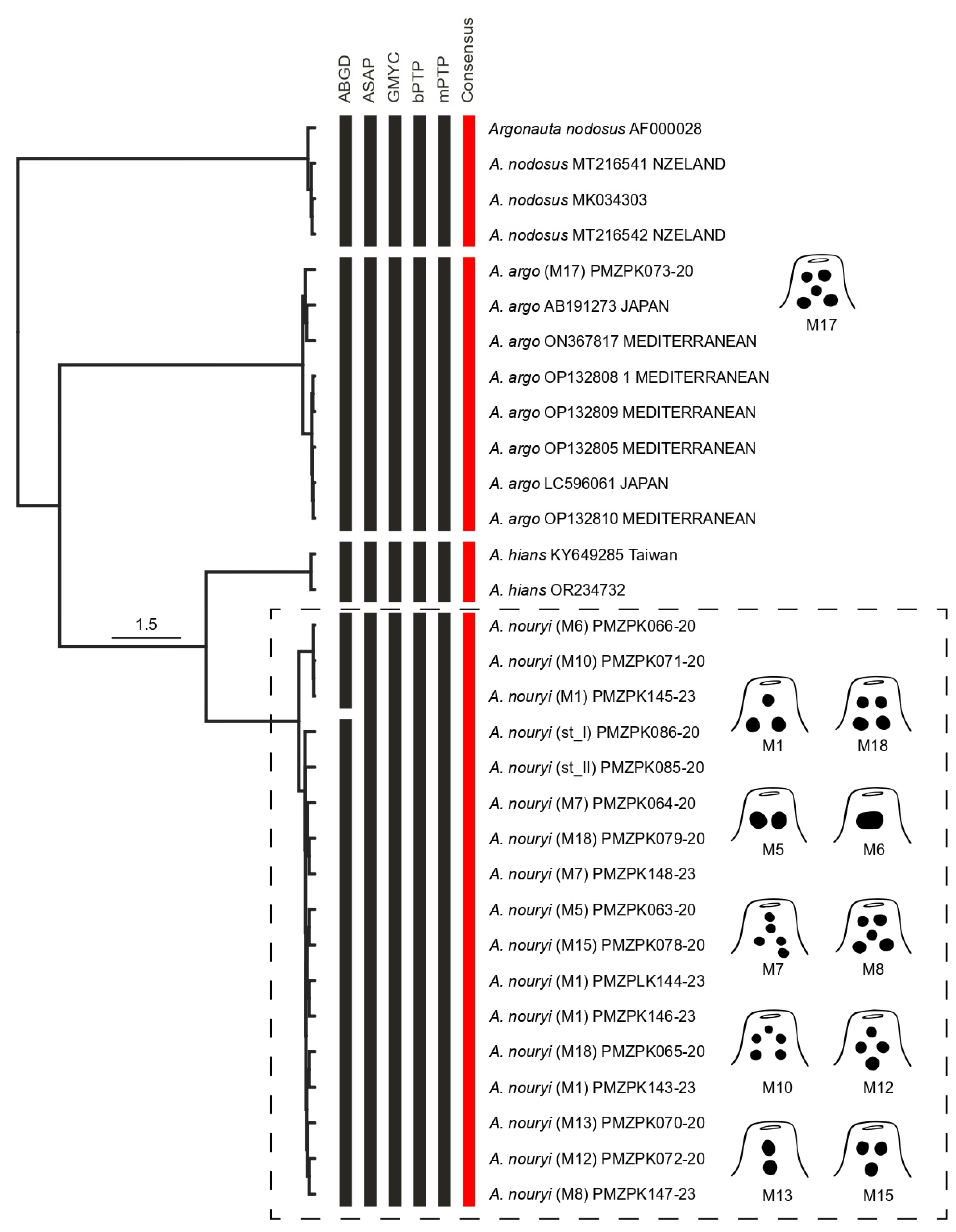
3.2. DNA Barcode Analysis and Species Delimitation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thiel, M.; Macaya, E.; Acuña, E.; Arntz, W.; Bastias, H.; Brokordt, K.; Camus, P.; Castilla, J.; Castro, L.; Cortés, M.; et al. The Humboldt Current System of Northern and Central Chile. In Oceanography and Marine Biology; CRC Press: Boca Raton, FL, USA, 2007; Volume 45, pp. 195–344. [Google Scholar]
- Chavez, F.P.; Bertrand, A.; Guevara-Carrasco, R.; Soler, P.; Csirke, J. The Northern Humboldt Current System: Brief History, Present Status and a View towards the Future. Prog. Ocean. 2008, 79, 95–105. [Google Scholar] [CrossRef]
- Csirke, J.; Argüelles, J.; Alegre, A.; Ayón, P.; Bouchon, M.; Castillo, G.; Castillo, R.; Cisneros, R.; Guevara-Carrasco, R.; Lau, L.; et al. Biología, Estructura Poblacional y Pesquería de Pota o Calamar Gigante (Dosidicus gigas) En El Perú. Bol. Inst. Mar. Perú 2018, 33, 302–364. [Google Scholar]
- Orosco, X.; Ayón, P. Variación de Paralarvas de Cefalópodos Asociada a Condiciones Oceanográficas En La Región Norte Del Sistema de La Corriente de Humboldt. Bol. Inst. Mar. Perú 2022, 37, 256–270. [Google Scholar] [CrossRef]
- FAO. Cephalopods of the World: An Annotated and Illustrated Catalogue of Cephalopod Species Known to Date; Jereb, P., Roper, C.F.E., Norman, M.D., Finn, J.K., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; Volume 3, ISBN 9789251079898. [Google Scholar]
- Finn, J.K. Taxonomy and Biology of the Argonauts (Cephalopoda: Argonautidae) with Particular Reference to Australian Material. Molluscan Res. 2013, 33, 143–222. [Google Scholar] [CrossRef]
- Sasaki, M. Report on Cephalopod Collected during 1906 by The United States Bureau of Fisheries Steaivier “Albatross” in the Northwestern Pacific. Proc. U. S. Natl. Mus. 1920, 57, 163–203. [Google Scholar] [CrossRef]
- Robson, G.C. XXXIX.—Notes on the Cephalopoda.—No. 16. On the Variation, Eggs, and Ovipository Habits of Floridan Octopods. Ann. Mag. Nat. Hist. 1932, 10, 368–374. [Google Scholar] [CrossRef]
- Voss, G.L. The Cephalopoda Obtained by: The Harvard-Havana Expedition off the Coast of Cuba in 1938-39. Bull. Mar. Sci. 1955, 5, 81–115. [Google Scholar]
- Nesis, K.N. Cephalopods of the World; T.F.H. Publications: Neptune, NJ, USA, 1987. [Google Scholar]
- Sweeney, M.J.; Roper, C.F.E.; Mangold, K.M.; Clarke, M.R.; Boletzky, S.V. (Eds.) Smithsonian Contributions to Zoology “Larval” and Juvenile Cephalopods: A Manual for Their Identification; Smithsonian Institution Press: Washington, DC, USA, 1992; Volume 513. [Google Scholar]
- Young, R.E.; Harman, R.F. “Larva”, “Paralarva” and “Subadult” in Cephalopod Terminology. Malacologia 1988, 29, 201–207. [Google Scholar]
- Young, R.E.; Harman, R.F.; Hochberg, F.G. Octopodid Paralarvae from Hawaiian Waters. Veliger 1989, 32, 152–165. [Google Scholar]
- Vidal, É.A.G.; Fuentes, L.; da Silva, L.B. Defining Octopus Vulgaris Populations: A Comparative Study of the Morphology and Chromatophore Pattern of Paralarvae from Northeastern and Southwestern Atlantic. Fish. Res. 2010, 106, 199–208. [Google Scholar] [CrossRef]
- Vecchione, M. Juvenile Ecology. In Cephalopod Life Cycles; Boyle, P.R., Ed.; Academic Press: London, UK, 1987; Volume 2, p. 475. [Google Scholar]
- De Silva-Dávila, R.; Hochberg, F.G.; Lindgren, A.R.; del Carmen Franco-Gordo, M. Paralarval Development, Abundance, and Distribution of Pterygioteuthis hoylei (Cephalopoda: Oegopsida: Pyroteuthidae) in the Gulf of California, México. Molluscan Res. 2013, 33, 50–64. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological Identifications through DNA Barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Rosas-Puchuri, U.; Cañedo, R.M.; Alfaro-Shigueto, J.; Ayon, P.; Zelada-Mázmela, E.; Siccha-Ramirez, R.; Velez-Zuazo, X. DNA Barcoding in the Southeast Pacific Marine Realm: Low Coverage and Geographic Representation despite High Diversity. PLoS ONE 2020, 15, e0244323. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Nuñez-Rodriguez, D.; Britzke, R.; Siccha-Ramirez, R.; Ramírez, R. DNA Barcoding for Assessing Biodiversity. In Conservation Genetics in the Neotropics; Pedro, M.G., Jr., Ed.; Springer International Publishing: Sao Carlos, Brazil, 2023; pp. 21–45. [Google Scholar]
- Fernández-Álvarez, F.; Sánchez, P.; Villanueva, R. Morphological and Molecular Assessments of Bobtail Squids (Cephalopoda: Sepiolidae) Reveal a Hidden History of Biodiversity. Front. Mar. Sci. 2021, 7, 632261. [Google Scholar] [CrossRef]
- Ibáñez, C.M.; Fenwick, M.; Ritchie, P.A.; Carrasco, S.A.; Pardo-Gandarillas, M.C. Systematics and Phylogenetic Relationships of New Zealand Benthic Octopuses (Cephalopoda: Octopodoidea). Front. Mar. Sci. 2020, 7, 182. [Google Scholar] [CrossRef]
- Sales, J.B.d.L.; Rodrigues-Filho, L.F.d.S.; Ferreira, Y.d.S.; Carneiro, J.; Asp, N.E.; Shaw, P.W.; Haimovici, M.; Markaida, U.; Ready, J.; Schneider, H.; et al. Divergence of Cryptic Species of Doryteuthis Plei Blainville, 1823 (Loliginidae, Cephalopoda) in the Western Atlantic Ocean Is Associated with the Formation of the Caribbean Sea. Mol. Phylogenet. Evol. 2017, 106, 44–54. [Google Scholar] [CrossRef]
- Xu, R.; Lü, Y.; Tang, Y.; Chen, Z.; Xu, C.; Zhang, X.; Zheng, X. DNA Barcoding Reveals High Hidden Species Diversity of Chinese Waters in the Cephalopoda. Front. Mar. Sci. 2022, 9, 830381. [Google Scholar] [CrossRef]
- De Silva-Dávila, R. Paralarvas de Cefalópodos En El Golfo de California, México; Universidad de Guadalajara-CUCSUR: San Patricio Melaque, México, 2013. [Google Scholar]
- Zaragoza, N.; Quetglas, A.; Moreno, A. Identification Guide for Cephalopod Paralarvae from the Mediterranean Sea; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2015. [Google Scholar]
- Sukhsangchan, C.; Sunthornket, P.; Phuynoi, S. Morphological Characteristics of Paralarvae of Cephalopods Found in Thai Waters. Mar. Biodivers. 2017, 47, 639–645. [Google Scholar] [CrossRef]
- Montero-Pau, J.; Gómez, A.; Muñoz, J. Application of an Inexpensive and High-Throughput Genomic DNA Method for the Molecular Ecology of Zooplanktonic Eggs. Limnol. Ocean. Methods 2008, 6, 218–222. [Google Scholar] [CrossRef]
- Prosser, S.; Martínez-Arce, A.; Elías-Gutiérrez, M. A New Set of Primers for COI Amplification from Freshwater Microcrustaceans. Mol. Ecol. Resour. 2013, 13, 1151–1155. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Valdivia, P.; Rosas-Puchuri, U.; Valdivia, N.L. SPdel: A Pipeline to Compare and Visualize Species Delimitation Methods for Single-Locus Datasets. Mol. Ecol. Resour. 2023, 23, 1959–1965. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble Species by Automatic Partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A General Species Delimitation Method with Applications to Phylogenetic Placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Paredes, C.; Huamán, P.; Cardoso, F.; Vivar, R.; Vera, V. Estado Actual Del Conocimiento de Los Moluscos Acuáticos En El Perú. Rev. Peru. Biol. 1999, 6, 5–47. [Google Scholar] [CrossRef]
- Alamo, V.; Valdivieso, V. Lista Sistemática de Moluscos Marinos Del Perú, 2nd ed.; Instituto del Mar del Perú: Callao, Peru, 1997. [Google Scholar]
- Ramírez, R.; Paredes, C.; Arenas, J. Moluscos Del Perú. Rev. Biol. Trop. 2003, 51, 225–284. [Google Scholar]
- Battaglia, P.; Pedà, C.; Rizzo, C.; Stipa, M.G.; Arcadi, E.; Longo, F.; Ammendolia, G.; Cavallaro, M.; Rao, I.; Villari, A.; et al. How Rare Are Argonautoidea Octopuses in the Mediterranean? New Data from Stranding Events, Stomach Contents and Genetics. Biology 2023, 12, 420. [Google Scholar] [CrossRef]
- Alejo-Plata, M.d.C.; León-Guzmán, S.S.; Díaz-Polo, R.; Torres-Huerta, A.M. Diversity, Abundance and Frequency of Argonauts (Cephalopoda: Argonautidae) in the Diet of Pelagic Fishes of Commercial Value in Oaxaca, Mexico. Rev. Biol. Mar. Ocean. 2019, 54, 107–117. [Google Scholar] [CrossRef]
- Martínez-Soler, E.; Gómez-Gutiérrez, J.; De Silva-Dávila, R.; González-Rodríguez, E.; Aburto-Oropeza, O. Cephalopod Paralarval Species Richness, Abundance and Size Structure during the 2014-2017 Anomalous Warm Period in the Southern Gulf of California. J. Plankton Res. 2021, 43, 224–243. [Google Scholar] [CrossRef]
- Martínez, E. Variación Temporal de La Estructura de La Comunidad de Paralarvas de Cefalópodos Del Parque Nacional Cabo Pulmo, B.C.S. En Respuesta a La Variabilidad Ambiental (2014–2017); La Paz, B.C.S.: La Paz, Mexico, 2019. [Google Scholar]
- Nesis, K.N. Cephalopoda. In South Atlantic Zooplankton; Boltovskoy, D., Ed.; Backhuys Publishers: Leiden, The Netherlands, 1999. [Google Scholar]
- Vecchione, M.; Roper, C.F.E.; Sweeney, M.J.; Lu, C.C. Distribution, Relative Abundance and Developmental Morphology of Paralarval Cephalopods in the Western North Atlantic Ocean; US Government Printing Office: Seattle, WA, USA, 2001.
- Vidal, E.A.; Haimovici, M.; Hackbart, V.C. Distribution of Paralarvae and Small Juvenile Cephalopods in Relation to Primary Production in an Upwelling Area off Southern Brazil. ICES J. Mar. Sci. 2010, 67, 1346–1352. [Google Scholar] [CrossRef]
- Franco-Santos, R.M.; Iglesias, J.; Domingues, P.M.; Vidal, E.A.G. Early Beak Development in Argonauta nodosa and Octopus vulgaris (Cephalopoda: Incirrata) Paralarvae Suggests Adaptation to Different Feeding Mechanisms. Hydrobiologia 2014, 725, 69–83. [Google Scholar] [CrossRef]
- Cloney, R.A.; Florey, E. Uhrastructure of Cephalopod Chromatophore Organs. Z. Fiir Zellforsch. 1968, 89, 250–280. [Google Scholar] [CrossRef]
- Reed, C.M. The Ultrastructure and Innervation of Muscles Controlling Chromatophore Expansion in the Squid, Loligo vulgaris. Cell Tissue Res. 1995, 282, 503–512. [Google Scholar] [CrossRef]
- Messenger, J.B. Cephalopod Chromatophores: Neurobiology and Natural History. Biol. Rev. 2001, 76, 473–528. [Google Scholar] [CrossRef]
- Vecchione, M.; Lipiński, M.R. Descriptions of the Paralarvae of Two Loliginid Squids in Southern African Waters. S. Afr. J. Mar. Sci. 1995, 15, 1–7. [Google Scholar] [CrossRef]
- González, Á.F.; Rocha, F.; Guerra, A.; Bückle, L.; Guerra, A.; Rocha, E.; González, A.E.; Bückle, L.E. Embryonic Stages of the Patagonian Squid Loligo Gahi (Mollusca: Cephalopoda). The Veliger 2001, 44, 109–115. [Google Scholar]
- Barón, P.J. The Paralarvae of Two South American Sympatric Squid: Loligo Gahi and Loligo Sanpaulensis. J. Plankton Res. 2003, 25, 1347–1358. [Google Scholar] [CrossRef]
- Troschel, F.H. Bemerkungen Über Die Cephalopoden von Messina. Arch. Naturgeschichte 1857, 23, 41–76. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Barcode Accession Number | Sex | Dorsal Mantle Length (mm) | Latitude | Longitude | Net | Year |
|---|---|---|---|---|---|---|
| PMZPK063_20 | M | 1.1 | 8°5.954 S | 83°45.008 W | Bongo 300 µm | 2017 |
| PMZPK064_20 | M | 1.0 | 7°33.569 S | 82°49.057 W | Bongo 300 µm | 2017 |
| PMZPK065_20 | F | 1.2 | 7°3.598 S | 83°19.075 W | Bongo 300 µm | 2017 |
| PMZPK066_20 | F | 1.0 | 7°31.346 S | 84°6.873 W | Bongo 300 µm | 2017 |
| PMZPK070_20 | F | 1.2 | 6°39.534 S | 81°56.751 W | Bongo 300 µm | 2017 |
| PMZPK071_20 | F | 1.0 | 6°39.534 S | 81°56.751 W | Bongo 300 µm | 2017 |
| PMZPK072_20 | F | 1.7 | 6°40.1 | 82°38.677 W | Bongo 300 µm | 2017 |
| PMZPK073_20 | F | 1.0 | 6°0.039 S | 82°57.959 W | Bongo 300 µm | 2017 |
| PMZPK078_20 | F | 0.9 | 6°54.661 S | 81°41.04 W | Bongo 300 µm | 2017 |
| PMZPK079_20 | F | 0.8 | 8°011 S | 83°34.898 W | Bongo 300 µm | 2017 |
| PMZPK085_20 | F | egg mass | 13°28.921 S | 77°0.538 W | Pelagic trawl | 2019 |
| PMZPK086_20 | F | egg mass | 13°28.921 S | 77°0.538 W | Pelagic trawl | 2019 |
| PMZPK143-23 | - | 2.2 | 9°48.171 S | 80°08.640 W | IKMT | 2022 |
| PMZPK144-23 | - | 1.0 | 14° 4.001 S | 77°29.002 W | CUFES | 2019 |
| PMZPK145-23 | F | juvenile (3.5 mm) | 12°03.440 S | 77°10.0405 W | Bongo 300 µm | 2023 |
| PMZPK146-23 | - | 2.5 | 13°26.627 S | 78°12.331 W | Bongo 300 µm | 2019 |
| PMZPK147-23 | - | 1.0 | 10°44.882 S | 80°36.264 W | Bongo 300 µm | 2019 |
| PMZPK148-23 | - | 1.3 | 6°30.058 S | 82°10.967 W | Bongo 300 µm | 2019 |
| Barcode Accession Number | Morphotype | ML (mm) | Sex | Chromatophore Pattern | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DHCP | VHCP | DM | VM | DE | VE | DG | Funnel | Arms | Hectocotyl | ||||
| PMZPK145-23 | M1 | 3.5 | F | - | - | - | - | - | - | - | 1 + 2 | BI:1b + 19dr/BII:1b + 11dr/BIII:1b + 11dr/BIV:1b + 14dr | |
| PMZPK146-23 | M1 | 2.5 | 2 + 4 | 2 + 2 | - | - | 2(2) | 1(2) | - | 1 + 2 | - | ||
| PMPZK063-20 | M5 | 1.1 | M | 2 + 4 | 2 + 2 | 4 + 4 + 2 + 1 | 4 + 3 + 3 + 2 + 1 | 2(2) | 1(2) | 9 | 2 | BI-BIII:1b + 1/BIV:1b + 1 + 1 | 10 |
| PMPZK066-20 | M6 | 1 | F | 2 + 4 | 2 + 2 | 5 + 2 + 1 | 5 + 3 + 2 + 2 + 2 | 1(2) | 1(2) | 8 | 1 | BI-BIII: 1b/BIV: 1b + 1 | - |
| PMPZK064-20 | M7 | 1 | M | 2 + 4 | 2 + 2 | 4 + 4 + 2 + 1~ | 5 + 1 * + 4 + 2 + 1 | 2(2) | 1(2) | 10 | 1 + 2 + 1 * | BI-BIII: 1b + 1 + 1dr (d)/BIV: 1b + 2 + 1dr(d) | 33 |
| PMZPK147-23 | M7 | 1.3 | 2 + 4 | 2 + 2 | - | - | 2(2) | 1(2) | - | 1 + 1 + 2 | - | ||
| PMZPK148-23 | M8 | 1 | 2 + 4 | 2 + 2 | - | - | 2(2) | 1(2) | - | 2 + 1 + 2 | - | ||
| PMPZK071-20 | M10 | 1 | F | 2 + 4 | 2 + 2 | 4 + 2 + 2 + 1 | 4 + 3 + 5 + 1 + 2 | 2(2) | 1(2) | 8 | 1 + 2 + 2 | BI:1b + 1 * + 1dr(d)/BII:1b + 1/BIII:1b/BIV: 1b + 1 | 10 |
| PMPZK072-20 | M12 | 1.7 | F | 2 + 4 | 2 + 2 | 5 + 2 + 1 | 54 + 1 + 2 | 2(2) | 1(2) | 10 | 1 + 2 + 1 | BI- BIII: 1b/BIV: 1b + 1 | - |
| PMZPK070-20 | M13 | 1.2 | F | 2 + 4 | 2 + 2 | 4 + 2 + 2 + 1 + 2 | 4 + 2 + 3 + 2 | 2(2) | 1(2) | 10 | 1 + 1 | BI: 1b + 1 + 1dr(d)/BII-BIII: 1b + 1/BIV:1b + 1 + 1 | - |
| PMPZK078-20 | M15 | 0.9 | F | 2 + 4 | 2 + 2 | 5 + 1 * + 2 | 4 + 3 + 2 + 1 | 2(2) | 1(2) | 11 | 2 + 1 | BI-BIII: 1b/BIV: 1b + 1 | - |
| PMPZK073-20 | M17 | 1 | F | 2 + 4 | 2 + 2 | 3 + 7 + 1 * + 2 + 1 | 4 + 4 + 2 | 1(2) | 1(2) | 9 | 2 + 1 + 2 | BI: 1b + 1 + 1dr(d)/BII-BIII: 1b + 1/BIV: 1b + 1dr + 1 | - |
| PMZPK065-20 | M18 | 1.2 | F | 2 + 4 | 2 + 2 | 4 + 3 + 2 | 4 + 3 + 2 | 2(2) | 1(2) | 9 | 2 + 2 | BI & BIV: 1b + 1/BII-BIII: 1b | - |
| PMPZK079-20 | M18 | 0.8 | F | 2 + 4 | 2 + 2 | 4 + 2 + 1 | 4 + 3 + 1 + 2 | 1(2) | 1(2) | 9 | 2 + 2 | BI & BIV 1b + 1/BII-BIII: 1b | - |
| PMZPK143-23 | M18 | 2.2 | F | 2 + 4 | 2 + 2 | 5 + 4 + 4 + 3 + 2 + 1 | 5 + 5 + 4 + 5 | 2(2) | 1(2) | - | 2 + 2 | BI: 1b + 4dr/BII: 1b + 4sr/BIII: 1b + 3sr/BIV: 1b + 5dr | - |
| PMZPK144-23 | M18 | - | 2 + 4 | 2 + 2 | - | - | 2(2) | 1(2) | - | 2 + 2 | - | - | |
| PMZPK086-20 | st_I | F | Egg mass of female with shell type I (Finn 2013) | - | - | ||||||||
| PMPZK085-20 | st_II | F | Egg mass of female with shell type II (Finn 2013) | - | - | ||||||||
| Consensus MOTUs | Mean | Max | NN | DtoNN |
|---|---|---|---|---|
| MOTU 01 (A. argo) | 0.22 | 0.64 | MOTU 04 | 8.80 |
| MOTU 02 (A. hians) | 0.16 | 0.16 | MOTU 04 | 4.63 |
| MOTU 03 (A. nodosus) | 0.16 | 0.32 | MOTU 02 | 11.01 |
| MOTU 04 (A. nouryi) | 0.28 | 0.97 | MOTU 02 | 4.63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orosco, X.; Ayón, P.; Sotil, G.; Ramirez, J.L. Species Delimitation of Argonauta Paralarvae Reveals an Extensive Morphological Variability in the Northern Humboldt Current System. Diversity 2024, 16, 320. https://doi.org/10.3390/d16060320
Orosco X, Ayón P, Sotil G, Ramirez JL. Species Delimitation of Argonauta Paralarvae Reveals an Extensive Morphological Variability in the Northern Humboldt Current System. Diversity. 2024; 16(6):320. https://doi.org/10.3390/d16060320
Chicago/Turabian StyleOrosco, Ximena, Patricia Ayón, Giovanna Sotil, and Jorge L. Ramirez. 2024. "Species Delimitation of Argonauta Paralarvae Reveals an Extensive Morphological Variability in the Northern Humboldt Current System" Diversity 16, no. 6: 320. https://doi.org/10.3390/d16060320
APA StyleOrosco, X., Ayón, P., Sotil, G., & Ramirez, J. L. (2024). Species Delimitation of Argonauta Paralarvae Reveals an Extensive Morphological Variability in the Northern Humboldt Current System. Diversity, 16(6), 320. https://doi.org/10.3390/d16060320